Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 19
Microarray Data Analysis of Gene Crosstalk Between Psoriasis and Non-Alcoholic Fatty Liver Disease
Authors Tan Y, Xu C ![]() , Xiao Y
, Xiao Y ![]() , Song W, Zhang Y, Yang Z
, Song W, Zhang Y, Yang Z ![]() , Chen A, Wu Q
, Chen A, Wu Q
Received 8 December 2025
Accepted for publication 11 March 2026
Published 21 March 2026 Volume 2026:19 587398
DOI https://doi.org/10.2147/CCID.S587398
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Monica K. Li
Ye Tan,1 Chen Xu,1 Yi Xiao,1 Wenjing Song,2 Yibo Zhang,1 Zhihan Yang,3 Aidong Chen,4 Qiwen Wu2
1Kangda College of Nanjing Medical University, Lianyungang, 222000, People’s Republic of China; 2Department of Laboratory Medicine, The First Affiliated Hospital of Wannan Medical College, Wuhu, 241001, People’s Republic of China; 3Medical School of Nantong University, Nantong, 226000, People’s Republic of China; 4The Key Laboratory of Targeted Intervention of Cardiovascular Disease, Nanjing Medical University, Nanjing, Jiangsu, 211166, People’s Republic of China
Correspondence: Qiwen Wu, Department of Laboratory Medicine, The First Affiliated Hospital of Wannan Medical College, 2 Zheshan West Road, Wuhu, Anhui, People’s Republic of China, Email [email protected] Aidong Chen, The Key Laboratory of Targeted Intervention of Cardiovascular Disease, Nanjing Medical University, Nanjing, Jiangsu, 211166, People’s Republic of China, Email [email protected]
Background: Numerous studies both domestically and internationally have indicated a potential close association between psoriasis and non-alcoholic fatty liver disease (NAFLD), yet the precise mechanism underlying this association remains incompletely elucidated.
Methods: Microarray datasets were retrieved from the GEO database, and differentially expressed genes (DEGs) were identified using the GEO2R tool. Functional system analysis was conducted utilizing clusterProfiler, STRING, Cytoscape, and cytoHubba.
Results: This study identified a total of 49 commonly differentially expressed genes, including 10 upregulated genes and 39 downregulated genes. Protein-protein interaction(PPI) network analysis identified 15 hub genes, and GeneMANIA analysis showed that they were mainly enriched in co-expression (75.18%).
Conclusion: The study reveals the comorbid molecular mechanism of psoriasis and NAFLD from the perspectives of gene expression, functional pathways, and transcriptional regulation, providing a new direction for the development of cross-disease therapeutic targets.
Keywords: non-alcoholic fatty liver disease, psoriasis, DEGs, bioinformatics
Introduction
Nonalcoholic fatty liver disease (NAFLD) and psoriasis are two common chronic diseases, characterized by liver lipid metabolism disorder and skin immune inflammation disorder, respectively.1,2 The incidence of both diseases is showing a rising trend globally. NAFLD encompasses a pathological process ranging from simple hepatic steatosis to non-alcoholic steatohepatitis (NASH),3 liver fibrosis, and even cirrhosis, and is closely associated with obesity, insulin resistance, and metabolic syndrome.4 Psoriasis is an immune-mediated skin disease mediated by Th1/Th17 cells,5 characterized by epidermal hyperplasia, abnormal keratinization, and inflammatory cell infiltration in the dermis. Its pathogenesis is closely related to genetic susceptibility, innate immune activation, and cytokine network dysregulation.6 Clinical observations and epidemiological studies have shown that the prevalence of NAFLD in patients with psoriasis is significantly higher than that in the general population,7 and the severity of NAFLD is positively correlated with the activity of psoriatic lesions, suggesting that the two may share potential molecular pathological mechanisms, such as chronic inflammation, oxidative stress, lipid metabolism abnormalities, and signaling pathway dysregulation.8
Despite significant phenotypic differences between NAFLD and psoriasis, their pathological processes involve abnormal activation of inflammatory pathways such as nuclear factor κB (NF-κB), mitogen-activated protein kinase (MAPK), and Janus kinase-signal transducer and activator of transcription (JAK-STAT),9,10 as well as dysfunction of metabolism-related transcription factors such as peroxisome proliferator-activated receptor (PPAR) and sterol regulatory element-binding protein (SREBP).11,12 For instance, pro-inflammatory cytokines such as tumor necrosis factor-α (TNF-α) and interleukin-17 (IL-17) drive excessive proliferation of keratinocytes in psoriatic lesions, while inducing lipid accumulation in hepatocytes and intrahepatic inflammation;13,14 whereas abnormal secretion of hepatogenic inflammatory factors (such as IL-6 and C-reactive protein) in NAFLD may exacerbate skin lesions through systemic inflammation.15 However, despite the discovery of various molecular targets and related pathways in diseases such as psoriasis and NAFLD, systematic research on common molecular targets for these diseases remains inadequate, especially the cross-disease differential gene screening and core regulatory network analysis based on gene expression profiles have not been fully explored.
Based on this, this study aims to utilize bioinformatics methods to integrate gene expression microarray data from non-alcoholic fatty liver disease (NAFLD) and psoriasis, screen for common differentially expressed genes (DEGs), construct a protein-protein interaction (PPI) network, identify key hub genes, and reveal the core biological processes and signaling pathways involved in these genes through in-depth functional enrichment analysis. The study further predicts the transcription factors (TFs) that regulate the hub genes and validates the expression patterns of core genes using independent datasets, with the aim of providing new molecular targets for elucidating the comorbid mechanism of NAFLD and psoriasis and laying the foundation for the development of cross-disease treatment strategies.
Materials and Methods
GEO Data Download and Compilation
The microarray datasets were sourced from the Gene Expression Omnibus (GEO) database (http://www.ncbi.nlm.nih.gov/geo/), a repository rich in high-throughput sequencing and expression microarray data. Using keywords such as “psoriasis” and “Nonalcoholic fatty liver disease”, relevant gene expression datasets were searched, with non-human specimens excluded from consideration. Ultimately, datasets bearing the accession numbers GSE30999, GSE14905, GSE89632, and GSE164760 were downloaded from the GEO database. Specifically, GSE30999 and GSE14905 pertain to psoriasis, whereas GSE89632 and GSE164760 relate to non-alcoholic fatty liver disease. (Table 1).
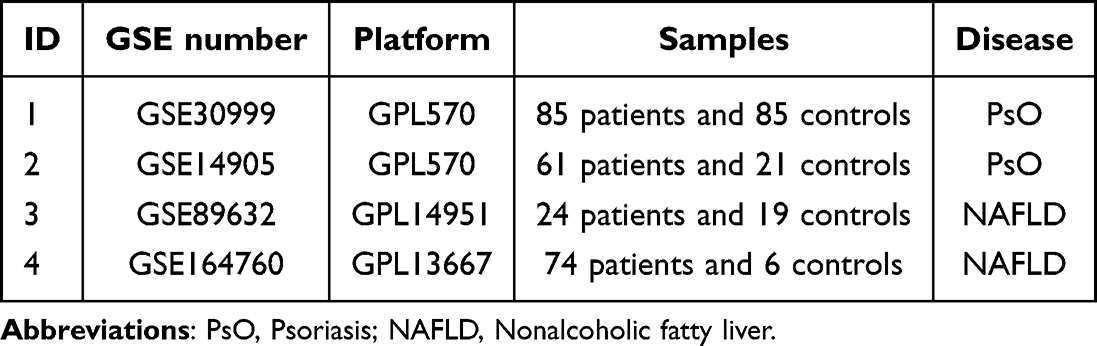 |
Table 1 Characteristics of the GEO Datasets Included in This Study |
Recognition of DEGs
The online analysis tool GEO2R was utilized to extract differentially expressed genes (DEGs) for the analysis of psoriasis and non-alcoholic fatty liver disease (NAFLD) from the dataset available at (https://www.ncbi.nlm.nih.gov/geo/geo2r/).16 The screening criteria for DEGs were set as follows: an adjusted p-value of less than 0.05 and a log2 fold change (FC) greater than 1. Gene differential expression was calculated using the “limma” package in R (version 4.4.3). We adopted the “ggplot2” package to create gene heatmaps and volcano plots, with log2FC serving as the horizontal axis and -log10(P.adj) as the vertical axis. Furthermore, an online Venn diagram tool (http://bioinformatics.psb.ugent.be/webtools/Venn/) was employed to visualize the overlapping DEGs between psoriasis and NAFLD (Supplementary Tables 1 and 2).
Functional Enrichment Analysis of DEGs
The enrichment analysis of gene sets offers valuable insights into the unique biological attributes of genes, encompassing biological processes, cellular components, and molecular functions. The Kyoto Encyclopedia of Genes and Genomes(KEGG)17 pathway enrichment analysis delves into the crucial pathways implicated in the onset and progression of diseases. In the realm of bioinformatics analysis, the R package clusterProfiler was employed to conduct KEGG and Gene Ontology(GO) enrichment analyses on differentially expressed genes associated with the progression of non-alcoholic fatty liver disease (NAFLD). Notably, the screened differentially expressed genes exhibited a significant positive correlation with fibrosis grading and activity scores, with a statistically significant p-value threshold of less than 0.05.
PPI Network Analysis and Hub Gene Identification
PPI network analysis was performed utilizing the online database STRING (http://string-db.org).18,19 Interactions with a composite score exceeding 0.4 were deemed statistically significant. The Cytoscape software (http://www.cytoscape.org)20 was employed to visualize the PPI network. Cluster analysis was conducted using Cytoscape’s Molecular Complex Detection (MCODE) plugin, with default parameters set as follows: K-core = 2, degree cutoff = 2, max depth = 100, and node score cutoff = 0.2. The CytoHubba plugin within Cytoscape was leveraged to identify hub genes, utilizing seven commonly adopted algorithms (MCC, MNC, Degree, EPC, Closeness, Radiality, Stress) to evaluate and select central genes. Ultimately, GeneMANIA (http://www.genemania.org/)21 was utilized to construct a co-expression network for these hub genes.
Validate the Expression of Hub Genes
Using Other Datasets The GSE16476022 and GSE3099923 datasets were employed to validate the mRNA expression of the identified hub genes. Specifically, the GSE164760 dataset comprised 6 normal liver tissue samples and 74 NASH liver tissue samples. Meanwhile, the GSE30999 dataset consisted of 85 non-lesional skin samples and 85 psoriasis tissue samples. A t-test was conducted to compare the datasets, with statistical significance indicated by a p-value of less than 0.05.
Prediction and Verification of TF
The Transcriptional Regulatory Relationships Unraveled by Sentence-based Text mining (TRRUST) is a meticulously curated database of transcriptional regulatory networks, which includes target genes corresponding to transcription factors and the regulatory interactions between them.24 The TRRUST database, being a manually annotated repository of transcriptional regulatory networks, comprises 8972 established regulatory interactions, accounting for 59.8% of the overall regulatory landscape. Utilizing the TRRUST database, we identified transcription factors that regulate hub genes and subsequently validated the expression levels of these transcription factors for hub genes in GSE30999 and GSE164760 using a t-test, with statistical significance determined by an adjusted p-value of less than 0.05.
Results
Differential Gene Identification
The research flowchart is depicted in Figure 1. Following standard processing of the datasets, the “limma” package within R software was utilized to visualize volcano plots (Figure 2A and B) and heatmaps (Figure 2C and D) for the differentially expressed genes across the two datasets. A total of 580 differentially expressed genes were identified in GSE14905, whereas 411 were identified in GSE89632. Through intersection analysis using a Venn diagram, 49 common differentially expressed genes were obtained. After excluding genes with discordant expression trends between GSE30999 and GSE89632, 39 down-regulated and 10 up-regulated differentially expressed genes were pinpointed (Figure 2E and F).
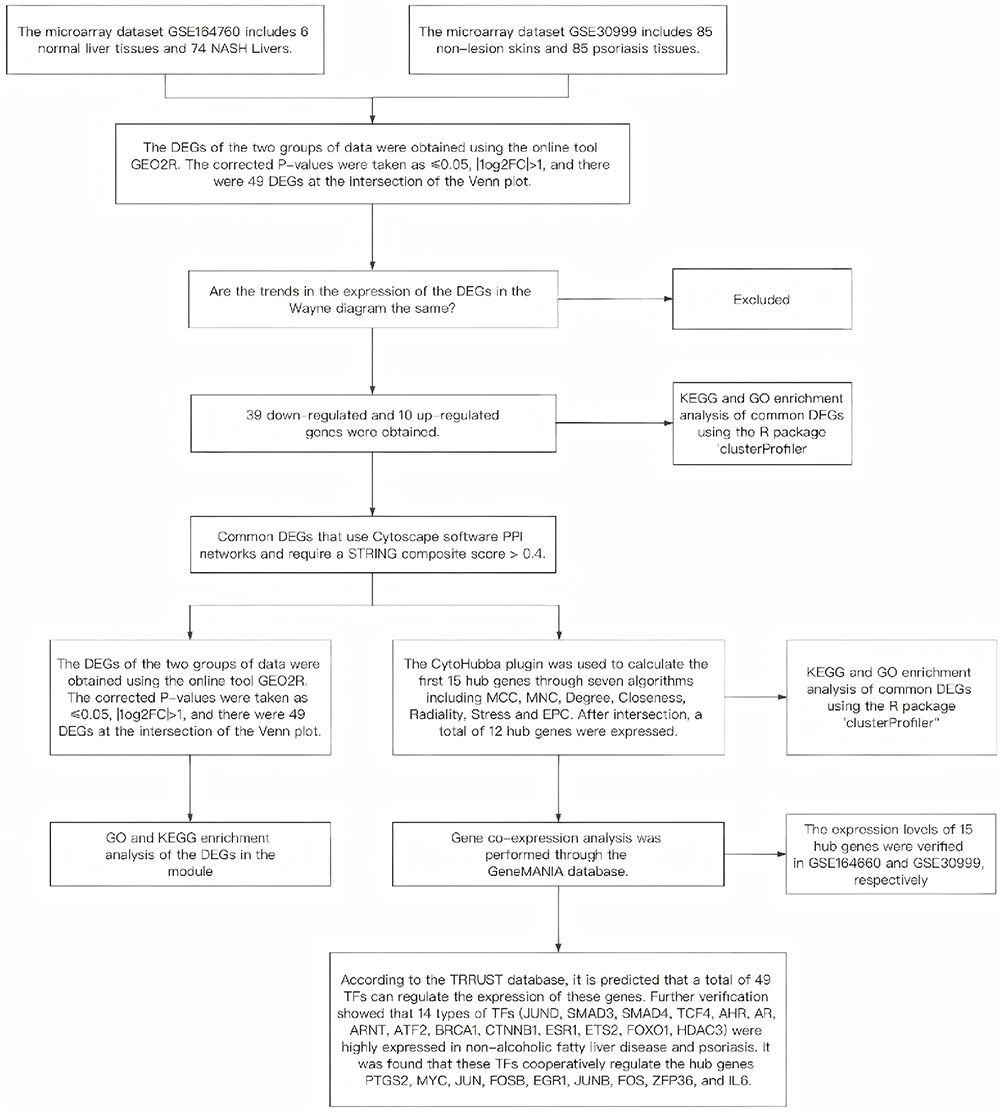 |
Figure 1 Research design flow chart. |
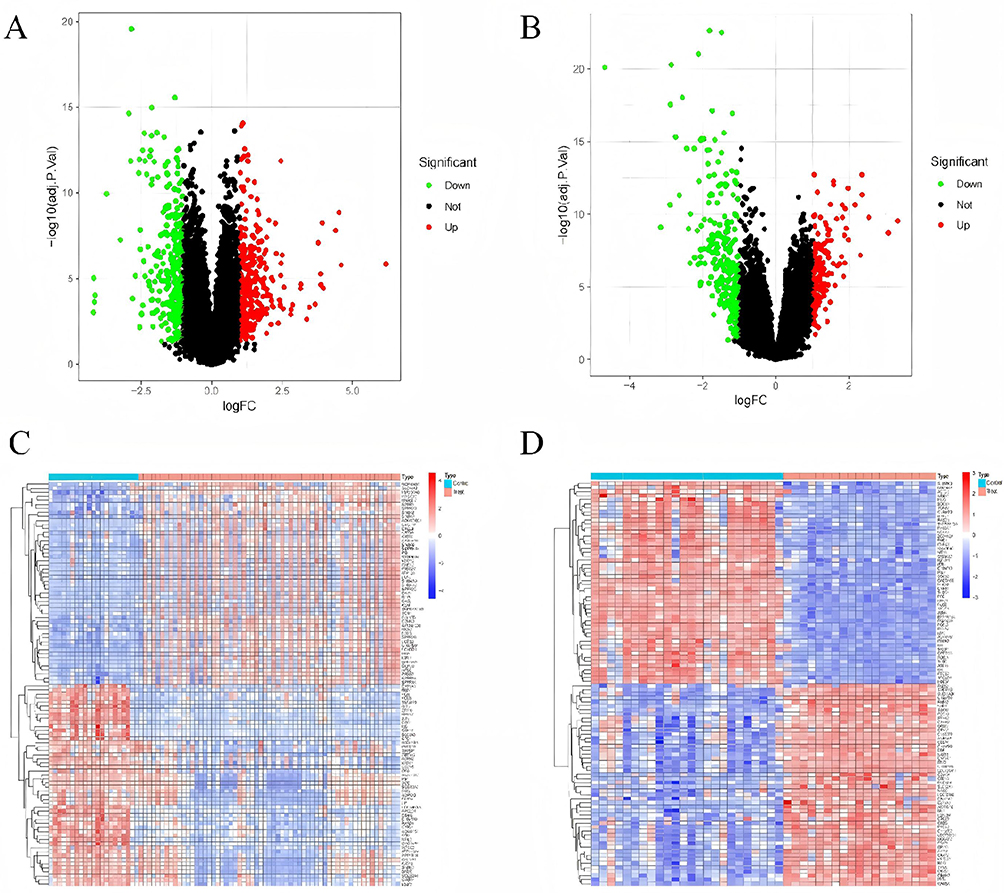 |
Figure 2 Continued. |
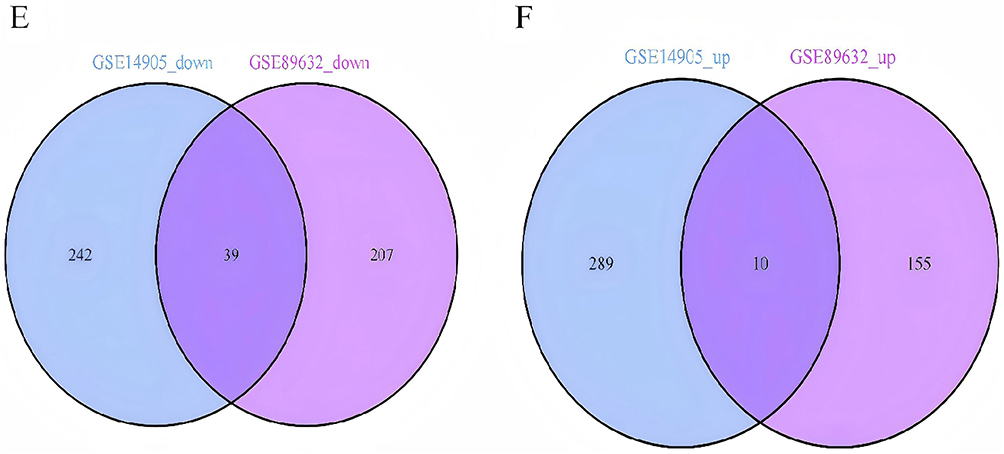 |
Figure 2 Dataset for psoriasis and non-alcoholic fatty liver disease. Volcano and Venn diagrams of differentially expressed genes (DEGs). (A) GSE14905 DEGs volcano map. (B) GSE89632 DEGs volcano map; Upregulated genes are marked with light red; The down-regulated gene is marked light green. (C) Heatmap showing GSE14905. (D) Heatmap showing GSE89632 and (E) Venn diagram showing GSE14905 and GSE89632 datasets downregulated by DEGs. (F) Venn plots of GSE14905 and GSE89632 datasets show upregulation of DEGs expression. |
Functional Analysis of DEGs
The selected Differentially Expressed Genes (DEGs) were analyzed more accurately using the “clusterProfiler” package in R to perform enrichment analysis on GO and KEGG pathways. A filtering condition of adjusted P < 0.05 was implemented, and the top 10 most significant GO (Figure 3B) and KEGG (Figure 3C) enrichments were chosen. Through Gene Ontology (GO) and KEGG enrichment analysis, it was revealed that the DEGs were significantly enriched in various biological processes, including responses to glucocorticoids and corticosteroids, integrated stress response signaling, steroid hormone response, positive regulation of miRNA transcription and metabolism, kidney development, miRNA transcription regulation, and renal system development. In terms of cellular components, the DEGs were predominantly associated with RNA polymerase II transcription regulatory complexes, paraspeckles, Golgi apparatus lumen, RNA polymerase II transcription repressor complexes, and CCR4−NOT complexes. Furthermore, the enrichment analysis uncovered that the DEGs were notably enriched in multiple functional categories pertaining to transcriptional regulation, such as DNA-binding transcription activator activity, RNA polymerase II specificity, DNA-binding transcription factor binding, nuclear glucocorticoid receptor binding, and RNA polymerase II-specific DNA-binding transcription factor binding. Notably, the important KEGG pathways enriched in the DEGs encompassed human T-cell leukemia virus type 1 infection, TNF signaling pathway, breast cancer, colorectal cancer, IL-17 signaling pathway, Kaposi’s sarcoma-associated herpesvirus infection, osteoclast differentiation, thyroid cancer, small cell lung cancer, and Toll-like receptor signaling pathway.
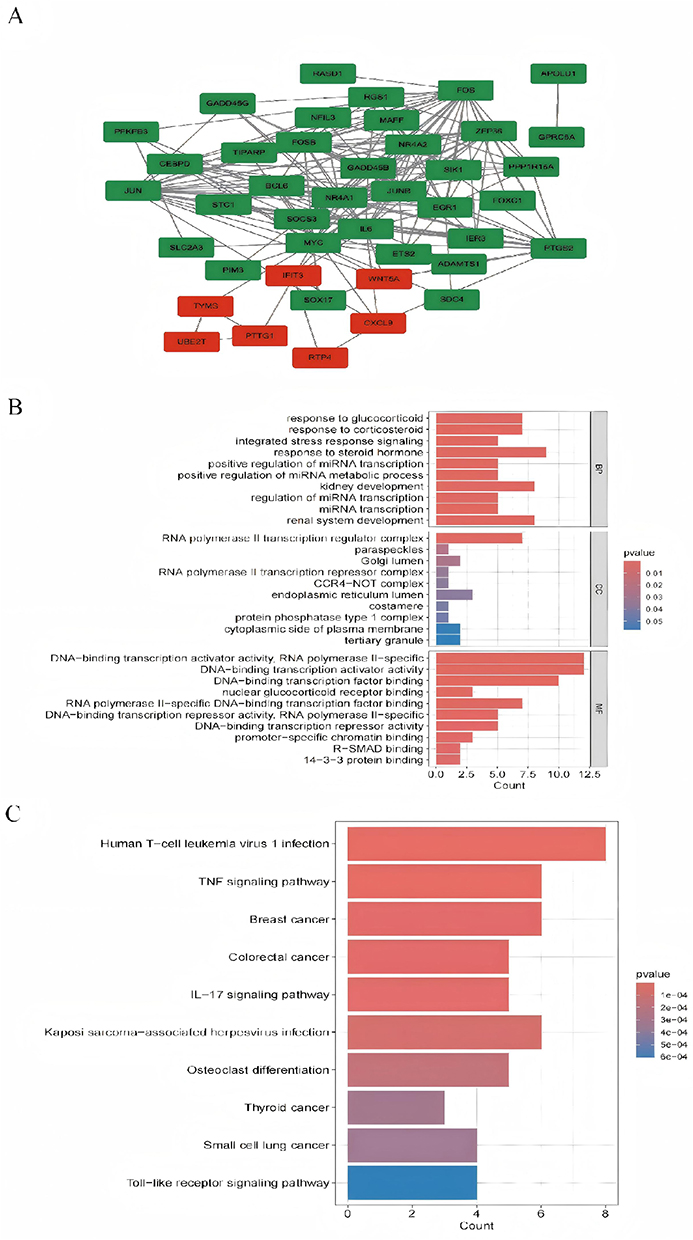 |
Figure 3 Protein–protein interaction network and enrichment analysis of common DEGs. (A) PPI network diagram. Red indicates up-regulated genes, and green indicates down-regulated genes. (B and C) The enrichment analysis results of GO and KEGG Pathway. Adjusted P-value < 0.05 was considered significant. |
PPI Network Analysis and Module Analysis
Using Cytoscape, we constructed a PPI network comprising co-expressed genes with a composite score exceeding 0.4. This network encompassed 183 interaction pairs and 42 nodes (Figure 3A). The MCODE plugin within Cytoscape identified two tightly interconnected subnetwork gene modules (Figure 4A and B). GO analysis demonstrated (Figure 4C) that the hub genes were predominantly associated with a range of biological processes, such as positive regulation of miRNA transcription, positive regulation of miRNA metabolic processes, response to steroid hormones, regulation of miRNA transcription, miRNA transcription itself, regulation of miRNA metabolic processes, miRNA metabolic processes, response to glucocorticoids, response to corticosteroids, and response to lipopolysaccharides.
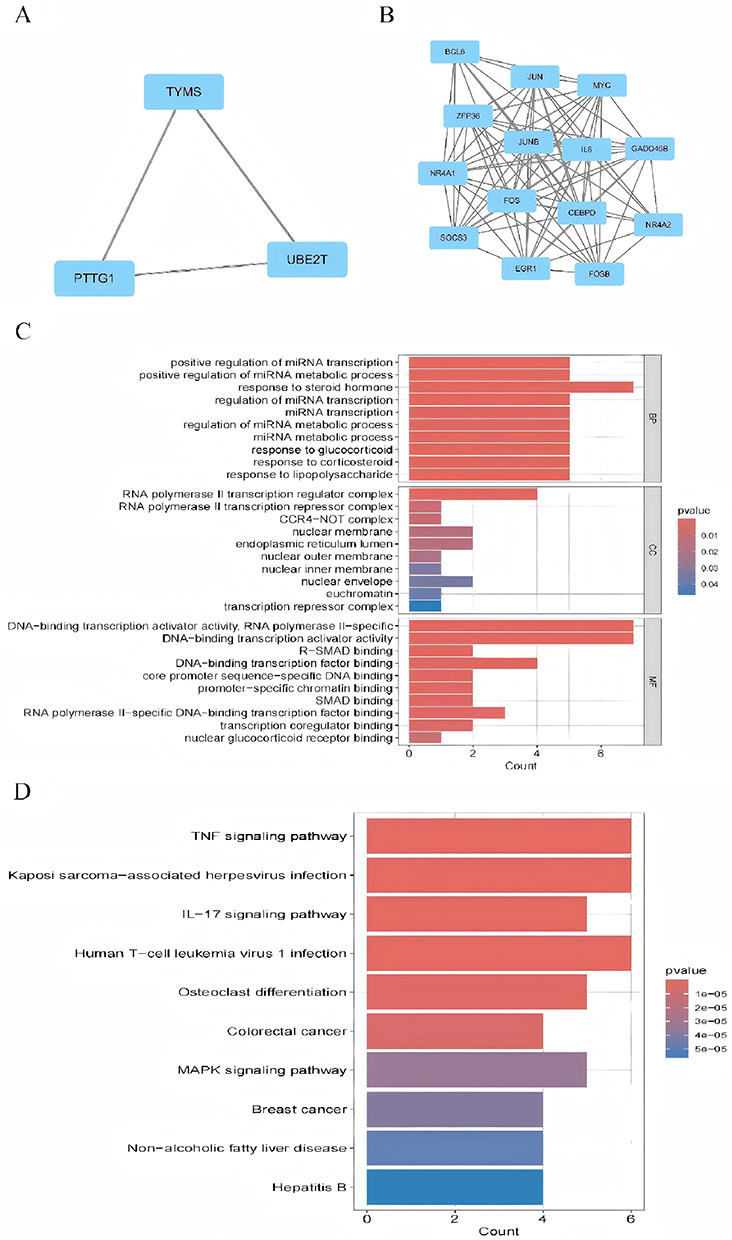 |
Figure 4 Presents the results of GO and KEGG pathway enrichment analysis for differentially expressed genes in psoriasis and nonalcoholic fatty liver disease, as outlined in the study. Panels (A and B) depict three closely related sub-network gene clustering modules. Panels (C and D) exhibit the enrichment analysis of modular genes in terms of GO and KEGG. The size of the circles in the figure signifies the number of genes, while the horizontal axis displays the frequency of occurrence of the participating genes among the total genes. Statistical significance is denoted by the p-value, with p<0.05 indicating significant enrichment. |
In terms of cellular components, hub genes are enriched in the RNA polymerase II transcription regulator complex, RNA polymerase II transcription repressor complex, CCR4−NOT complex, nuclear membrane, endoplasmic reticulum lumen, nuclear outer membrane, nuclear inner membrane, nuclear envelope, euchromatin, and transcription repressor complex. Molecular Function (MF) analysis has shown that hub genes are predominantly enriched in activities such as DNA-binding transcription activator activity specific to RNA polymerase II, DNA-binding transcription activator activity, R-SMAD binding, DNA-binding transcription factor binding, core promoter sequence-specific DNA binding, promoter-specific chromatin binding, SMAD binding, RNA polymerase II-specific DNA-binding transcription factor binding, transcription coregulator binding, and nuclear glucocorticoid receptor binding. Additionally, the results of the KEGG pathway analysis (Figure 4D) demonstrate that hub genes exhibit significant enrichment in the KEGG pathway enrichment analysis, primarily involving pathways such as TNF signaling, Kaposi’s sarcoma-associated herpesvirus infection, IL-17 signaling, human T-cell leukemia virus 1 infection, osteoclast differentiation, colorectal cancer, MAPK signaling, breast cancer, non-alcoholic fatty liver disease, and hepatitis B.
Hub Gene Identification
We employed the CytoHubba software to identify the top 15 hub genes using seven algorithms: MCC, MNC, Degree, Closeness, Radiality, Stress, and EPC. Following intersection analysis, we confirmed 12 common hub genes (Figure 5A and Table 2). Additionally, we utilized the GeneMANIA online database to analyze the co-expression network and associated functions of these hub genes. The results demonstrated that these 15 differentially expressed genes (DEGs) formed an intricate protein-protein interaction (PPI) network, comprising 75.18% co-expressed genes, 13.84% physical interactions, 5.95% predicted interactions, 2.96% co-localization, 1.07% pathways, and 1.01% shared protein domains (Figure 5B). Gene Ontology (GO) analysis revealed that these genes are primarily involved in the positive regulation of miRNA transcription, positive regulation of miRNA metabolic processes, and steroid hormone responses, among others (Figure 5C). Furthermore, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis indicated that these genes are mainly associated with the TNF signaling pathway, herpesvirus infection associated with Kaposi’s sarcoma, the IL-17 signaling pathway, and others (Figure 5D).
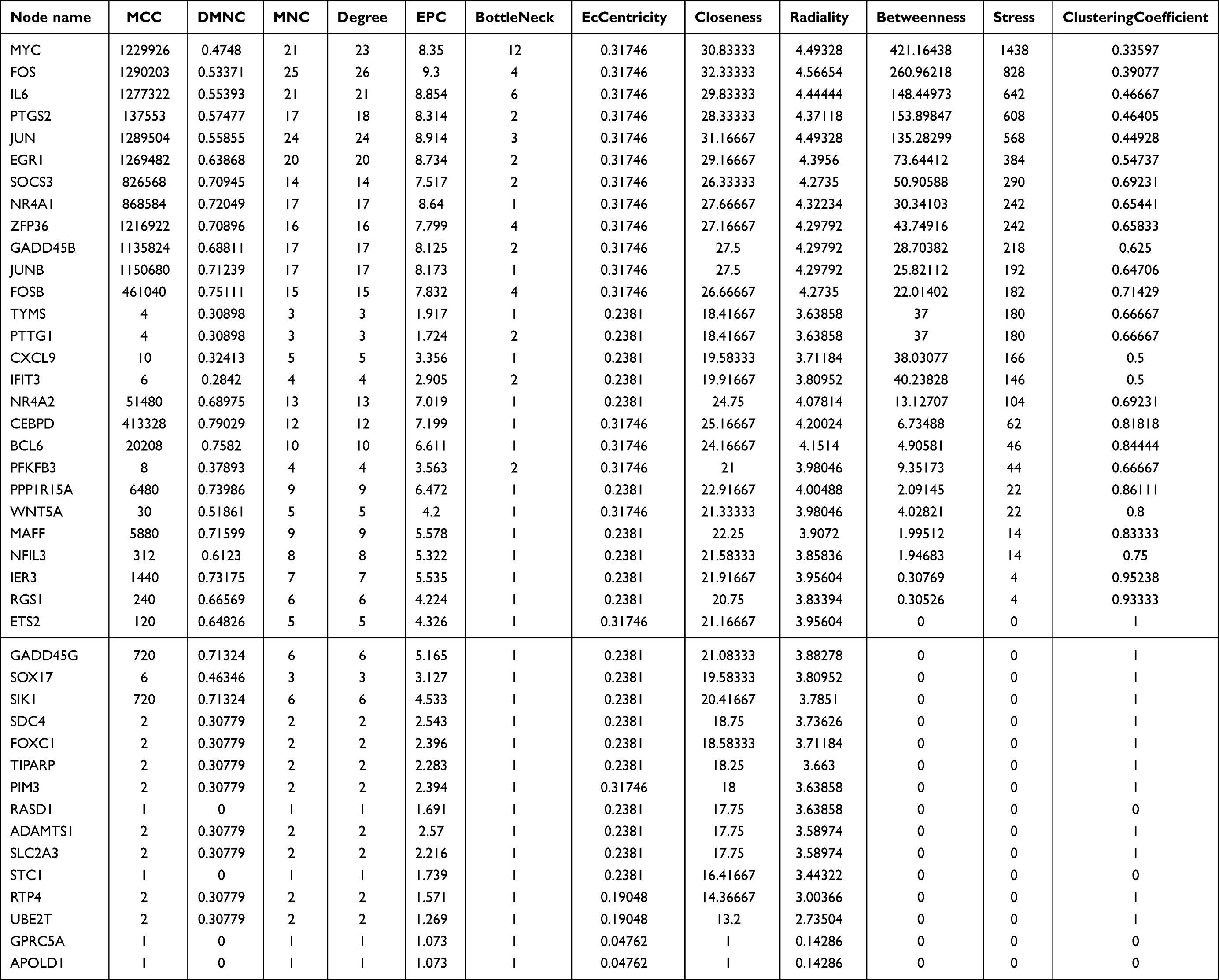 |
Table 2 Independent Scoring and Ranking of Candidate Genes by Seven Algorithms |
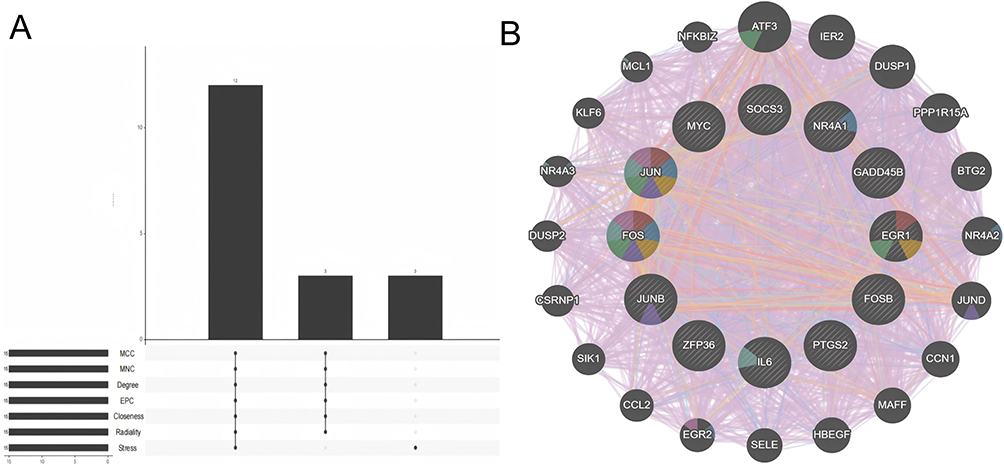 |
Figure 5 Continued. |
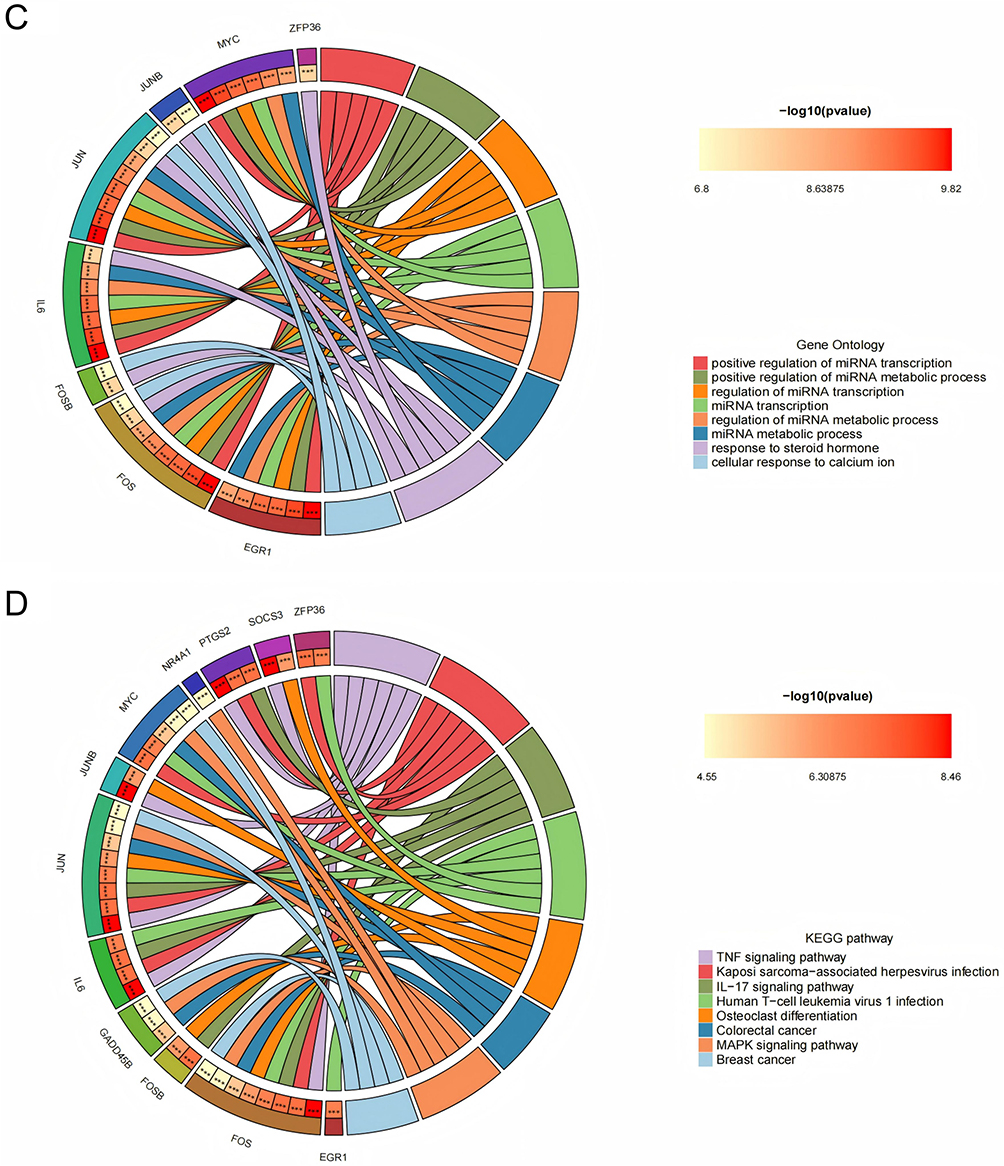 |
Figure 5 Analysis of core hub genes in psoriasis and NAFLD. (A) UpSet plot showing the intersection of top 15 genes from seven algorithms (MCC, MNC, Degree, EPC, Closeness, Radiality, Stress) for screening 12 core hub genes. (B) GeneMANIA analysis of core hub genes and their co-expressed gene interaction networks. (C) GO enrichment analysis of core hub genes. (D) KEGG enrichment analysis of core hub genes (enriched in PPAR/IL-17 signaling pathways); the outer circle is enrichment terms, inner circle is adjusted p-values. |
To validate the reliability of the expression levels of these hub genes, we utilized two additional datasets pertaining to non-alcoholic fatty liver disease (NAFLD) and psoriasis to assess the mRNA expression of the identified core genes. The results indicated that, when compared to normal tissues, all hub genes, with the exception of EGR1, were notably upregulated in psoriatic skin lesions. Likewise, in NAFLD, all core genes, excluding IL6, MYC, and PTGS2, demonstrated expression levels that surpassed those in normal tissues.
Predict TF and Validate
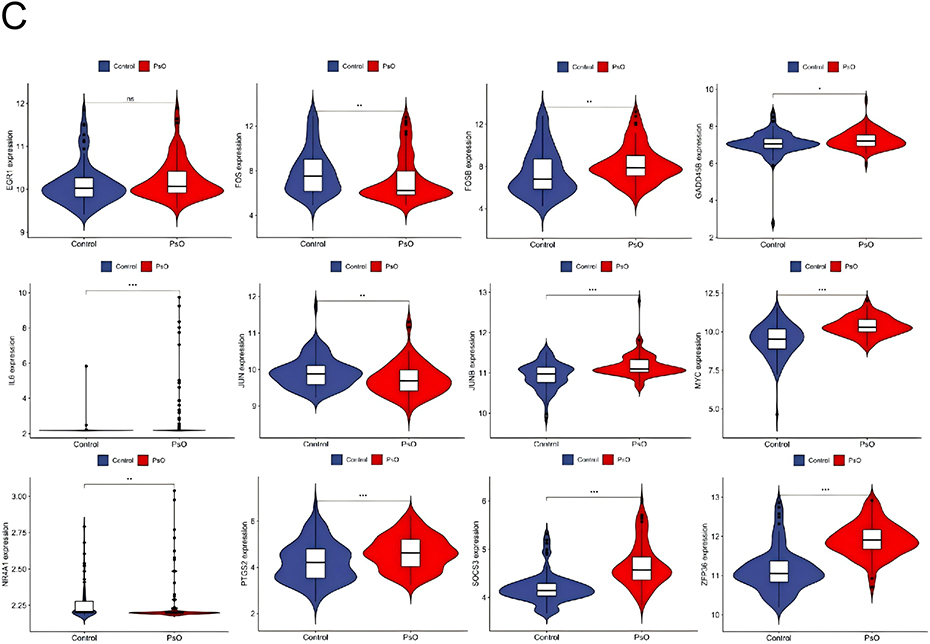
According to the TRRUST database, a total of 49 TFs are predicted to regulate the expression of these genes. Further verification revealed that 14 TFs (JUND, SMAD3, SMAD4, TCF4, AHR, AR, ARNT, ATF2, BRCA1, CTNNB1, ESR1, ETS2, FOXO1, HDAC3) exhibit high-level expression in non-alcoholic fatty liver disease and psoriasis, and it was found that these TFs jointly regulate hub genes PTGS2, MYC, JUN, FOSB, EGR1, JUNB, FOS, ZFP36, and IL6 (Figure 6A–C).
Figure 6 Continued. Figure 6 TFs regulate networks and their expression in GSE14905 and GSE89632. (A) Retrotransposon regulation, with green representing retrotransposon and yellow representing pivot genes. Expression levels of retrotransposons in (B and C) GSE14905 and GSE89632. The ere compared using the mean t-test. A P-value < 0.05 indicates statistical significance. *p < 0.05; **p < 0.01; p < 0.001.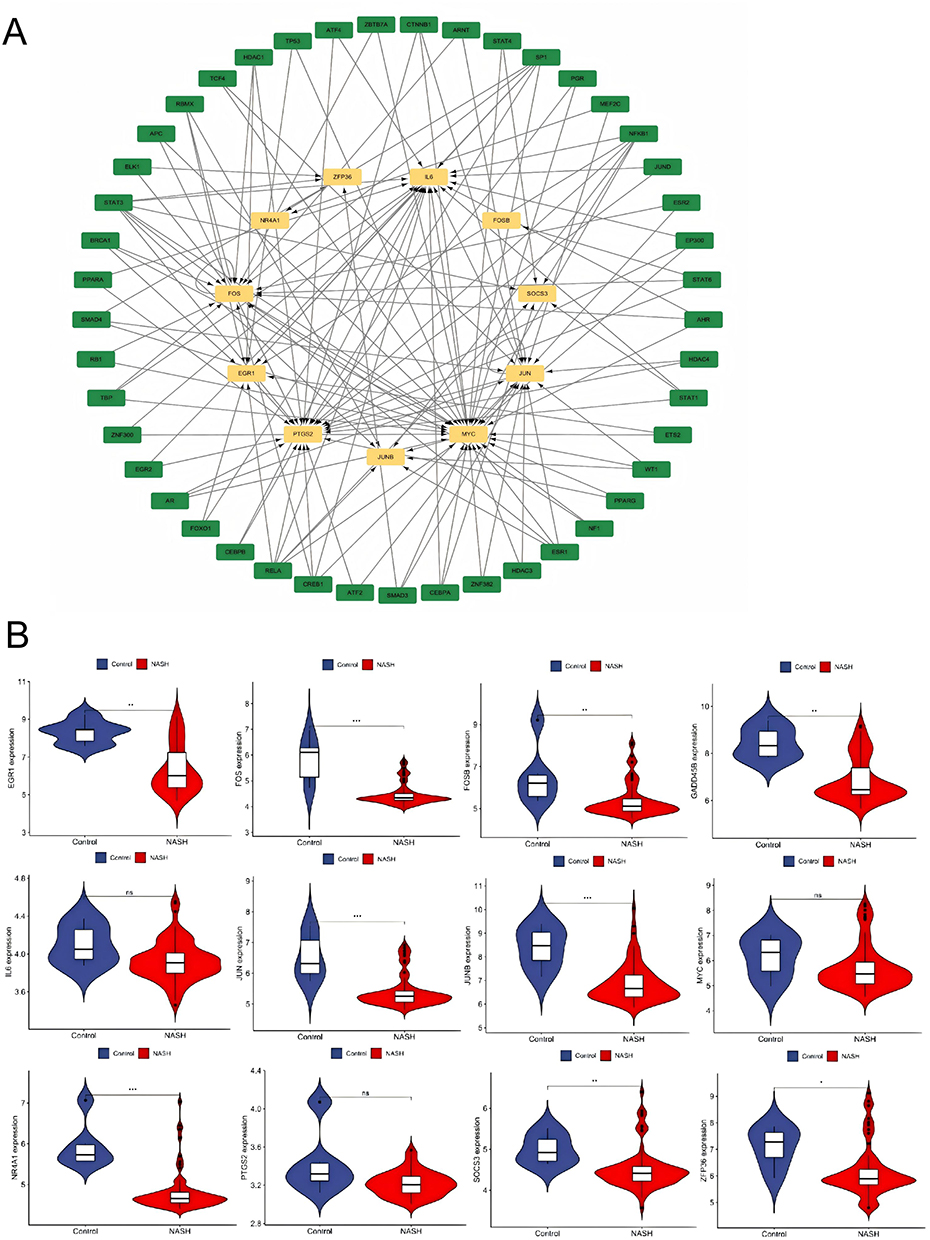
Discussion
Although NAFLD and psoriasis manifest as liver fat deposition and cutaneous immune inflammation, respectively,25 their pathogenesis is intricately linked to chronic inflammation, encompassing endothelial dysfunction, oxidative stress, and a cytokine network imbalance.26 NAFLD is influenced by genetics, metabolism, and gut microbiota, leading to severe disruptions in hepatic lipid metabolism. This, in turn, causes hepatocyte damage and the release of pro-inflammatory factors such as TNF-α and IL-6, which activate systemic immune responses. Conversely, psoriasis arises from the excessive activation of T helper (Th) 1 / Th17 cells, producing cytokines like IL-17 and TNF-α, which drive abnormal keratinocyte proliferation and epidermal inflammation.27 The shared pro-inflammatory cytokines in these two diseases activate crucial signaling pathways, such as JAK-STAT and NF-κB, establishing a bidirectional inflammation axis between the liver and skin. Hepatic inflammation in NAFLD can exacerbate cutaneous immune responses, while cutaneous inflammatory factors in psoriasis can reciprocate on the liver, promoting fat accumulation and liver damage.28 Metabolic abnormalities also play a pivotal role in the interplay between these diseases. The lipid metabolism disorder in NAFLD and the abnormal synthesis of epidermal barrier lipids in psoriasis interact, with studies showing a positive correlation between serum lipid levels and the severity of psoriatic inflammation,29 suggesting a role for lipid-inflammatory interactions in the progression of comorbidity.
Despite NAFLD and psoriasis belonging to the categories of metabolic liver disease and immune-mediated skin disease, respectively, their pathogenesis is closely tied to chronic inflammation, immune regulation, and metabolic abnormalities. This study aims to screen for common differentially expressed genes (DEGs) between the two diseases, uncover potential therapeutic targets, and delve into the mechanisms underlying their comorbidity. By utilizing gene microarray data from the GEO database for psoriasis (GSE30999, GSE14905) and NAFLD (GSE89632, GSE164760), we identified a total of 49 common DEGs through differential expression analysis and cross-validation, with 10 upregulated and 39 downregulated genes. Through protein-protein interaction (PPI) network and hub gene analysis, we ultimately pinpointed 12 core genes: FOS, EGR1, SOCS3, PTGS2, JUN, IL6, NR4A1, ZFP36, MYC, FOSB, JUNB, and GADD45B.
Among these core hub genes, IL6 and EGR1 serve as pivotal bridge nodes mediating the pathological crosstalk between psoriasis and NAFLD. Previous studies have extensively documented their roles in single diseases: IL6 is well-recognized in psoriasis for driving keratinocyte proliferation via the STAT3 axis and in NAFLD for promoting insulin resistance and hepatic stellate cell activation.15 However, our study advances this understanding by identifying IL6 not just as a local effector, but as a systemic connector regulated by a shared transcriptional network. In the comorbid state, circulating IL6 derived from psoriatic skin lesions may exacerbate hepatic inflammatory and metabolic disorders, while hepatocyte-secreted IL6 further activates the skin immune microenvironment, forming a positive feedback loop of “skin inflammation-hepatic lipid metabolism disorder”.
Regarding EGR1, it functions as a transcriptional regulator of lipid metabolism genes (eg, SREBP1, PPARα) in the liver and cell cycle progression in skin.30 Notably, our validation results showed that unlike other hub genes, EGR1 was not significantly upregulated in psoriatic lesions. This discrepancy can be biologically explained by the nature of EGR1 as an “immediate-early gene” with rapid but transient expression kinetics. In chronic psoriatic plaques (typical of validation datasets), the acute transcriptional surge of EGR1 may have subsided or been dampened by negative feedback loops, whereas its downstream effects on metabolic reprogramming persist. Furthermore, potential prior treatment with topical corticosteroids in clinical samples effectively inhibits EGR1 expression, which may mask its initial upregulation. Despite this, its identification as a hub gene in the discovery phase suggests it acts as a “trigger” in the early initiation of the comorbidity, disrupting the homeostasis of hepatic lipid metabolism and cutaneous immunity simultaneously.30
GO enrichment analysis revealed that the common DEGs in NAFLD and psoriasis were significantly enriched in biological processes such as “response to glucocorticoid”, “positive regulation of miRNA transcription”, and “integrated stress response signaling”, highlighting the crucial roles of steroid hormone metabolism and non-coding RNA-mediated transcriptional regulation in both diseases.31 KEGG pathway analysis indicated that DEGs were primarily enriched in pathways associated with “IL-17 signaling”, “non-alcoholic fatty liver disease”, and “human T-cell leukemia virus 1 infection”, underscoring the importance of immune regulation, lipid metabolism, and inflammation signaling networks linked to viral infections in comorbidities.32 These findings suggest that the identified DEGs are primarily implicated in the “transcription regulation-inflammation-metabolism” interconnected network, with transcription factors and cytokines playing pivotal roles.32 For instance, the hub genes MYC and PTGS2 were enriched in “positive regulation of miRNA transcription” in GO analysis, potentially impacting keratinocyte proliferation and hepatocyte lipid metabolism by regulating miRNA biosynthesis. IL6, as a key cytokine, was associated with the “response to glucocorticoid” pathway, hinting at its significant role in amplifying inflammation in both diseases.33 In psoriasis, the activation of the IL-17 signaling pathway promotes abnormal keratinocyte differentiation, whereas in NAFLD, genes related to the “non-alcoholic fatty liver disease” pathway regulate hepatocyte fatty acid β-oxidation, confirming the bidirectional relationship between metabolism and immunity. Notably, unlike previous studies that only focused on the independent roles of pro-inflammatory cytokines such as IL6 and TNF-α in psoriasis or NAFLD alone, this study innovatively integrates gene expression profiles of the two diseases, screens for shared DEGs and core hub genes, and clarifies the bidirectional regulatory molecular network of key genes represented by IL6 and EGR1 in the comorbidity of psoriasis and NAFLD.34 For the first time, we reveal the cross-disease regulatory characteristics of the “transcription factor-hub gene” axis (eg, JUND regulating IL6, SMAD4 targeting MYC) in mediating skin-liver pathological crosstalk, and construct a multi-dimensional regulatory model of psoriasis-NAFLD comorbidity from the perspectives of gene co-expression, functional pathway enrichment and transcriptional regulation.35 This study breaks the limitation of single-disease research on the two diseases, and provides a novel cross-disease molecular target screening strategy for the exploration of comorbidity mechanisms, which is different from the existing research that only focuses on a single inflammatory factor or signaling pathway.36
Furthermore, the regulatory network analysis reveals a hierarchical multi-layer topology driving the comorbidity, rather than a flat list of interactions. At the upstream apex, transcription factors JUND and SMAD3/4 function as “master regulators”. They do not merely bind to isolated targets but orchestrate a dense core regulatory module centered on the AP-1 complex (JUN, FOS, JUNB). This structure suggests a “feed-forward” mechanism: upstream JUND activates the expression of AP-1 components, which then physically interact (heterodimerize) to cooperatively bind the promoters of downstream effector genes such as IL6 and PTGS2.
This topological arrangement creates a signal amplification cascade. In psoriasis, the JUND-driven AP-1 accumulation ensures sustained high-level transcription of inflammatory cytokines (IL6), establishing a robust inflammatory state that resists minor fluctuations. Simultaneously, in the liver, the crosstalk between the SMAD3 (a key node in TGF-β signaling) and the MYC-related metabolic axis acts as a bridge between fibrosis and lipid synthesis. Specifically, JUN activates the MYC-mediated fatty acid synthesis pathway through synergistic action with SMAD3, inducing SREBP1 downstream genes to promote hepatic triglyceride accumulation.37
Within this network, ZFP36 acts as a crucial post-transcriptional regulatory node providing a counter-balance. As an RNA-binding protein,38 ZFP36 targets the mRNA 3’UTR regions of upstream drivers (like JUN) and downstream effectors (like IL6/MYC) to promote their degradation. This topology indicates a failed negative feedback loop in the disease state: while ZFP36 attempts to dampen the signal, the overwhelming activation of the JUND/SMAD axis likely bypasses this checkpoint, locking the system into a chronic cycle of inflammation and metabolic dysregulation.
Under the synergistic regulation of transcription factors (TFs) such as SMAD3, the promoter region of IL6 is enriched with binding sites for JUND. This, in turn, activates the differentiation process of Th17 cells via the JAK-STAT pathway in psoriasis, further prompting keratinocytes to secrete IL-17A and antimicrobial peptides, thereby exacerbating the inflammatory response in the epidermis. In NAFLD, IL6 induces insulin resistance in hepatocytes, inhibits PPARγ-mediated fatty acid oxidation, and promotes the activation and fibrosis of hepatic stellate cells.10 PTGS2, as a key enzyme in prostaglandin synthesis, catalyzes the conversion of arachidonic acid into PGE2 after being regulated by JUND. Its high expression is closely linked to the activation of TNF and IL-17 signaling pathways, participating in inflammation amplification in both diseases.
MYC, as a direct target gene of SMAD4, accelerates the cell cycle progression of keratinocytes by regulating Cyclin D1 in psoriasis. In NAFLD, it promotes de novo fatty acid synthesis by activating SREBP1. The abnormal expression of MYC aligns with the enrichment results of the “Non-alcoholic fatty liver disease” KEGG pathway. ZFP36, an RNA-binding protein,38 is regulated by TFs and binds to the mRNA 3’UTR regions of MYC and IL6, inhibiting their degradation and maintaining the continuous expression of pro-inflammatory and pro-metabolic genes, thereby forming a dual regulatory mechanism of “transcription-post-transcription”.
This study also has certain limitations that need to be acknowledged. First, the research conclusions are mainly based on bioinformatics analysis of public microarray datasets, and the regulatory relationship between transcription factors and hub genes is only predicted by bioinformatics databases without further verification by in vitro and in vivo wet experiments such as dual-luciferase reporter assay and ChIP-qPCR. Second, due to the lack of clinical sample data, the correlation between the expression levels of core hub genes and the clinical severity of psoriasis-NAFLD comorbidity has not been further analyzed. Third, the current study only focuses on the mRNA expression level, and the post-transcriptional and post-translational modification regulation of core genes in the two diseases has not been explored in depth. In subsequent studies, we propose to conduct specific intervention experiments targeting the identified core transcription factors, particularly JUND and SMAD3. By utilizing specific small-molecule inhibitors or gene-silencing techniques (eg, siRNA or CRISPR/Cas9) in dual-disease animal models (co-induced psoriasis and NAFLD), we aim to verify their regulatory effects on the comorbidity. Specifically, we will assess whether blocking these upstream TFs can simultaneously alleviate psoriatic skin lesions and hepatic steatosis, thereby confirming the “transcription factor-hub gene” axis as a viable therapeutic target for interrupting the cross-disease pathological network.39
Abbreviations
NAFLD, non-alcoholic fatty liver disease; DEGs, differentially expressed genes; NASH, non-alcoholic steatohepatitis; PPI, protein-protein interaction; NF-κB, nuclear factor κB; MAPK, mitogen-activated protein kinase; JAK-STAT, Janus kinase-signal transducer and activator of transcription; PPAR, peroxisome proliferator-activated receptor; SREBP, sterol regulatory element-binding protein; TNF-α, tumor necrosis factor-α; IL-17interleukin-17; GEO, Gene Expression Omnibus; TRRUST, Transcriptional Regulatory Relationships Unraveled by Sentence-based Text mining; KEGG, Kyoto Encyclopedia of Genes and Genomes.
Data Sharing Statement
The data are available from the corresponding author, Qiwen Wu.
Ethical Approval and Consent to Participate
This study utilized publicly available data from open-access databases and did not involve any sensitive personal information or commercial interests. According to Article 32 of the Ethical Review Measures for Human Life Science and Medical Research approved by the National Science and Technology Ethics Committee of the People’s Republic of China, ethical review may be exempted.
Acknowledgments
The authors acknowledge the support of the public resources.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the Youth Teacher Training Project of Education Department of Anhui Province (No. JWFX2024028) and Anhui Province Key Clinical Specialist Construction Programs.
Disclosure
The authors declare no competing interests in this work.
References
1. Xiao P, Ye Z, Li X, Feng Q, Su Y. Ginseng and its functional components in non-alcoholic fatty liver disease: therapeutic effects and multi-target pharmacological mechanisms. Front Pharmacol. 2025;16:1540255. doi:10.3389/fphar.2025.1540255
2. Wang X, Chen H, Zhu W, et al. Akebia trifoliata extracts attenuate liver injury via gut-liver axis in a murine model of nonalcoholic fatty liver disease with low-grade colitis. Food Res Int. 2025;208:116202. doi:10.1016/j.foodres.2025.116202
3. Younossi Z, Anstee QM, Marietti M, et al. The global burden of NAFLD and NASH is escalating, with trends indicating a significant increase in prevalence and associated health risks. Predictions suggest that the incidence of NASH-related liver cancer will rise, posing a substantial public health challenge. Risk factors such as obesity and diabetes are contributing to the growing number of cases, and prevention strategies are urgently needed. Nat Rev Gastroenterol Hepatol. 2018;15(1):11–18. doi:10.1038/nrgastro.2017.109
4. Moon AM, Singal AG, Tapper EB. Contemporary epidemiology of chronic liver disease and cirrhosis. Clin Gastroenterol Hepatol. 2020;18(12):2650–2666. doi:10.1016/j.cgh.2019.07.060
5. Griffiths CE, Barker JN. Pathogenesis and clinical features of psoriasis. Lancet. 2007;370(9583):263–271. doi:10.1016/S0140-6736(07)61128-3
6. Kamiya K, Kishimoto M, Sugai J, Komine M, Ohtsuki M. Risk factors for the development of psoriasis. Int J Mol Sci. 2019;20(18):4347. doi:10.3390/ijms20184347
7. Ruan Z, Lu T, Chen Y, et al. Association between psoriasis and nonalcoholic fatty liver disease among outpatient US adults. JAMA Dermatol. 2022;158(7):745–753. doi:10.1001/jamadermatol.2022.1609
8. Bellinato F, Maurelli M, Geat D, Girolomoni G, Gisondi P. Managing the patient with psoriasis and metabolic comorbidities. Am J Clin Dermatol. 2024;25(4):527–540. doi:10.1007/s40257-024-00857-0
9. Yao Y, Luo ZP, Li HW, et al. P38γ modulates the lipid metabolism in non-alcoholic fatty liver disease by regulating the JAK-STAT signaling pathway. FASEB J. 2023;37(1):e22716. doi:10.1096/fj.202200939RR
10. Wang F, Zhang X, Liu W, et al. Activated natural killer cell promotes nonalcoholic steatohepatitis through mediating JAK/STAT pathway. Cell Mol Gastroenterol Hepatol. 2022;13(1):257–274. doi:10.1016/j.jcmgh.2021.08.019
11. Chen H, Tan H, Wan J, et al. PPAR-γ signaling in nonalcoholic fatty liver disease: pathogenesis and therapeutic targets. Pharmacol Ther. 2023;245:108391. doi:10.1016/j.pharmthera.2023.108391
12. Kawamura S, Matsushita Y, Kurosaki S, et al. Inhibiting SCAP/SREBP exacerbates liver injury and carcinogenesis in murine nonalcoholic steatohepatitis. J Clin Invest. 2022;132(11):e151895. doi:10.1172/JCI151895
13. Ghoreschi K, Balato A, Enerbäck C, Sabat R. Therapeutics targeting the IL-23 and IL-17 pathway in psoriasis. Lancet. 2021;397(10275):754–766. doi:10.1016/S0140-6736(21)00184-7
14. Blauvelt A, Chiricozzi A. The immunologic role of IL-17 in psoriasis and psoriatic arthritis pathogenesis. Clin Rev Allergy Immunol. 2018;55(3):379–390. doi:10.1007/s12016-018-8702-3
15. Park J, Zhao Y, Zhang F, et al. IL-6/STAT3 axis dictates the PNPLA3-mediated susceptibility to non-alcoholic fatty liver disease. J Hepatol. 2023;78(1):45–56. doi:10.1016/j.jhep.2022.08.022
16. Barrett T, Wilhite SE, Ledoux P, et al. NCBI GEO: archive for functional genomics data sets--update. Nucleic Acids Res. 2013;41(Database issue):D991–D995. doi:10.1093/nar/gks1193
17. Kanehisa M, Goto S. KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000;28(1):27–30. PMID: 10592173; PMCID: PMC102409. doi:10.1093/nar/28.1.27
18. Franceschini A, Szklarczyk D, Frankild S, et al. STRING v9.1: protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013;41(Database issue):D808–15. PMID: 23203871; PMCID: PMC3531103. doi:10.1093/nar/gks1094
19. Charles S, Natarajan J. Identification of key gene modules and novel transcription factors in tetralogy of fallot using machine learning and network topological features. Medinformatics. 2024;1(1):27–34. doi:10.47852/bonviewMEDIN32021554
20. Smoot ME, Ono K, Ruscheinski J, Wang PL, Ideker T. Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics. 2011;27(3):431–432. PMID: 21149340; PMCID: PMC3031041. doi:10.1093/bioinformatics/btq675
21. Warde-Farley D, Donaldson SL, Comes O, et al. The GeneMANIA prediction server: biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010;38(Web Server issue):W214–20. PMID: 20576703; PMCID: PMC2896186. doi:10.1093/nar/gkq537
22. Chen Q, Bei S, Zhang Z, Wang X, Zhu Y. explore the identification of diagnostic biomarkers and the analysis of immune cell infiltration in ulcerative colitis, a chronic inflammatory bowel disease. The study builds upon recent advances in nanotechnology and immunotherapy, which have shown promise in targeting treatment and improving diagnostic accuracy for more personalized patient management. Sci Rep. 2023;13(1):6081. doi:10.1038/s41598-023-33388-5
23. Su W, Zhao Y, Wei Y, Zhang X, Ji J, Yang S. Exploring the pathogenesis of psoriasis complicated with atherosclerosis via microarray data analysis. Front Immuno. 2021;12:667690. doi:10.3389/fimmu.2021.667690
24. Han H, Cho JW, Lee S, et al. TRRUST v2: an expanded reference database of human and mouse transcriptional regulatory interactions. Nucleic Acids Res. 2018;46(D1):D380–D386. PMID: 29087512; PMCID: PMC5753191. doi:10.1093/nar/gkx1013
25. Muzica CM, Sfarti C, Trifan A, et al. Nonalcoholic fatty liver disease and type 2 diabetes mellitus: a bidirectional relationship. Can J Gastroenterol Hepatol. 2020;2020:6638306. doi:10.1155/2020/6638306
26. Nasiri-Ansari N, Androutsakos T, Flessa CM, et al. Endothelial cell dysfunction and nonalcoholic fatty liver disease (NAFLD): a concise review. Cells. 2022;11(16):2511. doi:10.3390/cells11162511
27. Bar-Or A, Fawaz L, Fan B, et al. Abnormal B-cell cytokine responses a trigger of T-cell-mediated disease in MS? Ann Neurol. 2010;67(4):452–461. doi:10.1002/ana.21939
28. Guo L, Kircik L, Armstrong AW. INDIVIDUAL ARTICLE: psoriasis and Obesity: optimizing pharmacologic treatment and lifestyle interventions. J Drugs Dermatol. 2025;24(1):491722s4–491722s14.
29. Zhang ZY, Jian ZY, Tang Y, Li W. The relationship between blood lipid and risk of psoriasis: univariable and multivariable mendelian randomization analysis. Front Immunol. 2023;14:1174998. doi:10.3389/fimmu.2023.1174998
30. Jeon Y, Jung Y, Kim MC, et al. Sargahydroquinoic acid inhibits TNFα-induced AP-1 and NF-κB signaling in HaCaT cells through PPARα activation. Biochem Biophys Res Commun. 2014;450(4):1553–1559. doi:10.1016/j.bbrc.2014.07.026
31. Wang X, Yang D. The regulation of RNA metabolism in hormone signaling and breast cancer. Mol Cell Endocrinol. 2021;529:111221. doi:10.1016/j.mce.2021.111221
32. Meissner EG, Lee YJ, Osinusi A, et al. The study examines the impact of sofosbuvir and ribavirin therapy on lipid metabolism in patients with chronic hepatitis C virus genotype 1, considering the potential adverse effects on peripheral and hepatic lipid profiles. Hepatology. 2015;61(3):790–801. doi:10.1002/hep.27424
33. Koga T, Sumiyoshi R, Kawakami A, Yoshizaki K. A benefit and the prospects of IL-6 inhibitors in idiopathic multicentric Castleman’s disease. Mod Rheumatol. 2019;29(2):302–305. doi:10.1080/14397595.2018.1532383
34. Sahrawat TR, Sharma R, Dange S, Patial R, Gahlawat SK. Interactome-based computational approach to identify association between cardiomyopathy and cardiovascular disease. Medinformatics. 2025;2(2):99–106. doi:10.47852/bonviewMEDIN42023102
35. Mantovani A, Gisondi P, Lonardo A, Targher G. Relationship between non-alcoholic fatty liver disease and psoriasis: a novel hepato-dermal axis? Int J Mol Sci. 2016;17(2):217. doi:10.3390/ijms17020217
36. Lowes MA, Suarez-Farinas M, Krueger JG. Immunology of psoriasis. Ann Rev Immunol. 2014;32(1):227–255. doi:10.1146/annurev-immunol-032713-120225
37. Liu Y, Sun Z, Dong R, et al. Rutin ameliorated lipid metabolism dysfunction of diabetic NAFLD via AMPK/SREBP1 pathway. Phytomedicine. 2024;126:155437. doi:10.1016/j.phymed.2024.155437
38. Zhang Z, Guo M, Li Y, et al. RNA-binding protein ZFP36/TTP protects against ferroptosis by regulating autophagy signaling pathway in hepatic stellate cells. Autophagy. 2020;16(8):1482–1505. doi:10.1080/15548627.2019.1687985
39. Candia R, Ruiz A, Torres‐Robles R, Chávez‐Tapia N, Méndez‐Sánchez N, Arrese M. Risk of non‐alcoholic fatty liver disease in patients with psoriasis: a systematic review and meta‐analysis. J Eur Acad Dermatol Venereol. 2015;29(4):656–662. doi:10.1111/jdv.12847
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Transcriptome Analysis Identifies Biomarkers for the Diagnosis and Management of Psoriasis Complicated with Depression
Xia X, Yu H, Li Y, Liang Y, Li G, Huang F
Clinical, Cosmetic and Investigational Dermatology 2023, 16:1287-1301
Published Date: 18 May 2023
Study on the Key Genes and Molecular Mechanisms of IL-27 Promoting Keratinocytes Proliferation Based on Transcriptome Sequencing
Wu Z, Yang Q, Xu K, Wu J, Yang B
Clinical, Cosmetic and Investigational Dermatology 2023, 16:1457-1472
Published Date: 7 June 2023
Association Between Psoriasis and Non-Alcoholic Fatty Liver Disease: A Two-Sample Mendelian Randomization Study
Li H, Su J, Zhu M, Zhu Y, Zhu S
Clinical, Cosmetic and Investigational Dermatology 2023, 16:3291-3294
Published Date: 14 November 2023
Network Analysis of Osteoarthritis Progression Using a Steiner Minimal Tree Algorithm
Xie Y, Shao F, Ji Y, Feng D, Wang L, Huang Z, Wu S, Sun F, Jiang H, Miyamoto A, Wang H, Zhang C
Journal of Inflammation Research 2024, 17:3201-3209
Published Date: 18 May 2024
Single-Cell Sequencing Combined with Transcriptome Sequencing to Explore the Molecular Mechanisms Related to Psoriasis
E C, Wang R, Meng Z, Zou Y
Clinical, Cosmetic and Investigational Dermatology 2024, 17:2197-2213
Published Date: 3 October 2024