Back to Journals » International Journal of Women's Health » Volume 17
MiR-484 Regulates the IL-6/STAT3 Signaling Pathway by Down-Regulating KLF12 to Inhibit the Malignant Progression of Cervical Cancer
Authors Bao K, Zhang X, Bao L, Tao X, Zhang L, Wang D
Received 15 August 2024
Accepted for publication 14 April 2025
Published 26 April 2025 Volume 2025:17 Pages 1165—1174
DOI https://doi.org/10.2147/IJWH.S491749
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Elie Al-Chaer
Keyong Bao,1 Xue Zhang,1 Lihong Bao,2 Xiaoyu Tao,1 Li Zhang,1 Dong Wang3
1Department of Gynecology, Affiliated Hospital of Inner Mongolia University for the Nationalities, Tongliao, Inner Mongolia, 028000, People’s Republic of China; 2Prevention College of Inner Mongolia University for Nationalities, Tongliao, Inner Mongolia, 028000, People’s Republic of China; 3Department of Oncology, Affiliated Hospital of Inner Mongolia University for the Nationalities, Tongliao, Inner Mongolia, 028000, People’s Republic of China
Correspondence: Dong Wang, Department of oncology, Affiliated Hospital of Inner Mongolia University for the Nationalities, 1742, Huolinhe Street, Tongliao, Inner Mongolia, 028000, People’s Republic of China, Email [email protected]
Background: This study aimed to investigate the function of miR-484 in C33A cells, as well as its mechanism.
Methods: The mRNA expression patterns of miR-484 and Krüppel-like factor 12 (KLF12) in C33A cells were detected by quantitative reverse transcription polymerase chain reaction. MiR-484 mimics and miR-484 negative controls were transfected into C33A cells. Luciferase reporter assay was used to confirm the binding of miR-484 and KLF12. The effects of miR-484 on cell viability, migration, invasion, and apoptosis were assessed using the cell counting kit-8 assay, wound healing assay, Transwell assay, flow cytometry, and Western blot. Rescue experiments were performed by overexpressing KLF12. Western blot was utilized to examine the expression of KLF12, interleukin-6 (IL-6), Janus Kinase 2, phosphorylated-Janus Kinase 2, signal transducer and activator of transcription 3 (STAT3), and phosphorylated-STAT3 proteins.
Results: MiR-484 expression was down-regulated in C33A cells, while KLF12 was up-regulated. The luciferase reporter assay confirmed the direct binding of miR-484 to KLF12. MiR-484 inhibited the proliferation, migration, and invasion of C33A cells while promoting apoptosis. Additionally, it could inhibit the expression of KLF12 protein and the activation of the IL-6/STAT3 signaling pathway. However, KLF12 overexpression could reverse the above effects.
Conclusion: MiR-484 can specifically inhibit the KLF12-mediated IL-6/STAT3 signaling pathway, thereby suppressing the malignant biological behavior of C33A cells.
Keywords: cervical cancer, miR-484, KLF12, IL-6/STAT3 signaling pathway
Introduction
Cervical cancer (CC) is a well-known gynecological malignancy for residents, with a widespread global prevalence. Annually, over 500,000 new cases of CC are reported, with approximately 260,000 deaths, posing a major challenge to women’s health globally.1 The pathogenesis of CC is primarily attributed to human papillomavirus (HPV) infection, particularly high-risk HPV types 16 and 18, among others.2 The introduction of HPV vaccines has progressively reduced the incidence and mortality of CC worldwide. However, due to economic disparities among different countries and regions, HPV vaccination coverage remains incomplete.3 Additionally, the high incidence and mortality of CC are related to drug resistance, immune escape, metastasis, and invasion.4 The five-year survival rate for CC patients is only 16.5%.5 Therefore, further research into its pathogenesis is necessary to identify novel potential therapeutic targets.
MicroRNA (miRNA) has been found to play a crucial role in various tumor processes, including proliferation, invasion, apoptosis, migration, and angiogenesis. Therefore, investigating the mechanisms of miRNA is essential for exploring new cancer therapeutic strategies.6 Multiple studies have shown that miRNA-484 is down-regulated in various cancers, including CC, gastric cancer, colorectal cancer, and lung cancer.7,8 For example, interference with long non-coding RNA plasmacytoma variant translocation 1 may suppress the proliferation, invasion and epithelial-mesenchymal transition of CC cells (CCC) through miR-484 overexpression.9 miR-484 inhibits gastric cancer. By restraining the expression of CCL-18, miR-484 blocks the phosphoinositide 3-kinase/protein kinase B pathway by suppressing the expression of CC chemokine ligand 18, thereby inhibiting the malignant process of gastric cancer.8 Therefore, miR-484 plays a significant role in various cancers; however, its mechanism of action in CC remains limited. Additionally, Krüppel-like factor 12 (KLF12) was predicted as the target gene through TargetScan. A literature review revealed that KLF12 is significantly up-regulated in ovarian cancer tissues and cells10 and has been reported to promote the proliferation of CCC.11 However, studies on KLF12 in CC are limited. Therefore, the purpose of this study was to investigate the function of miR-484 and KLF12 and their relationship with the malignant biological behavior of CC.
Previous research has demonstrated that the interleukin-6 (IL-6)/signal transducer and activator of transcription 3 (STAT3) signaling pathway is highly expressed in CC.12 A preliminary finding indicates that KLF12-specific siRNA can significantly reduce IL-6/STAT3 activation.13 However, whether miR-484 can regulate the IL-6/STAT3 pathway by targeting KLF12 and participate in CC progression remains unclear. This study aimed to explore the relationship between miR-484 and KLF12, their roles in CC progression, and their underlying mechanisms, thereby providing experimental evidence for the mechanistic research and treatment of CC.
Materials and Methods
Cell Culture and Transfection
Human normal cervical epithelial cells (H8) and human CCC (C33A) were derived from the American Type Culture Collection. All cells were cultivated in a complete medium containing 10% fetal bovine serum (Excell Bio, China), 1% penicillin-streptomycin (Macklin, China), and 89% Dulbecco’s modified Eagle’s medium (Gibco, China).
MiR-484 mimics and their corresponding negative controls (miR-NC mimics) were synthesized and constructed by Ruibo Biotechnology Co., Ltd. The KLF12 overexpression (oe-KLF12) plasmid and its corresponding negative control (oe-NC) were synthesized and constructed by GenePharma. After cell counting, the cells were inoculated into a 6-well plate at a density of 1×106 cells/well and allowed to adhere until they reached 60–70% confluence. Lipofectamine 3000 reagent (Invitrogen, USA) was used for transfection according to the manufacturer’s instructions.
Cell Counting Kit-8
Following digestion, successfully transfected and non-transfected C33A cells were seeded into 96-well plates. Cell Counting Kit-8 reagent (final concentration: 10%; Bioss, China) was added to each well 2 h before the completion of incubation. After incubation, optical density at 450 nm was measured using an enzyme marker, and cell viability was calculated.
Wound Healing Assay
Cells were seeded into a 6-well plate and cultured until they reached 100% confluence. A pipette tip was then vertically pressed against the marked line on the back of the plate to create a scratch. Images of the scratch width were captured under a microscope at 0 h and 24 h, respectively.
Transwell Assay
A suitable amount of Matrigel diluent was added to the Transwell chamber, which was placed in a 24-well plate. Transfected/non-transfected C33A cells were digested and resuspended. The upper chamber was supplemented with 1 × 10⁵ cells. Then, 600 μL of medium containing 20% fetal bovine serum was added to the lower chamber. Afterward, the cells were cultured in an incubator at 37°C for 24 h. The un-invaded cells in the upper layer were gently removed, and the cells in the lower layer were fixed with 4% paraformaldehyde. Subsequently, the cells were stained with crystal violet (0.2%–0.5%) for 10 min, followed by three washes with pure phosphate-buffered saline. Ultimately, photographs were taken under a microscope and the cell invasion rate was calculated.
Flow Cytometry
Cells from different groups were collected and resuspended in an appropriate medium, followed by transfer into the tubes. Cells were then stained with propidium iodide and Annexin V-fluorescein isothiocyanate (Beyotime, China), and incubation was conducted. Lastly, 400 μL of 1 × Annexin V Binding Buffer was added, and the analysis was performed on a machine.14
Quantitative Reverse Transcription Polymerase Chain Reaction
Cell precipitation from each group was collected. Total RNA was extracted using a kit (Vazyme, China), and the RNA concentration and purity were determined. The RNA was converted to cDNA using a reverse transcription kit (Vazyme, China). The relative expression levels of genes were examined using a quantitative reverse transcription polymerase chain reaction kit (Vazyme, China) and calculated based on the cycle threshold values. The primers sequences used in this study are listed in Table 1.
 |
Table 1 Primer Sequence |
Western Blot
Cell precipitation from each group was collected, and total protein was extracted using radioimmunoprecipitation assay (Beyotime, China). The extracted proteins were then separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes (Sigma-Aldrich, USA). After sealing and washing, the membranes were incubated overnight at 4°C with primary antibodies, including B-cell lymphoma 2 (Bcl-2, ab182858, 1:20,000, Abcam), Bcl-2-associated X protein (ab32503, 1:20,000, Abcam), KLF12 (ab129459, 1:20,000, Abcam), IL-6 (ab233706, 1:20,000, Abcam), Janus Kinase 2 (JAK2, phospho Y1007 + Y1008, ab32101, 1:20,000, Abcam), STAT3 (phospho Y705, ab76315, 1:20,000, Abcam), JAK2 (ab108596, 1:20,000, Abcam), STAT3 (ab68153, 1:20,000, Abcam), and GAPDH (bsm-33033M, 1:50,000, Bioss). On the next day, the membranes were incubated with secondary antibodies (bs-0295G-HRP and bs-0296G-HRP, 1:20,000, Bioss), and the protein bands were detected and analyzed using an enhanced chemiluminescent luminescent solution kit (NCM Biotech, China).
Luciferase Reporter Assay
Luciferase reporter gene vectors (Sangon Biotech, China) were constructed with the KLF12 sequence (WT-KLF12) containing miR-1294 binding site and its mutant sequence (MUT-KLF12). Then, these vectors were co-transfected with miR-NC and miR-484 mimics, respectively. Luciferase activity in C33A cells was measured 48 h after transfection.
Results
Expression and Interaction of miR-484 and KLF12 in C33A Cells
The expression levels of both were assessed in H8 and C33A cells to investigate the impact of miR-484 and KLF12 in CCC. The results revealed that compared to H8 cells, the miR-484 expression was markedly decreased in C33A cells (P < 0.01), while the KLF12 mRNA expression was significantly raised (P < 0.01, Figure 1A and B). TargetScan was used to identify the binding site between miR-484 and KLF12 (Figure 1D). To validate the predicted results, luciferase reporter gene vectors containing either WT-KLF12 or MUT-KLF12 were co-transfected with the plasmid and miR-NC or miR-484 mimics into C33A cells. The results showed that miR-484 mimics inhibited the luciferase activity of WT-KLF12 in C33A cells (P < 0.001), with no effect observed on MUT-KLF12 (Figure 1C). These results indicated a negative regulatory relationship between miR-484 and KLF12.
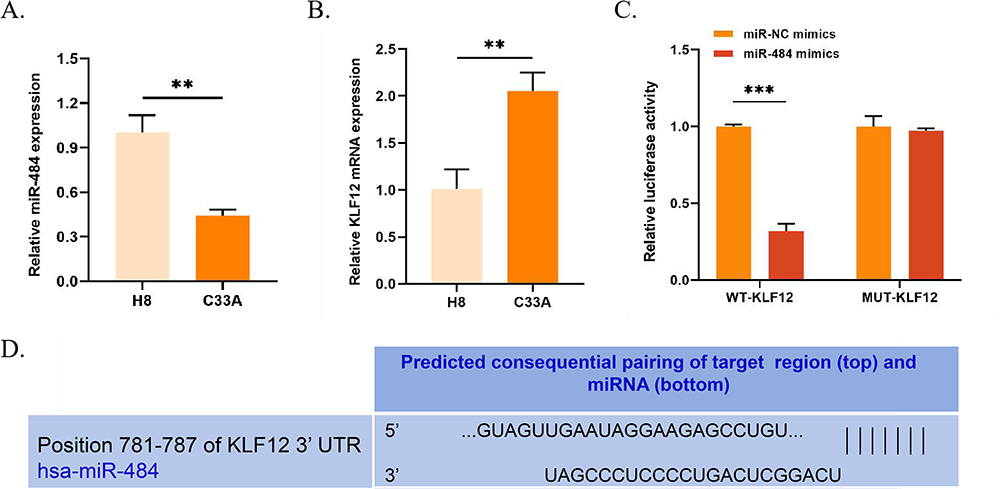 |
Figure 1 Expression and interaction of miR-484 and KLF12 in C33A cells. (A and B) The expression of miR-484 in H8 and KLF12 cells was assessed using qRT-PCR. (C) The luciferase reporter assay was used to assess the binding of miR-484 and KLF12. (D) TargetScan was utilized to detect the binding of miR-484 and KLF12. **P < 0.01, ***P < 0.001. Abbreviations: KLF12, Krüppel-like factor 12; qRT-PCR, quantitative reverse transcription polymerase chain reaction. |
Effects of miR-484 on the Malignant Biological Behavior of C33A Cells
C33A cells were transfected with miR-484 mimics or miR-NC mimics To further explore the relationship between miR-484 and the malignant progression of CCC. The results exhibited that the expression of miR-484 was remarkably higher in the miR-484 mimics group than that in the control group (P < 0.001), while no effect was observed in the miR-NC mimics group (Figure 2A). miR-484 mimics markedly reduced the proliferation, migration, and invasion of C33A cells (P < 0.01, Figure 2B–D). Additionally, the expression levels of Bcl-2-associated X protein were considerably up-regulated in the miR-484 mimics group (P < 0.05) while the expression levels of Bcl-2 protein were down-regulated (P < 0.05, Figure 2E). Overexpression of miR-484 mimics significantly increased the cell apoptosis rate compared to miR-NC mimics. Flow cytometry analysis showed a higher percentage of apoptotic cells in the miR-484 mimics group, indicated by increased populations in the Q2 and Q3 quadrants. The bar graph confirms a significant increase in apoptosis (P < 0.001, Figure 2F).
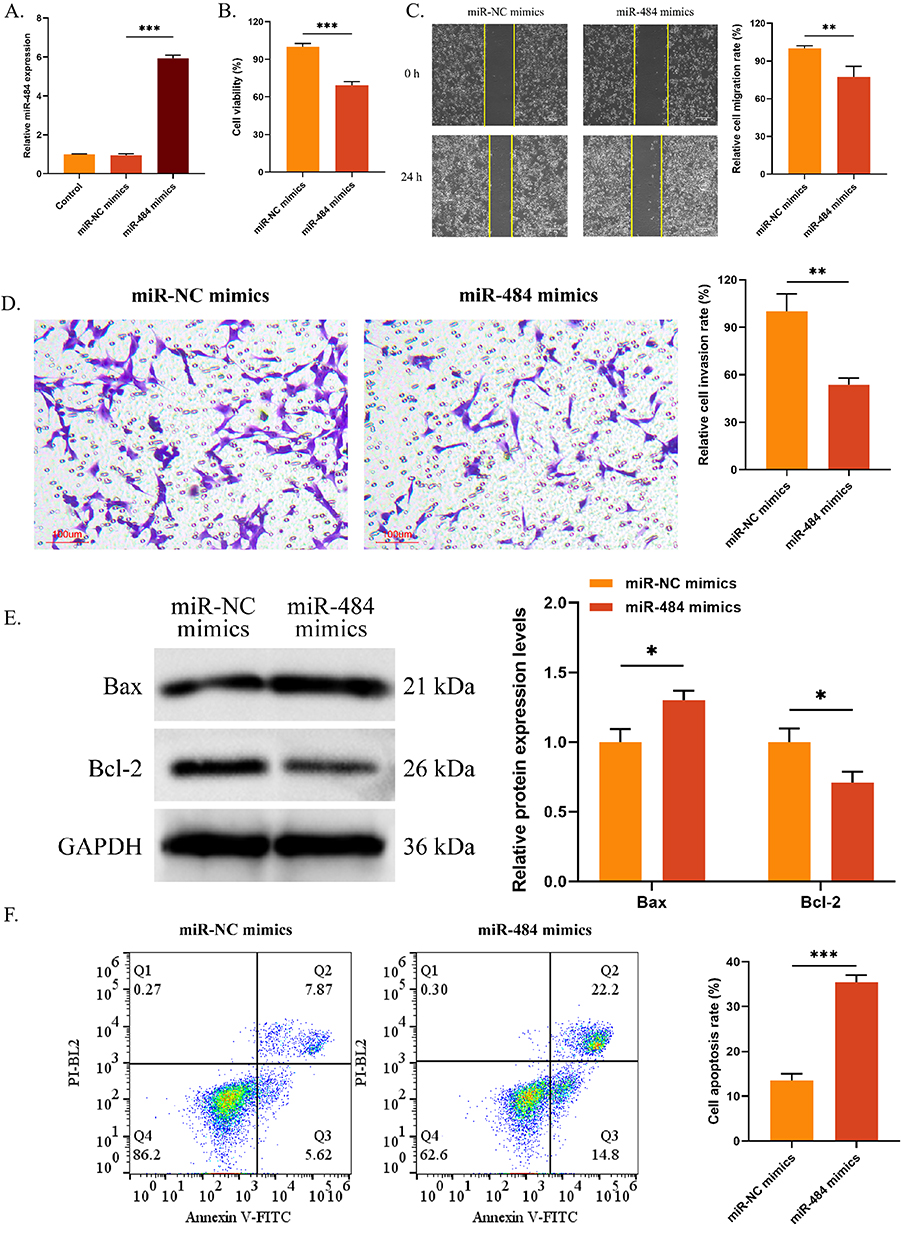 |
Figure 2 Effects of miR-484 on the malignant behaviors of C33A cells. (A) The transfection efficiency of miR-484 mimics was evaluated by qRT-PCR. (B) CCK-8 was used to detect the OD value. (C and D) Cell migration (C) and invasion (D) were measured by wound healing assay and Transwell assay, respectively. (E) The expression of Bax and Bcl-2 was assessed using Western blot. (F) Cell apoptosis was evaluated by flow cytometry. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: qRT-PCR, quantitative reverse transcription polymerase chain reaction; CCK-8, Cell Counting Kit-8; OD, optical density; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X protein. |
Effects of miR-484 on KLF12 Expression and the IL-6/STAT3 Pathway
Previously, an investigation was conducted on the interaction and negative synergistic effects between miR-484 and KLF12. Next, the impact of miR-484 on the expression of KLF12 protein was explored. miR-484 mimics or miR-NC mimics were transfected into C33A cells, and Western blot was employed to assess the expression of KLF12 protein. The results displayed that the protein expression of KLF12 was notably reduced in the miR-484 mimics group (P < 0.01, Figure 3). Knockdown of KLF12 expression has been reported to inhibit C33A cell growth by regulating the IL-6/STAT3 signaling pathway. However, the effect of miR-484 on the KLF12-mediated IL-6/STAT3 signaling pathway remains unclear. Therefore, Western blot was utilized to measure the expression levels of this pathway-associated proteins. The results revealed that miR-484 mimics remarkably decreased the expression of IL-6, phosphorylated (p)-JAK2, and p-STAT3 proteins (P < 0.01, Figure 3), whereas the expression of JAK2 and STAT3 remained unaffected. These findings indicate that miR-484 mimics inhibit the IL-6/STAT3 signaling pathway.
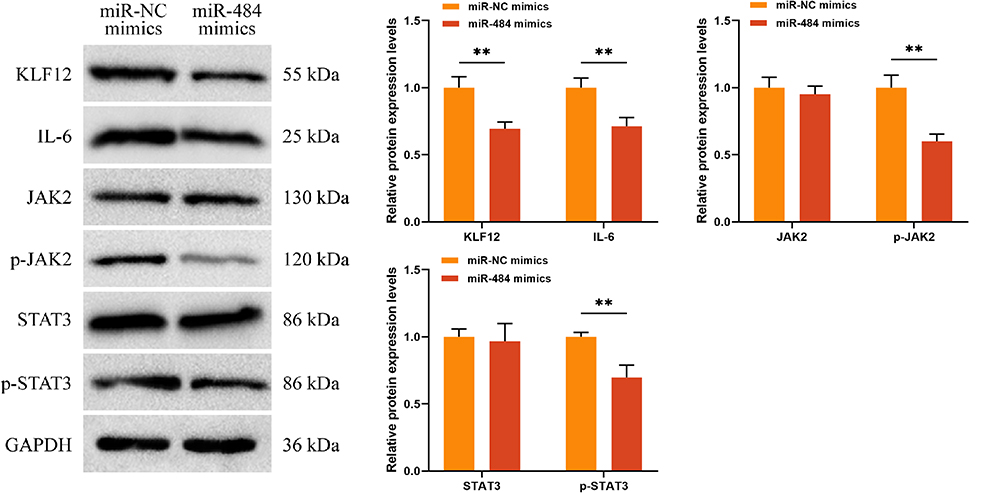 |
Figure 3 Effects of miR-484 on KLF12 expression and IL-6/STAT3 signaling pathway. Western blot was employed to measure the expression of KLF12, IL-6, p-JAK2, JAK2, p-STAT3, and STAT3 proteins. **P < 0.01. Abbreviations: KLF12, Krüppel-like factor 12; IL-6, interleukin-6; STAT3, signal transducer and activator of transcription 3; JAK2, Janus Kinase 2; p-JAK2, phosphorylated-JAK2; p-STAT3, phosphorylated-STAT3. |
MiR-484 Regulates the IL-6/STAT3 Signaling Pathway Through the Down-Regulation of KLF12 to Suppress the Malignant Biological Behavior of C33A Cells
The above results demonstrated that miR-484 mimics significantly inhibited C33A cell proliferation, migration, and invasion, promoted cell apoptosis, and markedly inhibited the protein expression of KLF12 as well as the activation of the KLF12-mediated IL-6/STAT3 signaling pathway. Rescue experiments were conducted to confirm whether miR-484 modulates the IL-6/STAT3 signaling pathway by regulating KLF12 to inhibit the malignant biological behavior of C33A cells. First, the transfection efficiency of KLF12 overexpression was examined by quantitative reverse transcription polymerase chain reaction. The results revealed that there was no significant difference in KLF12 mRNA between the control group and the oe-NC group (P < 0.05). Additionally, relative to the control group, the KLF12 mRNA and protein expression was significantly increased in the oe-KLF12 group (Figure S1). Subsequently, co-transfected miR-484 mimics and oe-KLF12 vectors were co-transfected into the cells. The malignant biological behavior of C33A cells was analyzed. The results indicated that oe-KLF12 reversed the inhibitory effects of miR-484 mimics on cell proliferation, migration, and invasion (P < 0.05, Figure 4A–C), as well as the promoting effect on apoptosis (P < 0.001, Figure 4D and E).
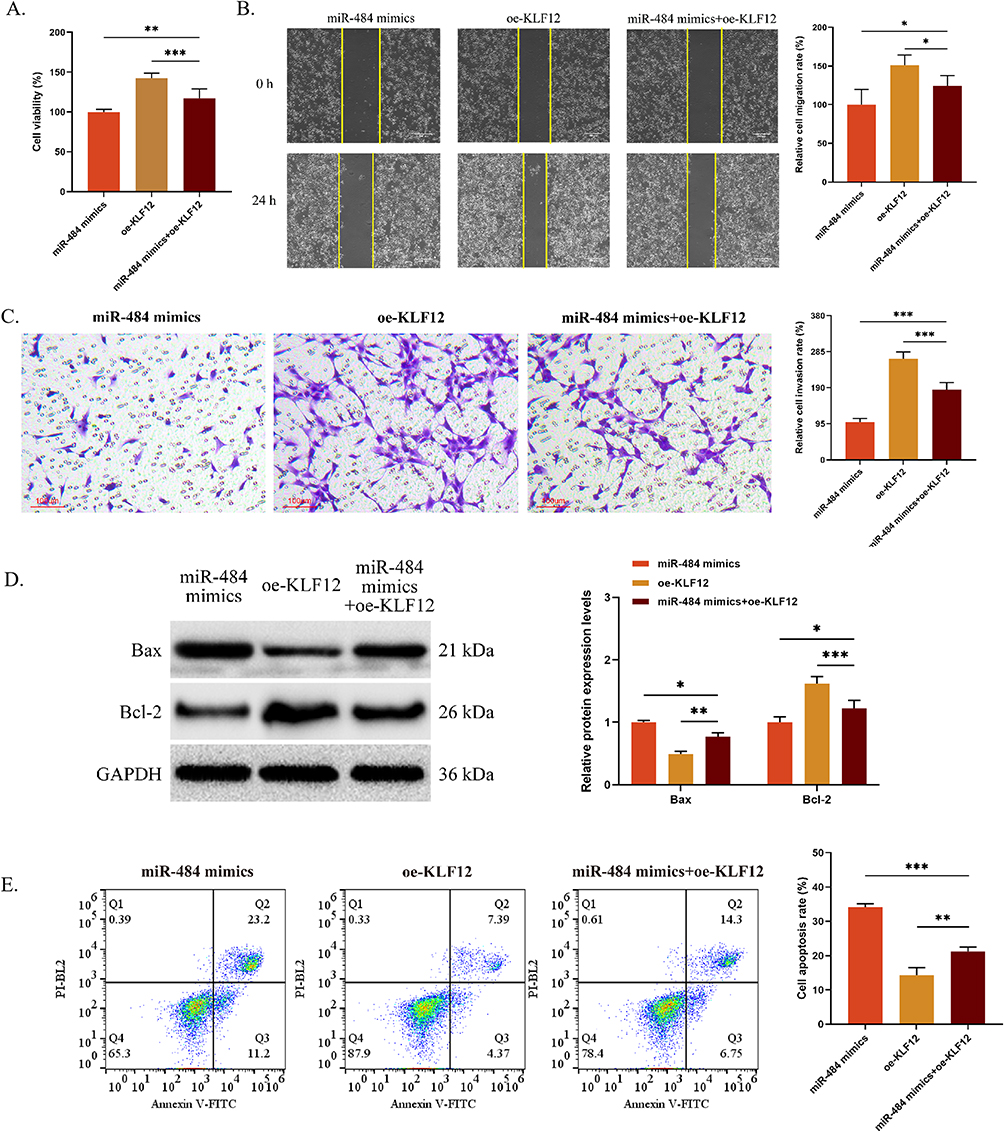 |
Figure 4 MiR-484 inhibits the malignant biological behaviors of C33A cells by down-regulating the KLF12 expression. (A) CCK-8 was used to detect the OD value. (B and C) Cell migration (B) and invasion (C) were determined using a wound healing assay and Transwell assay, respectively. (D) The expression of Bax and Bcl-2 proteins were assessed using Western blot. (E) Cell apoptosis was evaluated by flow cytometry. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: KLF12, Krüppel-like factor 12; CCK-8, Cell Counting Kit-8; OD, optical density; Bcl-2, B-cell lymphoma 2; Bax, Bcl-2-associated X protein. |
To validate our hypothesis, the pathway-associated proteins were detected. The above outcomes uncovered that miR-484 significantly inhibited the expression of KLF12 protein and the activation of the KLF12-mediated IL-6/STAT3 signaling pathway. Rescue experiments further revealed that oe-KLF12 reversed the inhibitory effect of miR-484 mimics on KLF12, IL-6 p-JAK2, and p-STAT3 protein levels in cells (Figure 5). Overall, miR-484 inhibited the malignant biological behavior of C33A cells by suppressing the expression of KLF12 and the IL-6/STAT3 signaling pathway.
 |
Figure 5 MiR-484 inhibits the IL-6/STAT3 signaling pathway by down-regulating the KLF12 expression. (A) The expression of KLF12, IL-6, p-JAK2, JAK2, p-STAT3, and STAT3 proteins was assessed using Western blot. **P < 0.01, ***P < 0.001. Abbreviations: IL-6, interleukin-6; STAT3, signal transducer and activator of transcription 3; KLF12, Krüppel-like factor 12; JAK2, Janus Kinase 2; p-JAK2, phosphorylated-JAK2; p-STAT3, phosphorylated-STAT3. |
Discussion
CC, a devastating disease, has emerged as the fourth leading cause of cancer-related death.15 Over the past few decades, significant progress has been made in the prevention and treatment of CC.16 In particular, the introduction of the HPV vaccine has played a crucial role in preventing CC. Hysterectomy, radiation therapy, and chemotherapy are the most commonly used treatments for early-stage CC. However, these methods fail to significantly enhance the 5-year survival rate of patients with middle and advanced-stage CC, and the prognosis of patients with CC is still unsatisfactory due to recurrence and metastasis.17–19 At present, the molecular basis of CC remains poorly understood. A deeper understanding of the molecular mechanisms of CC occurrence and development will help identify novel molecular therapeutic targets.
Currently, a substantial body of evidence demonstrates that miRNA plays a critical role in the malignant progression of CC. For example, miR-195-3p can inhibit the proliferation of CCC by directly targeting BCDIN3 domain containing RNA methyltransferase.20 MiR-362 inhibits CC progression by directly targeting B cell receptor associated protein 31 and activating the transforming growth factor-β/Sma- and Mad-related protein pathway.21 Consistent with previous studies, this study revealed that miR-484 was underexpressed in C33A cells, and miR-484 mimics could inhibit the proliferation, invasion, migration, and apoptosis of C33A cells. Furthermore, this study identified a therapeutic mechanism by which miR-484 regulated the development of CC.
KLF12 is up-regulated in a variety of cancers and promotes the proliferation of breast cancer cells by suppressing the P21 transcription.22 MiR-650 inhibits the metastasis of ovarian cancer by targeting KLF12 inhibition.10 Bioinformatics prediction and luciferase reporter assay confirmed that KLF12 was a target gene of miR-484, and there exists a negative regulatory relationship between them. Additionally, this study revealed that KLF12 was highly expressed in C33A cells, which was contrary to miR-484. To further validate the targeted relationship between miR-484 and KLF12, cells were transfected with miR-484 mimics. The results showed that miR-484 mimics significantly inhibited the expression of KLF12 protein. Therefore, we hypothesized that miR-484 may suppress the malignant biological behavior of CC by inhibiting the KLF12 expression. Subsequently, rescue experiments were performed, demonstrating that oe-KLF12 reversed the inhibitory effects of miR-484 mimics on the malignant progression of C33A cells.
Immunotherapy and targeted therapies, such as immune checkpoint inhibitors (eg, Pembrolizumab) and antibody-drug conjugates (eg, Tisotumab vedotin), have shown significant potential in treating recurrent and metastatic cervical cancer. Recent studies indicate that combining these therapies with chemotherapy has improved patient survival rates. Additionally, new pharmacological approaches, such as PARP inhibitors and Bevacizumab, are emerging as effective treatments.23 Despite these advances, current screening methods, such as Pap smears and HPV testing, still have limitations in detecting early-stage disease, and treatments like chemotherapy, radiation, and surgery have not significantly improved survival for patients with advanced or recurrent cervical cancer. Future studies could explore combining miR-484 with these novel therapies to further enhance treatment efficacy. miR-484’s ability to inhibit KLF12 and suppress the IL-6/STAT3 pathway could synergize with immune therapies and targeted treatments, offering a more comprehensive treatment strategy.24 Thus, miR-484 not only opens new avenues for cervical cancer therapy but also provides potential for improving the outcomes of novel therapeutic regimens.
Conclusion
The miR-484 expression is down-regulated in C33A cells, while the KLF12 expression is up-regulated. miR-484 can specifically inhibit the KLF12-mediated IL-6/STAT3 signaling pathway, thereby inhibiting the malignant biological behaviors of CCC.
Data Sharing Statement
The raw data used in this study have been uploaded in Figshare. The data used in this study is available at the following website.
Ethical Approval
This study and its experimental procedures were approved by the Affiliated Hospital of Inner Mongolia University for the Nationalities. The Ethics Committee of Affiliated Hospital of Inner Mongolia University for the Nationalities reviewed the informed consent procedure for the patients and confirmed that the study adhered to the ethical standards outlined in the 1964 Declaration of Helsinki.
Informed Consent
Written informed consent was obtained from all the participants prior to their enrollment in this study.
Acknowledgment
The authors thank the staff at the Affiliated Hospital of Inner Mongolia University for the Nationalities for their scientific advice and encouragement.
Funding
This study was supported by the Inner Mongolia Autonomous Region Natural Science Foundation Project (2022MS08077).
Disclosure
The authors declare that there are no competing interests in the submission of this manuscript, and the manuscript has been approved by all authors for publication.
References
1. Buskwofie A, David-West G, Clare CA. A review of cervical cancer: incidence and disparities. J Natl Med Assoc. 2020;112(2):229–232. doi:10.1016/j.jnma.2020.03.002
2. Twiggs LB, Hopkins M. High-risk HPV DNA testing and HPV-16/18 genotyping: what is the clinical application? J Low Genit Tract Dis. 2011;15(3):224–230. doi:10.1097/LGT.0b013e3181fb46d8
3. Tsu VD, Cernuschi T, LaMontagne DS. Lessons learned from HPV vaccine delivery in low-resource settings and opportunities for HIV prevention, treatment, and care among adolescents. J Acquir Immune Defic Syndr. 2014;66 Suppl 2(Supplement 2):S209–216. doi:10.1097/QAI.0000000000000175
4. Adiga D, Eswaran S, Pandey D, Sharan K, Kabekkodu SP. Molecular landscape of recurrent cervical cancer. Crit Rev Oncol Hematol. 2021;157:103178. doi:10.1016/j.critrevonc.2020.103178
5. Shrestha AD, Neupane D, Vedsted P, Kallestrup P. Cervical cancer prevalence, incidence and mortality in low and middle income countries: a systematic review. Asian Pac J Cancer Prev. 2018;19(2):319–324. doi:10.22034/APJCP.2018.19.2.319
6. Lee YS, Dutta A. MicroRNAs in cancer. Annu Rev Pathol. 2009;4(1):199–227. doi:10.1146/annurev.pathol.4.110807.092222
7. Miao J, Regenstein JM, Xu D, et al. The roles of microRNA in human cervical cancer. Arch Biochem Biophys. 2020;690:108480. doi:10.1016/j.abb.2020.108480
8. Liu J, Li SM. MiR-484 suppressed proliferation, migration, invasion and induced apoptosis of gastric cancer via targeting CCL-18. Int J Exp Pathol. 2020;101(6):203–214. doi:10.1111/iep.12366
9. Yan M, Pan XF, Liu Y, Zhao S, Gong WQ, Liu W. Long noncoding RNA PVT1 promotes metastasis via miR-484 sponging in osteosarcoma cells. Eur Rev Med Pharmacol Sci. 2020;24(5):2229–2238. doi:10.26355/eurrev_202003_20488
10. Lu X, Han Y, Han Y, et al. MicroRNA-650 suppresses KLF12 expression to regulate growth and metastasis of human ovarian cancer cells. Acta Biochim Pol. 2022;69(4):745–751. doi:10.18388/abp.2020_5987
11. Chen Y, Geng Y, Huang J, et al. CircNEIL3 promotes cervical cancer cell proliferation by adsorbing miR-137 and upregulating KLF12. Cancer Cell Int. 2021;21(1):34. doi:10.1186/s12935-020-01736-4
12. Hao Y, Yan Z, Zhang A, et al. IL-6/STAT3 mediates the HPV18 E6/E7 stimulated upregulation of MALAT1 gene in cervical cancer HeLa cells. Virus Res. 2020;281:197907. doi:10.1016/j.virusres.2020.197907
13. Bao K, Tao X, Zhang X. The expression of KLF12 in cervical cancer and the mechanism of regulating IL-6/STAT3 pathway affecting proliferation and apoptosis of cervical cancer cells. Chin J Immunol. 2021;37(16):1979–1983.
14. Qu Q, Liu L, Cui Y, et al. miR-126-3p containing exosomes derived from human umbilical cord mesenchymal stem cells promote angiogenesis and attenuate ovarian granulosa cell apoptosis in a preclinical rat model of premature ovarian failure. Stem Cell Res Ther. 2022;13(1):352. doi:10.1186/s13287-022-03056-y
15. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424. doi:10.3322/caac.21492
16. Zhang Y, Wu D, Wang D. Long non-coding RNA ARAP1-AS1 promotes tumorigenesis and metastasis through facilitating proto-oncogene c-Myc translation via dissociating PSF/PTB dimer in cervical cancer. Cancer Med. 2020;9(5):1855–1866. doi:10.1002/cam4.2860
17. Zhang M, Song Y, Yu L. LncRNA PTCSC3 suppressed cervical carcinoma cell invasion and proliferation via regulating miR-574-5p. Am J Transl Res. 2019;11(11):7186–7194.
18. Li YJ, Yang Z, Wang YY, Wang Y. Long noncoding RNA ZNF667-AS1 reduces tumor invasion and metastasis in cervical cancer by counteracting microRNA-93-3p-dependent PEG3 downregulation. Mol Oncol. 2019;13(11):2375–2392. doi:10.1002/1878-0261.12565
19. Wang AH, Jin CH, Cui GY, et al. MIR210HG promotes cell proliferation and invasion by regulating miR-503-5p/TRAF4 axis in cervical cancer. Aging. 2020;12(4):3205–3217. doi:10.18632/aging.102799
20. Jin M, Wang L, Zheng T, Yu J, Sheng R, Zhu H. MiR-195-3p inhibits cell proliferation in cervical cancer by targeting BCDIN3D. J Reprod Immunol. 2021;143:103211. doi:10.1016/j.jri.2020.103211
21. Yang S, Sun Y, Jiang D, et al. MiR-362 suppresses cervical cancer progression via directly targeting BAP31 and activating TGFbeta/Smad pathway. Cancer Med. 2021;10(1):305–316. doi:10.1002/cam4.3601
22. Li Y, Li S, Shi X, et al. KLF12 promotes the proliferation of breast cancer cells by reducing the transcription of p21 in a p53-dependent and p53-independent manner. Cell Death Dis. 2023;14(5):313. doi:10.1038/s41419-023-05824-x
23. D’Oria O, Bogani G, Cuccu I, et al. Pharmacotherapy for the treatment of recurrent cervical cancer: an update of the literature. Expert Opin Pharmacother. 2024;25(1):55–65. doi:10.1080/14656566.2023.2298329
24. Ferrari F, Giannini A. Approaches to prevention of gynecological malignancies. BMC Women’s Health. 2024;24(1):254. doi:10.1186/s12905-024-03100-4
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
RCN1 Binds KIF14 and Promotes the Malignant Growth of Cervical Cancer Through the PI3K-AKT Pathway
Li Y, Cai L, Zhou J, Zhang X, Zheng Y, Zhang J, Cheng H, Wang Q, Zhang B
International Journal of General Medicine 2025, 18:5047-5062
Published Date: 2 September 2025