Back to Journals » Journal of Multidisciplinary Healthcare » Volume 18
Metabolism-Mediated FGF5 Association with Stroke: Based on Mendelian Randomization and Bioinformatics Analysis
Authors Xu C, Xu Y, Gao L, Wang M, Wang G, Wang G ![]()
Received 12 April 2025
Accepted for publication 18 July 2025
Published 31 July 2025 Volume 2025:18 Pages 4481—4495
DOI https://doi.org/10.2147/JMDH.S529168
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Krzysztof Laudanski
Cong Xu,1 Yonghong Xu,2 Ling Gao,3 Min Wang,1 Guangyan Wang,3 Guangming Wang1,4
1School of Clinical Medicine, Dali University, Dali, Yunnan, 671000, People’s Republic of China; 2Department of General Surgery, Banan Hospital Affiliated to Chongqing Medical University, Banan, Chongqing, 401320, People’s Republic of China; 3Department of Clinical Laboratory, Chuxiong Yi Autonomous Prefecture People’s Hospital, Chuxiong, 675000, People’s Republic of China; 4Center of Genetic Testing, the First Affiliated Hospital of Dali University, Dali, 671000, People’s Republic of China
Correspondence: Guangming Wang, Email [email protected]
Background: Stroke is the second leading cause of death and the third leading cause of disability worldwide. The role of fibroblast growth factor 5 (FGF5) in the occurrence and development of stroke remains unclear. We used bidirectional Mendelian randomization (MR) analysis to evaluate the mediating role of metabolites and causal association between inflammatory factors and stroke.
Methods: We analyzed the stroke dataset from the FinnGen database (v11) (cases: 43,132; Control: 297,867). Data on metabolites and inflammatory factors were obtained from the genome-wide Association Studies (GWAS) catalog of the European Institute for Bioinformatics (EBI). Using expression data of FGF5 mRNA and protein in the Comprehensive Gene Expression Database (GEO) and clinical data, expression level and clinical relevance of FGF5 in stroke were explored. The protein-protein interaction (PPI) network of FGF5-related genes was constructed, and various bioinformatics analyses (including functional enrichment, immune infiltration analysis, etc) were conducted to evaluate its functional mechanism.
Results: FGF5 was significantly associated with stroke risk (inverse variance weighting method (IVW): odds ratio (OR) = 1.052, 95% confidence interval (CI): 1.021– 1.084, P< 0.01). Mediation analysis indicated that inflammatory factors influenced stroke risk through the metabolites 1-palmitoyl-phosphoglycerol (GPG) [effect: 0.00462 (− 0.0102, 0.001); mediated effect: 9.09% (− 20.2%, 1.97%)], 1-stearoyl-2-arachidonoyl-phosphoethanolamine (GPE) [effect: 0.00274 (− 0.00212, 0.0076); mediated effect: 5.39% (4.17%, 14.9%). Among them, the mediating effect of 1-palmitoyl phosphatidylglycerol (GPG) was not significant. Furthermore, FGF5 is associated with epithelial cell proliferation, peptidyl-tyrosine phosphorylation, CD4+ primary T cells and M0 macrophages.
Conclusion: This study, by integrating multiple omics methods, such as Mendelian randomization, expression profiling analysis, and bioinformatics, has for the first time established FGF5 as a novel potential biomarker for stroke risk. Inflammatory factors can mediate the molecular pathways of stroke occurrence through metabolites such as GPE. The value of FGF5 as a novel biomarker for the diagnosis/prognosis of stroke and the new mechanism of stroke-related metabolic regulatory network provide a theoretical basis for targeted intervention of stroke.
Keywords: inflammatory factors, stroke, Mendelian randomization, mediation analysis, metabolite
Introduction
Stroke is the second most common cause of death globally among non-communicable diseases (NCDs) and ranks third as a combined cause of mortality and disability, as reflected by the loss of disability-adjusted life years (DALY).1 With the improvement of secondary stroke prevention strategies, the incidence of recurrent stroke and transient ischemic attack has decreased in recent years.2 Stroke incidence is rising in younger populations, emphasizing the need for lower diagnostic thresholds and specialized testing.3 Stroke is defined as a sudden neurological deficit caused by acute focal injury of the central nervous system owing to vascular reasons, including ischemic stroke (approximately 85%), cerebral hemorrhage (approximately 10%), and subarachnoid hemorrhage (approximately 5%). Ischemic stroke is heterogeneous, including subtypes such as large artery atherosclerosis, cardiac embolism, and small vessel occlusion, and the subtypes have different pathophysiological mechanisms and clinical significance. The clinical severity range is very wide, from transient symptoms (TIA) to devastating disability or death, and the presence of complications such as affected brain regions, infarction area, and brain edema or hemorrhagic transformation has significant impact.4–6 Time is crucial for the treatment of stroke. Neuronal death progresses rapidly within minutes to hours after occlusion. The hyperacute stage (<6 h) is mainly characterized by energy exhaustion, excitotoxicity and peri-infarct depolarization. The following days included complex inflammatory cascades and the formation of edema. From the subacute to the chronic stage, it often develops into persistent neurological deficits that require rehabilitation.7–9 In this context, the development of early detection strategies using more accurate biomarkers represents a critical frontier, as it would be the most cost-effective and reliable method for aiding timely stroke intervention.
The pathophysiology of stroke provides key insights into potential biomarker candidates. Ischemic injury triggers microglia and glial cell activation, leading to neuroinflammation, cell death, and pro-inflammatory cytokine and chemokine release.10 Post-stroke immune cells can either exacerbate inflammation or inhibit the inflammatory cascade to support healing.11 Cerebral ischemia induces robust inflammatory and immune activation in the brain and its vasculature. Given the central role of this inflammation in stroke pathophysiology, targeting key inflammatory mediators offers significant potential for improving stroke diagnosis and treatment outcomes.12,13
Beyond direct inflammatory signals, the downstream metabolic alterations induced by ischemia and inflammation offer another rich source of potential biomarkers. Metabolomics, the study of small-molecule metabolites, has revealed significant associations between specific metabolic profiles and stroke. Previous Mendelian research identified 14 unknown and 15 known serum metabolites (eight hazards and seven safeguards) linked to lacunar stroke.14 For example, glycogen phosphorylase isoenzyme BB and the apolipoprotein A1-unique peptide differ between stroke and control groups.15,16 These metabolic alterations are not merely bystanders; metabolic imbalances can significantly alter metabolite expression and circulating levels, actively contributing to the emergence of aberrant molecules that may reflect or influence disease progression. To establish the causal role of these metabolites and inflammatory factors in stroke pathogenesis, it is essential to overcome the limitations inherent in observational epidemiology17 To robustly establish the causal involvement of these metabolites and inflammatory factors in stroke pathogenesis, overcoming limitations of observational epidemiology is essential. Mendelian randomization (MR) analysis, leveraging human genetic data such as genome-wide association studies (GWAS), provides a powerful tool for this purpose.18 MR uses genetic variants, primarily single nucleotide polymorphisms (SNPs), as instrumental variables to assess causal relationships between exposures and outcomes (like stroke), mitigating confounding and reverse causation concerns inherent in traditional studies.
Building upon this rationale, our study employed a multi-faceted approach. We first utilized MR analysis to explore genetic variations robustly linked to ischemic stroke incidence and specifically investigated the potential mediating roles of inflammatory factors and metabolites in this relationship through mediation analysis. This genetic epidemiology approach helps address some limitations of conventional studies by utilizing the random assortment of genetic variants during meiosis. To translate these genetic associations into biological and clinical relevance, we focused on a promising candidate emerging from our analyses: Fibroblast Growth Factor 5 (FGF5).We used the Gene Expression Omnibus (GEO) database to investigate fibroblast growth factor 5 (FGF5) expression and its correlations with clinical features. We analyzed FGF5 expression across different stroke subtypes, constructed a protein–protein interaction (PPI) network using FGF5 and its associated genes, and conducted gene ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway, immunoinfiltration, and drug susceptibility analyses. The predictive role of FGF5 in cerebral infarction development was also evaluated. This research uncovers new opportunities for early stroke detection and treatment by highlighting inflammatory variable contributions to disease onset.8
The aim of this integrated research strategy, spanning genetic causality (via MR), molecular expression profiling, functional network analysis (PPI, GO/KEGG), immune microenvironment assessment, and clinical prediction is to fill a critical gap: the lack of a comprehensive, multi-omics understanding of FGF5’s causal role, functional mechanisms, and translational potential in stroke pathogenesis and early detection.
Materials And Methods
Study Design
To assess the indirect impact of inflammatory factors on the risk of stroke through potential mediators (such as metabolites), we adopted a two-step Mendelian randomization method. A two-sample magnetic resonance investigation was adopted. The first stage included evaluating the pathogenic effects of metabolites and inflammatory variables on stroke and screening genetic characteristics highly correlated with the risk of stroke. The subsequent stage involves evaluating the impact of the selected inflammatory factors on the characteristics of the metabolites examined and determining the intermediate effect of each factor on the mediator.19 Subsequently, we conducted sensitivity, pleiotropy and heterogeneity analyses to ensure reliability and effectiveness.20
Data Sources
Data were collected from the FinnGen database (v11), which includes 297,867 controls and 43,132 patients of European descent. The inflammatory factor dataset was obtained from the European Bioinformatics Institute genome-wide association study (EBI GWAS; https://ftp.ebi.ac/UK/pub/databases/gwas_summary_statistics/gwas) Catalog (accession numbers GCST90274758–GCST90274848) and the Department of Public Health and Primary Care (https://www.phpc.cam.ac.uk/ceu/proteins), with the former containing data on 1,400 metabolites.21
Genetic Instrumental Variable Selection
We adjusted the linkage disequilibrium parameters for SNPs strongly associated with exposure to r²<0.001 and kb>1000.22 During reverse MR, we applied a P<1e-05 threshold to filter inflammatory components with adverse effects.23,24 We used stringent criteria (r²<0.1, kb>500) to address SNP contribution imbalances, ensuring independent instrumental variables. We screened SNPs using an F statistic critical value of 10.25–27
Statistical Analysis
Mendelian Randomization
Statistical analyses were conducted using R (v4.3.0; The R Foundation for Statistical Computing, Vienna, Austria). Two-sample MR was performed using the “TwoSampleMR”, “VariantAnnotation”, and “ieugwasr” R packages. Five analytical methods were employed: weighted median (WM), MR–Egger, inverse variance weighted (IVW), simple mode, and weighted mode.28 The IVW method was prioritized as the primary analysis due to its higher statistical power and consistency under the assumption of valid instruments (no horizontal pleiotropy).29 In MR analysis, IVW is considered a reliable and accurate method for estimating causal relationships.30 A statistically significant causal association between outcomes and exposure was defined by P<0.05. Odds ratio (OR) and 95% confidence interval (CI) estimates indicated that a positive beta (effect size) signified increased exposure factors, potentially enhancing stroke risk.
Sensitivity Analyses
Cochran’s Q statistic quantified heterogeneity.Significant heterogeneity (Cochran’s Q P-value < 0.05) suggests potential violations of MR assumptions, warranting further investigation using alternative methods and sensitivity analyses.31 MR-PRESSO technique, which identifies and corrects for outliers potentially indicative of horizontal pleiotropy, and the MR--Egger intercept test. A non-zero MR-Egger intercept (P < 0.05) provides evidence for directional pleiotropy.32 To test result stability, the “leave-one-out” method was applied, removing each SNP individually to assess the impact of each remaining SNP. The “product of coefficients” method was used to evaluate the indirect effects of inflammatory factors on stroke risk through potential mediators, and the delta method ascertained standard errors for indirect effects.27
Transcriptome Analysis
Data Set Filtering
Stroke microarray data in the MINiML format were downloaded from the GEO database (www.ncbi.nlm.nih.gov/geo/), applying the following criteria: a) sequenced using the same platform to generate expression profiles across two different spectra; b) human test samples; and c) >20 samples in each group. We included the datasets GSE22255 (40 samples, 20 of which were normal) and GSE37587 (68 samples).
Stroke Subtype Classification
Consensus clustering is widely used to classify disease subtypes. Consistency analysis was performed using the ConsensusClusterPlus R package (v1.54.0), with maximum cluster number=6. The clustering process was repeated 100 times, with 80% of the sample randomly selected in each iteration. The parameters clusterAlg=“hc” and innerLinkage=“ward.D2” were applied to optimize hierarchical clustering.
For clustering heatmap analysis, the pheatmap R package (v1.0.12) was used. Only genes with variance>0.1 were retained for the heatmap. Statistical analysis was conducted using R (v4.0.3). P<0.05 was considered statistically significant.
PPI Network Construction and Gene Pathways and Function Analysis
We used the CoreMine Medical database (https://coremine.com/medical) to investigate cerebral infarction, focusing on the eight initial genes associated with FGF5. We then explored the GO functions and KEGG pathway enrichment of FGF5 in the stroke context using the Metascape platform (https://metascape.org). The biological processes (GOTERM_BP_DIRECT), cellular components (GOTERM_CC_DIRECT), and molecular functions (GOTERM_MF_DIRECT) results were grouped under GO, whereas KEGG pathways were documented separately.
Immune Cell Infiltration Evaluation
The “CIBERSORT” software package was employed to analyze immune cell infiltration and the proportions of 22 immune cell types in the stroke and control groups. The immune cell proportions in each sample were represented in a bar chart, and a boxplot compared immune cell percentages between groups. Pearson correlation analysis assessed relationships between immune cell proportions and candidate diagnostic gene expression. The results were visualized using the “ggplot2” package.
Drug Sensitivity Analysis
The Coremine Medical database (https://coremine.com/medical) was used for drug sensitivity analysis, and drugs significantly associated with FGF5 and stroke (P<0.05) were identified. This analysis aimed to identify potential therapeutic agents targeting the FGF5 pathway in stroke. And we used the Comparative Toxicomics Database (CTD, https://ctdbase.org/) to verify these key drugs that were significantly associated with FGF5 in the context of cardiovascular disease, with particular focus on stroke.
Metabolic Pathways and Enrichment Analysis
We used the MetaboAnalyst 6.0 platform (http://dev.metaboanalyst.ca/) to investigate the selected metabolites and conducted metabolic pathway enrichment analysis using the KEGG database to identify metabolite groups or pathways potentially associated with cerebral infarction development and progression. Metabolites that yielded significant IVW results (P<0.05) were emphasized. Metabolites that yielded significant IVW results (P<0.05) in the initial MR analysis relating metabolites to stroke risk were emphasized. Pathways with a Holm-Bonferroni adjusted P-value < 0.05 were considered significantly enriched.
Results
This study aims to explore the causal relationship between inflammatory factors and their related metabolites and the risk of stroke through the combination of Mendelian randomization (MR) and bioinformatics analysis. The study further explored the potential mechanism of the core inflammatory factor Fibroblast growth factor 5 (FGF5) through transcriptomics and immune infiltration analysis.
Bidirectional Mendelian Risk Analysis of Inflammatory Factors for Stroke
Three inflammatory factors were strongly associated with stroke (Table 1). IL-6 may exert protective effects against stroke and reduce stroke risk.20 In contrast, Matrix metalloproteinase-1 (MMP-1) and FGF-5 are correlated with increased stroke risk (Figure 1A–F).
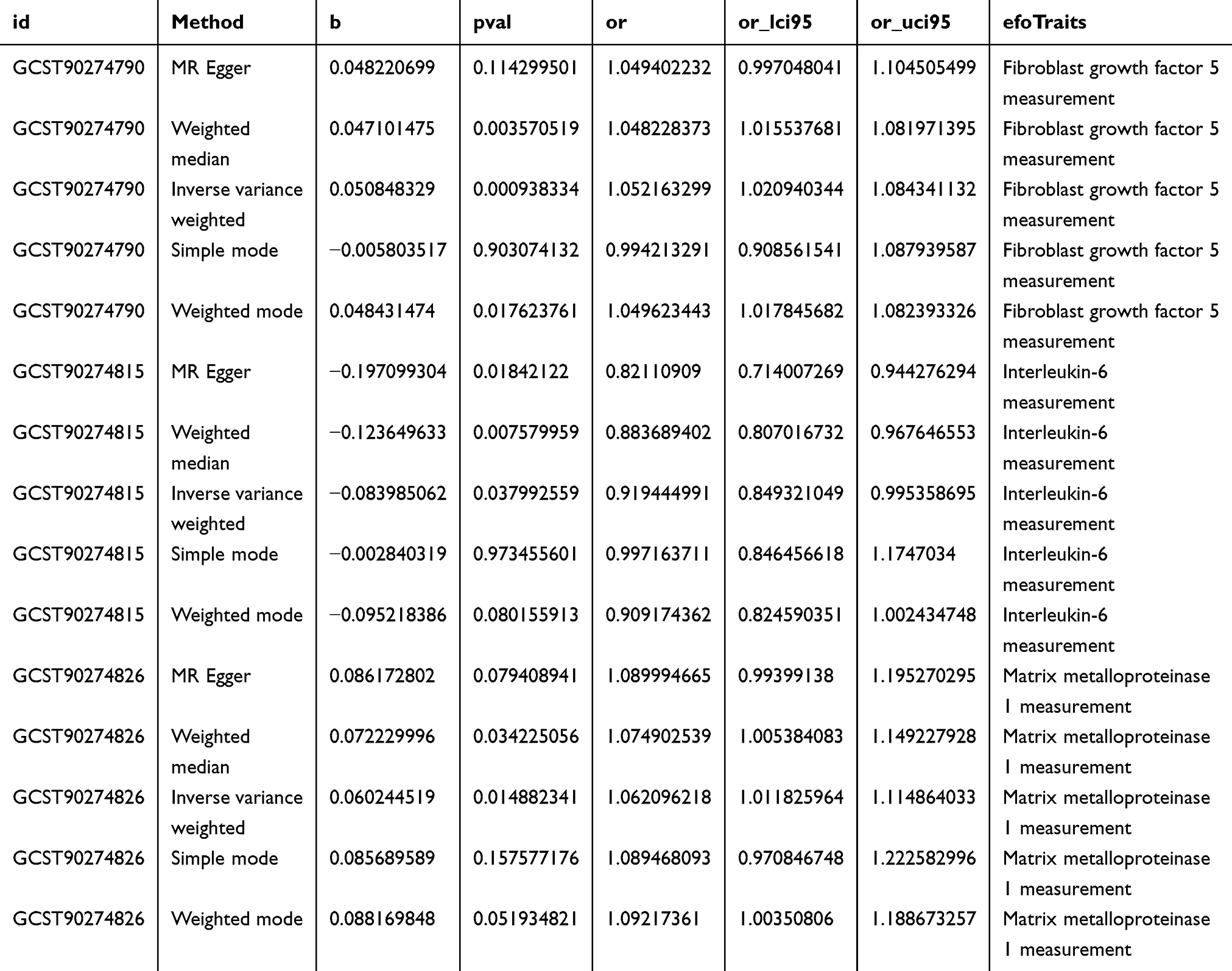 |
Table 1 A Strong Link Was Observed Between Three Key Inflammatory Indicators and Stroke Rates |
 |
Figure 1 Mendelian randomization of inflammatory factors. (A–C): Scatter plots of strong association of inflammatory factors with stroke. (D–F): Forest map of strong association of inflammatory factors with stroke. (G–I): Funnel plot for sensitivity analysis of strong association of inflammatory factors with stroke. (J–L): Four types of gut microbiota closely associated with stroke were mapped using the “leave-one-out” method. (M). Forest map of inflammatory factors. Forest maps show a causal relationship between inflammatory factors significantly associated with stroke; the analytical methods were inverse variance weighting, Mendelian randomization, Egger, weighted median, simple mode, and weighted mode. |
The sensitivity analysis revealed no bias in the MR analysis; neither heterogeneity nor horizontal pleiotropy was detected using Cochran’s Q, the MR–Egger intercept, or the MR-PRESSO method (Figure 1G–I). The SNP “leave-one-out” analysis showed no changes in the total error, confirming data reliability (Figure 1J–L).21,33
No reverse causal relationship was evident between stroke and FGF-5 expression: (IVW; OR=1.016, 95% CI: 0.952–1.085, P=0.63), consistent with the IL-6 (IVW; OR=1.039, 95% CI: 0.980–1.101, P=0.20) and matrix metalloproteinase (MMP)-1 (IVW: OR=0.999, 95% CI: 0.939–1.062, P=0.97; Figure 1M) results.
Inflammatory Factor and Metabolite Risk Analysis
We assessed the potential stroke risk associated with 1,400 metabolites using the IVW technique, in conjunction with MR–Egger, WM, and simple and weighted mode analyses. Thirty metabolites were linked to stroke risk, 20 positively and 10 negatively. After excluding redundant data, all variables remained statistically significant. Cochran’s Q-test indicated no heterogeneity in the data.34 The “leave-one-out” analysis showed that the causality estimate was unaffected by any single SNP removal.
Investigating the causal relationship between stroke risk and metabolites revealed that only two inflammatory factors—FGF-5 (IVW; OR=1.052, 95% CI: 1.021–1.084, P<0.01) and MMP-1 (IVW; OR=1.062, 95% CI: 1.012–1.115, P=0.02; Figure 2A–I)—were strongly associated with stroke risk in the Mendelian analysis. No heterogeneity or pleiotropy was evident, and no specific SNPs showed causal effects (Figure 2I–P).
 |
Figure 2 Mendelian randomization analysis of inflammatory factors and metabolites. (A–C): Scatter plots of the association of fibroblast growth factor 5 (FGF5) with 1-palmitoyl-glycero-3-phosphoglycerol (GPG), 1-stearoyl-2-arachidonoyl-glycero-3-phosphoethanolamine (GPE), and X-16087. (D–F): Forest map of strong association of FGF5 with 1-palmitoyl-GPG, 1-stearoyl-2-arachidonoyl-GPE, and X-16087. (G–I): Funnel plot for sensitivity analysis of strong association of FGF5 with 1-palmitoyl-GPG, 1-stearoyl-2-arachidonoyl-GPE, and X-16087. (J–L): Strong associations of FGF5 with 1-palmitoyl-GPG, 1-stearoyl-2-arachidonoyl-GPE, and X-16087 were mapped using the “leave-one-out” method. Abbreviations: CI, confidence interval; MR, Mendelian randomization; OR, odds ratio; SNP, single nucleotide polymorphism. |
Mediation by Metabolites in Causal Stroke–Inflammatory Factor Relationships
The two-step MR analysis identified three metabolites that mediate causal inflammatory factor–stroke relationships: 1-palmitoyl-phosphoglycerol (GPG) [effect: 0.00462 (−0.0102, 0.001); mediated effect: 9.09% (−20.2%, 1.97%)], 1-stearoyl-2-arachidonoyl-phosphoethanolamine (GPE) [effect: 0.00274 (−0.00212, 0.0076); mediated effect: 5.39% (4.17%, 14.9%)], and the unidentified metabolite X-16087 [effect: 0.00369 (−0.00118, 0.00856); mediated effect: 7.26% (2.33%, 16.8%)]. Therefore, FGF-5 modulates stroke risk by altering 1-palmitoyl-GPG, 1-stearoyl-2-arachidonoyl-GPE, and X-16087 expression.
Overall Effect of Inflammatory Factors on Stroke
FGF-5 gene expression showed a strong positive association with stroke risk (IVW; OR=1.052, 95% CI: 1.021–1.084, P<0.01), suggesting it as a potential risk factor. This association implies that tissue damage during stroke may result from an inflammatory process, consistent with the weighted mode (OR=1.050, 95% CI: 1.018–1.082, P = 0.02) and WM results (OR=1.048, 95% CI: 1.016–1.082, P<0.01; Figure 3A, B, E).
 |
Figure 3 Total effects between inflammatory factors and stroke with Mendelian randomization analysis. (A): Scatter plots of strong association of fibroblast growth factor 5 (FGF5) with stroke. (B): Forest map of strong association of FGF5 with stroke. (C): Funnel plot for sensitivity analysis of strong association of FGF5 with stroke. (D): Mapping of the strong association of FGF5 with stroke using the “leave-one-out” method. (E). FGF5 shows reverse causality with the total effects of stroke. 1-palmitoyl-GPG, 1-stearoyl-2-arachidonoyl-GPE, and X-16087 are positively correlated with the incidence of stroke. FGF5 has a positive causal relationship with 1-palmitoyl-GPG and 1-stearoyl-2-arachidonoyl-GPE. FGF5 has a negative causal relationship with X-16087. Abbreviations: CI, confidence interval; MR, Mendelian randomization; OR, odds ratio; SNP, single nucleotide polymorphism. |
Cochran’s Q-test revealed no significant heterogeneity, and MR–Egger analysis indicated no horizontal pleiotropy. Although some outlier SNPs were detected using the MR-PRESSO method, their presence did not affect the stability of the main findings. The instrumental variable distribution showed clear symmetry. The robustness of our results was further validated through the “leave-one-out” analysis, confirming the absence of outlier SNPs.(Figure 3C and D).35
Transcriptomic Analysis
FGF5 and Stroke Subtype Classification
To further elucidate the significance of FGF5 as a biomarker for the subtype classification of stroke, we have uncovered the biological heterogeneity of distinct stroke subtypes (C1/C2) via molecular profiling and demonstrated the pivotal role of FGF5 in this classification.
At k=2, the ConsensusClusterPlus consistent clustering results were visualized through a heatmap, where both axes represent samples and colors represent different categories. The heatmap illustrates FGF5 expression levels across the identified subgroups, with red and blue indicating high and low expressions, respectively. The differential FGF5 expression between clusters C1 and C2 was notable A significant difference in gene expression was observed when comparing C1 and C2 in the four-gene clustering heatmap. In additional heatmaps highlighting FGF5-associated gene expression in these subgroups, red and blue represent high and low expression, respectively (Figure 4A–C).
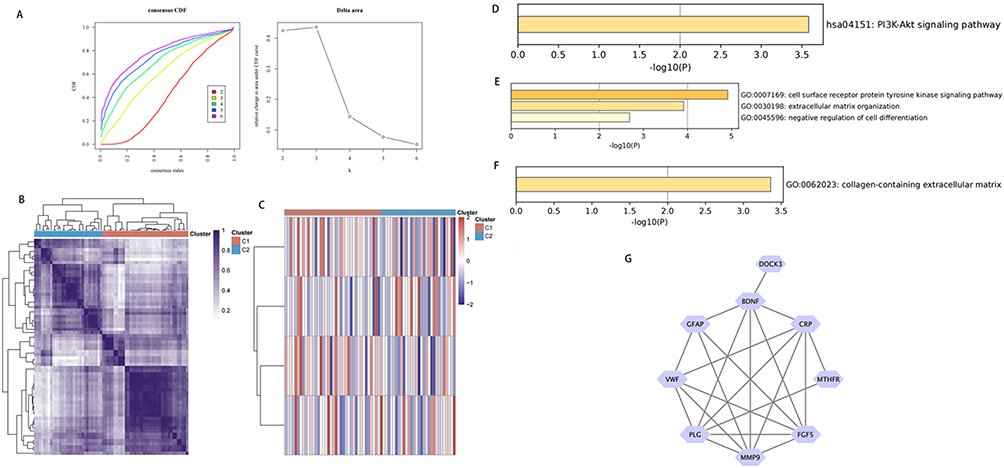 |
Figure 4 (A–C): Microarray subtype based on cerebral infarction associated fibroblast growth factor 5 (FGF5). (A) CDF curve and delta area of CDF curve. (B) Consistency of heat maps of clustering results when K=2. (C) Heat maps of FGF5 and its related genes in different subpopulations; Red indicates high expression and blue indicates low expression. CDF, cumulative distribution function. (D–G): GO and KEGG enrichment analysis. (D) BP enrichment analysis. (E) CC enrichment analysis. (F) KEGG enrichment analysis. (G) PPI network diagram. Abbreviations: GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; PPI, protein-protein interaction. |
Functional Enrichment Analysis and PPI Network
To elucidate the biological mechanisms of FGF5 and its associated genes in stroke pathology, we constructed a molecular network and conducted a comprehensive functional analysis. Our findings position FGF5 as a multifunctional node within the stroke pathological network.
Through the Core Mine website (https://coremine.com/medical/), we identified strongly FGF5-related genes in the stroke context, including C-reactive protein (CRP), cytokinesis protein 3 (DOCK3), plasminogen (PLG), brain-derived neurotrophic factor (BDNF), methylenetetrahydrofolate reductase (MTHFR), Von Willebrand factor (VWF), MMP9, and glial fibrillary acidic protein (GFAP). To explore the biological roles of FGF5 and these genes, we performed GO and KEGG functional enrichment analyses. In the biological process category, these genes were enriched in the cell surface receptor protein tyrosine kinase signaling pathway and extracellular matrix organization in negative cell differentiation regulation. In the cell component category, these genes were found in collagen-containing extracellular matrix. KEGG pathway analysis revealed significant PI3K-Akt signaling, EGFR tyrosine kinase inhibitor resistance, central carbon metabolism in cancer, melanoma, and proteoglycans in cancer pathway involvement (Figure 4D–G).
Different Immune Infiltration Levels in Stroke Patients with Diverse Risk Profiles
The reprogramming effect of FGF5 expression levels on the immune microenvironment in stroke has been validated. FGF5 modulates the immunosuppressive microenvironment of stroke through the regulation of immune cells and factors, thereby providing a foundational basis for immunotargeted therapeutic strategies.
Immune infiltration pattern analysis in stroke patients and healthy controls (Figure 5A) showed significant increase in M0 and M1 macrophage in stroke patients, whereas CD4+ naïve T cells were notably reduced in the control group (Figure 5B). Moreover, neutrophil levels were elevated in the high-risk group, suggesting immunosuppression, possibly linked to immune checkpoint expression. In our hepatocellular carcinoma analysis, we found significant differences in M1 and M2 macrophage expression between high- and low-risk subgroups. Significant differences in the proportions of three immune cell types between the experimental and control groups were revealed (Figure 5C). Specifically, FGF5 expression correlated positively with CD4+ naïve T cells and M0 macrophages, but negatively with CD8+ T cells (Figure 5D).
 |
Figure 5 Analysis of immune cell infiltration: (A) Comparison of types and proportions of immune cells between the stroke and normal groups; (B) Differences in immune cell infiltration between the stroke and the normal groups; (C) Lollipop plots illustrating fibroblast growth factor 5 and its associated genes in the context of immune cell infiltration in stroke; (D) Principal components analysis scores of the normal and stroke groups. |
Drug Sensitivity Analysis in Relation to FGF5 in Stroke
In order to transform the discovery of FGF-5 into therapeutic insights, we conducted a drug sensitivity analysis. Correlation analysis was conducted using the Coremine Medical database (https://coremine.com/medical) to study potential correlations between FGF5, stroke, and drug sensitivity. The top 10 significantly FGF5-associated drugs in stroke treatment were tissue plasminogen activator, warfarin, aspirin, salicylic acid, menogaril, clotrimazole, cholesterol, arginine/omega-3 fatty acids/nucleotides oral supplement, tetradecanoylphorbol acetate, and heparin (Table 2).
 |
Table 2 Drugs Significantly Associated with Fibroblast Growth Factor 5 Action in Stroke |
Among them, the verified drugs are Tissue Plasminogen Activator,36 Warfarin,37,38 Aspirin, Salicylic Acid,39,40 Heparin.41,42 However, for several other drugs, including arginine, clotrimazole, Menogarre and tetradecanyl phosphorus acetate, there is currently a lack of experimental evidence to support them in FGF5-related stroke models. Nevertheless, these drugs still show value as potential targets for future research.
Metabolite Enrichment Analysis
Functional enrichment analysis using known metabolites via the IVW method (PIVW<0.05) identified six significant metabolic pathways with key roles in cerebral infarction initiation and development, including glycerophospholipid, linoleic acid, glycerophospholipid, alpha-linolenic acid, and arachidonic acid metabolism, and glycosylphosphatidylinositol (GPI)-anchor biosynthesis (Table 3).
 |
Table 3 Metabolic Pathway Enrichment Analysis of Stroke-Associated Metabolites |
Discussion
The FGF family includes key mitotic factors for blood vessel formation during angiogenesis,43 with FGF-5 being the third member of this family.44 Notably, systolic and diastolic blood pressure shows a positive correlation (P<0.05) with FGF-5 protein expression.45 FGF-5 overexpression exacerbates oxidative stress, inflammation, and spinal cord injury by activating AMP-activated protein kinase signaling in a cAMP/protein kinase A-dependent manner.46 FGF5 is mainly described as a factor that plays a key role in the cardiovascular system. It significantly promotes cardiac remodeling by directly inducing myocardial hypertrophy and serving as a paracrine signal induced by mechanical stress. The continuously elevated level of circulating FGF5 is significantly associated with hypertensive cardiac hypertrophy and may promote the occurrence and development of heart failure. Regarding the direct impact of FGF5 on cerebrovascular diseases, the current material data is limited. However, studies on other members of the FGF family (such as FGF-21, FGF-23 and FGF-2) have shown that this family may play a role in stroke and neuropsychiatric sequelae, and the specific role of FGF5 remains to be further studied and confirmed.47,48
This aligns with our observation that FGF-5 contributes to increased stroke risk, suggesting that FGF-5 influences various blood vessel- and neural tissue-associated disorders.
FGF-5-associated genes, including CRP, DOCK3, PLG, BDNF, MTHFR, VWF, MMP9, and GFAP, were identified. FGF-5 and related genes were significantly enriched in several pathways, including PI3K-Akt signaling, cell surface receptor protein tyrosine kinase signaling, and extracellular matrix organization in negative cell differentiation regulation. Enrichment of stroke-associated proteins was observed in glycerophospholipid, linoleic acid, alpha-linolenic acid, and arachidonic acid metabolism, and in glycosylphosphatidylinositol (GPI)-anchor biosynthesis.
Drug sensitivity analysis revealed patient responses to various medications, including tissue plasminogen activator, warfarin, aspirin, salicylic acid, menogaril, clotrimazole, and cholesterol-lowering agents. Significant drug sensitivity was observed for arginine/omega-3 fatty acids/nucleotides oral supplement, tetradecanoylphorbol acetate, and heparin.
M0 and M1 macrophages were significantly elevated in stroke patients. CD4 naive T cells and M0 macrophages showed significant positive associations, whereas CD8 T cells demonstrated an inverse relationship with FGF5 expression. In addition to the direct association between FGF5 and immune cells, recent studies have also found that immune receptors and neuroregulatory genes may further affect cerebrovascular homeostasis through the immune-neural axis mediated by metabolites. Some scholars have studied that RNF8 regulates the degradation of galectin-3 (gal-3) through K48-polyubiquitination, affecting T-cell infiltration in the tumor microenvironment and the therapeutic sensitivity of PD-L1. ATAD5 affects B-cell class transition recombination (CSR) by regulating the unloading of proliferating cell nuclear antigens. Its haploid deficiency leads to a decrease in the expression of activation-induced deaminase (AID) and reduces IgH locus breaks (in the immune direction). RELN, as a key gene for cerebellar development, its mutation leads to motor coordination disorders (such as spinocerebellar ataxia) (neural direction). In addition, CACNA1A encodes voltage-gated calcium channel subunits. Mutations are associated with epileptic encephalopathy (DEE-SWAS) and ataxia (neural direction), and may also affect immune-neural interactions through calcium signaling. It indicates that the metabolite X-16087 may regulate cerebrovascular homeostasis through the immune-neural axis. This result supports our core hypothesis - that FGF5 may affect the occurrence of stroke through the interaction of the metabolism-immune-neural multi-pathway through the mediator molecule X-16087. Previous studies highlighted the importance of FGF5 and T cells in the development of various diseases. C2 cytotoxic T lymphocytes (CTLs), derived from human CTLs infiltrating renal cell carcinoma, target and destroy cancer cells and certain FGF5-overexpressing prostate and breast cancer cell lines.49 In our study, C2 cells recognized the human leukocyte antigen A3 major histocompatibility complex class I molecule, which presents nine FGF5-derived peptides via protein splicing. This highlights vertebrate proteome complexity and the immune system’s capacity to identify endogenous and exogenous peptides.50 In vitro, FGF5 promotes the proliferation, invasion, and migration of lung cancer cells, with its expression predominantly localized to the nucleolar fibrillar center.51 In the brain, atherosclerotic plaques often exhibit immunoreactive acidic FGF (aFGF), particularly in areas of new blood vessel formation within the plaque and in macrophage-rich regions. aFGF released by plaque macrophages may promote microvessel development as atherosclerosis progresses in humans.52
Recent developments in studies on other brain disorders may be promising for future stroke treatment, although fibroblast research is not sufficiently advanced to target stroke therapeutics. For instance, neurofibromas can progress into malignancies without exhibiting elevated mitotic activity. To incorporate auxiliary research, we first emphasize the role of single-cell analysis. Genetic testing alone cannot reliably identify malignancy in atypical tumors. However, these techniques may provide some utility, particularly in histological evaluations when SOX10 (nuclear) and S100 protein (cytoplasmic and nuclear) loss is evident, and CD34-positive fibroblast network absence is observed.53
Biomarkers of neuronal damage and membrane integrity are widely utilized to study metabolic alterations in the cerebral cortex following stroke.54 Recombinant FGFs have shown promise for stroke treatment in animal models. FGFs regulate metabolism-associated cells and hormones, serving as key homeostatic factors. After stroke, FGFs influence pro-inflammatory (M1) and anti-inflammatory (M2) cytokine release. Although energy substrate deficiency is commonly considered the primary cause of ischemic damage, a recent study suggested that metabolite signaling may influence stroke pathogenesis.33 In addition to regulating substrates transport across the plasma membrane through kinases and transporters, palmitoylation is crucial in controlling mitochondrial metabolism. Palmitoylation dysregulation may contribute to hypoxic damage and metabolic disorders. The role of palmitoylation in regulating subcellular substrate transport has been extensively studied in adipocytes and liver cells but not well understood in the bone and myocardium.55,56
The endocannabinoids arachidonic glycerol and arachidonic glycolamide have been implicated in physiological conditions, including inflammation, pain, neurological disorders, obesity, metabolic syndrome, and liver disease. They possess immunomodulatory properties and contain arachidonic acid, a precursor of numerous bioactive lipids that exhibit both pro-inflammatory and anti-inflammatory effects.57 The metabolite 1-arachidonic acyl-GPE is a potential mediator in atopic dermatitis.58 Its expression is strongly correlated with smoking status and smoker body mass index.59 Stearoyl-arachidonoyl-2-arachidonoyl-GPE is closely associated with 25-hydroxyvitamin D activity, which is important in the pathogenesis of cancer, cardiovascular disease, and bone health. A strong positive association between 1-stearoyl-2-arachidonoyl-GPE and stroke risk was reported.22 However, the impact of 1-palmitoyl-GPG and 1-stearoyl-2-arachidonoyl-GPE on cardiovascular and cerebrovascular disorders remains underexplored. In addition, arachidenyl derivatives play a dual role in stroke. In the brain, they activate targets such as CB2 receptor, TRPV1 and PPARα through endogenous cannabinoids (such as anandamide/OEA) to enhance the blood-brain barrier (BBB) against ischemic injury and exert a protective effect. In the systemic blood vessels, abnormal accumulation of arachidenyl phospholipids (such as pPE38:4/38:5) mediated by CEPT1 enzyme in diabetic carotid artery plaques leads to the enrichment of arachidonic acid (AA), which aggravates plaque vulnerability through the inflammatory pathway and promotes embolic stroke. This contradictory mechanism highlights the necessity of tissue-specific targeted intervention.48 Future research should examine the mediating role of these metabolites in stroke and reassess the mec hanisms underlying stroke development and pathogenesis.
Our results demonstrated consistency, with no horizontal pleiotropy, strengthening the robustness of the findings. Sensitivity assessment further validated the objectivity and reliability of the analytical outcomes. An important limitation of this study is that the samples are mainly derived from the European population. There are significant differences between the European population and other ethnic groups in terms of the frequency of genetic polymorphisms, etc.48 In addition, the metabolite lineage is also influenced by dietary culture. These differences limit the general applicability of the findings of this study, especially among the populations in Asia and Africa. Future studies should incorporate multi-ethnic cohorts (such as the African American population in the UK Biobank and the Chinese CHARLS cohort) to verify whether the FGF-5- metabolite-stroke pathway has cross-ethnic conservation. Second, using stringent thresholds in our analyses may have reduced the accuracy of some findings. Last, age range and dietary characteristic data, which are not readily accessible from the original data source, are lacking. Consequently, the study outcomes may vary across demographic groups or exposure levels. Future research should consider stratifying populations by age and incorporating dietary data to enhance the generalizability of the findings.
Conclusion
Through MR analysis, a genetic link between metabolites, inflammatory markers, and stroke risk was demonstrated. FGF-5 may reduce stroke risk by modulating metabolite levels (1-palmitoyl-GPG and 1-stearoyl-2-arachidonoyl-GPE). These metabolic signaling pathways may be more important in stroke pathophysiology than traditionally studied inflammatory factors. These results offer novel insights into the mechanisms underlying stroke and highlight potential candidate biomarkers for targeted therapeutic interventions. Future research aimed at developing stroke treatments should explore the therapeutic potential of targeting these metabolic pathways, particularly in European populations.
Data Sharing Statement
All data generated or analyzed during this study are included in the published article.
Ethics Statement
The studies involving human participants were reviewed and approved by the Ethics Committee of the Affiliated Hospital of Dali University. The patients/participants provided their written informed consent to participate in this study. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article. Ethics review approval number is DFY20220415001.
Acknowledgments
We would like to acknowledge the key construction disciplines of The First Affiliated Hospital of Dali University.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study received funding from The National Natural Science Foundation of China [No: 82160244]; Scientific Research Fund project of Education Department of Yunnan Province, No:2025Y1140; Dali City Industrial Information and Technology Bureau 2024 science and technology plan project,No: 2024KBG145.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Feigin VL, Brainin M, Norrving B, et al. World stroke organization: global stroke fact sheet 2025. Int J Stroke Feb. 2025;20(2):132–144. doi:10.1177/17474930241308142
2. Mulder M, Cras TY, Shay J, Dippel DWJ, Burke JF. Comparison of American and European guideline recommendations for diagnostic workup and secondary prevention of ischemic stroke and transient ischemic attack. Circulation. 2024;150(10):806–815. doi:10.1161/CIRCULATIONAHA.124.069651
3. Feigin VL, Owolabi MO. Pragmatic solutions to reduce the global burden of stroke: a world stroke organization-lancet neurology commission. Lancet Neurol. 2023;22(12):1160–1206. doi:10.1016/S1474-4422(23)00277-6
4. Sacco RL, Kasner SE, Broderick JP, et al. An updated definition of stroke for the 21st century: a statement for healthcare professionals from the American heart association/American stroke association. Stroke. 2013;44(7):2064–2089. doi:10.1161/STR.0b013e318296aeca
5. Feigin VL, Norrving B, Mensah GA. Global burden of stroke. Circ Res. 2017;120(3):439–448. doi:10.1161/CIRCRESAHA.116.308413
6. Campbell BCV, De Silva DA, Macleod MR, et al. Ischaemic stroke. Nat Rev Dis Primers. 2019;5(1):70. doi:10.1038/s41572-019-0118-8
7. Powers WJ, Rabinstein AA, Ackerson T, et al. Guidelines for the early management of patients with acute ischemic stroke: 2019 update to the 2018 guidelines for the early management of acute ischemic stroke: a guideline for healthcare professionals from the American heart association/American stroke association. Stroke. 2019;50(12):e344–e418. doi:10.1161/STR.0000000000000211
8. Moskowitz MA, Lo EH, Iadecola C. The science of stroke: mechanisms in search of treatments. Neuron. 2010;67(2):181–198. doi:10.1016/j.neuron.2010.07.002
9. Jayaraj RL, Azimullah S, Beiram R, Jalal FY, Rosenberg GA. Neuroinflammation: friend and foe for ischemic stroke. J Neuroinflamm. 2019;16(1):142. doi:10.1186/s12974-019-1516-2
10. Liu P, Chen Y, Zhang Z, et al. Noncanonical contribution of microglial transcription factor NR4A1 to post-stroke recovery through TNF mRNA destabilization. PLoS Biol. 2023;21(7):e3002199. doi:10.1371/journal.pbio.3002199
11. Tariq MB, Lee J, McCullough LD. Sex differences in the inflammatory response to stroke. Semin Immunopathol. 2023;45(3):295–313. doi:10.1007/s00281-022-00969-x
12. Li Y, Liao J, Xiong L, et al. Stepwise targeted strategies for improving neurological function by inhibiting oxidative stress levels and inflammation following ischemic stroke. J Control Release. 2024;368:607–622. doi:10.1016/j.jconrel.2024.02.039
13. Lambertsen KL, Finsen B, Clausen BH. Post-stroke inflammation-target or tool for therapy? Acta Neuropathol. 2019;137(5):693–714. doi:10.1007/s00401-018-1930-z
14. Guo MN, Hao XY, Tian J, et al. Human blood metabolites and lacunar stroke: a Mendelian randomization study. Int J Stroke. 2023;18(1):109–116. doi:10.1177/17474930221140792
15. Park KY, Ay I, Avery R, et al. New biomarker for acute ischaemic stroke: plasma glycogen phosphorylase isoenzyme BB. J Neurol Neurosurg Psychiatry. 2018;89(4):404–409. doi:10.1136/jnnp-2017-316084
16. Zhao X, Yu Y, Xu W, et al. Apolipoprotein A1-unique peptide as a diagnostic biomarker for acute ischemic stroke. Int J Mol Sci. 2016;17(4):458. doi:10.3390/ijms17040458
17. Li W, Shao C, Li C, et al. Metabolomics: a useful tool for ischemic stroke research. J Pharm Anal. 2023;13(9):968–983. doi:10.1016/j.jpha.2023.05.015
18. Chen Y, Lu T, Pettersson-Kymmer U, et al. Genomic atlas of the plasma metabolome prioritizes metabolites implicated in human diseases. Nat Genet. 2023;55(1):44–53. doi:10.1038/s41588-022-01270-1
19. Carter AR, Sanderson E, Hammerton G, et al. Mendelian randomisation for mediation analysis: current methods and challenges for implementation. Eur J Epidemiol. 2021;36(5):465–478. doi:10.1007/s10654-021-00757-1
20. Zheng T, Liu C, Wang Y, et al. Inflammatory cytokines mediating the effect of oral lichen planus on oral cavity cancer risk: a univariable and multivariable Mendelian randomization study. BMC Oral Health. 2024;24(1):375. doi:10.1186/s12903-024-04104-0
21. Si S, Li J, Tewara MA, Xue F. Genetically determined chronic low-grade inflammation and hundreds of health outcomes in the UK biobank and the finngen population: a phenome-wide Mendelian randomization study. Front Immunol. 2021;12:720876. doi:10.3389/fimmu.2021.720876
22. Cheng Z, Ye F, Liang Y, et al. Blood lipids, lipid-regulatory medications, and risk of bladder cancer: a Mendelian randomization study. Front Nutr. 2023;10:992608. doi:10.3389/fnut.2023.992608
23. Wen B, Wei S, Huang D, et al. The connection between 91 inflammatory cytokines and frailty mediated by 1400 metabolites: an exploratory two-step Mendelian randomization analysis. Arch Gerontol Geriatr. 2025;133:105774. doi:10.1016/j.archger.2025.105774
24. Zhang YC, Fan KY, Wang Q, et al. Genetically determined levels of mTOR-dependent circulating proteins and risk of multiple sclerosis. Neurol Ther. 2023;12(3):751–762. doi:10.1007/s40120-023-00455-y
25. Klarin D, O’Donnell CJ, Kathiresan S. Mendelian randomization study of ACLY and cardiovascular disease. N Engl J Med. 2020;383(7):e50. doi:10.1056/NEJMc1908496
26. Guo J, Peng C, He Q, Li Y. Type 2 diabetes and the risk of synovitis-tenosynovitis: a two-sample Mendelian randomization study. Front Public Health. 2023;11:1142416. doi:10.3389/fpubh.2023.1142416
27. Meng C, Sun L, Shi J, et al. Exploring causal correlations between circulating levels of cytokines and colorectal cancer risk: a Mendelian randomization analysis. Int J Cancer. 2024;155(1):159–171. doi:10.1002/ijc.34891
28. Sun J, Wang M, Kan Z. Causal relationship between gut microbiota and polycystic ovary syndrome: a literature review and Mendelian randomization study. Front Endocrinol. 2024;15:1280983. doi:10.3389/fendo.2024.1280983
29. Burgess S, Dudbridge F, Thompson SG. Combining information on multiple instrumental variables in Mendelian randomization: comparison of allele score and summarized data methods. Stat Med. 2016;35(11):1880–1906. doi:10.1002/sim.6835
30. Brion MJ, Shakhbazov K, Visscher PM. Calculating statistical power in Mendelian randomization studies. Int J Epidemiol. 2013;42(5):1497–1501. doi:10.1093/ije/dyt179
31. Ren S, Xue C, Xu M, Li X. Mendelian randomization analysis reveals causal effects of polyunsaturated fatty acids on subtypes of diabetic retinopathy risk. Nutrients. 2023;15(19):4208. doi:10.3390/nu15194208
32. Bowden J, Davey Smith G, Haycock PC, Burgess S. Consistent estimation in Mendelian randomization with some invalid instruments using a weighted median estimator. Genet Epidemiol. 2016;40(4):304–14.
33. Fan L, Lei H, Lin Y, et al. Hotair promotes the migration and proliferation in ovarian cancer by miR-222-3p/CDK19 axis. Cell Mol Life Sci. 2022;79(5):254. doi:10.1007/s00018-022-04250-0
34. Zhao K, Zhao Q, Guo Z, et al. Hsa-Circ-0001275: a potential novel diagnostic biomarker for postmenopausal osteoporosis. Cell Physiol Biochem. 2018;46(6):2508–2516. doi:10.1159/000489657
35. Ziats MN, Rennert OM. Identification of differentially expressed microRNAs across the developing human brain. Mol Psychiatry. 2014;19(7):848–852. doi:10.1038/mp.2013.93
36. Prabhakaran S, Rivolta J, Vieira JR, et al. Symptomatic intracerebral hemorrhage among eligible warfarin-treated patients receiving intravenous tissue plasminogen activator for acute ischemic stroke. Arch Neurol. 2010;67(5):559–563. doi:10.1001/archneurol.2010.25
37. Majeed A, Kim YK, Roberts RS, Holmström M, Schulman S. Optimal timing of resumption of warfarin after intracranial hemorrhage. Stroke. 2010;41(12):2860–2866. doi:10.1161/STROKEAHA.110.593087
38. Orken DN, Kenangil G, Uysal E, Forta H. Cerebral microbleeds in ischemic stroke patients on warfarin treatment. Stroke. 2009;40(11):3638–3640. doi:10.1161/STROKEAHA.109.559450
39. Clopidogrel/aspirin combination therapy in high-risk atrial fibrillation patients. Cardiovasc J Afr. 2009;20(3):210.
40. Sacco RL, Prabhakaran S, Thompson JL, et al. Comparison of warfarin versus aspirin for the prevention of recurrent stroke or death: subgroup analyses from the Warfarin-Aspirin recurrent stroke study. Cerebrovasc Dis. 2006;22(1):4–12. doi:10.1159/000092331
41. LaMonte MP, Brown PM, Hursting MJ. Stroke in patients with heparin-induced thrombocytopenia and the effect of argatroban therapy. Crit Care Med. 2004;32(4):976–980. doi:10.1097/01.CCM.0000119426.34340.E2
42. Kelly J, Lewis R. Heparin thromboprophylaxis after acute stroke. Age Ageing. 2001;30(1):89–90. doi:10.1093/ageing/30.1.89
43. Oh YM, Lee SW, Kim WK, et al. Age-related Huntington’s disease progression modeled in directly reprogrammed patient-derived striatal neurons highlights impaired autophagy. Nat Neurosci. 2022;25(11):1420–1433. doi:10.1038/s41593-022-01185-4
44. Wu DP, Zhao YD, Yan QQ, Liu LL, Wei YS, Huang JL. Circular RNAs: emerging players in brain aging and neurodegenerative diseases. J Pathol. 2023;259(1):1–9. doi:10.1002/path.6021
45. Hezroni H, Perry RBT, Ulitsky I. Long noncoding RNAs in development and regeneration of the neural lineage. Cold Spring Harb Symp Quant Biol. 2019;84:165–177. doi:10.1101/sqb.2019.84.039347
46. Schneider MF, Müller V, Müller SA, Lichtenthaler SF, Becker PB, Scheuermann JC. LncRNA RUS shapes the gene expression program towards neurogenesis. Life Sci Alliance. 2022;5(10):e202201504. doi:10.26508/lsa.202201504
47. Tao Y, Liu Q, Wang Y, et al. Multiomics insight into the role of glucagon-like peptide-1 receptor agonists in heart failure. ESC Heart Fail. 2025. doi:10.1002/ehf2.15310
48. Marthi A, Donovan K, Haynes R, et al. Fibroblast growth factor-23 and risks of cardiovascular and noncardiovascular diseases: a meta-analysis. J Am Soc Nephrol. 2018;29(7):2015–2027. doi:10.1681/ASN.2017121334
49. Luitse MJ, Velthuis BK, Kappelle LJ, van der Graaf Y, Biessels GJ. Chronic hyperglycemia is related to poor functional outcome after acute ischemic stroke. Int J Stroke. 2017;12(2):180–186. doi:10.1177/1747493016676619
50. Thomson DW, Dinger ME. Endogenous microRNA sponges: evidence and controversy. Nat Rev Genet. 2016;17(5):272–283. doi:10.1038/nrg.2016.20
51. Boehme AK, Esenwa C, Elkind MS. Stroke risk factors, genetics, and prevention. Circ Res. 2017;120(3):472–495. doi:10.1161/circresaha.116.308398
52. Gretarsdottir S, Thorleifsson G, Manolescu A, et al. Risk variants for atrial fibrillation on chromosome 4q25 associate with ischemic stroke. Ann Neurol. 2008;64(4):402–409. doi:10.1002/ana.21480
53. Chauhan G, Arnold CR, Chu AY. Identification of additional risk loci for stroke and small vessel disease: a meta-analysis of genome-wide association studies. Lancet Neurol. 2016;15(7):695–707. doi:10.1016/S1474-4422(16)00102-2
54. French CR, Seshadri S, Destefano AL, et al. Mutation of FOXC1 and PITX2 induces cerebral small-vessel disease. J Clin Invest. 2014;124(11):4877–4881. doi:10.1172/jci75109
55. Malik R, Traylor M, Pulit SL, et al. Low-frequency and common genetic variation in ischemic stroke: the METASTROKE collaboration. Neurology. 2016;86(13):1217–1226. doi:10.1212/wnl.0000000000002528
56. Akil AA, Yassin E, Al-Maraghi A, Aliyev E, Al-Malki K, Fakhro KA. Diagnosis and treatment of type 1 diabetes at the Dawn of the personalized medicine era. J Transl Med. 2021;19(1):137. doi:10.1186/s12967-021-02778-6
57. Yuan S, Xu F, Li X, et al. Plasma proteins and onset of type 2 diabetes and diabetic complications: proteome-wide Mendelian randomization and colocalization analyses. Cell Rep Med. 2023;4(9):101174. doi:10.1016/j.xcrm.2023.101174
58. Fajans SS, Bell GI, Polonsky KS. Molecular mechanisms and clinical pathophysiology of maturity-onset diabetes of the young. N Engl J Med. 2001;345(13):971–980. doi:10.1056/NEJMra002168
59. Zhu J, Jiang Y. Editorial: the role of diabetes in the pathophysiology and prognosis of ischemic stroke. Front Endocrinol. 2023;14:1207537. doi:10.3389/fendo.2023.1207537
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Identification of 7-HOCA as a Potential Biomarker in Glioblastoma: Evidence from Genome-Wide Association Study and Clinical Validation
Zhao Z, Xing N, Sun G
International Journal of General Medicine 2024, 17:6185-6197
Published Date: 13 December 2024
The Causal Relationship Between Circulating Metabolites and the Risk of Atopic Dermatitis: A Two-Sample Mendelian Randomization Study
Chen J, Jian D, Bai B
Clinical, Cosmetic and Investigational Dermatology 2025, 18:567-577
Published Date: 13 March 2025
Multidimensional Analysis of Frailty and Its Influencing Factors in Hospitalized Elderly Stroke Patients
Zhou H, Han Y, Xie D, Zheng K, Zhu H, Zhou Z, Ji Y
Clinical Interventions in Aging 2025, 20:1741-1755
Published Date: 11 October 2025