Back to Journals » Journal of Pain Research » Volume 18
Modulating Gamma Oscillations in the Anterior Cingulate Cortex with Intermittent Theta Burst Stimulation: Implications for Chronic Non-Specific Low Back Pain
Authors Jiang H ![]() , Wang Q
, Wang Q ![]() , Ye Z
, Ye Z ![]() , Ding Y
, Ding Y
Received 19 August 2025
Accepted for publication 23 December 2025
Published 30 December 2025 Volume 2025:18 Pages 7299—7309
DOI https://doi.org/10.2147/JPR.S561653
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Überall
Hui Jiang,1,* Qiaohua Wang,1,* Zhengfei Ye,2 Yi Ding3
1Huangshi Hospital of Traditional Chinese Medicine, Huangshi, Hubei, People’s Republic of China; 2Department of Rehabilitation, Huangshi Hospital of Traditional Chinese Medicine, Huangshi, Hubei, People’s Republic of China; 3Huangshi Central Hospital, Huangshi, Hubei, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhengfei Ye; Yi Ding, Email [email protected]; [email protected]
Abstract: Chronic nonspecific low back pain (CNSLBP), a debilitating condition associated with abnormal anterior cingulate cortex (ACC) activity and central sensitization, may be addressed through intermittent theta-burst stimulation (iTBS), a non-invasive neuromodulation technique. iTBS is designed to restore the ACC’s excitatory/inhibitory balance by suppressing abnormal gamma oscillations and enhancing synaptic plasticity. Clinical evidence shows that iTBS can reduce ACC gamma power, strengthen corticostriatal circuit connectivity, and provide short-term pain relief, although efficacy varies with protocol, pain subtype, and individual differences. In conclusion, iTBS represents a promising approach for managing CNSLBP by modulating neuroplasticity. Future efforts should focus on optimizing stimulation parameters, integrating multimodal data for personalized treatment, and validating long-term benefits through large-scale trials to shift the therapeutic paradigm towards remodeling maladaptive neural pathways.
Keywords: low back pain, anterior cingulate cortex, nerve oscillations, gamma oscillations, intermittent theta burst stimulation, chronic pain
Introduction: The Complexity of CNSLBP and the Potential for Neuromodulation
Chronic nonspecific low back pain (CNSLBP), defined as persistent pain lasting >3 months without identifiable structural pathology, affects up to 15.4% of middle-aged and elderly individuals and constitutes the majority (≈62%) of chronic low back pain cases worldwide, imposing enormous socioeconomic burden and frequently co-occurring with anxiety, depression, sleep disturbances1,2 and heightened fall risk, particularly in geriatrics where it correlates with reduced functional capacity, physical performance, and balance as evidenced by validated assessments like the Five Times Sit-to-Stand Test (FTSST), which demonstrates excellent reliability (ICC=0.99) and strong associations with pain intensity, disability, and lower extremity strength.3 Conventional pharmacologic (NSAIDs, opioids) and physical therapies, including core stabilization exercises alone or combined with aerobic protocols, predominantly offer symptomatic relief—yielding improvements in functional capacity,4 pain, depression, and fall risk yet consistently fail to modify the underlying central sensitization—a maladaptive neuroplastic change characterized by hyperexcitability of spinal dorsal horn neurons and supraspinal pain-processing circuits, most prominently the anterior cingulate cortex (ACC).5–8
A critical knowledge gap persists: although neuroimaging and electrophysiological studies robustly demonstrate that ACC hyperactivity, aberrant functional connectivity, and pathologically elevated gamma oscillations (23–80 Hz) directly encode ongoing pain intensity and sustain central sensitization through disrupted excitatory–inhibitory (E/I) balance and excessive synaptic potentiation,9–12 no current treatment possesses the spatiotemporal precision to selectively normalize these ACC-mediated oscillatory and circuit-level abnormalities. Traditional high-frequency repetitive transcranial magnetic stimulation (HF-rTMS), despite evidence of analgesic efficacy, requires protracted sessions (20–40 min), limiting clinical scalability,13 whereas pharmacological approaches targeting central mechanisms remain hampered by off-target effects and inability to modulate specific neural rhythms,5 even exercise-based interventions, while superior when aerobic components are added to core stabilization for enhancing geriatric outcomes like reduced fall risk and better physical performance, lack direct impact on these neural signatures.4,14,15
This therapeutic void raises a pivotal research question: Can targeted, non-invasive neuromodulation of ACC gamma oscillatory networks reverse central sensitization and provide durable relief in CNSLBP? Intermittent theta-burst stimulation (iTBS) emerges as the most promising candidate to address this challenge.16 By delivering brief (≈3 min), high-intensity bursts of 50 Hz gamma stimulation nested within a 5 Hz theta rhythm — a pattern that physiologically mimics endogenous hippocampal–prefrontal theta–gamma coupling — iTBS potently induces NMDA receptor- and CaMKII-dependent long-term potentiation (LTP)-like plasticity while restoring E/I balance far more efficiently than conventional HF-rTMS.17–19 When applied over the left dorsolateral prefrontal cortex (DLPFC), iTBS exerts downstream modulation of the interconnected ACC,20 directly counteracting injury-induced Cx36-mediated electrical coupling, pathological gamma synchronization, and the resultant amplification of nociceptive signaling observed in both animal models and human chronic pain states.21,22 Thus, iTBS represents a paradigm-shifting, mechanistically grounded intervention uniquely capable of remodeling the maladaptive neuroplasticity that perpetuates CNSLBP, offering superior efficiency, tolerability, and potential for personalized oscillatory targeting compared to existing neuromodulation and pharmacological strategies13,16,23 (Figure 1).
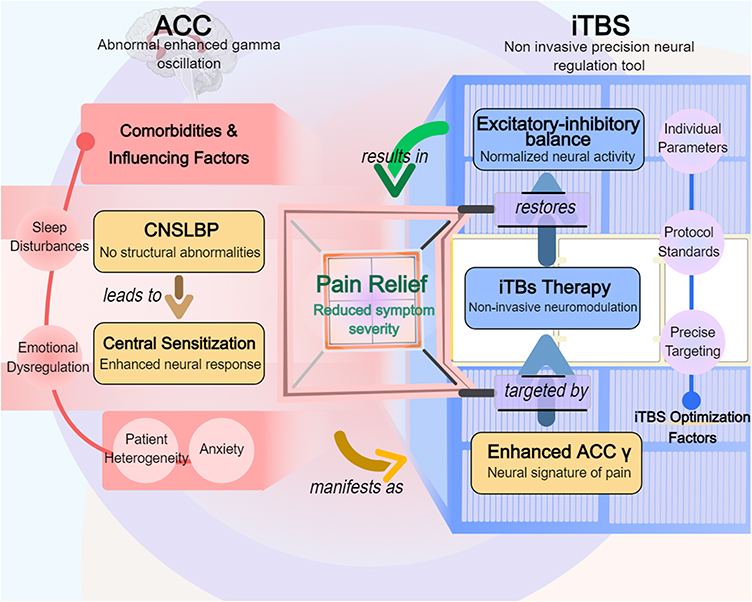 |
Figure 1 Systematically elucidates the pathophysiological mechanisms of chronic nonspecific low back pain (CNSLBP) and the intervention logic of intermittent theta pulse stimulation (iTBS). The core pathological manifestation is abnormal enhancement of gamma oscillations in the anterior cingulate cortex (ACC), which triggers chronic pain through central sensitization and is accompanied by comorbidities such as sleep disorders, emotional dysregulation, and anxiety. iTBS, as a non-invasive precision neurostimulation tool, targets the ACC to regulate the excitatory-inhibitory (E/I) balance, restore neural activity normalization, and thereby alleviate symptom severity (pain relief). Treatment optimization requires consideration of patient heterogeneity, combined with individualized parameters (such as precise targeting localization and standardized stimulation protocols) to enhance efficacy. |
The Central Role of γ Oscillations of the ACC in CNSLBP
Functional Anatomy of the ACC and Multidimensional Integration of Pain Information Processing
The ACC functions as a pivotal “hub” connecting the limbic system and the higher cortical regions, with its anatomical connectivity network underpinning its essential role in multidimensional pain coding. Structurally, the ACC is implicated in pain processing through two principal pathways: (1) The ventral pathway, wherein the ACC establishes an “emotion-reward loop” with the ventral tegmental area (VTA) and the nucleus accumbens (NAc), facilitating the integration of negative emotions and motivational deficits associated with pain.24 For instance, optogenetic inhibition of ACC projections to the NAc has been shown to significantly diminish avoidance behavior in response to painful stimuli in mice.24 (2) The dorsal pathway, characterized by the ACC forming a “sensory-motor loop” with the dorsomedial striatum (DMS) and the ventral posterior nucleus of the thalamus (VPL/VPM), plays a crucial role in modulating pain-related motor inhibition and decision-making bias.24,25 Functional imaging studies have demonstrated that the strength of functional connectivity between the ACC and the DMS during the anticipatory phase of pain is significantly greater in patients with central sensitization-related low back pain (CSRLBP) compared to healthy individuals (p < 0.001), and this connectivity is positively correlated with the extent of pain-induced motor restriction.25,26
At the molecular level, the imbalance between E/I within the ACC constitutes a critical mechanism underlying the persistence of chronic pain. (1) Inhibitory neuronal apoptosis: Nerve injury triggers the activation of the TNF-α-necroptosis pathway, characterized by the phosphorylation of RIP1, RIP3, and MLKL, which results in the necrotic apoptosis of parvalbumin (PV)-expressing inhibitory interneurons (PV-INs). This process directly diminishes inhibitory synaptic inputs.27 Concurrently, the phagocytosis of microglial cells mediated by NLRP3 inflammasomes further contributes to the depletion of PV-INs, thereby aggravating the E/I imbalance.28 (2) Impaired GABAergic transmission: In models of chronic inflammation, there is a down-regulation of presynaptic vesicular GABA transporter (VGAT) expression in the ACC, leading to a decreased frequency of GABA release. Notably, the subunits of GABAA receptors remain unchanged, indicating a dysfunction in presynaptic inhibitory mechanisms.29 (3) Pyramidal cell hyperexcitability: Following nerve injury, there was a significant increase in the in vivo discharge frequency of ACC pyramidal cells, accompanied by an up-regulation of c-Fos expression, indicating enhanced intrinsic excitability. In contrast, inhibitory neuronal activity did not exhibit a significant change, leading to a dysregulated excitatory/inhibitory (E/I) ratio.30 This imbalance contributes to the ACC’s heightened responsiveness to low-threshold sensory inputs, thereby establishing the neural foundation for “pain memory”.12
Neural Mechanisms of γ Oscillations: From Synaptic Dynamics to Network Coding
γ oscillations function as temporal encoders for the processing of pain information within the ACC. Their generation is contingent upon the precise interactions between two distinct neuronal types: (1) fast-spiking interneurons (FS-INs), which facilitate rhythmic synchronization of the local network through GABAergic synaptic release,(2) pyramidal neurons, which receive synchronization inhibition to produce clustered discharges that establish phase locking within the γ band.31–33 In a healthy state, γ oscillations in the ACC demonstrate task-dependent dynamic regulation; acute painful stimuli elicit a transient increase in γ power, which subsequently returns to baseline through a negative feedback loop mediated by NMDA receptors.34
In patients with CNSLBP, the spatiotemporal characteristics of γ oscillations exhibit significant abnormalities: (1) Power Anomaly: Consistent with the above, resting-state γ power is elevated and correlates with pain sensitivity;35 (2) Rhythm Disturbance: Phase-amplitude coupling (PAC) analysis reveals a decreased coupling strength between γ oscillations in the ACC and the theta frequency band (4–8 Hz), leading to less efficient information integration across frequency bands;34 (3) Propagation Abnormality: Dynamic causal modeling (DCM) indicates that the effective connectivity strength of γ-band connections from the ACC to the insula is weakened in CNSLBP patients, while connections to the amygdala are abnormally enhanced, exacerbating the comorbidity of pain and anxiety.36,37
The molecular underpinnings of the aberrant γ activity are intricately linked to potassium channel dysfunction. Chronic pain induces a reduction in Kv4.2 channel expression within ACC pyramidal neurons, resulting in an elevation of their membrane potential oscillation frequency from 8.2 Hz to 12.5 Hz.38,39 Concurrently, the miR-17-92 cluster exacerbates neuronal hyperexcitability by destabilizing Kv1.1 channel mRNA.39,40 This “dual potassium channel defect” diminishes the synchronization threshold for γ oscillations, thereby increasing the frequency of spontaneous γ burst events.41–44
The Vicious Circle of γ Oscillations and CNSLBP: From Local Abnormalities to Whole-Brain Network Dysregulation
Abnormal γ oscillations in the ACC not only indicate dysfunction within local neural circuits but also contribute to the reorganization of whole-brain networks via long-range projections. Specifically, three key pathways are affected: (1) The cortico-striatal loop, wherein glutamatergic projections from the ACC to the dorsomedial striatum (DMS) are significantly enhanced in chronic pain conditions. Optogenetic inhibition of this pathway has been shown to increase the mechanical pain threshold (p < 0.001).25 During this process, γ oscillations in the ACC facilitate synchronized firing among DMS dopaminergic neurons, thereby augmenting pain-related motor inhibition;25,35 (2) The limbic-thalamic loop, characterized by elevated γ band coherence between the ACC and the ventral posterior lateral nucleus of the thalamus (VPL) in patients with CSRLBP. This increased coherence is associated with thalamic gating failures, resulting in the misinterpretation of non-injurious sensory inputs as pain signals.45–47 (3) Default Mode Network (DMN): Aberrant γ activity in the ACC disrupts the resting state of the DMN by inhibiting the alpha (α) oscillations (8–12 Hz) of the posterior cingulate gyrus (PCC), thereby impairing functional integration. This disruption manifests as cognitive deficits related to pain, such as declines in working memory.9,48,49 Current research collectively indicates that disturbed γ oscillations in the ACC act as “amplifiers” of the expansion of CNSLBP from localized plasticity abnormalities to pathological networks across the entire brain, thereby providing a critical opportunity for targeted intervention.50
Mechanism and Clinical Translation of iTBS Modulation of ACC γ Oscillations
Bidirectional Regulatory Mechanisms of iTBS: From Synaptic Plasticity to Network Remodeling
The modulatory influence of iTBS on ACC γ oscillations arises from its spatiotemporal-specific regulation of neuroplasticity. At the molecular level, iTBS alters the E/I balance within the ACC through a dual mechanism: (1) Rapid modulation: The γ pulses (50 Hz) associated with iTBS facilitate the swift remodeling of dendritic spines in pyramidal neurons by activating voltage-gated calcium channels. This activation promotes the release of glutamate from the presynaptic membrane, thereby triggering NMDA receptor-mediated calcium influx.19,20,51,52 This mechanism has been validated in animal models through electrophysiological recordings53 and human imaging data.13 (2) Slow modulation: The theta rhythm of iTBS (5 Hz) augments GABAergic transmission and mitigates aberrant γ synchronization in parvalbumin-positive (PV+) interneurons through the entrainment of theta oscillatory phases within the ACC.19 Animal studies have demonstrated that following iTBS intervention, there is a reduction in γ power within the ACC and an increase in GAD67 (GABA synthase) expression.52,54,55
At the systemic level, iTBS exerts its efficacy by modulating the dynamic connectivity of the ACC with brain-wide networks. Functional magnetic resonance imaging (fMRI) studies have demonstrated that a single iTBS intervention leads to a reduction in the strength of the ACC’s functional connectivity with the default mode network (DMN), while simultaneously enhancing its theta-band coherence with the dorsolateral prefrontal cortex (DLPFC). This reorganization of network connectivity is significantly correlated with pain relief (p<0.001).55 Furthermore, dynamic causal modeling (DCM) has revealed that iTBS restores normal coding of the affective dimension of pain by suppressing abnormal γ connectivity from the ACC to the amygdala and enhancing theta-gamma cross-frequency coupling from the ACC to the insula.56
Parameter Optimization: From Empirical Settings to Individualized Precision Regulation
The efficacy of iTBS is significantly influenced by the precision of parameter settings, necessitating optimization across three dimensions. First, target localization: research indicates that the strength of resting-state functional connectivity (rsFC) between the left dorsolateral prefrontal cortex and the ACC is predictive of iTBS’s impact on hypothalamic-pituitary-adrenal (HPA) axis modulation.20 Specifically, a stronger baseline anticorrelation between the DLPFC and ACC correlates with a more pronounced reduction in cortisol levels (AUCi) following iTBS intervention, highlighting the importance of integrating individualized target selection with functional connectivity features to enhance intervention specificity. Second, stimulation intensity: empirical evidence suggests that an iTBS intensity set at 120% of the resting motor threshold (RMT) achieves safe and effective modulation, thereby improving therapeutic efficacy compared to lower-intensity protocols, such as 80% of the active motor threshold,13 without increasing the risk of adverse effects. Furthermore, the modulation of γ oscillations can be achieved by integrating iTBS with γ-frequency transcranial alternating current stimulation (γ-tACS). The current intensity for γ-tACS is generally set at a peak amplitude of 1 mA and a frequency range of 50–80 Hz.18,52 Third, the temporal pattern: iTBS utilizes an intermittent theta burst pattern, characterized by 3 pulses with an intra-cluster frequency of 50 Hz and an inter-cluster frequency of 5 Hz. For effective synergistic modulation of γ oscillations, γ-tACS should be initiated approximately 10 seconds before the commencement of iTBS and maintained throughout the process.52 This spatiotemporal coupling facilitates the synchronization of cortical γ rhythms with iTBS-induced synaptic plasticity, significantly enhancing long-term potentiation (LTP) effects and extending the increase in motor evoked potentials (MEP) beyond 30 minutes.18 Notably, the modulation of GABAergic interneurons through γ-tACS can be effectively quantified using short-interval intracortical inhibition (SICI), which demonstrates an inhibitory effect that positively correlates with the change in motor evoked potential (MEP) amplitude following iTBS.52 This correlation suggests that γ oscillations enhance synaptic remodeling by modulating GABA-Aergic neurotransmitter homeostasis. Consequently, by integrating individualized target localization, optimized stimulation intensity, and synergistic timing design, iTBS can achieve targeted intervention in the neuromodulation of ACC-related networks and γ oscillations, thereby offering a precise strategy for therapeutic interventions in disease treatment.
Clinical Translational Evidence: Efficacy, Heterogeneity, and Safety
(1) Efficacy: Numerous studies have demonstrated that the analgesic effect of iTBS is both context-dependent and time-limited. In a randomized controlled trial involving 19 patients with chronic orofacial pain, it was observed that visual analogue scores (VAS) significantly decreased (p<0.05) immediately following a single iTBS treatment; however, this effect was not sustained at the two-week follow-up.57 Similarly, another study reported that a single session of iTBS in patients with neuropathic pain led to a reduction in VAS scores within 60 minutes, but the analgesic effect diminished rapidly. Furthermore, the efficacy of iTBS was significantly lower compared to 20 Hz conventional high-frequency repetitive transcranial magnetic stimulation (rTMS).23 A separate investigation into neuropathic pain following spinal cord injury revealed a reduction in VAS from baseline after four consecutive weeks of iTBS treatment (p<0.05 between groups), indicating that the efficacy of repeated iTBS sessions was superior to that of a single application.58 These findings suggest that the efficacy of iTBS may be influenced by the treatment regimen, such as the frequency of sessions, as well as the specific type of pain being treated.
(2) Heterogeneity: In a study involving 15 patients with central pain, iTBS was reported to provide significant pain relief, although the proportion of effective responders was not specified.59 Conversely, research by André-Obadia et al suggests that the efficacy of iTBS may be contingent upon the efficiency of integrating pain mechanisms and neural networks.23 Furthermore, Yang et al demonstrated that iTBS, when used as an initial stimulus in combination with other treatments, was more effective than when applied in isolation, underscoring the synergistic differences in treatment regimens.58 Discrepancies among studies may be attributed to patient heterogeneity, such as variations in pain etiology and cortical plasticity, as well as differences in the selection of stimulation parameters, including localization accuracy and session design.
(3) Safety: Multiple lines of evidence indicate that iTBS is generally well tolerated. In the Accelerated iTBS Protocol Trial conducted by Drabek et al, no serious adverse events were reported among the first 10 patients with chronic pain, and the low dropout rate was comparable to that observed with conventional repetitive transcranial magnetic stimulation (rTMS).60 Blumberger et al found that the iTBS group experienced significantly higher treatment-related pain scores compared to the 10Hz rTMS group (3.8 vs 3.4/10, p=0.011), although there was no significant difference in dropout rates (8% vs 6%) or headache incidence (65% vs 64%).13 However, long-term safety data in the context of chronic pain require further refinement, as the available evidence is predominantly derived from single-center, small-sample studies.60 Consequently, while iTBS demonstrates a transient analgesic effect in chronic pain, its efficacy is significantly influenced by disease subtype and treatment regimen. Despite its safety advantages, the optimal indications and standardized parameters for iTBS need to be elucidated through multicenter, large-sample studies.57
Critical Limitations and Methodological Challenges in Current Neuromodulation Research for CNSLBP
The investigation of neuromodulation as a treatment for CNSLBP is constrained by significant limitations in both preclinical and clinical research domains, alongside pervasive methodological flaws. In animal studies, a critical challenge is the translational gap. Commonly used neuropathic pain models (eg, sciatic nerve ligation) fail to fully recapitulate the complex psychopathological and functional dimensions of human CNSLBP. Behavioral assessments primarily rely on reflexive measures like paw withdrawal thresholds, which do not capture the subjective experience of spontaneous pain, affective distress, and disability characteristic of CNSLBP.6,61,62
In human trials, the methodological shortcomings are substantial. A primary issue is the prevalence of underpowered studies with small, heterogeneous samples that include patients with diverse pain etiologies and comorbidities, compromising statistical power and generalizability.63,64 The inadequate blinding of participants and investigators is another major flaw, particularly for techniques that can produce perceptible sensations, making it difficult to control for placebo effects effectively.65 There is also a notable lack of standardization and personalization in stimulation parameters. Most studies apply stimulation to standardized scalp positions, ignoring individual variations in brain anatomy and functional connectivity, which likely contributes to the inconsistent therapeutic outcomes reported across studies.66,67 Finally, the field is hampered by a dearth of long-term follow-up data, leaving the crucial question of whether neuromodulation can induce lasting neuroplastic changes and sustained clinical benefits largely unanswered.68,69
A critical assessment of the current research paradigm reveals a fundamental disconnect between mechanistic inquiry and clinical validation. Many trials remain focused on preliminary efficacy testing without rigorously correlating neurophysiological biomarkers with clinical improvement to establish causal mechanisms.70 Moreover, the almost exclusive use of sham-controlled designs fails to demonstrate comparative effectiveness against established active treatments (eg, exercise therapy), thus questioning the added clinical added value of neuromodulation. Future research must prioritize large-scale, long-term, neuroimaging-guided, individualized RCTs and head-to-head comparisons with best-standard non-pharmacological treatments to overcome these hurdles.
Challenges and Future Directions: From Single-Target Interventions to Multimodal Synergies
A significant challenge in the contemporary management of chronic pain lies in the complexity of modulating multidimensional pathological networks through a singular intervention. Research indicates that targeted neuromodulation, informed by central sensitization6 and synaptic plasticity,71 offers symptomatic relief; however, it exhibits limited efficacy in addressing individual heterogeneity and the mechanisms of peripheral-central interaction. Future approaches should focus on integrating multimodal data, such as genomic information, functional imaging, and dynamic biomarkers, to develop cross-scale combination therapies. For instance, combining transcranial magnetic stimulation (TMS) with glutamatergic drugs72 could synergistically modulate the cerebral-peripheral axis. Additionally, optimizing intervention parameters of the ACC-mPFC loop in real-time through closed-loop neurofeedback73 is essential. Furthermore, patient stratification should be facilitated by artificial intelligence, and the long-term efficacy of these combined strategies must be validated in multicenter randomized trials. Ultimately, this will contribute to the establishment of an individualized pain management system grounded in systems biology.
Future Direction: Interdisciplinary Breakthroughs and Clinical Translation of iTBS Precision Regulation
Innovations in Precision Localization Techniques: From Brain Region Delineation to Sub-Millimeter Functional Network Targeting
The efficacy of iTBS is fundamentally rooted in the accurate localization of functional subregions within the ACC. The Individual Brain Functional Segmentation (pBFS) methodology, developed by Hesheng Liu’s team at the Changping Laboratory, employs machine learning algorithms to partition the brain into 213 distinct functional subregions, such as Brodmann areas 24a, 24b, and 24c of the ACC. This approach, when combined with resting-state functional magnetic resonance imaging (rs-fMRI) and Dynamic Causality Modeling (DCM), facilitates the identification of patient-specific aberrant network nodes.4 For example, in cases of post-stroke cognitive impairment (PSCI), precise targeting of the prefrontal-parietal cognitive network (FCN) has demonstrated a 33% enhancement in the hit rate of iTBS-induced fields and a 93% response rate.4 To further refine sub-millimeter stimulus localization, with an error margin of less than 0.5 mm, the integration of multimodal data, such as diffusion tensor imaging (DTI) for white matter fiber tracking and electroencephalography (EEG) γ power topography, is essential for constructing individualized “heat maps of neurological function”.74,75
Intelligent Regulation of Parameters: From Static Schemes to Dynamic Adaptive Systems
The customization of iTBS parameters necessitates moving beyond the conventional empirical model to establish a “stimulus-response” dynamic framework. The FPGA closed-loop system developed at Stanford University has successfully achieved millisecond-level monitoring of γ power and optimization of pulse timing. Specifically, when the γ power in the ACC exceeds 65 dB/Hz, the system automatically initiates a 50 Hz γ pulse sequence, resulting in a 42% reduction in pain intensity within 30 seconds.74 Future research directions include: (1) the development of a multi-parameter co-optimization algorithm that allows for the simultaneous adjustment of intensity (80–120% RMT), frequency (45–65 Hz), and pulse interval (15–25 ms) using the coyote optimization algorithm (COA), while avoiding locally optimal solutions through a chaotic weighting strategy;4 and (2) genotype-driven dose prediction, where the COMT Val158Met and BDNF Val66Met polymorphisms collectively predict the optimal stimulus intensity. Patients with the Met/Met genotype may require an increase in intensity up to 110% RMT to surpass the synaptic plasticity threshold.75,76
Multimodal Joint Interventions: From Single Target to Network-Level Synergistic Regulation
To establish a comprehensive “multi-target-multimodal” intervention network, iTBS should be integrated with other neuromodulation techniques and rehabilitation strategies. For instance: (1) iTBS combined with γ transcranial alternating current stimulation (γ-tACS) phase synchronization, where γ-tACS enhances the synaptic specificity of iTBS by externally modulating the phase of ACC γ oscillations with an error margin of less than 5 milliseconds.77 (2) iTBS paired with virtual reality (VR) attention training, where VR-induced theta-γ cross-frequency coupling facilitates information integration within the ACC-prefrontal network, leading to a decrease in pain catastrophizing scores and an extended duration of efficacy.4 (3) iTBS in conjunction with spinal cord magnetic stimulation (sTMS), targeting the bidirectional pathway between the spinal cord and ACC. The synchronized stimulation of the motor cortex (M1) and the lumbar region of the spinal cord enhances motor inhibition and pain perception through long-range synaptic plasticity linkage.78
From Animal Models to Clinical Translation: Mechanistic Consistency and Limitations
The clinical translation potential of CNSLBP neuro-modulation strategies depends on the synergistic validation of mechanistic studies in animal models and human clinical evidence. Mechanistic consistency is reflected in the cross-species commonality of disrupted excitatory/inhibitory (E/I) balance in the ACC. Both rodent models and human studies indicate that apoptosis of parvalbumin-positive interneurons (PV-INs) and glutamatergic hyperexcitability are key drivers of abnormal gamma oscillations. For example, neural injury in mice upregulates Cx36 gap junction protein in the ACC, enhancing γ synchrony.35 Similarly, iTBS exhibits cross-species universality in regulating GABAergic transmission: animal studies show that iTBS induces upregulation of GAD67 expression,52 consistent with enhanced short-interval intracortical inhibition (SICI) observed in human studies.13
However, translational challenges arise from species differences. First, pathological complexity: human CNSLBP not only manifests as nociceptive hypersensitivity but is also accompanied by psychological comorbidities such as anxiety and depression, as well as social behavioral factors, which are difficult to replicate in animal models. Second, network hierarchy differences: Rodent studies focus on molecular mechanisms, such as downregulation of Kv4.2 channels,39 while human neuroimaging reveals interactions between higher-order networks like the ACC and the DMN, where weakened ACC-DMN functional connectivity after iTBS correlates with analgesic effects.55 Third, intervention parameter calibration requires cross-species adjustment: mouse iTBS intensity (30–50% of human thresholds) must be adapted to skull impedance differences, while human protocols rely on resting-state functional connectivity (rs-fMRI) for individualized targeting.4 To bridge these gaps, future studies should establish a bidirectional translational framework. Animal models can simulate “anxiety-pain comorbidity” by manipulating the limbic circuit via optogenetics79,80 to approximate human phenotypes; clinically, multimodal biomarkers (eg, EEG γ maps, genomic analysis) should be integrated to enable patient stratification and iTBS parameter optimization.81,82 By integrating the depth of animal mechanisms with the breadth of clinical data, it is anticipated that precise neurostimulation strategies can be developed to advance CNSLBP treatment from symptom relief to neural plasticity restoration.
Technology Translation and Industrial Ecology: From Lab to Wearable Devices
The primary objective of future advancements is to facilitate the miniaturization and enhancement of intelligence in iTBS technology, aligning with prevailing trends across various technological domains. With the swift progression of artificial intelligence and Internet of Things (IoT) technologies, the trajectory of innovation in next-generation biosensing systems is increasingly oriented toward intelligence, miniaturization, and wireless portability (Q. Wang et al, 2023). For instance, researchers at Tsinghua University have developed a wireless patch, measuring 0.3 mm in thickness and weighing 5 g, that integrates fNIRS-EEG dual-modal sensing. This device is capable of continuously monitoring and regulating ACC γ oscillations in real time, while maintaining a power consumption of less than 1 mW.83,84
Conclusion
Pathologically enhanced gamma oscillations in the ACC constitute a mechanistic hallmark of central sensitization in CNSLBP. By physiologically entraining theta–gamma coupling, iTBS over the left DLPFC offers a time-efficient, mechanistically precise means to normalize this oscillatory dysfunction and restore cortical E/I balance. Although current evidence confirms short- to medium-term efficacy and excellent tolerability, durable disease modification will require personalized targeting, optimized protocols, and multimodal integration. These advances hold transformative potential to shift CNSLBP management from symptomatic palliation to reversal of maladaptive neuroplasticity.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was funded by the Natural Science Foundation of Hubei Province, grant number No.2023AFD024.
Disclosure
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Iizuka Y, Iizuka H, Mieda T. et al. Prevalence of Chronic Nonspecific Low Back Pain and Its Associated Factors among Middle-Aged and Elderly People: an Analysis Based on Data from a Musculoskeletal Examination in Japan. Asian Spine journal. 2017;11(6):989–997. doi:10.4184/asj.2017.11.6.989
2. Cohen SP, Vase L, Hooten WM. Chronic pain: an update on burden, best practices, and new advances. Lancet. 2021;397(10289):2082–2097. doi:10.1016/S0140-6736(21)00393-7
3. Özüdoğru A, Canlı M, Ceylan İ, Kuzu Ş, Alkan H, Karaçay BÇ. Five Times Sit-to-Stand Test in people with non-specific chronic low back pain—a cross-sectional test–retest reliability study. Ir J Med Sci. 2023;192(4):1903–1908. doi:10.1007/s11845-022-03223-3
4. Ren J, Su W, Zhou Y, et al. Efficacy and safety of high-dose and personalized TBS on post-stroke cognitive impairment: a randomized controlled trial. Brain Stimulation. 2025;18(2):249–258. doi:10.1016/j.brs.2025.02.009
5. Chenot JF, Greitemann B, Kladny B, Petzke F, Pfingsten M, Schorr SG. Non-Specific Low Back Pain. Dtsch Arztebl Int. 2017;2017:1. doi:10.3238/arztebl.2017.0883
6. Ji RR, Nackley A, Huh Y, Terrando N, Maixner W. Neuroinflammation and Central Sensitization in Chronic and Widespread Pain. Anesthesiology. 2018;129(2):343–366. doi:10.1097/ALN.0000000000002130
7. Pak DJ, Yong RJ, Kaye AD, Urman RD. Chronification of Pain: mechanisms, Current Understanding, and Clinical Implications. Curr Pain Headache Rep. 2018;22(2):9. doi:10.1007/s11916-018-0666-8
8. Pimenta DC, Cardenas-Rojas A, Camargo L, et al. Exercise effects on cortical excitability in pain populations: a systematic review and meta-analysis. Physiother Res Int. 2024;29(3):e2102. doi:10.1002/pri.2102
9. Baliki MN, Mansour AR, Baria AT, Apkarian AV. Functional reorganization of the default mode network across chronic pain conditions. PLoS One. 2014;9(9):e106133. doi:10.1371/journal.pone.0106133
10. Gross J, Schnitzler A, Timmermann L, Ploner M, Fries P. Gamma Oscillations in Human Primary Somatosensory Cortex Reflect Pain Perception. PLoS Biol. 2007;5(5):e133. doi:10.1371/journal.pbio.0050133
11. Schulz E, May ES, Postorino M, et al. Prefrontal Gamma Oscillations Encode Tonic Pain in Humans. Cereb Cortex. 2015;25(11):4407–4414. doi:10.1093/cercor/bhv043
12. Bliss TVP, Collingridge GL, Kaang BK, Zhuo M. Synaptic plasticity in the anterior cingulate cortex in acute and chronic pain. Nat Rev Neurosci. 2016;17(8):485–496. doi:10.1038/nrn.2016.68
13. Blumberger DM, Vila-Rodriguez F, Thorpe KE, et al. Effectiveness of theta burst versus high-frequency repetitive transcranial magnetic stimulation in patients with depression (THREE-D): a randomised non-inferiority trial. Lancet. 2018;391(10131):1683–1692. doi:10.1016/S0140-6736(18)30295-2
14. Hejazi HS, Khanmohammadi R, Olyaei G, Qanbari S. The effects of combining sensorimotor training with transcranial direct current stimulation on the anticipatory and compensatory postural adjustments in patients with chronic low back pain. Disabil Rehabil. 2025;47(6):1512–1524. doi:10.1080/09638288.2024.2375756
15. Kuzu Ş, Canli M, Valamur İ, Özüdoğru A, Alkan H, Hartavi A. Effects of aerobic exercise in addition to core stabilization exercises on functional capacity, physical performance and fall risk in geriatric individuals with chronic non-specific low back pain. BMC Sports Sci Med Rehabil. 2025;17(1):218. doi:10.1186/s13102-025-01271-7
16. Moisset X, Goudeau S, Poindessous-Jazat F, Baudic S, Clavelou P, Bouhassira D. Prolonged Continuous Theta-burst Stimulation is More Analgesic Than ‘Classical’ High Frequency Repetitive Transcranial Magnetic Stimulation. Brain Stimulation. 2015;8(1):135–141. doi:10.1016/j.brs.2014.10.006
17. Di Lazzaro V, Pilato F, Dileone M, et al. The physiological basis of the effects of intermittent theta burst stimulation of the human motor cortex. J Physiol. 2008;586(16):3871–3879. doi:10.1113/jphysiol.2008.152736
18. Guerra A, Suppa A, Bologna M, et al. Boosting the LTP-like plasticity effect of intermittent theta-burst stimulation using gamma transcranial alternating current stimulation. Brain Stimulation. 2018;11(4):734–742. doi:10.1016/j.brs.2018.03.015
19. Li C, Huang Y, Bai Y, Tsai S, Su T, Cheng C. Critical role of glutamatergic and GABAergic neurotransmission in the central mechanisms of theta‐burst stimulation. Human Brain Mapp. 2019;40(6):2001–2009. doi:10.1002/hbm.24485
20. De Wandel L, Pulopulos MM, Labanauskas V, De Witte S, Vanderhasselt M, Baeken C. Individual resting‐state frontocingular functional connectivity predicts the intermittent theta burst stimulation response to stress in healthy female volunteers. Human Brain Mapp. 2020;41(18):5301–5312. doi:10.1002/hbm.25193
21. Wang J, Wang J, Xing GG, Li X, Wan Y. Enhanced Gamma Oscillatory Activity in Rats with Chronic Inflammatory Pain. Front Neurosci. 2016;10:489. doi:10.3389/fnins.2016.00489
22. Buzsáki G, Wang XJ. Mechanisms of Gamma Oscillations. Annu Rev Neurosci. 2012;35(1):203–225. doi:10.1146/annurev-neuro-062111-150444
23. André-Obadia N, Magnin M, Garcia-Larrea L. Theta-burst versus 20 Hz repetitive transcranial magnetic stimulation in neuropathic pain: a head-to-head comparison. Clin Neurophysiol. 2021;132(10):2702–2710. doi:10.1016/j.clinph.2021.05.022
24. Guo F, Du Y, Qu FH, Lin SD, Chen Z, Zhang SH. Dissecting the Neural Circuitry for Pain Modulation and Chronic Pain: insights from Optogenetics. Neurosci Bull. 2022;38(4):440–452. doi:10.1007/s12264-022-00835-8
25. Zhuang X, Huang L, Gu Y, et al. The anterior cingulate cortex projection to the dorsomedial striatum modulates hyperalgesia in a chronic constriction injury mouse model. Arch Med Sci. 2019;17(5):1388–1399. doi:10.5114/aoms.2019.85202
26. Baliki MN, Petre B, Torbey S, et al. Corticostriatal functional connectivity predicts transition to chronic back pain. Nat Neurosci. 2012;15(8):1117–1119. doi:10.1038/nn.3153
27. Duan Y, Li Q, Zhou Y, Chen S, Li Y, Zang Y. Activation of the TNF-α-Necroptosis Pathway in Parvalbumin-Expressing Interneurons of the Anterior Cingulate Cortex Contributes to Neuropathic Pain. IJMS. 2023;24(20):15454. doi:10.3390/ijms242015454
28. Li QY, Duan YW, Zhou YH, Chen SX, Li YY, Zang Y. NLRP3-Mediated Piezo1 Upregulation in ACC Inhibitory Parvalbumin-Expressing Interneurons Is Involved in Pain Processing after Peripheral Nerve Injury. IJMS. 2022;23(21):13035. doi:10.3390/ijms232113035
29. Koga K, Shimoyama S, Yamada A, et al. Chronic inflammatory pain induced GABAergic synaptic plasticity in the adult mouse anterior cingulate cortex. Mol Pain. 2018;14:1744806918783478. doi:10.1177/1744806918783478
30. Zhu DY, Cao TT, Fan HW, et al. The increased in vivo firing of pyramidal cells but not interneurons in the anterior cingulate cortex after neuropathic pain. Mol Brain. 2022;15(1):12. doi:10.1186/s13041-022-00897-9
31. Huysmans E, Ickmans K, Van Dyck D, et al. Association Between Symptoms of Central Sensitization and Cognitive Behavioral Factors in People With Chronic Nonspecific Low Back Pain: a Cross-sectional Study. J Manipulative Physiological Therap. 2018;41(2):92–101. doi:10.1016/j.jmpt.2017.08.007
32. Nijs J, Apeldoorn A, Hallegraeff H, et al. Low back pain: guidelines for the clinical classification of predominant neuropathic, nociceptive, or central sensitization pain. Pain Physician. 2015;18(3):E333–346.
33. Nijs J, Torres-Cueco R, van Wilgen CP, et al. Applying modern pain neuroscience in clinical practice: criteria for the classification of central sensitization pain. Pain Physician. 2014;17(5):447–457.
34. Shyu BC, Vogt BA. Short-Term Synaptic Plasticity in the Nociceptive Thalamic-Anterior Cingulate Pathway. Mol Pain. 2009;5:1744–8069–5–51. doi:10.1186/1744-8069-5-51
35. Tan LL, Oswald MJ, Kuner R. Neurobiology of brain oscillations in acute and chronic pain. Trends Neurosci. 2021;44(8):629–642. doi:10.1016/j.tins.2021.05.003
36. Jiang Y, Oathes D, Hush J, et al. Perturbed connectivity of the amygdala and its subregions with the central executive and default mode networks in chronic pain. Pain. 2016;157(9):1970–1978. doi:10.1097/j.pain.0000000000000606
37. Wang Z, Wang Y, Ji Y, et al. Hypoconnectivity of the Amygdala in Patients with Low-Back-Related Leg Pain Linked to Individual Mechanical Pain Sensitivity: a Resting-State Functional MRI Study. JPR. 2023;16:3775–3784. doi:10.2147/JPR.S425874
38. Kang SJ, Kwak C, Lee J, et al. Bidirectional modulation of hyperalgesia via the specific control of excitatory and inhibitory neuronal activity in the ACC. Mol Brain. 2015;8(1):81. doi:10.1186/s13041-015-0170-6
39. Li XY, Ko HG, Chen T, et al. Alleviating neuropathic pain hypersensitivity by inhibiting PKMzeta in the anterior cingulate cortex. Science. 2010;330(6009):1400–1404. doi:10.1126/science.1191792
40. Peng W, Wang P, Tan C, et al. High-frequency terahertz stimulation alleviates neuropathic pain by inhibiting the pyramidal neuron activity in the anterior cingulate cortex of mice. Elife. 2024;13:RP97444. doi:10.7554/eLife.97444
41. Brewer GJ, Boehler MD, Jones TT, Wheeler BC. NbActiv4 medium improvement to Neurobasal/B27 increases neuron synapse densities and network spike rates on multielectrode arrays. J Neurosci Methods. 2008;170(2):181–187. doi:10.1016/j.jneumeth.2008.01.009
42. De Graaf TA, Gross J, Paterson G, Rusch T, Sack AT, Thut G. Alpha-Band Rhythms in Visual Task Performance: phase-Locking by Rhythmic Sensory Stimulation. PLoS One. 2013;8(3):e60035. doi:10.1371/journal.pone.0060035
43. Spaak E, De Lange FP, Jensen O. Local Entrainment of Alpha Oscillations by Visual Stimuli Causes Cyclic Modulation of Perception. J Neurosci. 2014;34(10):3536–3544. doi:10.1523/JNEUROSCI.4385-13.2014
44. Wang X, Popratiloff A, Motahari Z, LaMantia AS, Mendelowitz D. Disrupted Coordination of Hypoglossal Motor Control in a Mouse Model of Pediatric Dysphagia in DiGeorge/22q11.2 Deletion Syndrome. eNeuro. 2020;7(5):520. doi:10.1523/ENEURO.0520-19.2020
45. Keum JY, Toi PT, Park S, Chun H, Park JY. Direct imaging of neural activity reveals neural circuits via spatiotemporal activation mapping. Neuroscience. 2024. doi:10.1101/2024.07.31.606112
46. Nüssel M, Zhao Y, Knorr C, et al. Deep Brain Stimulation, Stereotactic Radiosurgery and High-Intensity Focused Ultrasound Targeting the Limbic Pain Matrix: a Comprehensive Review. Pain Ther. 2022;11(2):459–476. doi:10.1007/s40122-022-00381-1
47. Yan Y, Zhu M, Cao X, et al. Thalamocortical Circuit Controls Neuropathic Pain via Up-regulation of HCN2 in the Ventral Posterolateral Thalamus. Neurosci Bull. 2023;39(5):774–792. doi:10.1007/s12264-022-00989-5
48. Clancy KJ, Andrzejewski JA, Simon J, Ding M, Schmidt NB, Li W. Posttraumatic Stress Disorder Is Associated with α Dysrhythmia across the Visual Cortex and the Default Mode Network. eNeuro. 2020;7(4):53. doi:10.1523/ENEURO.0053-20.2020
49. Petschow C, Scheef L, Paus S, et al. Central Pain Processing in Early-Stage Parkinson’s Disease: a Laser Pain fMRI Study. PLoS One. 2016;11(10):e0164607. doi:10.1371/journal.pone.0164607
50. Cordeiro Matos S, Zhang Z, Séguéla P. Peripheral Neuropathy Induces HCN Channel Dysfunction in Pyramidal Neurons of the Medial Prefrontal Cortex. J Neurosci. 2015;35(38):13244–13256. doi:10.1523/JNEUROSCI.0799-15.2015
51. Guerra A, D’Onofrio V, Asci F, et al. Assessing the interaction between L‐dopa and γ ‐transcranial alternating current stimulation effects on primary motor cortex plasticity in Parkinson’s disease. Eur J Neurosci. 2023;57(1):201–212. doi:10.1111/ejn.15867
52. Guerra A, Asci F, D’Onofrio V, et al. Enhancing Gamma Oscillations Restores Primary Motor Cortex Plasticity in Parkinson’s Disease. J Neurosci. 2020;40(24):4788–4796. doi:10.1523/JNEUROSCI.0357-20.2020
53. Lu Q, Huang S, Zhang T, et al. Age-related differences in long-term potentiation-like plasticity and short-latency afferent inhibition and their association with cognitive function. Gen Psych. 2024;37(1):e101181. doi:10.1136/gpsych-2023-101181
54. Huang YZ, Edwards MJ, Rounis E, Bhatia KP, Rothwell JC. Theta Burst Stimulation of the Human Motor Cortex. Neuron. 2005;45(2):201–206. doi:10.1016/j.neuron.2004.12.033
55. Meng HJ, Cao N, Zhang J, Pi YL. Intermittent theta burst stimulation facilitates functional connectivity from the dorsal premotor cortex to primary motor cortex. PeerJ. 2020;8:e9253. doi:10.7717/peerj.9253
56. Chung SW, Rogasch NC, Hoy KE, Sullivan CM, Cash RFH, Fitzgerald PB. Impact of different intensities of intermittent theta burst stimulation on the cortical properties during TMS-EEG and working memory performance. Hum Brain Mapp. 2018;39(2):783–802. doi:10.1002/hbm.23882
57. Kohútová B, Fricová J, Klírová M, Novák T, Rokyta R. Theta Burst Stimulation in the Treatment of Chronic Orofacial Pain: a Randomized Controlled Trial. Physiol Res. 2017;2017:1041–1047. doi:10.33549/physiolres.933474
58. Yang C, Bi Y, Hu L, et al. Effects of different transcranial magnetic stimulations on neuropathic pain after spinal cord injury. Front Neurol. 2023;14:1141973. doi:10.3389/fneur.2023.1141973
59. Kim JK, Park HS, Bae JS, Jeong YS, Jung KJ, Lim JY. Effects of multi-session intermittent theta burst stimulation on central neuropathic pain: a randomized controlled trial. NeuroRehabilitation. 2020;46(1):127–134. doi:10.3233/NRE-192958
60. Drabek M, Hodkinson D, Horvath S, et al. B rain connectivity-guided, Optimised theta burst transcranial magnetic stimulation to improve Central Pain Modulation in knee Osteoarthritis Pain (BoostCPM): protocol of a pilot randomised clinical trial in a secondary care setting in the UK. BMJ Open. 2023;13(10):e073378. doi:10.1136/bmjopen-2023-073378
61. Corti EJ, Marinovic W, Nguyen AT, Gasson N, Loftus AM. Motor cortex excitability in chronic low back pain. Exp Brain Res. 2022;240(12):3249–3257. doi:10.1007/s00221-022-06492-7
62. Qanbari S, Khanmohammadi R, Olyaei G, Hosseini Z, Hejazi HS. Effects of combining sensory-motor exercises with transcranial direct current stimulation on cortical processing and clinical symptoms in patients with lumbosacral radiculopathy: an exploratory randomized controlled trial. PLoS One. 2024;19(12):e0314361. doi:10.1371/journal.pone.0314361
63. Olechowski C, Gener M, Aiyer R, Mischel N. Transcranial magnetic stimulation for the treatment of chronic low back pain: a narrative review. Front Pain Res. 2023;4:1092158. doi:10.3389/fpain.2023.1092158
64. Kirupaharan S, Milev R, Bressee J, et al. Changes in pain following bilateral intermittent theta-burst, transcranial magnetic stimulation for depression: a retrospective chart review. Can J Pain. 2024;8(1):2300026. doi:10.1080/24740527.2023.2300026
65. Bialosky JE, George SZ, Horn ME, Price DD, Staud R, Robinson ME. Spinal Manipulative Therapy–Specific Changes in Pain Sensitivity in Individuals With Low Back Pain (NCT01168999). J Pain. 2014;15(2):136–148. doi:10.1016/j.jpain.2013.10.005
66. Kong Q, Li T, Reddy S, Hodges S, Kong J. Brain stimulation targets for chronic pain: insights from meta-analysis, functional connectivity and literature review. Neurotherapeutics. 2024;21(1):e00297. doi:10.1016/j.neurot.2023.10.007
67. Ciampi de Andrade D, García-Larrea L. Beyond trial-and-error: individualizing therapeutic transcranial neuromodulation for chronic pain. Eur J Pain. 2023;27(9):1065–1083. doi:10.1002/ejp.2164
68. Shi Y, Wu W. Multimodal non-invasive non-pharmacological therapies for chronic pain: mechanisms and progress. BMC Med. 2023;21(1):372. doi:10.1186/s12916-023-03076-2
69. Yang J, Fu R, Hao Z, et al. The immediate effects of iTBS on the muscle activation pattern under challenging balance conditions in the patients with chronic low back pain: a preliminary study. Front Neurosci. 2023;17:1135689. doi:10.3389/fnins.2023.1135689
70. Chang TT, Chang YH, Du SH, Chen PJ, Wang XQ. Non-invasive brain neuromodulation techniques for chronic low back pain. Front Mol Neurosci. 2022;15:1032617. doi:10.3389/fnmol.2022.1032617
71. Zhuo M. Silent synapses in pain-related anterior cingulate cortex. Mol Pain. 2023;19:17448069231179011. doi:10.1177/17448069231179011
72. Benbow T, Cairns BE. Dysregulation of the peripheral glutamatergic system: a key player in migraine pathogenesis? Cephalalgia. 2021;41(11–12):1249–1261. doi:10.1177/03331024211017882
73. Etkin A, Egner T, Kalisch R. Emotional processing in anterior cingulate and medial prefrontal cortex. Trends Cognitive Sci. 2011;15(2):85–93. doi:10.1016/j.tics.2010.11.004
74. Cole EJ, Stimpson KH, Bentzley BS, et al. Stanford Accelerated Intelligent Neuromodulation Therapy for Treatment-Resistant Depression. Am J Psychiatry. 2020;177(8):716–726. doi:10.1176/appi.ajp.2019.19070720
75. Klooster DCW The Potential of Multimodal Neuroimaging to Personalize Transcranial Magnetic Stimulation Treatment Protocols. Phd Thesis 1 (Research TU/e / Graduation TU/e). Technische Universiteit Eindhoven; 2019.
76. Oteo I, Lukas JC, Leal N, et al. Pathophysiological idiosyncrasies and pharmacokinetic realities may interfere with tacrolimus dose titration in liver transplantation. Eur J Clin Pharmacol. 2011;67(7):671–679. doi:10.1007/s00228-011-0998-3
77. Lynch CJ, Breeden AL, Gordon EM, Cherry JBC, Turkeltaub PE, Vaidya CJ. Precision Inhibitory Stimulation of Individual-Specific Cortical Hubs Disrupts Information Processing in Humans. Cereb Cortex. 2019;29(9):3912–3921. doi:10.1093/cercor/bhy270
78. Patel D, Banerjee R, Farooque K, et al. Cortico-Spinal Intermittent Theta Burst Stimulation Propelling Sensorimotor Function Recovery in Complete Spinal Cord Injury: randomized Control Trial Protocol. Rehabil Med Phys Ther. 2024;14(1):e66531. doi:10.1101/2024.04.13.24305754
79. Ansari AH, Pal A, Ramamurthy A, Kabat M, Jain S, Kumar S. Fibromyalgia Pain and Depression: an Update on the Role of Repetitive Transcranial Magnetic Stimulation. ACS Chem Neurosci. 2021;12(2):256–270. doi:10.1021/acschemneuro.0c00785
80. Muir J, Lopez J, Bagot RC. Wiring the depressed brain: optogenetic and chemogenetic circuit interrogation in animal models of depression. Neuropsychopharmacology. 2019;44(6):1013–1026. doi:10.1038/s41386-018-0291-6
81. Aydemir O. Olfactory Recognition Based on EEG Gamma-Band Activity. Neural Comput. 2017;29(6):1667–1680. doi:10.1162/NECO_a_00966
82. Sims D, Sudbery I, Ilott NE, Heger A, Ponting CP. Sequencing depth and coverage: key considerations in genomic analyses. Nat Rev Genet. 2014;15(2):121–132. doi:10.1038/nrg3642
83. Baraeinejad B, Fallah Shayan M, Vazifeh AR, et al. Design and Implementation of an Ultralow-Power ECG Patch and Smart Cloud-Based Platform. IEEE Transact Instrument Measurement. 2022;6:5. doi:10.36227/techrxiv.17003401.v6
84. Lou Z, Tao J, Wei B, et al. Near-Infrared Organic Photodetectors toward Skin-Integrated Photoplethysmography-Electrocardiography Multimodal Sensing System. Adv Sci. 2023;10(36):e2304174. doi:10.1002/advs.202304174
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Interventional Pain Procedures: A Narrative Review Focusing On Safety and Complications. PART 2 Interventional Procedures For Back Pain
Lo Bianco G, Tinnirello A, Papa A, Marchesini M, Day M, Palumbo GJ, Terranova G, Di Dato MT, Thomson SJ, Schatman ME
Journal of Pain Research 2023, 16:761-772
Published Date: 9 March 2023