Back to Journals » Journal of Inflammation Research » Volume 19
MicroRNAs in Asthma: A Replication-Based Review of Human Evidence
Authors Hadikhani P ![]() , Piparia S, Sharma R
, Piparia S, Sharma R ![]() , McGeachie M, Tantisira KG
, McGeachie M, Tantisira KG
Received 23 December 2025
Accepted for publication 23 April 2026
Published 28 May 2026 Volume 2026:19 585667
DOI https://doi.org/10.2147/JIR.S585667
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Dr Cynthia Koziol-White
Parham Hadikhani,1 Shraddha Piparia,1 Rinku Sharma,2 Michael McGeachie,2 Kelan G Tantisira1,3
1Department of Pediatrics, Division of Pediatric Respiratory Medicine, University of California San Diego, San Diego, CA, USA; 2Channing Division of Network Medicine, Department of Medicine, Brigham and Women’s Hospital and Harvard Medical School, Boston, MA, USA; 3Rady Children’s Hospital, San Diego, CA, USA
Correspondence: Parham Hadikhani, Email [email protected]
Abstract: Asthma is a heterogeneous inflammatory airway disease characterized by diverse endotypes, variable clinical severity, and differential treatment responses driven by genetic, epigenetic, and environmental influences. MicroRNAs (miRNAs), as post-transcriptional regulators of gene expression, are central modulators of airway inflammation, remodeling, and immune dysregulation and represent promising candidates for biomarker development and therapeutic targeting. Despite the growing literature on dysregulated miRNAs in asthma, findings remain inconsistent and frequently lack independent validation. Using a structured search strategy covering 2009– 2025, we identified 299 records and applied predefined replication criteria to prioritize reproducible associations. Two complementary validation Criteria were implemented: cross-study replication across independent cohorts and discovery–replication study designs addressing clinical heterogeneity. This approach yielded 49 cross-study replicated and 10 discovery–replication validated journal articles, substantially narrowing the field to a focused set of high-confidence miRNA candidates. Among the most consistently replicated miRNAs, miR-146a and miR-1246 emerged as central regulators linked to NF-κB signaling, epithelial dysfunction, and corticosteroid responsiveness. Replicated miRNAs demonstrated compartment-specific expression across airway epithelium, smooth muscle, immune cells, and circulating fractions, and were associated with clinically relevant dimensions including disease severity, exacerbation frequency, lung function variability, and treatment response. Genetic, epigenetic, and environmental influences further support their biological plausibility. By consolidating independent replication evidence, this review defines a curated set of robust asthma-associated miRNAs supported by convergent data across populations and phenotypes. These findings provide a basis for future efforts in endotype-aware biomarker development and inform future strategies toward miRNA-guided precision diagnostics and therapeutic interventions in asthma.
Keywords: asthma, MicroRNAs, epigenetic regulation, biomarkers, asthma severity, airway inflamma-tion, asthma endotypes
Introduction
Asthma is a common complex chronic respiratory disease that is heterogeneous, with a variety of phe-notypes, endotypes, and severity scales.1 It affects about 23.4 million individuals in the United States, including 7 million children,2 and over 300 million people globally live with asthma,3 which represents a significant and persistently evolving public health burden. The climate and environmental changes seen over the past several years have led to an increase in asthma incidence and mortality. In addition to these factors, changes associated with improved hygiene practices and western lifestyles, including increased urbanization, have also been implicated in the rising prevalence of asthma.4 Severe asthma affects approximately 5% of patients and is associated with increased morbidity, a poorer quality of life, and greater healthcare costs.5,6
The development and progression of asthma is driven by several factors, including exposures to environ-mental agents, genetic polymorphisms, and associated epigenetic changes, all of which contribute to the clinical heterogeneity and the responses to therapy.7 One such driving factor is microRNA (miRNA), regulation of gene expression. miRNAs are small non-coding RNA molecules which bind messenger RNA transcripts using base-pair complementarity, and then interfere with the translation of those mRNA into proteins by isolating and degrading them. miRNAs are present in biological fluids and are more stable compared to messenger RNA (mRNA), enabling for their post-transcriptional regulatory functions and potential as biomarkers.8,9 The stability of circulating miRNAs is largely attributed to their association with proteins such as HDL and Argonaute or their encapsulation within small vesicles such as exosomes. In addition to their intracellular roles, miRNAs also act as mediators of intercellular communication, functioning as long-range regulators that facilitate cell-to-cell signaling. miRNAs packaged in exosomes and emitted from activated immune cells, such as specific T-cell populations, serve as a window to the airway inflammation status.10,11
In terms of functionality, miRNAs impact many biological processes relevant to asthma, such as airway epithelial differentiation, cytokine production, immune modulation, and airway smooth muscle (ASM) remodeling.12 A number of studies have found that dysregulation of specific miRNAs promotes inflamma-tory persistence, steroid resistance, and phenotypic heterogeneity across asthma subtypes.6,13–16 These characteristics make miRNAs interesting mediators, potential biomarkers, and even therapeutic targets.
Because miRNAs regulate multiple pathways involved in airway inflammation, immune signaling, and airway remodeling, understanding their dysregulation may provide insight into asthma pathogenesis and identify potential biomarkers or therapeutic targets. However, despite the growing number of studies reporting altered miRNAs in asthma, important gaps remain. Many findings come from small and diverse cohorts and have not been independently replicated, leading to inconsistent results across studies. Differ-ences in sample types, measurement methods, analysis approaches, and patient populations further limit direct comparison and make it difficult to determine whether reported miRNAs reflect true disease biology or study-specific effects.1,12,16 In addition, previous reviews have mainly listed miRNAs associated with asthma without clearly separating those linked to important clinical features such as disease severity, exacerbations, lung function, or treatment response.12,15,17 To date, no review has systematically required replication as a key inclusion criterion to identify a high-confidence set of miRNAs supported by evidence from independent studies. The present review was designed to fill this gap by focusing on reproducibility and clinical relevance.
miRNA Formation and Biogenic Pathways
miRNAs are synthesized through a series of tightly coordinated steps involving nuclear and cytoplasmic processing (Figure 1). They are transcribed mainly by RNA polymerase II as long primary transcripts (pri-miRNAs) derived from intronic, exonic, or intergenic regions.18,19 Some are organized in clusters and co-transcribed as polycistronic units with shared seed regions and functions.19
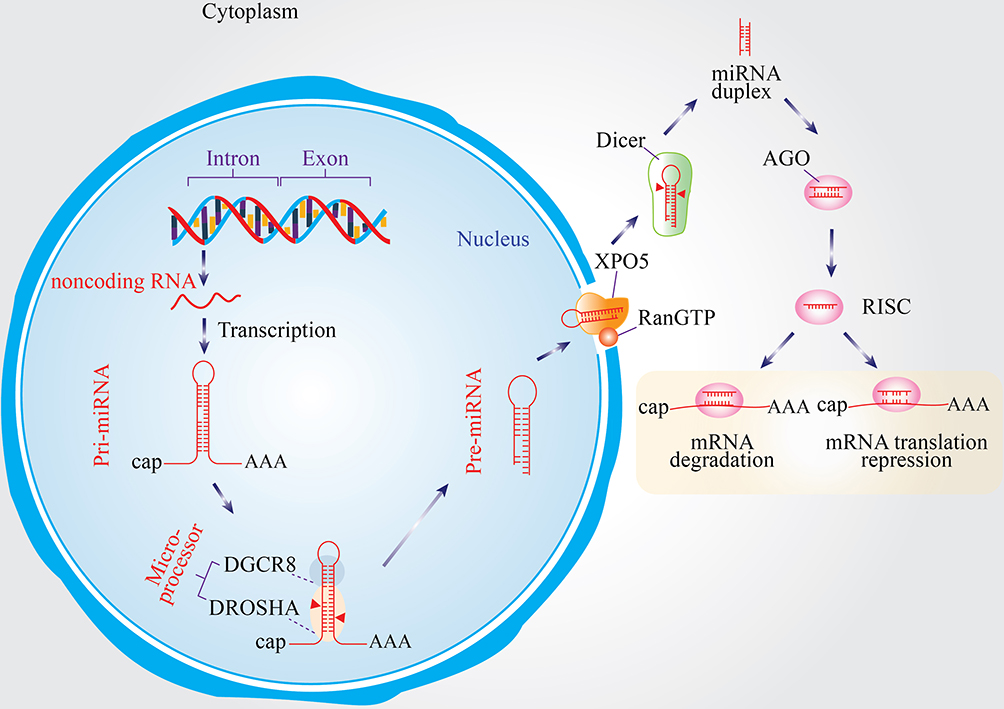 |
Figure 1 Biogenesis and mechanism of miRNAs. pri-miRNAs are transcribed and processed by the DROSHA–DGCR8 complex into pre-miRNAs in the nucleus. After export to the cytoplasm via XPO5/RanGTP, Dicer cleaves pre-miRNAs into duplexes, from which one strand is incorporated into the AGO-containing RISC complex to guide mRNA degradation or translational repression. |
In the nucleus, pri-miRNAs form a distinctive hairpin structure and are processed by the Drosha–DGCR8 Microprocessor complex into ∼70-nt precursor miRNAs (pre-miRNAs). The pre-miRNAs are shuttled to the cytoplasm by Exportin-5 (XPO5) in a RanGTP-dependent manner. Once in the cytoplasm, Dicer processes pre-miRNA into a ∼22-nt duplex, where the guide strand is assembled into the RNA-induced silencing complex (RISC) with Argonaute (AGO) proteins, and the passenger strand is degraded. The miRNA–RISC complex interacts with mRNAs that are complementary to the miRNA, ultimately inducing either degradation of the mRNA or translational repression.20–22
miRNA maturation is governed by two main pathways: the canonical (Drosha–DGCR8 → Exportin-5 → Dicer → RISC) and non-canonical pathways, the latter which can bypass Drosha or Dicer (eg, mirtrons, tRNase Z–based processing).22 These alternative pathways augment regulatory complexity and expression diversity of miRNAs.
In addition to acting intracellularly, miRNAs are released in exosomes and other extracellular vesicles (EVs) allowing for intercellular communication between epithelial, immune, and smooth muscle cells. miRNAs have a unique stability in biofluids and demonstrate specific expression patterns among cell-types, traits which make them attractive potential biomarkers and therapeutic targets in asthma.23–28
Aims of This Review
Building on prior summaries of miRNAs in asthma,3,12,17,29–32 this review aims to move beyond sim-ply listing reported associations by emphasizing replication across independent studies and integrating mechanistic and clinical perspectives. Specifically, we aim to:
- Describe the functions of asthma-associated miRNAs in airway inflammation, remodeling, and immune regulation.
- Summarize phenotypic and tissue-specific miRNA expression patterns across clinical and experi-mental models.
- Explore how environmental and lifestyle factors affect miRNA regulation and disease outcomes.
- Highlight reproducible miRNAs validated across independent cohorts, supported by mechanistic evidence, and with potential clinical utility.
Methods
Literature Search Strategy
To identify biologically robust miRNAs implicated in asthma, we conducted a structured literature search using PubMed. The search included studies published between 2009 and 2025. The following search query was applied:
(“microRNA”[MeSH] OR “miRNA”[Title/Abstract] OR “micro RNA”[Title/Abstract]) AND (“asthma”[MeSH] OR “asthmatic”[Title/Abstract])
A total of 299 records were initially identified. Search results were restricted to human studies to ensure clinical relevance. Titles and abstracts were screened for relevance, followed by full-text review of potentially eligible studies to confirm replication status and study design (Figure 2). This review employed predefined eligibility criteria to prioritize reproducibility and biological robustness.
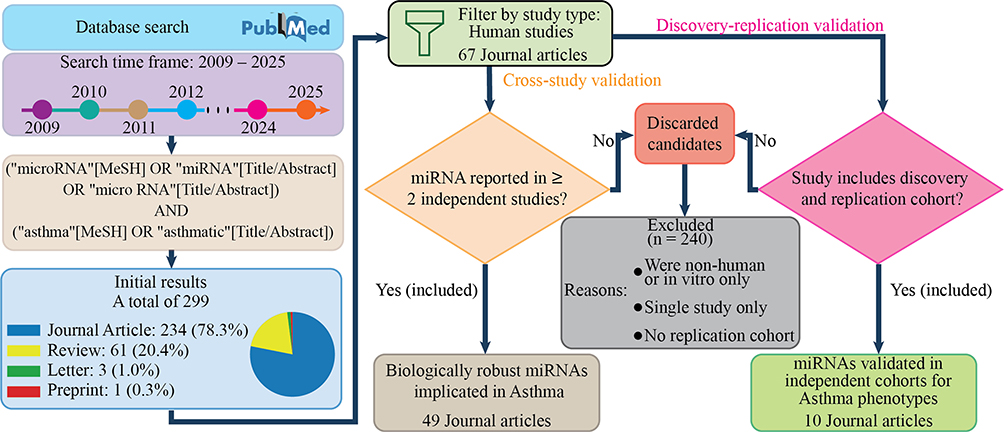 |
Figure 2 Overview of the structured literature search and dual replication-based selection strategy (2009–2025). A total of 299 records were identified through PubMed and restricted to human studies. After screening and application of predefined replication criteria, 240 studies were excluded. Two complementary validation approaches were applied: (1) cross-study replication (miRNAs independently reported in ≥2 publications), resulting in 49 included journal articles; and (2) discovery–replication validation (miRNAs validated in independent replication cohorts), resulting in 10 included journal articles. |
Eligibility Criteria
Studies were included if they:
- involved human subjects with asthma or asthma-related phenotypes;
- evaluated miRNA expression in relation to asthma diagnosis, severity, exacerbations, lung function, or treatment response; and
- provided evidence of independent replication, either through cross-study validation or discov-ery–replication cohort designs.
Studies were excluded if they:
- involved non-human models or in vitro experiments only;
- reported findings from a single cohort without independent replication;
- lacked sufficient methodological detail to determine replication status; or
- focused exclusively on non-asthma respiratory diseases without asthma-specific analysis.
Based on these criteria, 240 studies were excluded, primarily due to absence of replication evidence, non-human design, or lack of independent validation.
Replication Criteria
To minimize cohort-specific or methodological bias and to enhance biological confidence, we priori-tized miRNAs supported by replication across independent populations. Two complementary validation strategies were applied:
- Cross-study replication: miRNAs independently identified in at least two separate peer-reviewed publications involving non-overlapping patient cohorts, distinct study populations, and independent analytical pipelines.
- Discovery–replication design: miRNAs identified in a discovery cohort and subsequently validated in an independent replication cohort within the same study.
Replication was defined as confirmation of directionally consistent association across independent study populations; it was not intended to imply causal validation.
Abstracts and full texts were manually reviewed to identify explicit evidence of replication, including terminology such as “replication,” “validation,” “independent cohort,” “discovery and validation,” or “multiple cohorts.” Eligible studies included those examining asthma diagnosis (case–control compar-isons), disease severity, exacerbation frequency, lung function parameters, incident asthma prediction, and treatment response.
Application of this dual replication Criteria resulted in 49 journal articles meeting cross-study replication criteria and 10 journal articles meeting discovery–replication validation criteria.
Risk of Bias and Methodological Considerations
A formal risk-of-bias assessment was not performed because this review did not combine data or calculate pooled effect sizes. Instead, we used independent replication across different cohorts as an indicator of robustness and reproducibility. Differences in sample type, measurement platforms, data processing methods, and patient characteristics were taken into account during the qualitative interpretation of the findings. This structured, replication-focused Criteria was designed to emphasize reproducibility and strengthen biological plausibility rather than to provide an exhaustive catalog of all reported miRNA associations in asthma.
Search Results
The search approach presented in the Method section led to the identification of significant asthma-related miRNA studies and a core list of biologically relevant and consistently replicated candidates. The top ten most replicated miRNAs are highlighted in Figure 3, emphasizing the reproducibility of these findings. In particular, miR-146a and miR-1246 demonstrate high levels of replication across studies, underscoring their potential as key regulators in asthma pathogenesis. By concentrating on replication, we reduce study-to-study variability and establish a robust foundation for subsequent analyses of mechanistic functions, tissue-specific expression patterns, and clinical relevance.
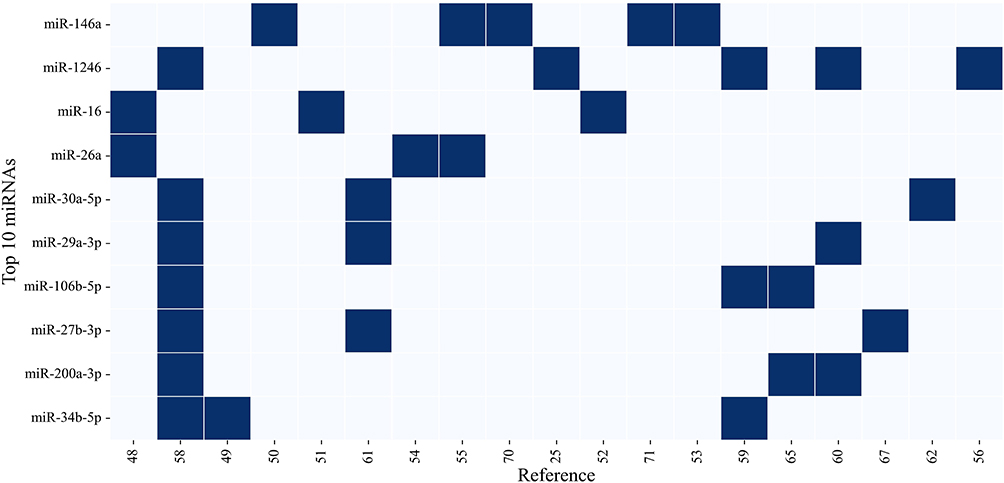 |
Figure 3 Top 10 most replicated miRNAs in asthma literature. Deep blue squares indicate miRNA presence across publications, highlighting independent replication. miRNAs are ranked top to bottom by citation frequency. |
Table 1 presents asthma-associated miRNAs that have been consistently replicated across multiple inde-pendent studies comparing asthmatic patients to healthy controls. These miRNAs were identified through external validation—meaning they were independently discovered and reported in at least two separate publications, each with its own cohort and methodology. For each miRNA, the table provides the reference studies, replication counts, and sample sizes for asthmatic and control cohorts. This cross-study replication approach allows readers to evaluate the reproducibility of each miRNA’s association with asthma presence across different research groups, populations, and experimental conditions.
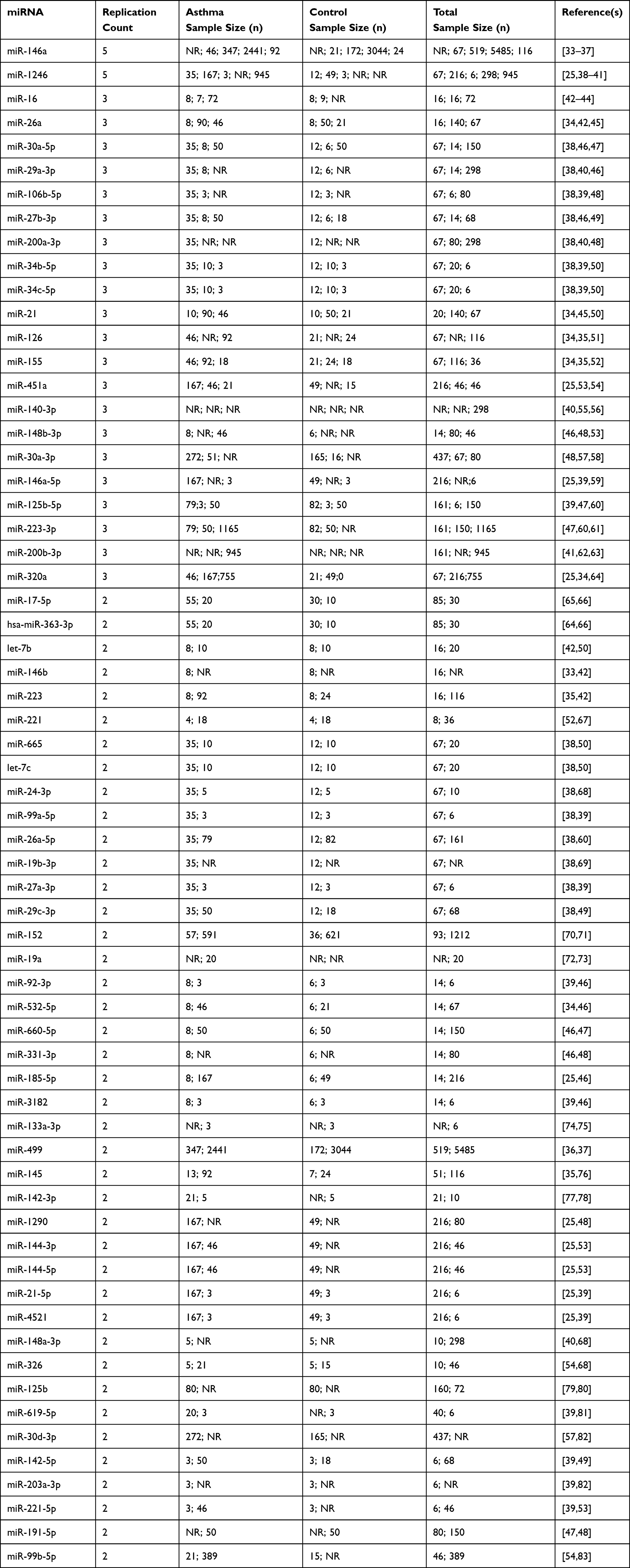 |
Table 1 Asthma-Associated miRNAs Replicated Across Multiple Studies, Including Reference Studies, Replication Counts, and Sample Sizes for Asthmatic and Control Cohorts, as Well as the Total Number of Participants. NR Indicates That the Sample Size Was Not Reported in the Original Study |
Table 2 takes a complementary but distinct approach, presenting miRNAs replicated through prospective discovery–replication study designs that examine asthma’s clinical heterogeneity—including variation in severity, symptoms, exacerbations, lung function, and treatment response. Unlike Table 1’s cross-study validation, Table 2 focuses on miRNAs that were identified in a discovery cohort and then independently validated in a separate replication cohort within the same study or research program. This rigorous within-study validation approach spans multiple clinically important phenotypic dimensions:
- disease severity (severe uncontrolled vs mild asthma),
- exacerbation frequency (frequent vs infrequent exacerbators), with cross-study replication (external replication) in independent adult COPD cohorts,
- asthma symptoms and disease progression through incident asthma prediction in children with recurrent wheeze,
- lung function measures including bronchodilator response, eosinophilia, airway hyperresponsiveness, and airflow obstruction, and
- treatment response to inhaled corticosteroids and bronchodilators, including studies examining vitamin D effect modification and functional validation through NF-κB pathway assays.
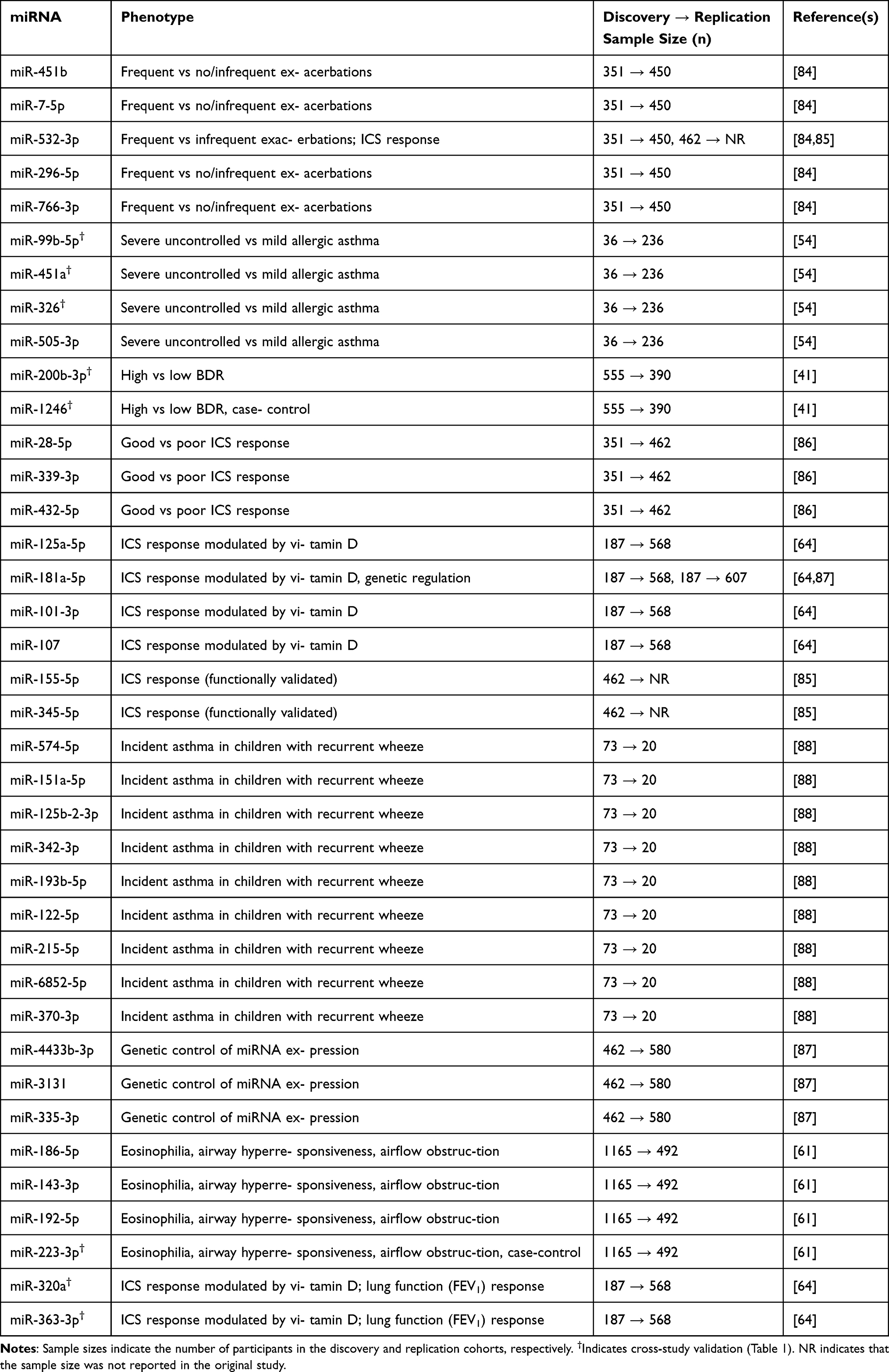 |
Table 2 miRNAs Validated Through Discovery → Replication Designs Examining Asthma Severity, Exacerbations, Lung Function, and Treatment Response |
Sample sizes are presented in a discovery cohort → replication cohort format to emphasize this sequential validation process. Several studies achieved cross-age validation, replicating findings from pediatric discovery cohorts in independent adult replication cohorts or vice versa. Notably, several miRNAs (marked with †) appear in both tables, having been validated through both external cross-study replication (Table 1) and prospective discovery–replication designs (Table 2). This dual validation across independent replication strategies indicates that the same miRNAs implicated in asthma susceptibility also contribute to the molecular pathways driving exacerbation severity, underscoring their mechanistic and clinical relevance.
By consolidating both types of replication evidence, we identify miRNAs with the strongest empirical support across complementary validation approaches. Table 1 summarizes miRNAs that demonstrate reproducibility across independent research efforts and diverse populations, while Table 2 presents miR-NAs validated through prospective discovery–replication study designs addressing asthma heterogeneity. Together, these tables summarize replicated miRNA associations across studies and clinical phenotypes. “NA” indicates that the number of participants is not available (not reported or not found).
miRNA Effects on Mechanisms of Asthma
Asthma is defined by complex interactions between structural airway cells, immune cells, and circulating mediators. miRNAs are important post-transcriptional regulators in these interactions, regulating airway inflammation, remodeling, and immune dysregulation. Recent studies are revealing compartment-specific functions of miRNAs and their role in disease severity, steroid responsiveness, and systemic immune effects.89 Table 3 summarizes asthma-associated miRNAs, their major targets, and biological roles across different biological compartments.
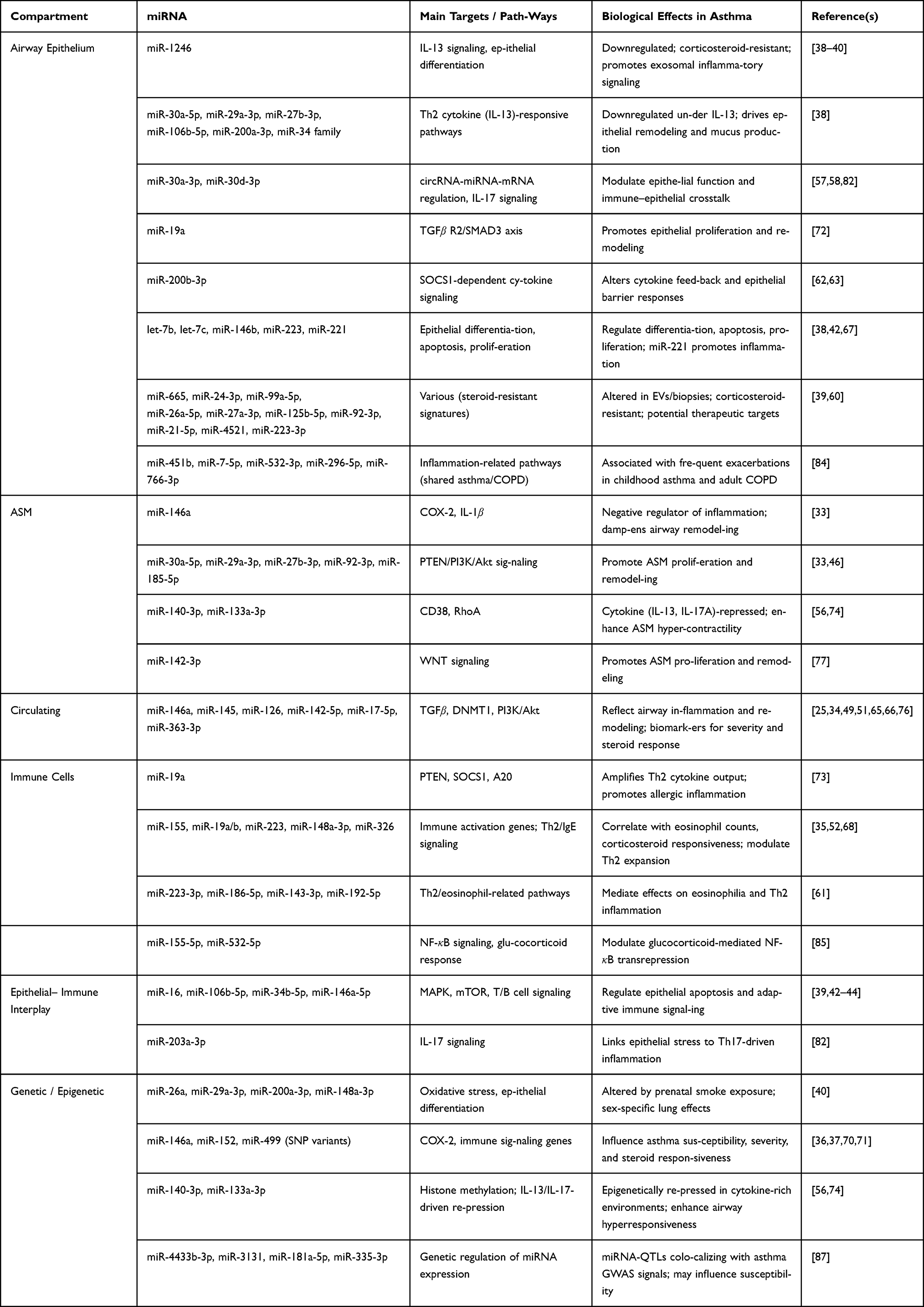 |
Table 3 Asthma-Associated miRNAs, Their Major Targets, and Biological Roles Across Compartments |
Regulation of Airway Inflammation and Remodeling
Asthma-related miRNAs regulate remodeling of the airway by targeting pathways regulated the epithelial differentiation, smooth muscle proliferation, and cytokine signaling.90
- Airway epithelium: The bronchial epithelium serves as a primary interface between environmental triggers and the immune system. Dysregulated miRNAs in epithelial cells influence epithelial integrity, cytokine signaling, and repair responses. For example, miR-1246 is consistently down-regulated in asthmatic bronchial epithelial cells, affecting IL-13–mediated pathways and epithelial differentiation. Importantly, this repression is resistant to corticosteroid therapy, linking miR-1246 to steroid-insensitive asthma. Exosomal miR-1246 can propagate pro-inflammatory signals to neighbor-ing cells, amplifying airway inflammation.38–40 Other miRNAs, including miR-30a-5p, miR-29a-3p, miR-106b-5p, miR-27b-3p, miR-200a-3p, and the miR-34 family, are downregulated under IL-13 stimulation, linking Th2 cytokine activity to epithelial remodeling.38 Additionally, miR-30a-3p and miR-30d-3p operate within circRNA–miRNA–mRNA regulatory networks, modulating airway epithelial function, IL-17 signaling, and immune–epithelial interplay in asthma.57,58,82In contrast, miR-19a promotes epithelial proliferation via suppression of TGFβ R2 and reduced SMAD3 phosphorylation, while dysregulation of miR-200b-3p alters SOCS1-dependent cytokine signaling.62,63,72Additionally, members of the let-7 family, particularly let-7b and let-7c, as well as miR-146b, miR-223, and miR-221, are highly expressed in airway biopsies and epithelial or macrophage cells, where they regulate epithelial differentiation, apoptosis, proliferation, and immune responses.38,42,67Notably, repression of let-7c, miR-665, miR-24-3p, miR-99a-5p, and miR-26a-5p in steroid-resistant asthmatic epithelium highlights corticosteroid-resistant miRNA signatures that could inform alter-native therapeutic strategies. These observations are supported by studies showing that miRNAs including miR-99a-5p, miR-26a-5p, miR-27a-3p, miR-125b-5p, miR-92-3p, miR-21-5p, miR-4521, and miR-223-3p are altered in EVs or bronchial biopsies from asthmatic patients.39,60 miR-221 also contributes to airway inflammation and has been proposed as a therapeutic target for controlling airway inflammation in both murine and human models.67
- Airway Smooth Muscle: In ASM, miRNAs regulate proliferation, contractility, and inflammatory responses, all of which contribute to airway remodeling and hyperresponsiveness. miR-146a serves as a negative regulator of inflammation by suppressing COX-2 and IL-1β, whereas dysregulation of miRNAs associated with the PTEN/PI3K/Akt axis (eg, miR-30a-5p, miR-29a-3p, miR-27b-3p, miR-92-3p, miR-185-5p) promotes ASM proliferation and remodeling.33,46 Additionally, miR-140-3p and miR-133a-3p modulate ASM contractility via CD38 and RhoA signaling; their cytokine-induced repression (IL-13, IL-17A) enhances smooth muscle hypercontractility.55,56,74 miR-142-3p contributes to ASM proliferation by modulating WNT signaling.77
- Circulating: Systemic miRNAs reflect and propagate airway inflammation. Circulating miR-146a, miR-145, miR-126, and miR-142-5p influence inflammation and remodeling by targeting pathways such as TGF-β, DNMT1, and PI3K/Akt signaling, serving as potential biomarkers for disease severity and therapeutic response.25,34,49,51,76 Additional miRNAs, including miR-451b, miR-7-5p, miR-532-3p, miR-296-5p, and miR-766-3p, have been associated with frequent exacerbations in both childhood asthma and adult COPD, suggesting shared inflammatory pathways.84 Moreover, members of the miR-17–92 cluster, including miR-17-5p and miR-363-3p, have been detected in circulating extracellular vesicles and are associated with asthma, including obesity-associated low type-2 phenotypes, linking metabolic inflammation with airway disease mechanisms.65,66 In addition, miR-320a has been associated with inhaled corticosteroid (ICS) response in analyses stratified by vitamin D status.64
Together, these findings underscore miRNAs as central regulators linking cytokine signaling, structural cell remodeling, and steroid responsiveness in asthma.
Immune Cell Regulation
miRNAs also coordinate interactions between epithelial, smooth muscle, and immune compartments, shaping adaptive immune responses and allergic inflammation.
- Epithelial: Epithelial miRNAs modulate immune signaling by regulating apoptosis, cytokine secretion, and T/B cell activation. For instance, miR-16, miR-106b-5p, miR-34b-5p, and miR-146a-5p control MAPK and mTOR pathways and influence T/B cell signaling cascades,39,42–44 miR-203a-3p regulates IL-17 signaling and Th cell differentiation, linking epithelial signals to Th17-mediated inflammation.82
- Immune cells: In immune cells, miRNAs fine-tune cytokine production and inflammatory responses. miR-19a amplifies Th2 cytokine output by targeting negative regulators such as PTEN, SOCS1, and A20, thereby exacerbating allergic inflammation.73 Circulating miRNAs, including miR-155, miR-19a/b, and miR-223, reflect systemic immune dysregulation and correlate with eosinophil counts and corticosteroid responsiveness.35,52,68 Moreover, miR-155-5p and miR-532-5p have been functionally validated to modulate glucocorticoid-mediated NF-κB transrepression, reinforcing the mechanistic link between miRNAs and steroid responsiveness.85 Hub miRNAs, including miR-223-3p, miR-186-5p, miR-143-3p, and miR-192-5p, mediate effects on eosinophilia and Th2 inflammation,61 further highlighting their role in shaping allergic immune responses. Other circulating miRNAs, such as miR-148a-3p and miR-326, modulate Th2 expansion, IgE signaling, and general immune dysregulation, contributing to allergic inflammation and asthma pathophysiology.68
Genetic and Epigenetic Influences
The expression and function of miRNAs in asthma are shaped not only by inflammation associated with disease, but also by genetic predispositions, epigenetic changes and exposures during development. These factors may predispose the airways to increased reactivity and contribute to variability in asthma severity and treatment response.91
- Prenatal and developmental exposures: Environmental exposures in critical windows of lung development can introduce permanent changes in miRNA expression. For instance, intrauterine exposure to tobacco smoke is associated with changes in the expression of miR-26a, miR-29a-3p, miR-200a-3p, and miR-148a-3p. These miRNAs regulate pathways related to epithelial differentia-tion, oxidative stress responses, and airway remodeling, and their dysregulation has sex-specific effects on lung structure and function.40 Early-life perturbation may predispose individuals to asthma by inducing abnormal airway responses prior to experiencing environmental triggers in the postnatal period.
- Genetic variation: The presence of single nucleotide polymorphisms (SNPs) can change miRNA maturation, stability, or target recognition, subsequently altering asthma risk and severity measures, as well as steroid responses. For instance, SNPs in miR-146a, miR-152, and miR-499 regulate inflammatory signaling, immune activation, and Th2/Th17 polarization.36,37,70 One SNP can effect the binding efficiency of miR-146a to its target COX-2 that can trigger airway inflammation. miR-152 targets HLA-G in airway epithelial cells, with maternal asthma status and the +3142 SNP affecting offspring gene regulation and transgenerational asthma risk, highlighting miR-152 as a potential biomarker.71 Recent studies have identified miRNA expression quantitative trait loci (miRNA-QTLs) that colocalize with asthma GWAS signals, including miR-4433b-3p, miR-3131, miR-181a-5p, and miR-335-3p75. These findings suggest that genetic variation directly influencing miRNA expression contributes to asthma susceptibility and may inform personalized therapeutic strategies.
- Epigenetic regulation: Cytokine-driven epigenetic mechanisms can dynamically regulate miRNA expression in response to the inflammatory milieu. IL-13 and IL-17A, key cytokines in Th2 and Th17-driven asthma, can repress miR-140-3p and miR-133a-3p through histone modifications and promoter methylation. These epigenetic changes create self-reinforcing feedback loops that promote airway hyperresponsiveness, smooth muscle proliferation, and extracellular matrix deposition.56,74 This demonstrates how miRNA expression is finely tuned by both the local cytokine environment and chromatin accessibility, linking inflammation to structural remodeling.
miRNA Profiles Across Asthma Phenotypes and Tissues
Asthma is a heterogeneous disease with distinct phenotypes and variable clinical manifestations. miR-NAs exhibit phenotype and tissue-specific expression patterns, linking structural cell behavior, immune responses, and systemic inflammation. Understanding these profiles provides insight into disease mech-anisms and identifies potential biomarkers or therapeutic targets.92 Table 4 summarizes phenotype and tissue-specific miRNA patterns distinguishing asthma subtypes and reflecting disease processes.
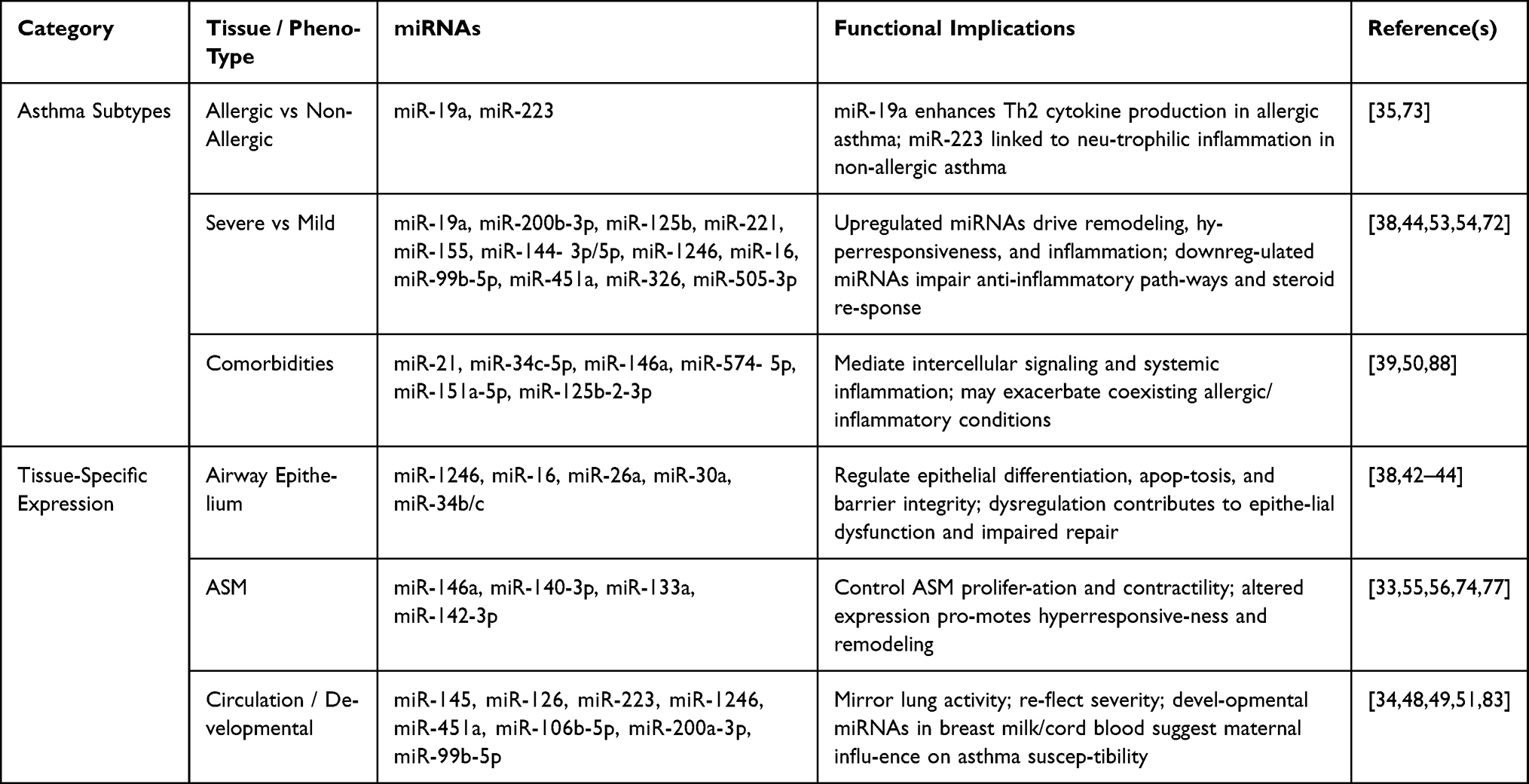 |
Table 4 miRNA Profiles Across Asthma Phenotypes and Tissue Compartments |
Asthma Subtypes and Phenotypes
Asthma-associated miRNAs show differential regulation across allergic, non-allergic, mild, and severe disease phenotypes:
- Allergic vs Non-Allergic Asthma: Allergic asthma is characterized by Th2-driven inflamma-tion. miR-19a enhances Th2 cytokine production, amplifying allergic responses and eosinophilic inflammation. In contrast, miR-223 is more relevant in non-allergic asthma, where neutrophilic inflammation predominates.35,73 These distinctions highlight the role of miRNAs in defining inflammatory endotypes and clinical heterogeneity.
- Severe vs Mild Asthma: Severe asthma exhibits pronounced airway remodeling, steroid resistance, and chronic inflammation. Upregulation of miR-19a, miR-200b-3p, miR-125b, miR-221, miR-155, and miR-144-3p/5p contributes to structural changes, hyperresponsiveness, and persistent inflammation. Conversely, downregulation of regulatory miRNAs such as miR-1246 and miR-16 impairs anti-inflammatory pathways and promotes corticosteroid insensitivity.38,53,54,72 Additional validated biomarkers include miR-99b-5p, miR-451a, miR-326, and miR-505-3p, which distinguish severe uncontrolled asthma from mild allergic asthma, highlighting their potential use in risk stratification and treatment decision-making.54 These miRNA signatures may serve as molecular markers for identifying patients with high-risk or treatment-resistant asthma.
- Comorbidities: Systemic inflammation associated with asthma comorbidities is reflected in exo-somal miRNAs such as miR-21, miR-34c-5p, and miR-146a. These miRNAs mediate intercellular signaling and may exacerbate coexisting allergic or inflammatory conditions, suggesting a role in the systemic aspects of asthma pathophysiology.39,50 Furthermore, miRNAs such as miR-574-5p, miR-151a-5p, and miR-125b-2-3p have been shown to predict incident asthma in children with recurrent wheeze, suggesting their utility as early biomarkers for disease development.88
Tissue-Specific Expression
Distinct miRNA profiles contribute to compartment-specific regulation of airway biology:
- Airway epithelium: miRNAs such as miR-1246, miR-16, miR-26a, miR-30a, and miR-34b/c regulate epithelial differentiation, apoptosis, and barrier integrity. Dysregulation in this compart-ment contributes to epithelial dysfunction, impaired repair, and susceptibility to environmental triggers.38–40,42–44
- ASM: miRNAs including miR-146a, miR-140-3p, miR-133a, and miR-142-3p control ASM prolifer-ation and contractility. Altered expression promotes airway hyperresponsiveness and remodeling, key features of severe asthma.33,55,56,74,77
- Circulation: Serum and plasma miRNAs (miR-145, miR-126, miR-223, miR-1246, miR-451a) mirror cellular activity within the lung and correlate with asthma severity. Developmental miRNAs in breast milk and cord blood (miR-106b-5p, miR-200a-3p, miR-99b-5p) suggest maternal influences on miRNA-mediated programming of asthma susceptibility in offspring.48,83
The Influence of Environmental and Lifestyle Factors
Environmental triggers and lifestyle exposures can modify miRNA expression, shaping asthma phenotypes and treatment responses as shown in Table 5.
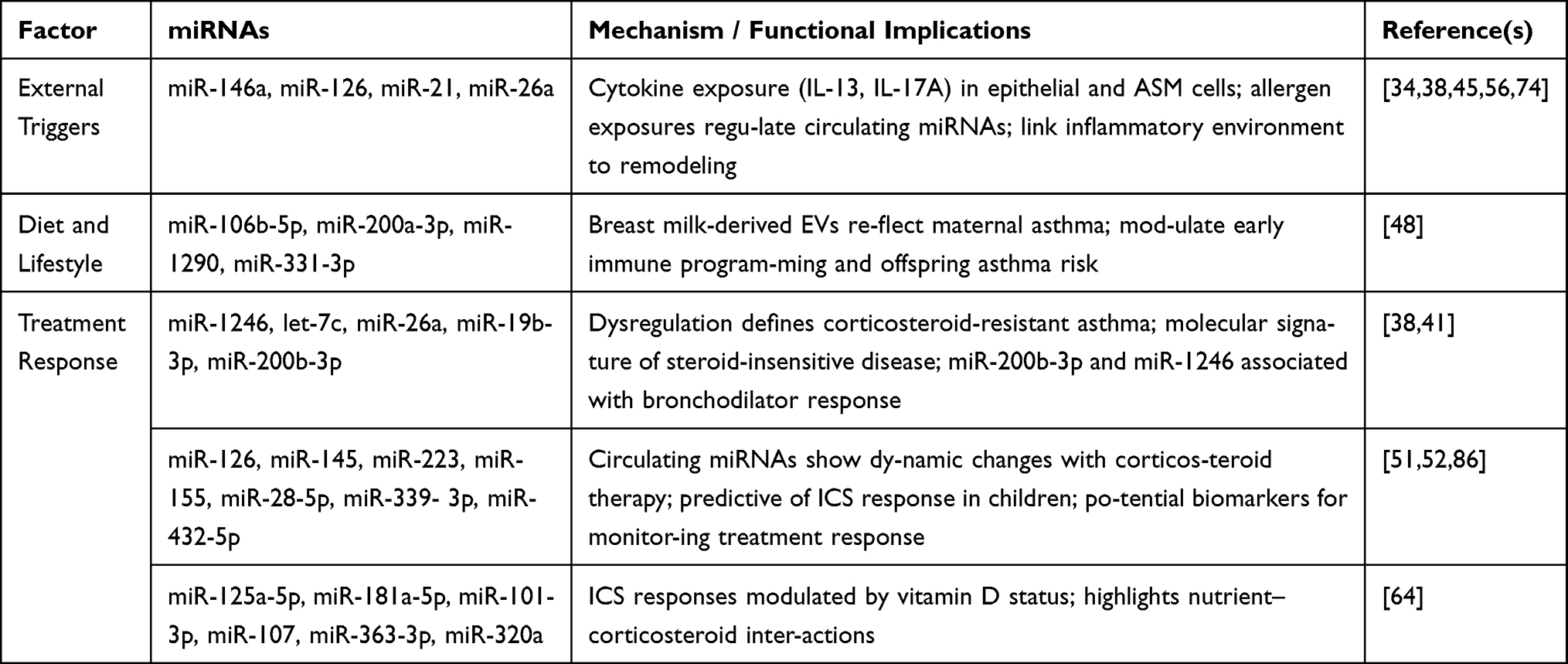 |
Table 5 Environmental and Lifestyle Influences on Asthma-Associated miRNAs |
External Triggers
Cytokine exposure (IL-13, IL-17A) recapitulates disease-associated miRNA dysregulation in epithelial and ASM cells, linking inflammatory environments directly to structural and functional remodeling.38,56,74 Allergen exposures (ovalbumin, house dust mite) regulate circulating miRNAs, including miR-146a, miR-126, miR-21, and miR-26a, highlighting their role as mediators of immune-environment interactions.34,45
Diet and Lifestyle
Maternal factors influence offspring miRNA profiles. Breast milk-derived EVs containing miRNAs (miR-106b-5p, miR-200a-3p, miR-1290, miR-331-3p) reflect maternal asthma status and may modulate early immune programming, contributing to asthma risk.48 Broader interactions between diet, metabolism, and miRNA regulation remain an emerging area of research.
Treatment Response
Corticosteroid resistance in asthma is associated with dysregulation of miRNAs such as miR-1246, let-7c, miR-26a, and miR-19b-3p, defining molecular signatures of steroid-insensitive disease.38 miR-200b-3p and miR-1246 have been replicated across pediatric and adult cohorts as associated with bronchodilator response.41 Circulating miRNAs, including miR-126, miR-145, miR-223, and miR-155, exhibit dynamic changes in response to corticosteroid therapy, supporting their potential as biomarkers for treatment monitoring.51,52 Additional predictive markers of ICS response in children include miR-28-5p, miR-339-3p, and miR-432-5p85. Furthermore, miR-125a-5p, miR-181a-5p, miR-101-3p, miR-107, and miR-320a showed associations with ICS response modified by vitamin D status, highlighting potential interactions between micronutrients and corticosteroid efficacy. In addition, miR-363-3p emerged as a replicated circulating marker in the validation analysis, supporting its potential relevance to ICS response and lung function in childhood asthma.64
Clinical Translation
miRNAs have emerged as promising tools for both the diagnosis and treatment of asthma due to their central role in regulating airway inflammation, remodeling, and immune responses. Their stability in circulation and tissue-specific expression profiles make them particularly attractive for clinical applications. Table 6 lists clinically relevant miRNAs for asthma diagnosis and treatment.
 |
Table 6 Clinical Translation of Asthma-Associated miRNAs: Biomarkers and Therapeutic Targets |
miRNAs as Biomarkers
Circulating and exosomal miRNAs provide sensitive and specific biomarkers for asthma diagnosis, phenotyping, and monitoring of disease progression:
- Exosomal miRNAs: Exosomal miRNAs such as miR-21, miR-34b/c-5p, let-7b/c, and miR-665 have demonstrated 72% predictive power in distinguishing asthma patients from healthy controls.50 Their encapsulation in exosomes protects them from degradation and reflects active intercellular signaling in asthma pathophysiology.miR-200b-3p and miR-1246 have been replicated across independent pediatric and adult cohorts, demonstrating robust associations with bronchodilator response.41 This replication underscores their potential utility as candidate biomarkers for asthma diagnosis and treatment monitoring, although prospective validation studies are still needed before clinical implementation.
- Plasma miRNAs: Circulating miRNAs offer even higher diagnostic accuracy. For instance, plasma miR-146a ratios achieved the area under the ROC curve (AUC) of 0.92, while exosomal miR-125b levels reached AUC values as high as 0.9995.34,79,80
- Disease differentiation: Certain miRNAs can distinguish asthma from other airway diseases. For example, miR-619-5p is downregulated in eosinophilic COPD compared to asthma and non-eosinophilic COPD, reflecting distinct miRNA-mediated molecular pathways and highlighting its potential utility as a biomarker for differentiating asthma from COPD subtypes.81
MiRNAs as Therapeutic Targets
Therapeutically modulating miRNAs offers the potential to correct dysregulated pathways underlying asthma pathogenesis:
- Anti-inflammatory strategies: Mimics of miR-146a can suppress ASM-mediated inflammation by inhibiting targets such as COX-2 and IL-1β, reducing cytokine-driven remodeling.33
- Modulation of airway hyperresponsiveness: Restoration of miR-140-3p and miR-133a miti-gates smooth muscle proliferation and contractility, improving airway dynamics and attenuating hyperresponsiveness.56,74
- Immune modulation: Inhibition of miR-19a, a driver of Th2 cytokine production, reduces allergic inflammation and eosinophilic responses. Similarly, targeting circulating miRNAs such as miR-142-3p and miR-126 can modulate systemic pathways including PTEN/AKT and DNMT1, offering opportunities for broader immune regulation.51,78
The dual role of miRNAs as biomarkers and therapeutic targets underscores their translational potential in asthma. By combining molecular profiling with functional interventions, miRNA-based strategies could enable early diagnosis, stratification of disease subtypes, prediction of treatment response, and development of novel precision therapeutics.
Discussion
This review applied a dual replication Criteria cross-study validation and prospective discovery-replication designs to identify miRNAs with the strongest empirical support in asthma. By prioritizing reproducibility over breadth, we aimed to distinguish robust, biologically meaningful candidates from findings potentially influenced by small cohorts, platform variability, or study-specific confounders.3,12,17,29–32 This approach narrows the field considerably, highlighting a limited subset of miRNAs with consistent validation across independent populations and phenotypes.
The externally validated miRNAs identified through our replication-based Criteria represent a focused set of high-confidence candidates supported by evidence from both cross-study validation and discov-ery–replication designs. These replicated miRNAs provide insight into key pathogenic processes in asthma, including airway inflammation, smooth muscle remodeling, and immune regulation. In addition, the reviewed studies highlight tissue- and phenotype-specific expression patterns as well as the influence of environmental and lifestyle factors on miRNA regulation. Collectively, these findings suggest potential translational applications of replicated miRNAs as biomarkers for disease detection, severity assessment, exacerbation risk prediction, and treatment response.
Among these, miR-146a and miR-1246 emerge as central candidates. miR-146a functions as a negative regulator of NF-κB-driven inflammation in airway smooth muscle and immune cells through suppression of COX-2 and IL-1β 47, 53, 65, 72, 73. miR-1246 demonstrates corticosteroid-resistant downregulation in bronchial epithelium and is linked to IL-13 signaling and exosomal propagation of inflammatory sig-nals.38–40 Their consistent replication across compartments suggests that they occupy regulatory nodes rather than representing incidental downstream markers. Nevertheless, replication alone does not establish causality; many highly replicated miRNAs may reflect consequences of established inflammation rather than primary drivers of disease.89,90 Disentangling mediators from markers will require longitudinal designs and functional intervention studies.
A key insight from the mechanistic literature is strong compartment specificity.89,90 miRNA function differs across bronchial epithelium, airway smooth muscle, immune cells, and circulating fractions. Circu-lating miRNAs are clinically attractive due to stability and accessibility,8,9 but they represent composite signals from multiple tissues. Reported diagnostic AUC values for plasma and exosomal miRNAs34,79,80 are promising yet derived from relatively small studies and require large-scale, multicenter validation. Moreover, lack of standardized extracellular vesicle isolation and quantification protocols limits cross-study comparability.23,26
Phenotypic heterogeneity further complicates interpretation. miR-19a is closely associated with Th2-driven allergic inflammation,73 whereas miR-223 is more strongly linked to neutrophilic, non-allergic pheno-types.35 No single miRNA is likely to capture the full endotypic spectrum of asthma,92 supporting the de-velopment of multi-marker, endotype-aware panels. Discovery–replication validated candidates associated with exacerbation frequency, ICS response, lung function, and incident asthma prediction64,84–88 strengthen translational relevance. Notably, exacerbation-associated miRNAs replicated in adult COPD cohorts84 suggest shared inflammatory pathways across obstructive airway diseases; however, such cross-disease replication may reflect generalized inflammation rather than asthma-specific biology.
Genetic and epigenetic data reinforce the biological plausibility of several candidates. miRNA-QTLs colocalizing with asthma GWAS loci,87 along with SNP associations involving miR-146a, miR-152, and miR-499,36,37,70,71 link germline variation to regulatory function. Environmental and developmental influences, including prenatal smoke exposure40 and cytokine-driven epigenetic repression of miR-140-3p and miR-133a-3p,55,56,74 highlight dynamic regulation within inflammatory environments. These multi-layered influences underscore that miRNA dysregulation reflects the interaction of genetic susceptibility, environmental exposure, and immune signaling.
Mechanistic studies support therapeutic hypotheses, including miR-146a augmentation to dampen airway smooth muscle inflammation,33 restoration of miR-140-3p and miR-133a-3p to reduce hypercontractil-ity,56,74 inhibition of miR-19a to attenuate Th2 amplification,73 and modulation of glucocorticoid signaling via miR-155-5p and miR-532-5p68. However, translation remains constrained by delivery challenges, tissue specificity, off-target risks, and the absence of clinical trials in asthma.
Limitations
Several limitations should be considered. First, by prioritizing replication as an inclusion criterion, this review may have excluded potentially important miRNAs identified in single but methodologically rigorous studies.1,16 While replication strengthens confidence in robustness, it may inadvertently overlook emerging candidates that warrant further investigation. Second, the included studies are heterogeneous with respect to biospecimen source (eg, blood, airway epithelium, exosomes), patient characteristics, and profiling platforms, which limits direct comparability and may influence observed replication patterns.16,26,27 Third, the predominance of cross-sectional study designs constrains causal inference, as most associations reflect correlations at a single time point; well-powered longitudinal studies remain relatively scarce.88 Fourth, generalizability may be restricted by limited ancestry representation in several cohorts, potentially reducing applicability across diverse populations.34–36,41,87 Finally, although mechanistic insights have been supported by experimental work, much of this evidence derives from in vitro systems or murine models,33,56,67,74,78 which may not fully recapitulate the complexity of human airway biology.
Future Directions
Future investigations should prioritize integrative, multi-layered approaches that combine miRNA profiling with transcriptomic, epigenomic, and proteomic datasets to better define causal regulatory networks in asthma. The application of single-cell and spatial transcriptomic technologies will be particularly valuable in resolving cell-type–specific miRNA activity across epithelial, immune, and smooth muscle compart-ments, thereby clarifying context-dependent effects that bulk analyses may obscure. Longitudinal cohort studies are also needed to determine the temporal stability of miRNA signatures and their predictive value for exacerbations, disease progression, and treatment response. Standardization of sample processing, normalization strategies, and analytic pipelines will be essential to improve reproducibility and enable meta-analytic comparisons across studies. Mechanistic validation using functional assays and in vivo models should accompany observational findings to distinguish causal regulators from secondary inflam-matory signals. Finally, translational efforts must address delivery systems, tissue specificity, and safety considerations for miRNA-based therapeutics, while prospective clinical trials are required to evaluate the feasibility of miRNA-guided precision medicine strategies in asthma management.
Conclusion
This review examined the evidence linking microRNAs (miRNAs) to asthma using replication based criteria that included both cross study validation and discovery and replication study designs. By focusing on reproducibility across independent studies, we identified a smaller group of miRNAs with consistent evidence and reduced the influence of findings that may reflect study specific variability.
Several miRNAs, including miR-146a, miR-1246, miR-19a, and miR-223, show repeated associations with processes involved in asthma such as airway inflammation, immune regulation, and airway smooth muscle remodeling. The literature also indicates that miRNA expression differs across tissues and asthma phenotypes, suggesting that individual miRNAs may contribute to specific biological contexts within the disease.
Some of the replicated miRNAs are also associated with clinical features including asthma susceptibility, exacerbation risk, lung function, and treatment response. However, additional studies are needed to confirm these findings across larger and more diverse populations.
Overall, the evidence summarized here suggests that a limited number of miRNAs show consistent associations with asthma across studies. Further work integrating multi omics data, single cell approaches, and longitudinal cohorts will help clarify their biological roles and potential clinical relevance.
Data Sharing Statement
Data sharing is not applicable to this article as no new data were created or analysed in this study.
Author Contributions
Parham Hadikhani: Conceptualization, Literature review, Data curation, Formal analysis, Visualization, Writing – original draft. Shraddha Piparia: Validation, Writing – review & editing. Rinku Sharma: Validation, Writing – review & editing. Michael McGeachie: Validation, Writing – review & editing. Kelan Tantisira: Conceptualization, Supervision, Validation, Writing – review & editing.
All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by NIH R01 HL162570, R01 HL161362, and R01 HL127332.
Disclosure
Parham Hadikhani reports grants from NIH R01 HL162570, grants from NIH R01 HL161362, grants from NIH R01 HL127332, during the conduct of the study; Kelan Tantisira reports grants from National Institutes of Health, during the conduct of the study.
References
1. Elrebehy MA, Abulsoud AI, El-Dakroury WA, et al. Tuning into mirnas: a comprehensive analysis of their impact on diagnosis, and progression in asthma. Pathol Pract. 2024;254:155147.
2. Tarlo SM, Balmes J, Balkissoon R, et al. Diagnosis and management of work-related asthma: american college of chest physicians consensus statement. Chest. 2008;134(3 Suppl):1S–22. doi:10.1378/chest.08-0201
3. Sharma R, Tiwari A, McGeachie MJ. Recent mirna research in asthma. Curr Allergy Asthma Rep. 2022;22(12):231–258. doi:10.1007/s11882-022-01050-1
4. Tantisira KG, Weiss ST. Childhood infections and asthma: at the crossroads of the hygiene and barker hypotheses. Respir res. 2001;2(6):324. doi:10.1186/rr81
5. Côté A, Godbout K, Boulet L-P. The management of severe asthma in 2020. Biochem Pharmacol. 2020;179:114112. doi:10.1016/j.bcp.2020.114112
6. Mirra D, Cione E, Spaziano G, et al. Circulating micrornas expression profile in lung inflammation: a preliminary study. J Clin Med. 2022;11(18):5446. doi:10.3390/jcm11185446
7. Sheikhpour M, Maleki M, Ebrahimi Vargoorani M, Amiri V. A review of epigenetic changes in asthma: methylation and acetylation. Clin Clin Epigenet. 2021;13(1):65. doi:10.1186/s13148-021-01049-x
8. Huang W. Micrornas: biomarkers, diagnostics, and therapeutics. In: Bioinforma in MicroRNA Research. Springer; 2017:57–67.
9. Dexheimer PJ, Cochella L. Micrornas: from mechanism to organism. Front Cell Dev Biol. 2020;8:409.
10. Vázquez-Mera S, Martelo-Vidal L, Miguéns-Suárez P, Saavedra-Nieves P, Arias P, González-Fernández C. Serum exosome inflamma-miRs are surrogate biomarkers for asthma phenotype and severity. Allergy. 2023;78(1):141–155. doi:10.1111/all.15480
11. Soccio P, Moriondo G, Lacedonia D, et al. Mirna and exosomal mirna as new biomarkers useful to phenotyping severe asthma. Biomolecules. 2023;13(10):1542. doi:10.3390/biom13101542
12. Kierbiedz´-Guzik N, Sozan´ska B. mirnas as modern biomarkers in asthma therapy. Int J Mol Sci. 2023;24(14):11499. doi:10.3390/ijms241411499
13. Gil-Martínez M, Rodrigo-Muñoz J, Sastre B, et al. Serum micrornas catalog asthma patients by phenotype. J Investig Allergol Clin Immunol. 2022;32(6):471–478. doi:10.18176/jiaci.0753
14. Guan Y, Ma Y, Tang Y, Liu X, Zhao Y, An L. Mirna-221-5p suppressed the th17/treg ratio in asthma via rorγt/foxp3 by targeting socs1. Allergy Asthma Clin Immunol. 2021;17(1):123. doi:10.1186/s13223-021-00620-8
15. Gil-Martínez M, Lorente-Sorolla C, Naharro S, Rodrigo-Muñoz JM, Del Pozo V. Advances and highlights of mirnas in asthma: biomarkers for diagnosis and treatment. Int J Mol Sci. 2023;24(2):1628. doi:10.3390/ijms24021628
16. Kyyaly MA, Vorobeva EV, Kothalawala DM, et al. Micrornas—a promising tool for asthma diagnosis and severity assessment: a systematic review. J Pers Med. 2022;12(4):543. doi:10.3390/jpm12040543
17. Al-Noshokaty TM, Fathi D, Abulsoud AI, et al. Harnessing the power of mirnas: the molecular architects of asthma pathogenesis and potential targets for therapeutic innovation. Pathol Pract. 2024;253:155054. doi:10.1016/j.prp.2023.155054
18. Baskar R, Dai J, Wenlong N, Yeo R, Yeoh K-W. Biological response of cancer cells to radiation treatment. Front Mol Biosci. 2014;1:24. doi:10.3389/fmolb.2014.00024
19. De Rie D, Abugessaisa I, Alam T, et al. An integrated expression atlas of mirnas and their promoters in human and mouse. Nat Biotechnol. 2017;35(9):872–878. doi:10.1038/nbt.3947
20. Mahmoud MM, Sanad EF, Hamdy NM. Micrornas’ role in the environment-related non-communicable diseases and link to multidrug resistance, regulation, or alteration. Environ Sci Pollut Res. 2021;28(28):36984–37000. doi:10.1007/s11356-021-14550-w
21. Erturk E, Onur OE, Akgun O, Tuna G, Yildiz Y, Ari F. Mitochondrial mirnas (mitomirs): their potential roles in breast and other cancers. Mitochondrion. 2022;66:74–81. doi:10.1016/j.mito.2022.08.002
22. Achkar NP, Cambiagno DA, Manavella PA. mirna biogenesis: a dynamic pathway. Trends Plant Sci. 2016;21(12):1034–1044. doi:10.1016/j.tplants.2016.09.003
23. O’Brien J, Hayder H, Zayed Y, Peng C. Overview of microrna biogenesis, mechanisms of actions, and circulation. Front Endocrinol. 2018;9:402. doi:10.3389/fendo.2018.00402
24. Liu X, Chen X, Yu X, et al. Regulation of micrornas by epigenetics and their interplay involved in cancer. J Exp Clin Cancer Res. 2013;32(1):96. doi:10.1186/1756-9966-32-96
25. Rodrigo-Muñoz JM, Cañas JA, Sastre B, et al. Asthma diagnosis using integrated analysis of eosinophil micrornas. Allergy. 2019;74(3):507–517. doi:10.1111/all.13570
26. Taka S, Tzani-Tzanopoulou P, Wanstall H, Papadopoulos NG. Micrornas in asthma and respiratory infections: identifying common pathways. Allergy Asthma Immunol Res. 2019;12(1):4–23. doi:10.4168/aair.2020.12.1.4
27. Wang J, Chen J, Sen S. Microrna as biomarkers and diagnostics. J Cell Physiol. 2016;231(1):25–30. doi:10.1002/jcp.25056
28. Mori MA, Ludwig RG, Garcia-Martin R, Brandão BB, Kahn CR. Extracellular mirnas: from biomarkers to mediators of physiology and disease. Cell Metab. 2019;30(4):656–673. doi:10.1016/j.cmet.2019.07.011
29. Tinè M, Padrin Y, Bonato M, et al. Extracellular vesicles (evs) as crucial mediators of cell-cell interaction in asthma. Int J Mol Sci. 2023;24(5):4645. doi:10.3390/ijms24054645
30. Zhu H, Shi J, Li W. Bioinformatics analysis of cerna network of autophagy-related genes in pediatric asthma. Medicine. 2023;102(48):e36343. doi:10.1097/MD.0000000000036343
31. Kierbiedz´-Guzik N, Sozan´ska B. The molecular basis of asthma exacerbations triggered by viral infections: the role of specific mirnas. Int J Mol Sci. 2024;26(1):120. doi:10.3390/ijms26010120
32. Zimbru R-I, Zimbru E-L, Bojin F-M, et al. Connecting the dots: how micrornas link asthma and atherosclerosis. Int J Mol Sci. 2025;26(8):3570. doi:10.3390/ijms26083570
33. Comer BS, Camoretti-Mercado B, Kogut PC, Halayko AJ, Solway J, Gerthoffer WT. Microrna-146a and microrna-146b expression and anti-inflammatory function in human airway smooth muscle. Am J Physiol Cell Mol Physiol. 2014;307(9):L727–L734. doi:10.1152/ajplung.00174.2014
34. Milger K, Götschke J, Krause L, et al. Identification of a plasma miRNA biomarker signature for allergic asthma: a translational approach. Allergy. 2017;72(12):1962–1971. doi:10.1111/all.13205
35. Weidner J, Ekerljung L, Malmhäll C, Miron N, Rådinger M. Circulating micrornas correlate to clinical parameters in individuals with allergic and non-allergic asthma. Respir Res. 2020;21(1):107. doi:10.1186/s12931-020-01351-x
36. Trinh HKT, Pham DL, Kim S-C, Kim R-Y, Park H-S, Kim S-H. Association of the mir-196a2, mir-146a, and mir-499 polymorphisms with asthma phenotypes in a Korean population. Mol Diagn Ther. 2017;21(5):547–554. doi:10.1007/s40291-017-0280-1
37. Dong J, Sun D, Lu F. Association of two polymorphisms of miRNA-146a rs2910164 (G > C) and miRNA-499 rs3746444 (T > C) with asthma: a meta-analysis. J Asthma. 2021;58(8):995–1002. doi:10.1080/02770903.2020.1759085
38. Solberg OD, Ostrin EJ, Love MI, et al. Airway epithelial mirna expression is altered in asthma. Am J Respir Crit Care Med. 2012;186:965–974.
39. Schindler VE, Alhamdan F, Preußer C, et al. Side-directed release of differential extracellular vesicle-associated microrna profiles from bronchial epithelial cells of healthy and asthmatic subjects. Biomedicines. 2022;10(3):622. doi:10.3390/biomedicines10030622
40. Rosenberg L, Liu C, Sharma R, et al. Intrauterine smoke exposure, microrna expression during human lung development, and childhood asthma. Int J Mol Sci. 2023;24(9):7727. doi:10.3390/ijms24097727
41. Sharma R, Tiwari A, Kho AT, et al. Circulating micrornas associated with bronchodilator response in childhood asthma. BMC Pulm Med. 2024;24(1):553. doi:10.1186/s12890-024-03372-4
42. Williams AE, Larner-Svensson H, Perry MM, et al. Microrna expression profiling in mild asthmatic human airways and effect of corticosteroid therapy. PLoS One. 2009;4(6):e5889. doi:10.1371/journal.pone.0005889
43. Chen Y, Qiao J. Protein–protein interaction network analysis and identifying regulation micrornas in asthmatic children. Allergol Et Immunopathologia. 2015;43(6):584–592. doi:10.1016/j.aller.2015.01.008
44. Yu B, Yao L, Liu C, Tang L, Xing T. Upregulation of microrna-16 alters the response to inhaled β -agonists in patients with asthma though modulating expression of adrb2. Mol Med Rep. 2019;19:4027–4034.
45. Jiang C, Yu H, Sun Q, et al. Extracellular microrna-21 and microrna-26a increase in body fluids from rats with antigen induced pulmonary inflammation and children with recurrent wheezing. BMC Pulm Med. 2016;16(1):50. doi:10.1186/s12890-016-0216-2
46. Alexandrova E, Miglino N, Hashim A, et al. Small rna profiling reveals deregulated phosphatase and tensin homolog (pten)/phosphoinositide 3-kinase (pi3k)/akt pathway in bronchial smooth muscle cells from asthmatic patients. J Allergy Clin Immunol. 2016;137(1):58–67. doi:10.1016/j.jaci.2015.05.031
47. Vorobeva EV, Kyyaly MA, Sones CL, et al. Circulating micrornas as potential diagnostic tools for asthma and for indicating severe asthma risk. Int J Mol Sci. 2025;26(14):6676. doi:10.3390/ijms26146676
48. Bozack AK, Colicino E, Rodosthenous RS, et al. Breast milk-derived extracellular vesicle mirnas are associated with maternal asthma and atopy. Epigenomics. 2022;14(12):727–739. doi:10.2217/epi-2022-0090
49. Coskunpinar E, Akcesme B, Tas SK, Aynaci A. Investigation of mirnas that are effective in the pathogenesis of asthma. J Asthma. 2023;60(12):2145–2152. doi:10.1080/02770903.2023.2225605
50. Levänen B, Bhakta NR, Paredes PT, et al. Altered microrna profiles in bronchoalveolar lavage fluid exosomes in asthmatic patients. J Allergy Clin Immunol. 2013;131(3):894–903. doi:10.1016/j.jaci.2012.11.039
51. Zhao M, Li Y-P, Geng X-R, et al. Expression level of mirna-126 in serum exosomes of allergic asthma patients and lung tissues of asthmatic mice. Curr Drug Metabol. 2019;20(10):799–803. doi:10.2174/1389200220666191011114452
52. Khoie ZR, Dezfuli NK, Varahram M, et al. Serum exosomal expression of mir-155 and mir-221 in moderate-to-severe asthmatic patients. Iran J Allergy Asthma Immunol. 2025;2025:1–11.
53. Gil-Martínez M, Lorente-Sorolla C, Rodrigo-Muñoz JM, et al. Analysis of differentially expressed micrornas in serum and lung tissues from individuals with severe asthma treated with oral glucocorticoids. Int J Mol Sci. 2023;24(2):1611. doi:10.3390/ijms24021611
54. Escolar-Peña A, Delgado-Dolset MI, Pablo-Torres C, et al. Specific microrna profile associated with inflammation and lipid metabolism for stratifying allergic asthma severity. Int J Mol Sci. 2024;25(17):9425. doi:10.3390/ijms25179425
55. Chiba Y, Ando Y, Kato Y, Hanazaki M, Sakai H. Down-regulation of mir-140-3p is a cause of the interlukin-13-induced up-regulation of rhoa protein in bronchial smooth muscle cells. Small GTPases. 2022;13(1):1–6. doi:10.1080/21541248.2021.1872318
56. Chiba Y, Matsumoto M, Hanazaki M, Sakai H. Downregulation of mir-140-3p contributes to upregulation of cd38 protein in bronchial smooth muscle cells. Int J Mol Sci. 2020;21(21):7982. doi:10.3390/ijms21217982
57. Chen D, Wu W, Yi L, et al. A potential circrna-mirna-mrna regulatory network in asthmatic airway epithelial cells identified by integrated analysis of microarray datasets. Front Mol Biosci. 2021;8:703307. doi:10.3389/fmolb.2021.703307
58. Wu W, Gao J, Chen D, et al. Epithelial microrna-30a-3p targets runx2/hmgb1 axis to suppress airway eosinophilic inflammation in asthma. Respir Res. 2022;23(1):17. doi:10.1186/s12931-022-01933-x
59. Fussbroich D, Kohnle C, Schwenger T, et al. A combination of lcpufas regulates the expression of mirna-146a-5p in a murine asthma model and human alveolar cells. Prostaglandins Other Lipid Mediat. 2020;147:106378. doi:10.1016/j.prostaglandins.2019.106378
60. Roffel MP, Boudewijn IM, van Nijnatten JL, et al. Identification of asthma-associated micrornas in bronchial biopsies. Eur Respir J. 2022;59(3):2101294. doi:10.1183/13993003.01294-2021
61. Sharma R, Mendez K, Begum S, et al. mirnaome-metabolome wide association study reveals effects of mirna regulation in eosinophilia and airflow obstruction in childhood asthma. EBioMedicine. 2025;112:105534. doi:10.1016/j.ebiom.2024.105534
62. Magnaye KM, Naughton KA, Huffman J, et al. A-to-i editing of mir-200b-3p in airway cells is associated with moderate-to-severe asthma. Eur Respir J. 2021;58(1):2003862. doi:10.1183/13993003.03862-2020
63. Liu F, Zhang J, Zhang D, et al. Follistatin-related protein 1 in asthma: mir-200b-3p interactions affect airway remodeling and inflammation phenotype. Int Immunopharma-Col. 2022;109:108793. doi:10.1016/j.intimp.2022.108793
64. Jiang M, Zhang Y, Liu T, et al. Circulating mirnas and childhood asthma ics response: a stratified analysis in the intervention arm of an rct with vitamin d effect modification. Thorax. 2025:8011788–8011795. doi:10.1136/thorax-2024-222618
65. Trejo-Jasso CA, Rocha-González HI, Montes-Martínez E, et al. Circulating mirnas are down-regulated in asthmatic patients; case-control study. Int J Med Sci. 2025;22(15):4003. doi:10.7150/ijms.111022
66. Alhamdan F, Greulich T, Daviaud C, et al. Identification of extracellular vesicle microRNA signatures specifically linked to inflammatory and metabolic mechanisms in obesity-associated low type-2 asthma. Allergy. 2023;78(11):2944–2958. doi:10.1111/all.15824
67. Qin H-B, Xu B, Mei J-J, et al. Inhibition of mirna-221 suppresses the airway inflammation in asthma. Inflammation. 2012;35(4):1595–1599. doi:10.1007/s10753-012-9474-1
68. Whittle E, Leonard M, Gant T, Tonge D. Multi-method molecular characterisation of human dust-mite-associated allergic asthma. Sci Rep. 2019;9:8912.
69. Aripova A, Akparova A, Bersimbaev R. The potential role of mirna-19b-3p and mirna-320c in patients with moderate bronchial asthma. MicroRNA. 2020;9(5):373–377. doi:10.2174/2211536609666201221122715
70. Zhou -P-P, Li Y, Ma Z-D, Li Z-Y, Chen F-Y, Jiang Y-X. Single nucleotide polymorphisms in the promoter region of mir-133a-1 and in pre-mir-152 rs1707 may contribute to the risk of asthma in a Chinese han population. Eur Rev Med Pharmacol Sci. 2016;20:12.
71. Nicodemus-Johnson J, Laxman B, Stern RK, et al. Maternal asthma and microrna regulation of soluble hla-g in the airway. J Allergy Clin Immunol. 2013;131(6):1496–1503. doi:10.1016/j.jaci.2013.01.037
72. Haj-Salem I, Fakhfakh R, Bérubé J-C, et al. MicroRNA-19a enhances proliferation of bronchial epithelial cells by targeting TGFβR2 gene in severe asthma. Allergy. 2015;70(2):212–219. doi:10.1111/all.12551
73. Simpson LJ, Patel S, Bhakta NR, et al. A microrna upregulated in asthma airway t cells promotes th2 cytokine production. Nat immunol. 2014;15(12):1162–1170. doi:10.1038/ni.3026
74. Chiba Y, Tanoue G, Suto R, et al. Interleukin-17a directly acts on bronchial smooth muscle cells and augments the contractility. Pharmacol Rep. 2017;69(3):377–385. doi:10.1016/j.pharep.2016.12.007
75. Chen Y, Mao Z-D, Shi Y-J, et al. Comprehensive analysis of mirna–mrna–lncrna networks in severe asthma. Epigenomics. 2019;11(2):115–131. doi:10.2217/epi-2018-0132
76. Lacedonia D, Palladino GP, Foschino-Barbaro MP, Scioscia G, Carpagnano GE. Expression profiling of miRNA-145 and miRNA-338 in serum and sputum of patients with COPD, asthma, and asthma–COPD overlap syndrome phenotype. Int J Chronic Obstruct Pulmon Dis. 2017;12:1811–1817. doi:10.2147/COPD.S130616
77. Bartel S, Carraro G, Alessandrini F, Krauss-Etschmann S, Ricciardolo FLM, Bellusci S. miR-142-3p is associated with aberrant WNT signaling during airway remodeling in asthma. Am J Physiol Cell Mol Physiol. 2018;315(2):L328–L333. doi:10.1152/ajplung.00113.2018
78. Huang H, Sun B, Li B, Wei B. mir-142-3p regulates airway inflammation through pten/akt in children and mice with asthma. Immunol Investig. 2025;54(3):297–316. doi:10.1080/08820139.2024.2438339
79. Zhao M, Juanjuan L, Weijia F, et al. Expression levels of microrna-125b in serum exosomes of patients with asthma of different severity and its diagnostic significance. Curr Drug Metabol. 2019;20(10):781–784. doi:10.2174/1389200220666191021100001
80. Tunçer F, Sahiner ÜM, Ocak M, et al. Comparison of mirna expression in patients with seasonal and perennial allergic rhinitis and non-atopic asthma. Turkish J Pediatr. 2022;64(5):859–868. doi:10.24953/turkjped.2022.410
81. Asensio VJ, Tomás A, Iglesias A, et al. Eosinophilic copd patients display a distinctive serum mirna profile from asthma and non-eosinophilic copd. Arch de Ia. 2020;56:234–241.
82. Shaik NA, Nasser K, Mohammed A, et al. Identification of mirna–mrna–tfs regulatory network and crucial pathways involved in asthma through advanced systems biology approaches. PLoS One. 2022;17(10):e0271262. doi:10.1371/journal.pone.0271262
83. Mirzakhani H, Wang AL, Sharma R, et al. Early-life microrna signatures in cord blood associated with allergic rhinitis and asthma development. J Allergy Clin Immunol. 2025;156(1):129–138. doi:10.1016/j.jaci.2024.12.1077
84. Tiwari A, Hobbs BD, Li J, et al. Blood mirnas are linked to frequent asthma exacerbations in childhood asthma and adult copd. Non-Cod RNA. 2022;8(2):27. doi:10.3390/ncrna8020027
85. Li J, Panganiban R, Kho AT, et al. Circulating micrornas and treatment response in childhood asthma. Am J Respir Crit Care Med. 2020;202:65–72.
86. Sharma R, Tiwari A, Kho AT, et al. Systems genomics reveals microrna regulation of ics response in childhood asthma. Cells. 2023;12(11):1505. doi:10.3390/cells12111505
87. Hecker J, Tiwari A, Sharma R, et al. Serum microrna expression quantitative trait loci in children with asthma colocalize with asthma-related gwas results. NPJ Genomic Med. 2025;10(1):55. doi:10.1038/s41525-025-00510-7
88. Li J, Tiwari A, Mirzakhani H, et al. Circulating microrna: incident asthma prediction and vitamin d effect modification. J Person Med. 2021;11(4):307. doi:10.3390/jpm11040307
89. Liu X, Gao J, Yang L, Yuan X. Roles of exosomal mirnas in asthma: mechanisms and applications. J Asthma Allergy. 2024;17:935–947. doi:10.2147/JAA.S485910
90. Ariel D, Upadhyay D. The role and regulation of micrornas in asthma. Curr Opinion Allergy Clin Immunol. 2012;12(1):49–52. doi:10.1097/ACI.0b013e32834ecb7f
91. Wang S-C, Tsai C-W, Chang W-S, et al. Genetic variants in mir-145 gene are associated with the risk of asthma in Taiwan. Sci Rep. 2022;12:15155.
92. Heffler E, Allegra A, Pioggia G, Picardi G, Musolino C, Gangemi S. Microrna profiling in asthma: potential biomarkers and therapeutic targets. Am J Respir Cell Mol Biol. 2017;57:642–650.
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Bibliometric Analysis of microRNA: A Comprehensive Evaluation of Its Contribution to Acute Coronary Syndromes
Gao Y, Li T, Mu X, Ren X, Wang J, Fan J, Zhu H, Wang X
Journal of Multidisciplinary Healthcare 2025, 18:3121-3137
Published Date: 3 June 2025
Early Detection of Asthma: Exploring Inflammatory Biomarkers in Symptomatic Adults with Normal Spirometry
Laroche J, Boulay MÈ, Lechasseur A, Guertin J, Boulet LP, Bergeron C, Lemière C, Lougheed MD, Vandemheen KL, Morissette MC, Aaron SD, Côté A
Journal of Asthma and Allergy 2025, 18:1675-1688
Published Date: 3 December 2025
Precision Medicine in Asthma: The Role of Biomarkers
Quek E, Horn N, Siddiqui S
ImmunoTargets and Therapy 2025, 14:1479-1513
Published Date: 28 December 2025
The Effect of Vitamin D Levels on Asthma Exacerbation in Asthmatic Patients in a Tertiary Hospital: Retrospective Study
Alhubaishi AA, Almutairi NK, Alghamdi RA, AlMolaiki MA, Alsaleh NA, Alnajjar LI, Alarfaj SJ, Aljahdali H
Journal of Asthma and Allergy 2026, 19:572118
Published Date: 27 February 2026