Back to Journals » Journal of Inflammation Research » Volume 19
Microplastics, Gut Dysbiosis, and Inflammatory Pathways in Ulcerative Colitis
Received 16 March 2026
Accepted for publication 24 June 2026
Published 10 July 2026 Volume 2026:19 609110
DOI https://doi.org/10.2147/JIR.S609110
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Alberto Caminero
Yalong Chen,1,2 Xudong Tian2
1College of Integrated Traditional Chinese and Western Medicine, Gansu University of Traditional Chinese Medicine, Lanzhou, Gansu, People’s Republic of China; 2Gastrospleen Disease Diagnosis and Treatment Center, Gansu Provincial Hospital of Traditional Chinese Medicine, Lanzhou, Gansu, People’s Republic of China
Correspondence: Xudong Tian, Gastrospleen Disease Diagnosis and Treatment Center, Gansu Provincial Hospital of Traditional Chinese Medicine, No. 418 Guazhou Road, Lanzhou, 730050, People’s Republic of China, Tel/Fax +86 18894129095, Email [email protected]
Abstract: Microplastics (MPs) are pervasive environmental pollutants characterized by their widespread distribution. They can enter the human body through multiple routes, including inhalation and dietary intake, accumulate in the gastrointestinal tract, and subsequently disrupt intestinal microecological homeostasis, thereby contributing to digestive diseases. Ulcerative colitis (UC), a chronic inflammatory bowel disease of unclear etiology, has been closely associated with gut microbiota dysbiosis, which is considered one of its central pathogenic mechanisms. This review comprehensively review the fundamental characteristics of MPs and their interactions with the gut microbiota and microbial metabolites. It further elucidates the key mechanisms by which MPs mediate the initiation and progression of UC, with particular emphasis on intestinal mucosal barrier dysfunction and immune dysregulation. Mechanistically, MPs disrupt SCFAs-producing microbial networks and activate epithelial inflammatory pathways, particularly TLR4–NF-κB signaling, thereby amplifying mucosal inflammation in UC. This review is the first to completely connect the regulatory axis of MPs – gut microbiota – metabolites – intestinal barrier – immune inflammation, clarify the core pathological chain of UC induced by MPs, make up for the shortcomings of existing reviews that only conduct single-dimensional analysis and lack integration of complete mechanisms, and provide a new theoretical framework for this field. Additionally, current research limitations are discussed, and future research directions and potential intervention strategies are proposed, aiming to provide novel theoretical insights into the etiology and prevention of UC.
Keywords: gut microbiota, immunity, inflammatory, microplastics, ulcerative colitis
Introduction
Microplastics (MPs) are generally defined as plastic fibers, particles, or films with diameters ranging from 1 μm to 5 mm, and also include nanoplastics with sizes below 1 μm. MPs are generated through the physical, chemical, and biological degradation of plastic waste and are characterized by high persistence, widespread distribution, and ecological toxicity.1 Based on their origin, MPs are classified into primary and secondary MPs. Primary MPs are intentionally manufactured at microscopic sizes, commonly used in personal care and cosmetic products, whereas secondary MPs arise from the degradation of larger plastic products through biological, physical, and chemical processes and represent the major source of environmental MPs.2 Over recent decades, plastic pollution has emerged as a major global environmental challenge. Approximately 21% of plastic waste is recycled worldwide, whereas the remaining 79% is discarded into the environment.3 MPs are now ubiquitously present in aquatic systems, soils, and the atmosphere, as well as in food and drinking water. They can enter the human body through multiple exposure pathways, including ingestion, inhalation, and dermal contact, thereby posing substantial threats to human health and socioeconomic sustainability.4 The gastrointestinal tract is considered a principal target organ for MPs accumulation5,6 (the structural characteristics of MPs and their distribution in human tissues are illustrated in Figure 1). MPs have been detected in human blood, placenta, intestinal tissues, and other organs.7 Notably, accumulation has even been reported in the skeletal system, with a deposition abundance of 61.1±44.2 particles per gram identified in intervertebral discs, with an average particle size of 159.5±103.8 μm. In addition, MPs have been detected in the human stomach.8 Analysis of gastric contents from 26 cadavers revealed widespread MPs presence, predominantly fibers (52.04%), followed by fragments (39.80%) and films (8.16%), underscoring the potential health risks associated with MPs exposure. The impacts of MPs on intestinal homeostasis, digestive and respiratory systems, and immune function have become a multidisciplinary research focus spanning environmental science, microbiology, and medicine.9 Existing studies have confirmed that MPs exposure can induce tissue injury through multiple pathways, including physical irritation, adsorption of pathogenic bacteria and toxins, release of harmful substances such as plasticizers and heavy metals, and disruption of the in vivo microecological balance.10,11 However, most current studies only focus on the environmental toxicity of MPs or a single pathogenesis of intestinal inflammation, lacking systematic integration of the complete regulatory axis of MPs exposure–gut microbiota dysbiosis–microbial metabolite imbalance–intestinal barrier damage–immune inflammation activation. Meanwhile, the core pathological chain underlying the progression of ulcerative colitis (UC) mediated by MPs remains unclear. Based on the above evidence, this review proposes a scientific hypothesis that MPs can disrupt intestinal microecological balance, damage the intestinal mucosal barrier, and trigger intestinal immune disorder, thereby driving the onset and progression of UC. This paper aims to comprehensively summarize the physicochemical characteristics, environmental exposure and human accumulation patterns of MPs, and elaborate the interactions between MPs, gut microbiota and microbial metabolites, providing novel theoretical evidence for the etiological exploration and clinical prevention and treatment of UC.
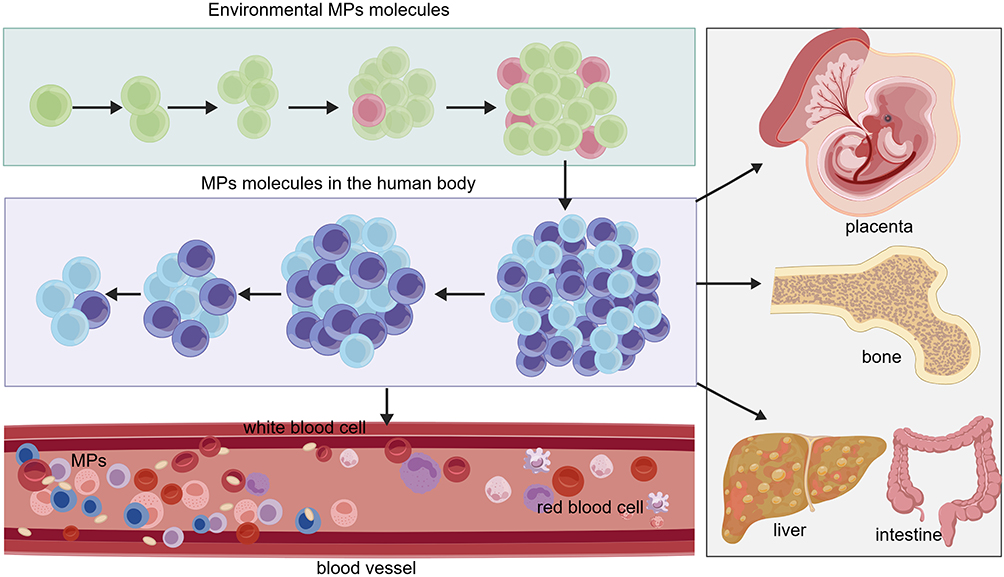 |
Figure 1 Environmental MPs enter the human body and accumulate in the bloodstream, where they interact with blood cells (including red blood cells and white blood cells). Subsequently, circulating MPs can translocate to and accumulate in multiple organs and tissues, such as the placenta, bone, liver, and intestine. |
UC is a chronic, nonspecific inflammatory bowel disease primarily involving the rectal and colonic mucosa and submucosa, with an etiology that remains incompletely understood. Clinically, it is characterized by persistent or recurrent abdominal pain, diarrhea, and mucopurulent bloody stools.12 UC is typically marked by a protracted course and a high rate of relapse. In severe cases, it may lead to adverse outcomes such as intestinal perforation and malignant transformation. Recent data indicate that among 667 patients with UC who received standard therapy over a five-year period, nearly 57.0% experienced new disease progression, 5.4% required colectomy, and 2.1% were diagnosed with malignancy,13 underscoring its status as a major global threat to gastrointestinal health. Current evidence suggests that UC pathogenesis arises from the interplay of genetic susceptibility, environmental exposures, gut microbiota dysbiosis, and immune dysregulation. Among these factors, disruption of gut microbial homeostasis has been identified as a central driving mechanism. A decline in beneficial commensals accompanied by overgrowth of pathogenic bacteria not only compromises the integrity of the intestinal mucosal barrier but also triggers aberrant immune responses, resulting in an imbalance between pro-inflammatory and anti-inflammatory mediators. This process establishes a vicious cycle of “microbiota dysbiosis-mucosal barrier disruption-immune dysregulation” that perpetuates intestinal inflammation.14 In recent years, accumulating evidence has linked environmental pollutant exposure to an increased risk of UC. A meta-analysis encompassing 32 human studies demonstrated significant associations between inflammatory bowel disease risk and exposure to heavy metals, air pollutants, per- and polyfluoroalkyl substances, and pesticides.15 As emerging environmental contaminants, MPs disturb the composition and metabolic function of gut microbiota, sharply decrease the abundance of SCFAs-producing beneficial bacteria and trigger gut dysbiosis. The subsequent microbial imbalance further impairs the integrity of intestinal mucosal barrier, activates classic inflammatory signaling pathways including TLR4-NF-κB, induces excessive secretion of pro-inflammatory cytokines and immune dysfunction, and consequently provokes persistent mucosal inflammatory damage to facilitate UC progression. However, their precise mechanistic roles have not yet been systematically elucidated. Accordingly, this review comprehensively examines the mechanisms by which MPs regulate the gut microbiota to mediate UC development and progression, with the aim of providing theoretical support for etiological research and the optimization of preventive and therapeutic strategies for UC.
Exposure Pathways and Accumulation Characteristics of MPs
The mechanistic schematic illustrating the exposure pathways and accumulation characteristics of MPs is presented in Figure 2.
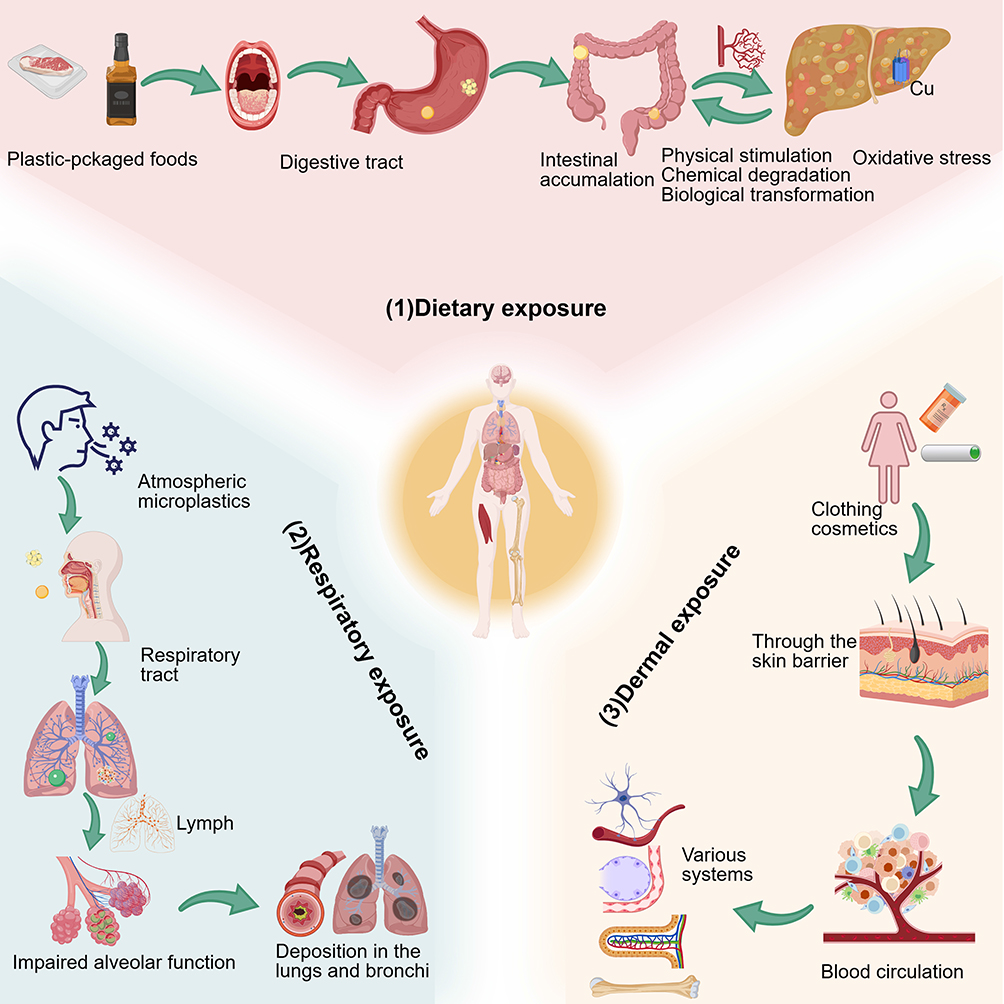 |
Figure 2 Humans are exposed to MPs via three primary routes: (1) dietary exposure through plastic-packaged foods, leading to intestinal accumulation and triggering physical stimulation, chemical degradation, biological transformation, and oxidative stress; (2) respiratory exposure via atmospheric MPs, resulting in deposition in the lungs/bronchi, impaired alveolar function, and systemic distribution through the lymphatic system; and (3) dermal exposure through clothing, cosmetics, and personal care products, enabling MPs to cross the skin barrier, enter blood circulation, and accumulate in multiple tissues. |
Human Exposure Pathways of MPs
MPs can enter the human body through multiple exposure pathways. With the advent of the “fast-food era”, dietary intake has become the predominant route of exposure. MPs have been detected in takeaway food containers, food packaging materials, drinking water (including bottled and tap water), seafood (such as fish and shellfish), processed foods (eg, chips and candies), and various other packaging products. Regular consumption or use of these items may therefore result in the ingestion of measurable quantities of MPs.16 A dietary exposure assessment conducted in Qingdao, China, reported an average intake abundance of 1.17±1.07 particles per person, with fibrous MPs accounting for 91.5% of the detected particles.17 Inhalation represents another important exposure route. Atmospheric MPs can be inhaled into the respiratory tract, and some particles may subsequently deposit in the gastrointestinal tract and accumulate in the intestine. It has been reported that the substantial increase in face mask consumption during the COVID-19 pandemic may have markedly elevated human exposure to MPs, thereby posing additional health risks.18 In addition, dermal contact-through cosmetics and textile fibers-constitutes a potential exposure pathway. MPs may penetrate the skin barrier, enter the systemic circulation, and subsequently distribute to organs such as the gastrointestinal tract and lungs. Collectively, although multiple exposure routes exist, MPs primarily exert systemic pathological effects through the gut-systemic axis. This further underscores the critical role of interactions between MPs and the intestinal microecological environment in the pathogenesis of digestive diseases.
Accumulation and Metabolic Characteristics of MPs in the Intestine
As the primary site of MPs accumulation in the human body, the intestine exhibits deposition patterns closely associated with exposure dose, particle size, and polymer type. Smaller MPs are more likely to penetrate the intestinal mucosal barrier, enter the systemic circulation, and subsequently distribute to extraintestinal organs. A study investigating the systemic toxicity of polystyrene MPs of different sizes in C57BL/6J mice demonstrated that smaller particles exhibited greater organ distribution. However, larger particles (5 μm) are associated with more pronounced intestinal mucosal barrier dysfunction and microbial dysbiosis.19 Within the intestinal environment, MPs exert toxic effects through multiple mechanisms, including physical irritation, chemical degradation, and biotransformation. Evidence indicates that MPs can enhance copper accumulation in the liver, pancreas, and intestine, thereby triggering severe oxidative stress responses.20 Additionally, biofilm-developed MPs have been shown to induce intestinal oxidative reactions and disrupt gut microbial homeostasis.21 Regardless of the specific mechanism, disturbance of gut microbiota equilibrium represents a central event in MPs-associated intestinal pathology. Consistent with findings reported by Xia et al22 continuous exposure to polyethylene MPs for 21 days resulted in significant alterations in intestinal histomorphology and pronounced microbial dysbiosis. These effects exhibited a dose-dependent trend across three MPs concentrations (1, 5, and 10 mg/L). Specifically, microbial diversity indices were markedly reduced, with the Chao index decreasing from 429 to 326 and the Shannon index declining from 7.05 to 6.02. Moreover, the relative abundance of Proteobacteria was significantly increased. The expansion of such potentially pathogenic taxa may suppress beneficial microbial populations, thereby promoting intestinal oxidative stress and impairing epithelial barrier integrity. Although existing studies have clarified the damaging mechanism of MPs on the intestine, most of them are based on animal experiments, which differ from the actual human exposure scenario, and the clinical transformation value of the research results still needs further verification.
Core Mechanisms by Which MPs Regulate the Gut Microbiota to Mediate the Initiation and Progression of Ulcerative Colitis
The core mechanisms by which MPs regulate the gut microbiota to mediate the initiation and progression of UC can be summarized into four principal aspects: remodeling of microbial community structure, disruption of microbial metabolic functions, impairment of intestinal mucosal barrier integrity, and dysregulation of intestinal immune homeostasis. A schematic overview of these mechanisms is presented in Figure 3. The following sections provide a detailed discussion of each of these four key mechanisms.
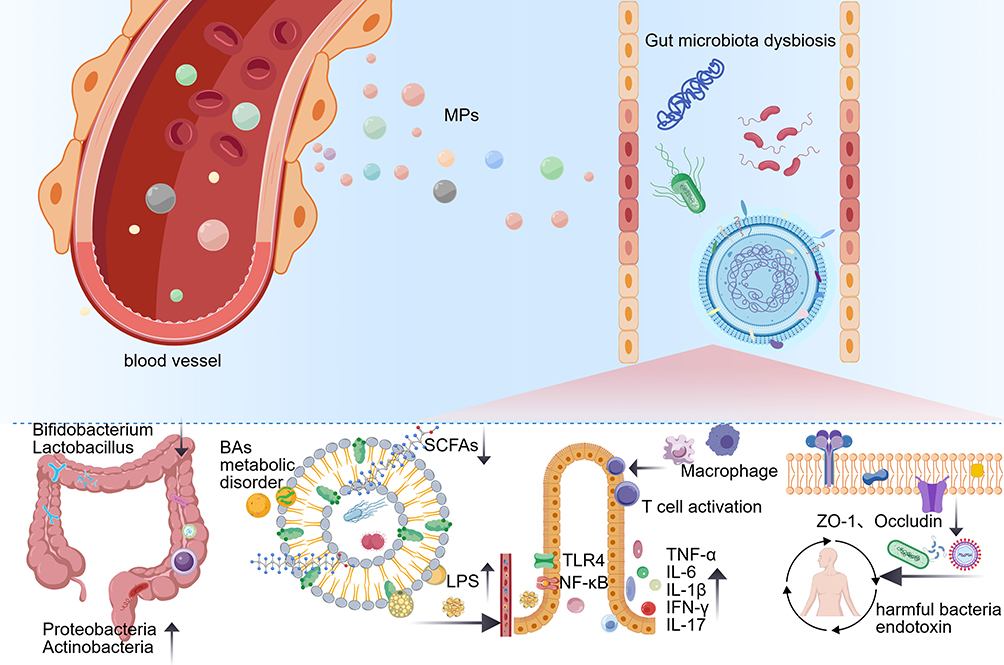 |
Figure 3 MPs translocate into the intestine and trigger gut microbiota dysbiosis, characterized by reduced beneficial bacteria (eg, Bifidobacterium, Lactobacillus) and increased pathogenic bacteria (eg, Proteobacteria, Actinobacteria). This dysbiosis impairs microbial metabolic function (including reduced short-chain fatty acids (SCFAs) and disrupted bile acids (BAs) metabolism), promotes lipopolysaccharide (LPS) release, and activates the TLR4/NF-κB pathway, leading to intestinal immune imbalance (macrophage and T cell activation, elevated pro-inflammatory cytokines). Additionally, MPs disrupt the intestinal mucosal barrier by downregulating tight junction proteins (eg, ZO-1, Occludin), increasing permeability to harmful bacteria and endotoxins, and exacerbating intestinal inflammation and dysfunction. |
MPs-Mediated Remodeling of Gut Microbial Community Structure
Homeostasis of the gut microbiota is fundamental for maintaining intestinal health, requiring a balanced abundance of beneficial and potentially harmful bacteria. Exposure to MPs can disrupt this microbial equilibrium through both physical and chemical mechanisms, thereby creating conditions conducive to the initiation and progression of UC. On the physical level, MPs can exert mechanical stimulation within the intestine, adhere to gut bacteria, and alter the local microenvironment, ultimately affecting microbial growth and proliferation. For instance, studies have shown that oral administration of polystyrene MPs in bees significantly reduced overall body weight and survival rate by impacting the gut microbiota, particularly in the rectum. Scanning electron microscopy revealed that 1 μm and 10 μm MPs tightly adhered to intestinal bacteria, and microbial analysis showed a marked decrease in beneficial bacteria such as Lactobacillus and Bifidobacterium, accompanied by an increase in pathogenic bacteria, leading to disrupted microbial community structure.23 Similarly, Jiang et al reported that the abundance of Actinobacteria increased from 0.72% in controls to 3.74% in the MPs group, while the relative abundance of Firmicutes decreased from 36.17% to 30.90%. At the genus level, MPs significantly reduced populations of Lactobacillus, Bacteroides, and Blautia, confirming the substantial impact of MPs on gut microbial composition.24 A meta-analysis of 28 studies further indicated that MPs exposure enriches taxa such as Clostridia, Proteobacteria, and Chlamydiae, while reducing Bacteroidetes, thereby promoting intestinal inflammatory infiltration.25 On the chemical level, MPs can adsorb pathogenic bacteria and toxins from the environment, introducing them into the intestine and triggering inflammation and microbial dysbiosis. MPs themselves may undergo enzymatic degradation, generating small molecules that selectively inhibit beneficial bacteria while promoting pathogenic bacterial growth. MPs have been shown to carry and release heavy metals and persistent organic pollutants (POPs), which stimulate the release of pro-inflammatory cytokines such as TNF-α and IL-6, damage the intestinal mucosa, and exacerbate oxidative stress and inflammation. When MPs accumulate beyond a certain threshold, they further disrupt the balance between beneficial and harmful bacteria, aggravating intestinal inflammation.26 Additionally, a significant proportion of foodborne micro- and nanoplastics (MNPs) has been detected in human intestinal and fecal samples. MNPs can alter gut microbial composition and metabolic function, being degraded by a series of enzymatic reactions-including biotransformation, fragmentation, and assimilation-into small molecules such as N2, CH4, and CO2, which create an environment favorable for pathogenic bacteria while inhibiting beneficial bacteria.27,28 Collectively, these studies indicate that MPs can mediate structural remodeling of the gut microbiota, reduce intestinal anti-inflammatory and antioxidant capacity, and increase susceptibility to intestinal inflammation. Notably, particle size plays a critical role: smaller MPs exert stronger effects on microbial composition, likely due to their greater ability to penetrate the intestinal mucosa and adhere to bacteria.29
MPs-Mediated Disruption of Gut Microbial Metabolic Functions
The metabolic products of the gut microbiota are central to its physiological functions. MPs can disrupt these metabolites by altering microbial community structure, with key products including short-chain fatty acids (SCFAs), lipopolysaccharides (LPS), and bile acids (BAs), all of which play critical roles in UC pathogenesis. SCFAs, produced by beneficial bacteria such as Bifidobacterium, act as a bridge between the intestinal microecology and host intestinal homeostasis. They suppress intestinal inflammation through activation of G-protein-coupled receptors, promote mucosal barrier repair, and feedback to regulate microbial homeostasis, making them essential for intestinal health.30 Exposure to MPs reduces the abundance of beneficial bacteria, directly decreasing SCFAs synthesis. This results in insufficient energy supply for intestinal epithelial cells, impaired mucosal barrier repair, weakened anti-inflammatory effects, and heightened susceptibility to intestinal inflammation. Polylactic acid MPs, commonly found in food packaging or disposable containers, have been shown to increase the abundance of H. muridarum and Helicobacter japonicus by more than 1000-fold, while populations of Prevotella sp. MGM1 and Bacteroides bouchesdurhonensis decreased significantly. Isotope-tracing studies revealed that partially degraded polylactic acid MPs fragments entered epithelial cell tricarboxylic acid (TCA) cycles, altering intestinal metabolism. SCFAs levels, the primary energy source for epithelial cells, were significantly reduced: acetate and propionate decreased by 20% and butyrate by 4%, leading to barrier disruption.31 Other studies indicate that oral MPs exposure may not always change SCFAs levels but can still alter natural killer (NK) cell populations and microbial composition, inducing severe intestinal inflammation. MPs-exposed mice exhibited more than double the number of NK cells compared to controls, with NK cells secreting IFN-γ and TNF-α, thereby exacerbating inflammation. Microbial shifts included a marked decrease in Bacteroides and an increase in inflammation-associated Allobaculum and Firmicutes.32 LPS, a cell wall component of Gram-negative bacteria, is normally present at low levels in the intestine, with the mucosal barrier preventing systemic entry.33 MPs promote the overgrowth of pathogenic bacteria, thereby increasing intestinal LPS levels. LPS can penetrate the mucosal barrier, enter circulation, and activate TLR4/NF-κB signaling pathways, triggering excessive secretion of pro-inflammatory cytokines such as TNF-α, IL-6, and IL-1β, thereby aggravating intestinal inflammation and promoting UC. For example, mice exposed to MPs for 21 days displayed acute inflammation by histopathology and blood tests, with activation of the NF-κB/NLRP3 pathway, neutrophil and macrophage infiltration, increased intestinal permeability, and elevated LPS levels, which further amplified TLR4/NF-κB/NLRP3-mediated inflammation.34 MPs also affect microbial BAs metabolism, disrupting primary and secondary BAs levels, exacerbating microbial dysbiosis and inflammation. MPs have been shown to regulate BAs synthesis and transport-related gene expression, altering the ratio of primary to secondary BAs in feces and impairing normal BAs metabolism, which leads to intestinal barrier and liver damage.35 Multi-omics analyses further indicated that MPs-mediated microbial dysbiosis disturbs BAs metabolism: 16S rRNA sequencing revealed a reduction in Bacteroides and Proteobacteria and enrichment of Actinobacteria. Conjugated BAs in the colon increased approximately 1.5–2 times relative to controls, with a positive correlation between BAs levels and Actinobacteria, and a negative correlation with Proteobacteria.36 These findings suggest that MPs may exacerbate colonic inflammation by disrupting specific microbial–BAs interactions.
MPs-Gut Microbiota Axis-Mediated Impairment of Intestinal Mucosal Barrier Function
The intestinal mucosal barrier is a critical defense system that maintains gut health, encompassing physical and chemical, including epithelial cells and tight junction proteins. Its integrity is essential, and barrier disruption is a key pathological event in UC. MPs can act on the gut microbiota to form MPs–microbiota axis, impairing mucosal barrier function through multiple mechanisms. Physical barrier: MPs-mediated microbial dysbiosis increases the production of harmful metabolites such as reactive oxygen species (ROS), which damage intestinal epithelial cells by promoting apoptosis, reducing proliferation, and downregulating tight junction proteins such as ZO-1 and Occludin. This loosens epithelial cell connections, increases intestinal permeability, and allows pathogenic bacteria and endotoxins to enter systemic circulation, triggering systemic inflammation. For example, polystyrene MPs have been shown to disrupt the physical barrier in quail ceca, alter microbial diversity, and damage ultrastructures, including microvilli breakage and rearrangement, with marked reductions in ZO-1 and Occludin expression.37 Similarly, exposure of Cipangopaludina cathayensis to polystyrene MPs and roxithromycin (ROX) resulted in mucosal damage, villus atrophy, epithelial cell shedding, and disrupted microbial structure; transcriptomic data also indicated oxidative stress and epithelial injury.38 Chemical barrier: Dysbiosis reduces mucus secretion and thickness, weakening the barrier against pathogen-epithelial contact and exacerbating mucosal injury. Polystyrene MPs exert dose-dependent intestinal and hepatic barrier injury by modulating the gut microbiota, reducing beneficial bacteria such as Firmicutes, Bifidobacterium, and Akkermansia, while increasing Proteobacteria and Bacteroidota. In treated groups (200 ppb and 500 ppb), mucus secretion decreased by 44.67%±0.52% and 49.50%±0.71%, respectively. Histology revealed significant reductions in goblet cell numbers, elevated iFABP (276.50±10.73 pg/mL), and decreased sIgA (0.60±0.03 mg/g), indicating increased intestinal permeability and chemical barrier disruption.39 Notably, bacterial biofilms on MPs can further modify their physical and chemical properties, influencing mucosal integrity. The intestinal mucosal barrier is a key link in the pathogenesis of UC. MPs can damage the intestinal mucosal barrier function from three dimensions (physical and chemical) by regulating the intestinal flora: at the physical level, they damage intestinal epithelial cells and the expression of tight junction proteins, increasing intestinal mucosal permeability; at the chemical level, they reduce intestinal mucus secretion and weaken the protective effect of the barrier.
MPs-Gut Microbiota Axis-Mediated Dysregulation of Intestinal Immune Homeostasis
The intestinal immune system is closely linked to the gut microbiota. Under normal conditions, the microbiota mediates immune tolerance through three interconnected pathways, maintaining immune homeostasis: (1) establishing a foundational microbial community through birth mode and diet, (2) dynamic microbiota–immune interactions that guide tolerance programming, and (3) direct stimulation of lymphoid tissue maturation and T cell differentiation by microbial antigens. Disruption at any of these steps can lead to immune deficiencies and compromise intestinal immunity.40 MPs can disrupt this balance by modulating the gut microbiota, breaking intestinal immune tolerance, and promoting the initiation and progression of UC. An integrated analysis of 118 experimental studies revealed that even when the microbiota possesses partial capacity to degrade MPs, MPs still exert a significant impact on intestinal microecology and indirectly influence the immune system.41 MPs promote the proliferation of pathogenic bacteria and enhance LPS release. LPS, a major component of the bacterial outer membrane, triggers cascades of interleukin-mediated responses by activating antigen-presenting cells such as macrophages and dendritic cells, leading to the secretion of pro-inflammatory cytokines. Simultaneously, Th1 and Th17 cells are activated, producing IFN-γ and IL-17, further aggravating intestinal inflammation. For example, in MPs-exposed mice, the abundance of harmful bacteria such as Desulfovibrio increased, accompanied by elevated serum levels of LPS, CRP, TNF-α, and CEA compared to controls. The expression of pro-inflammatory cytokines, including IL-1β and TNF-α, was significantly upregulated in the ileum and colon, contributing to inflammation initiation.42 Microbial dysbiosis activates the gut mucosal immune system, causing abnormal activation of lymphocytes and macrophages, immune imbalance, and exacerbation of inflammation and barrier injury, forming a vicious cycle of “microbiota dysbiosis-immune dysregulation-inflammatory stimulation” that drives UC pathology. Polystyrene MPs alter microbial composition, reducing the Firmicutes/Bacteroidetes ratio and increasing harmful bacteria such as Escherichia-Shigella and Enterobacteriaceae. Longer exposure times further increased pathogenic bacterial abundance. ELISA and flow cytometry analyses demonstrated significant downregulation of sIgA and suppressed T lymphocyte differentiation under high-concentration MPs, with reductions in CD4+ and CD8+ T cells becoming evident after 28 days. The first 14 days, during which beneficial bacteria temporarily compensate, showed minimal immune changes, indicating a strong correlation between microbial shifts and immune alterations. High-throughput sequencing and PCR-DGGE analyses indicated pronounced dysbiosis, including increased α-Proteobacteria and Actinobacteria, and significant reductions in Clostridium (2–3 times lower than controls) and Bacteroides (4 times lower than controls), both of which are crucial for immune system development and pathogen defense.43 These findings highlight the critical role of the MPs-microbiota axis in impairing the intestinal mucosal barrier and promoting UC-related inflammation. MPs also alter microbial composition and LPS metabolic pathways: Firmicutes abundance decreased, Bacteroides increased, LPS downstream genes ERK1 and NF-κB mRNA were downregulated, and immunoglobulin levels were elevated, collectively mediating intestinal immune responses.44 These findings demonstrate that MPs, via the MPs–microbiota axis, can profoundly disturb intestinal immune homeostasis, creating a pro-inflammatory environment that contributes to UC pathogenesis.
Gut Microbiota-Mediated Metabolism of MPs and Attenuation of UC Progression
Currently, environmental protection measures aim to reduce plastic and MPs usage at the source. However, in the “fast-food era”, the widespread use of plastics and MPs is inevitable due to convenience. Therefore, strategies to accelerate MPs metabolism in the body and minimize their harmful effects are of critical importance.
Recent studies indicate that certain probiotic strains in the gut can enhance MPs degradation, providing a potential intervention avenue. Probiotic supplementation has been shown to restore microbiota balance and improve intestinal barrier function, preventing unnecessary immune activation and thereby reducing MPs toxicity in vivo.45 Some probiotics can adsorb MPs in the intestine and facilitate their excretion via feces, effectively reducing intestinal accumulation. For instance, an assessment of 784 bacterial strains revealed that Lactobacillus paracasei DT66 and Lactiplantibacillus plantarum DT88 achieved the highest MPs adsorption, increasing fecal excretion of polystyrene MPs in mice by 34% and reducing residual intestinal MPs by 67%, concurrently alleviating intestinal inflammation.46 Similarly, lactic acid bacteria efficiently adsorb polypropylene, polyethylene, and polyvinyl chloride MPs, with adsorption rates of 78.57%, 71.59%, and 66.57%, respectively.47 Clostridium dalinum has also been shown to promote toxin clearance and restore intestinal barrier function by regulating metabolites, increasing ZO-1 and Occludin expression to ~97% (approximately 70% higher than MPs-exposed groups) and suppressing TLR4/NF-κB signaling to mitigate inflammation.48 Engineered strains, such as Escherichia coli Nissle 1917, can reduce MPs absorption by intestinal cells, thereby protecting mucosal barrier integrity.49 Collectively, these studies demonstrate that various probiotics can promote MPs metabolism and ameliorate UC-related intestinal pathology, though research in this area remains limited and warrants further investigation. Table 1 summarizes the mechanisms by which probiotics promote MPs metabolism or regulate gut microbiota to alleviate the progression of UC.
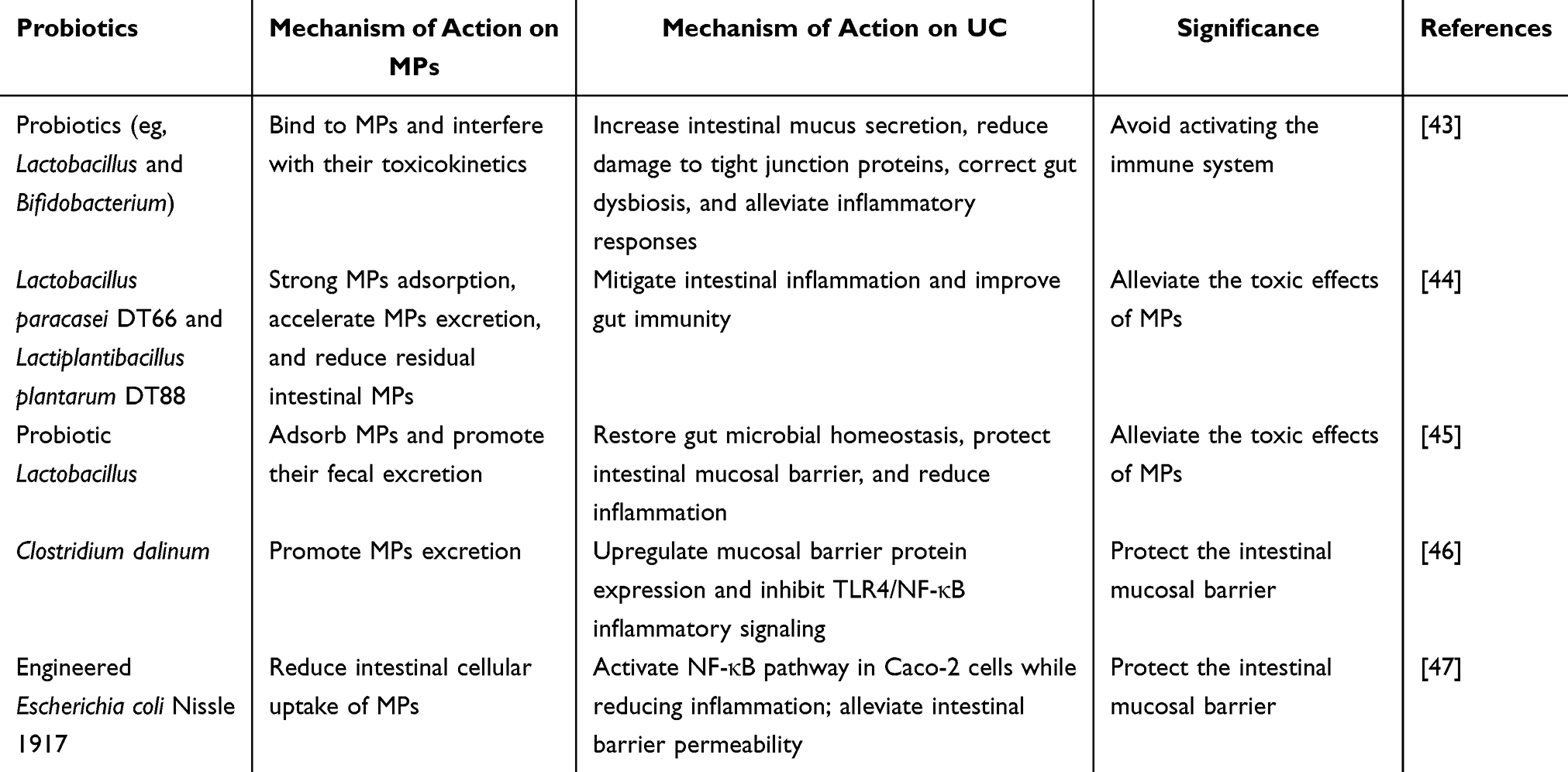 |
Table 1 The Mechanisms by Which Probiotics Promote MPs Metabolism or Regulate Gut Microbiota to Alleviate the Progression of UC |
Conclusion and Perspectives
MPs, as an emerging environmental pollutant, can enter the human body through multiple routes such as dietary intake and inhalation, and accumulate in the intestine. They contribute to gut microbiota dysbiosis and metabolic disturbances, impair intestinal mucosal barrier function, and disrupt intestinal immune homeostasis, forming a pathological pathway of “MPs exposure → gut microbiota dysbiosis → mucosal barrier damage → immune imbalance”, which plays a critical role in the pathogenesis and progression of UC. This review summarizes the core mechanisms by which MPs promote UC development, aiming to provide insights into additional etiological factors of UC. Notably, Most studies suggest that MPs are related to UC via multifaceted regulation of gut microbiota composition; however, whether MPs can serve as a modeling agent for UC in future experimental studies remains unclear, and classical UC modeling systems are still lacking. Future research could evaluate the efficacy of MPs combined with dextran sulfate sodium (DSS) in constructing UC models so as to establish an experimental system integrating MPs exposure with classical UC animal models, thereby more accurately elucidating the role of MPs in the onset and progression of UC.
Furthermore, most current studies are focused on animal experiments, including fish, shrimp, and birds, with data demonstrating correlations between MPs exposure levels, gut microbiota characteristics, and disease severity. However, clinical studies in humans are still limited, there are significant interspecies differences in intestinal structure, gut microbiota composition, and immune regulation between experimental animals and humans, limiting the direct extrapolation of animal findings to human pathogenesis. In addition, a marked discrepancy exists between the high exposure doses used in animal studies and the lower levels encountered in real-world human exposure. Therefore, current high-dose experiments mainly serve to elucidate the potential toxic mechanisms of MPs and should not be directly interpreted as reflecting UC risk in the general population. Consequently, the actual role of MPs in human UC pathogenesis and the dose–response relationship remain uncertain. Large-scale, multicenter clinical studies are needed to clarify the association between MPs exposure and UC risk or disease progression, and conduct studies using environmentally relevant low-dose and dose-gradient exposure models to establish dose–response relationship, providing evidence for the potential role of MPs in UC. Finally, current preventive strategies against MPs are largely limited to environmental protection measures. Given the unavoidable use of plastics in the contemporary convenience-oriented society, future interventions could focus on accelerating MPs metabolism in vivo by leveraging the interaction between MPs and gut microbiota. In summary, continued research on the interactions between MPs and gut microbiota is expected to open new avenues for intestinal health protection and the prevention and treatment of inflammatory bowel diseases.
Data Sharing Statement
Data sharing is not applicable to this article as no data were created or analyzed in this study.
Acknowledgments
We would like to thank Yanlong Li (Gastrospleen Disease Diagnosis and Treatment Center, Gansu Provincial Hospital of Traditional Chinese Medicine, Lanzhou, Gansu, People’s Republic of China) for his guidance on article review and editing. All figures are originally drawn by the authors, with no duplicate publication, unauthorized use or secondary reuse.
Author Contributions
Yalong Chen: Investigation, Visualization, Writing-original draft, Writing – review and editing; Xudong Tian: Conceptualization, Supervision, writing-review & editing. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (82560887), the Major Project of Gansu Provincial Joint Research Fund (24JRRA897), the Gansu Provincial Science and Technology Plan Funding - Youth Science and Technology Fund Project (23JRRA1728). The authors gratefully acknowledge the sponsorship of these funds.
Disclosure
The authors declare that there are no conflicts of interest.
References
1. Rafa N, Ahmed B, Zohora F, et al. Microplastics as carriers of toxic pollutants: source, transport, and toxicological effects. Environ Pollut. 2024;343:123190. doi:10.1016/j.envpol.2023.123190
2. Kumar M, Chaudhary V, Chaudhary V, et al. Impacts of microplastics on ecosystem services and their microbial degradation: a systematic review of the recent state of the art and future prospects. Environ Sci Pollut Res. 2024;31(55):63524–12.
3. Luo Q, Tan H, Ye M, et al. Microplastics as an emerging threat to human health: an overview of potential health impacts. J Environ Manag. 2025;387:125915. doi:10.1016/j.jenvman.2025.125915
4. Gupta A, Ghosh A, Yadav A, et al. Microplastics: the imperative influencer in blueprint of blue economy. J Environ Manag. 2024;372:123300. doi:10.1016/j.jenvman.2024.123300
5. Sun X, Zhuang Y, Wang Y, et al. Polyethylene terephthalate microplastics affect gut microbiota distribution and intestinal damage in mice. Ecotoxicol Environ Saf. 2025;294:118119. doi:10.1016/j.ecoenv.2025.118119
6. Ding R, Chen Y, Shi X, et al. Size-dependent toxicity of polystyrene microplastics on the gastrointestinal tract: oxidative stress related-DNA damage and potential carcinogenicity. Sci Total Environ. 2024;912:169514. doi:10.1016/j.scitotenv.2023.169514
7. Yang Q, Peng Y, Wu X, et al. Microplastics in human skeletal tissues: presence, distribution and health implications. Environ Int. 2025;196:109316. doi:10.1016/j.envint.2025.109316
8. Özsoy S, Gündogdu S, Sezigen S, et al. Presence of microplastics in human stomachs. Forensic Sci Int. 2024;364:112246. doi:10.1016/j.forsciint.2024.112246
9. Chartres N, Cooper CB, Bland G, et al. Effects of microplastic exposure on human digestive, reproductive, and respiratory health: a rapid systematic review. Environ Sci Technol. 2024;58(52):22843–22864. doi:10.1021/acs.est.3c09524
10. Lin W, Zhou X, Zeng Y, et al. Microplastics as vectors for microcystin-LR: enhanced bioavailability, oxidative damage, and immune dysregulation in zebrafish spleen. Fish Shellfish Immunol. 2025;167:110883. doi:10.1016/j.fsi.2025.110883
11. Fontes BLM, de Souza E Souza LC, Da Silva De Oliveira APS, et al. The possible impacts of nano and microplastics on human health: lessons from experimental models across multiple organs. J Toxicol Environ Health B. 2024;27(4):153–187. doi:10.1080/10937404.2024.2330962
12. Pravda J. Ulcerative colitis: timeline to a cure. World J Gastroenterol. 2025;31(26):108375. doi:10.3748/wjg.v31.i26.108375
13. Wan J, Shen J, Zhong J, et al. Natural course of ulcerative colitis in China: differences from the West? United Eur Gastroenterol J. 2024;12(9):1167–1178. doi:10.1002/ueg2.12634
14. Bu F, Chen K, Chen S, et al. Gut microbiota and intestinal immunity interaction in ulcerative colitis and its application in treatment. Front Cell Infect Microbiol. 2025;15:1565082. doi:10.3389/fcimb.2025.1565082
15. Estevinho MM, Midya V, Cohen-Mekelburg S, et al. Emerging role of environmental pollutants in inflammatory bowel disease risk, outcomes and underlying mechanisms. Gut. 2025;74(3):477–486. doi:10.1136/gutjnl-2024-332523
16. Li Y, Chen L, Zhou N, et al. Microplastics in the human body: a comprehensive review of exposure, distribution, migration mechanisms, and toxicity. Sci Total Environ. 2024;946:174215. doi:10.1016/j.scitotenv.2024.174215
17. Huang L, Li J, Yang D, et al. The occurrence and exposure of microplastics in bivalves from Qingdao, China. Mar Pollut Bull. 2024;207:116880. doi:10.1016/j.marpolbul.2024.116880
18. Zhao B, Rehati P, Yang Z, et al. The potential toxicity of microplastics on human health. Sci Total Environ. 2024;912:168946. doi:10.1016/j.scitotenv.2023.168946
19. Zhang Z, Chen W, Chan H, et al. Polystyrene microplastics induce size-dependent multi-organ damage in mice: insights into gut microbiota and fecal metabolites. J Hazard Mater. 2024;461:132503. doi:10.1016/j.jhazmat.2023.132503
20. Zhang C, Ye L, Wang C, et al. Toxic effect of combined exposure of microplastics and copper on goldfish (Carassius auratus): insight from oxidative stress, inflammation, apoptosis and autophagy in hepatopancreas and intestine. Bull Environ Contam Toxicol. 2022;109(6):1029–1036. doi:10.1007/s00128-022-03585-5
21. Yu Z, Qiu D, Zhou T, et al. Biofilm enhances the interactive effects of microplastics and oxytetracycline on zebrafish intestine. Aquat Toxicol. 2024;270:106905. doi:10.1016/j.aquatox.2024.106905
22. Xia X, Ma X, Liang N, et al. Damage of polyethylene microplastics on the intestine multilayer barrier, blood cell immune function and the repair effect of Leuconostoc mesenteroides DH in the large-scale loach (Paramisgurnus dabryanus). Fish Shellfish Immunol. 2024;147:109460. doi:10.1016/j.fsi.2024.109460
23. Wang K, Zhu L, Rao L, et al. Nano- and micro-polystyrene plastics disturb gut microbiota and intestinal immune system in honeybee. Sci Total Environ. 2022;842:156819. doi:10.1016/j.scitotenv.2022.156819
24. Jiang P, Yuan G, Jiang B, et al. Effects of microplastics (MPs) and tributyltin (TBT) alone and in combination on bile acids and gut microbiota crosstalk in mice. Ecotoxicol Environ Saf. 2021;220:112345. doi:10.1016/j.ecoenv.2021.112345
25. Souza-Silva TGD, Oliveira IA, Silva GGD, et al. Impact of microplastics on the intestinal microbiota: a systematic review of preclinical evidence. Life Sci. 2022;294:120366. doi:10.1016/j.lfs.2022.120366
26. Bora SS, Gogoi R, Sharma MR, et al. Microplastics and human health: unveiling the gut microbiome disruption and chronic disease risks. Front Cell Infect Microbiol. 2024;14:1492759. doi:10.3389/fcimb.2024.1492759
27. Wang J, Wang Y, Li Z, et al. Gut microbiota, a key to understanding the knowledge gaps on micro-nanoplastics-related biological effects and biodegradation. Sci Total Environ. 2024;944:173799. doi:10.1016/j.scitotenv.2024.173799
28. Zhao Z, Liu W, Wu L, et al. Effects of waxy maize starch and malate-debranched waxy maize starch on gut microbiota of humans in vitro and mice in vivo. Microorganisms. 2025;13(9):2218. doi:10.3390/microorganisms13092218
29. Demarquoy J. Microplastics and microbiota: unraveling the hidden environmental challenge. World J Gastroenterol. 2024;30(16):2191–2194. doi:10.3748/wjg.v30.i16.2191
30. Hays KE, Pfaffinger JM, Ryznar R. The interplay between gut microbiota, short-chain fatty acids, and implications for host health and disease. Gut Microbes. 2024;16(1):2393270. doi:10.1080/19490976.2024.2393270
31. Bao L, Cui X, Zeng T, et al. Incorporation of polylactic acid microplastics into the carbon cycle as a carbon source to remodel the endogenous metabolism of the gut. Proc Natl Acad Sci USA. 2025;122(19):e1877863174. doi:10.1073/pnas.2417104122
32. Hasegawa Y, Okamura T, Ono Y, et al. Oral exposure to high concentrations of polystyrene microplastics alters the intestinal environment and metabolic outcomes in mice. Front Immunol. 2024;15:1407936. doi:10.3389/fimmu.2024.1407936
33. Mishra S, Jain S, Agadzi B, et al. A cascade of microbiota-leaky gut-inflammation- is it a key player in metabolic disorders? Curr Obes Rep. 2025;14(1):32. doi:10.1007/s13679-025-00624-0
34. Chen X, Xuan Y, Chen Y, et al. Polystyrene nanoplastics induce intestinal and hepatic inflammation through activation of NF-κB/NLRP3 pathways and related gut-liver axis in mice. Sci Total Environ. 2024;935:173458. doi:10.1016/j.scitotenv.2024.173458
35. Wen J, Sun H, Yang B, et al. Environmentally relevant concentrations of microplastic exposure cause cholestasis and bile acid metabolism dysregulation through a gut-liver loop in mice. Environ Sci Technol. 2024;58(4):1832–1841. doi:10.1021/acs.est.3c07108
36. Chen J, Cheng Y, Fu R, et al. Multiomics reveals nonphagocytosable microplastics induce colon inflammatory injury via bile acid-gut microbiota interactions and barrier dysfunction. ACS Appl Mater Interfaces. 2025;17(31):44138–44159. doi:10.1021/acsami.5c07250
37. Jing L, Zhang Y, Zhang Q, et al. Polystyrene microplastics disrupted physical barriers, microbiota composition and immune responses in the cecum of developmental Japanese quails. J Environ Sci. 2024;144:225–235. doi:10.1016/j.jes.2023.08.020
38. Wang M, Liu X, Wang J, et al. Three-dimensional synergistic mechanism ofphysical injury, microbiota dysbiosis, and gene transfer in the gut of Cipangopaludina cathayensisunder microplastics and roxithromycin exposure. J Environ Manage. 2025;394:127514. doi:10.1016/j.jenvman.2025.127514
39. Li H, Xu S, Zhou F, et al. Polystyrene microplastics exposure: disruption of intestinal barrier integrity and hepatic function in infant mice. Ecotoxicol Environ Saf. 2024;288:117357. doi:10.1016/j.ecoenv.2024.117357
40. Wang Q, Meng Q, Chen Y, et al. Interaction between gut microbiota and immunity in health and intestinal disease. Front Immunol. 2025;16:1673852. doi:10.3389/fimmu.2025.1673852
41. Bao S, Yi J, Xian B, et al. Global analysis of the adverse effects of micro- and nanoplastics on intestinal health and microbiota of fish. J Hazard Mater. 2024;470:134157. doi:10.1016/j.jhazmat.2024.134157
42. Gan H, Chen S, Yao K, et al. Simulated microplastic release from cutting boards and evaluation of intestinal inflammation and gut microbiota in mice. Environ Health Perspect. 2025;133(3–4):47004. doi:10.1289/EHP15472
43. Zhang Z, Xu M, Wang L, et al. Continuous oral exposure to micro- and nanoplastics induced gut microbiota dysbiosis, intestinal barrier and immune dysfunction in adult mice. Environ Int. 2023;182:108353. doi:10.1016/j.envint.2023.108353
44. Sun H, Chen N, Yang X, et al. Effects induced by polyethylene microplastics oral exposure on colon mucin release, inflammation, gut microflora composition and metabolism in mice. Ecotoxicol Environ Saf. 2021;220:112340. doi:10.1016/j.ecoenv.2021.112340
45. Bazeli J, Banikazemi Z, Hamblin MR, et al. Could probiotics protect against human toxicity caused by polystyrene nanoplastics and microplastics? Front Nutr. 2023;10:1186724. doi:10.3389/fnut.2023.1186724
46. Teng X, Zhang T, Rao C. Novel probiotics adsorbing and excreting microplastics in vivo show potential gut health benefits. Front Microbiol. 2025;15:1522794. doi:10.3389/fmicb.2024.1522794
47. Zhao L, Dou Q, Chen S, et al. Adsorption abilities and mechanisms of Lactobacillus on various nanoplastics. Chemosphere. 2023;320:138038. doi:10.1016/j.chemosphere.2023.138038
48. Zheng-Qiang L, Jun L, Rui A, et al. A probiotic for preventing microplastic toxicity: clostridium dalinum mitigates microplastic-induced damage via microbiota-metabolism-barrier interactions. Curr Res Food Sci. 2025;11:101200. doi:10.1016/j.crfs.2025.101200
49. Chen W, Guo Q, Li H, et al. Engineered probiotics mitigate gut barrier dysfunction induced by nanoplastics. Adv Sci. 2025;12(22):e2417283. doi:10.1002/advs.202417283
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Xianglian Zhixie Tablet Antagonizes Dextran Sulfate Sodium-Induced Ulcerative Colitis by Attenuating Systemic Inflammation and Modulating Gut Microbiota

Li Y, Wang T, Ma B, Yu S, Pei H, Tian S, Tian Y, Liu C, Zhao X, Zuo Z, Wang Z
Journal of Inflammation Research 2023, 16:4331-4346
Published Date: 28 September 2023
Microbial Disruptions in Inflammatory Bowel Disease: A Comparative Analysis
Ma J, Wang K, Wang J, Zeng Q, Liu K, Zheng S, Chen Y, Yao J
International Journal of General Medicine 2024, 17:1355-1367
Published Date: 6 April 2024
Portulaca Oleracea L. as a Potential Therapeutic Drug Intervention in Ulcerative Colitis: Mechanisms of Action and Clinical Studies
Liu XY, Liu ZX, Tan WW, Zhang WB, Zhang YL, Zheng L, Que RY, Wen HZ, Dai YC
Drug Design, Development and Therapy 2024, 18:5931-5946
Published Date: 11 December 2024
Coix Seed Oil Alleviates DSS-Induced Ulcerative Colitis via Intestinal Barrier Repair and Ferroptosis Regulation
Zeng YX, Li NR, Deng BY, Gu YF, Lu SF, Liu Y
Journal of Inflammation Research 2025, 18:2557-2581
Published Date: 20 February 2025
Si-Ni Decoction as a Potential Treatment for Ulcerative Colitis: Modulation of Gut Microbiota and AKT1 Inhibition Through Network Pharmacology and in vivo Validation
Shi L, Chen L, Jin G, Yang Y, Zhu F, Zhou G
Journal of Inflammation Research 2025, 18:6263-6280
Published Date: 14 May 2025