Back to Journals » Eye and Brain » Volume 15
Meridional Attentional Asymmetries in Astigmatic Eyes
Authors de Lestrange-Anginieur E
Received 6 February 2023
Accepted for publication 6 May 2023
Published 15 May 2023 Volume 2023:15 Pages 63—76
DOI https://doi.org/10.2147/EB.S407481
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Margaret Wong-Riley
Elie de Lestrange-Anginieur1– 5
1School of Optometry, The Hong Kong Polytechnic University, Kowloon, Hong Kong; 2Centre for Myopia Research, School of Optometry, The Hong Kong Polytechnic University, Kowloon, Hong Kong; 3Research Centre for SHARP Vision (RCSV), The Hong Kong Polytechnic University, Kowloon, Hong Kong; 4Laboratory of Experimental Optometry, Centre for Myopia Research, School of Optometry, The Hong Kong Polytechnic University, Kowloon, Hong Kong; 5Centre for Eye and Vision Research (CEVR), Kowloon, Hong Kong
Correspondence: Elie de Lestrange-Anginieur, The Hong Kong Polytechnic University, Hung Hom, Kowloon, Hong Kong, Tel +8522766 4186, Fax +852 2764 6051, Email [email protected]
Purpose: To investigate the impact of attention orientation in young myopic adults with astigmatism.
Methods: The effect of attention on foveal meridional performance and anisotropy was measured in corrected myopes with various levels of astigmatism (with-the-rule astigmatism ≤ − 0.75D, Axis: 180 ± 20) using orientation-based attention. Attention was manipulated by instructing subjects to attend to either the horizontal or the vertical line of a central pre-stimulus (a pulsed cross) along separate blocks of trials. For each attention condition, meridional acuity and reaction times were measured via an annulus Gabor target situated remotely from the cross and presented at random horizontally and vertically in a two-alternative forced-choice employing two interleaved staircase procedures (one-up/one-down). Attention modulations were estimated by the difference in performance between horizontal and vertical attention.
Results: Foveal meridional performance and anisotropy were strongly affected by the orientation of attention, which appeared critical for the enhancement of reaction times and resolution. Under congruent orienting of attention, foveal meridional anisotropy was correlated with the amount of defocus for both reaction time and resolution, demonstrating greater vertical performance than horizontal performance as myopia increased. Compatible with an attentional compensation of blur through optimal orienting of attention, vertical attention enhanced reaction times compared to horizontal attention and was accompanied by an increase in overall acuity when myopia increased. Increased astigmatism was associated with smaller attention effects and asymmetry, suggesting potential deficits in the compensation of blur in astigmatic eyes.
Conclusion: Collectively, attention to orientation plays a significant role in horizontal-vertical foveal meridional anisotropy and can modulate the asymmetry of foveal perception imposed by the optics of the eye in episodes of uncorrected vision. Further work is necessary to understand how attention and refractive errors interact during visual development. These results may have practical implications for methods to enhance vision with attention training in myopic astigmats.
Keywords: myopia, astigmatism, meridional anisotropy, attention, visual performance
Introduction
Refractive anomalies are a leading cause of visual impairment and have been recognized as a major health concern worldwide.1 Reported as amongst the most common refractive errors,2 the prevalence of myopia and astigmatism has been increasing in many parts of the world, including East Asia3,4 and the United States,3,5–7 representing a serious burden to both patients and the economy.8,9 These refractive anomalies are often associated,10 leading researchers to surmise that astigmatism may be a by-product of the over-elongation of the eyeball in myopia.11 The presence of astigmatism is manifested by meridional-varying refractive power, leading to a constant retinal blur, which is not correctable without the use of eyeglasses or contact lenses.12 With the-rule (WTR) astigmatism, in which the horizontal refractive power is weaker than the vertical refractive power, is the most common form of astigmatism in Asian myopic adults.12,13 While there is a broad consensus on the myopiagenic influence of environmental blurs,14–17 to date, the etiopathogenesis of myopia, and its relationship with astigmatism, are still highly debated. A relatively neglected area in myopia research has been the contribution of the brain in the emmetropization process, despite converging evidence of the possible influence of brain signals from animal studies.18,19 Such a contribution, though still unresolved, could take place through the autonomic neural mediation of the myopiagenic eye responses to environmental blur. Therefore, an important question is whether, and to what extent, the brain can detect blur and counteract its visual consequences. Although the answer remains an enigma, it appears that blur exposure is often dealt with through ocular compensatory responses of the eyeball in myopes.20 While this compensation improves the image quality of retinal blurred stimulations, the downside is that it operates irreversibly, causing a permanent retinal blur.21 When this occurs, the resulting degradation of retinal images falls on post-retinal processes, which remodel the perceived image depending on the recruited neuronal pathways (or visual channels) of the stimulation.22–24 In this remodelling, the role of attention is important as attending to visual feature(s) allows the selective prioritization of information processing by increased firings of neurons preferring the attended feature(s) and suppression of firing in other neurons.25 This attention-related neuronal modulation has been shown to enhance a variety of visual tasks, including visual acuity.26–28 This process coincides with the phenomenon of blur adaptation, which has been shown to improve defocused visual acuity and modulate contrast sensitivity.29 It has been suggested that an increased neuronal response of attended features could influence adaptation.30 Thus, under complex blur environments, the adaptability of neural adjustment driven by blur characteristics31 may benefit from task-oriented attention. More importantly, attention involves a wide network of neuronal connections,32 which could enable the mediation of the oculomotor response regulating retinal environmental blurs,33 in addition to neural coding of stimuli. However, it remains unknown whether attention can selectively enhance or suppress blur.28
One reason is that the features on which attention selectively acts remain only partially determined,25 as most studies have focused on modulation of the neuronal response through attention to low-level features, including spatial locations,34 colour,35 orientation,30 or spatial frequency.36 Considering that blur involves a combination of low-level features, attention may influence the processing of blur. In this respect, previous studies have shown that attention and defocus blur jointly modulate visual performance in near-peripheral vision, such that an increase in attention could counter a reduction in optical quality, and vice-versa.26 It has also been reported that reduced visual acuity can be accompanied by increased attention efficiency in peripheral vision.26
Although it is not clear how this interaction works in ametropic eyes, evidence of an association between myopia and visuospatial attention has been systematically reported37–40 as demonstrated by the observation that reaction times in a visual search task tended to be more affected by distractors in myopes than emmetropes.40 Use of a peripheral detection task on Gabor revealed stronger attenuation of contrast sensitivity in myopes, with the deployment of attention to the fovea.37 Myopes were also found to perform better in a foveal discrimination task (half-face matching task) requiring a narrow focus of attention.38 To explain the augmented impact of attention with myopia, it was proposed that myopes may use a narrower attentional window.37,40 Based on the zoom lens model,41 narrowing spatial attention could provide increased resolution around the attended location, and, thus, enable a flexible compensation of uncorrected blurred acuity in myopes. To date, however, whether myopes can effectively compensate through attention for the effects of uncorrected refractive errors remains to be determined.
In this study, it was reasoned that, if attention does counteract the effects of optical blur by enhancing visual processing, attention (modulations) would differ across the meridionally-varying blur of astigmatic myopes. Doing so would enable rebalancing of the visual performance across anisotropic optical meridians. For instance, the meridian with greatest refractive errors could be more enhanced by attention than the perpendicular meridian with lower refractive errors in order to minimize the asymmetrical blur inputs of astigmatism. Given the orientation dependence of astigmatic blur,11,42–44 such modulations could be realized through feature-based orientation.45 In this respect, a possible neural retuning with refractive errors is indicated by the alterations of performance anisotropy in adult myopic astigmats,46 as compared to adult myopes and emmetropes, under full optical correction. Thus, in WTR myopic astigmats, it has been shown that there is significant superiority of acuity performance for isolated gratings along the vertical as compared to the horizontal (the horizontal effect).46 Conversely, in emmetropes, the foveal vertical-horizontal asymmetry was found to be reversed and minimal,46 leaving the well-known oblique effect47 – an inferior performance for oblique orientation compared to cardinal (vertical/horizontal) orientation – as the predominant anisotropy observable for isolated gratings. While it appears that no studies have investigated the attentional origin of this neural retuning, anisotropic effects of attention have been previously demonstrated in peripheral vision48–50 with a dominance of attention in the lower visual field as compared to the upper visual field48,50 and in the vertical meridian as compared to the horizontal49 for non-foveal stimuli. Also relevant to the current study is the observation of a modulatory effect of attentional load on the oblique effect for orientation discrimination tasks in normal subjects,51 underscoring the potential of attention to act on spatially localized anisotropies in performance.
To test the existence of an astigmatic-blur-related attentional anisotropy, myopic subjects with various levels of astigmatism, but identical astigmatism axis was recruited, ensuring that all the subjects had the greatest blur along the same meridian. It was hypothesized that : 1) Attention to orientation selectively increases foveal resolution to the attended meridian, enabling separate compensation of horizontal and vertical blurred acuity (ie, meridional blur); and 2) attentional modulation is superior to the predominant blurred meridian. The study revealed that attention orientation produces both selective and global enhancements of meridional performance. For resolution acuity, global attention enhancement coincided with a preferential attention orientation, which varied with the amount of individual defocus. In agreement with a meridional attention compensation of blur, differential attention-related modulations between horizontal and vertical blurred acuity were found for both reaction time and resolution acuity. Astigmatism was found to reduce attention effect and asymmetry, highlighting possible attentional deficits with higher astigmatism.
Methods
Subjects
Visually corrected astigmatic subjects (Snellen equivalent > 20/20 in the tested eye) with various levels of refractive errors (Sphere: −3.47 ± 1.35D, Cylinder: −1.72 ± 0.78D) were recruited from a pool of thirty optometrist students, who underwent both subjective and objective refraction using an open-autorefractor (Shin-Nippon NVision-K 5001, Rexxam Co., Ltd., Kagawa, Japan). Given the limited proportion of low-moderate myopic young adults with WTR astigmatism in Hong Kong (>28% in the age range of 21–30 years, including hyperopes and high myopes),13 of the thirty potential participants, only nine subjects (Table 1, four males and five females, aged 21–24) matched the eligibility criteria: (1) spherical error: −0.50 D to −5.50 D; (2) cylindrical error < −0.75 D; (3) cylindrical axis: 180 ± 20 in the dominant eye; (4) anisometropia ≤2 D; (5) absence of ocular and systemic diseases; strabismus, or amblyopia treatment; and (6) no history of myopic control treatment. Given that all the subjects were compound myopes having WTR astigmatism, the most defocused meridian was near the vertical, where optical resolution is poorer. The sample size was chosen to provide a representative range of refractive errors in compound myopic astigmatism and achieve a statistically significant effect of attention. The experimental procedures were approved by the human ethics committee of The Hong Kong Polytechnic University (HSEARS20170103003), and the research was conducted according to the principles expressed in the Declaration of Helsinki. Written informed consent was obtained from each participant.
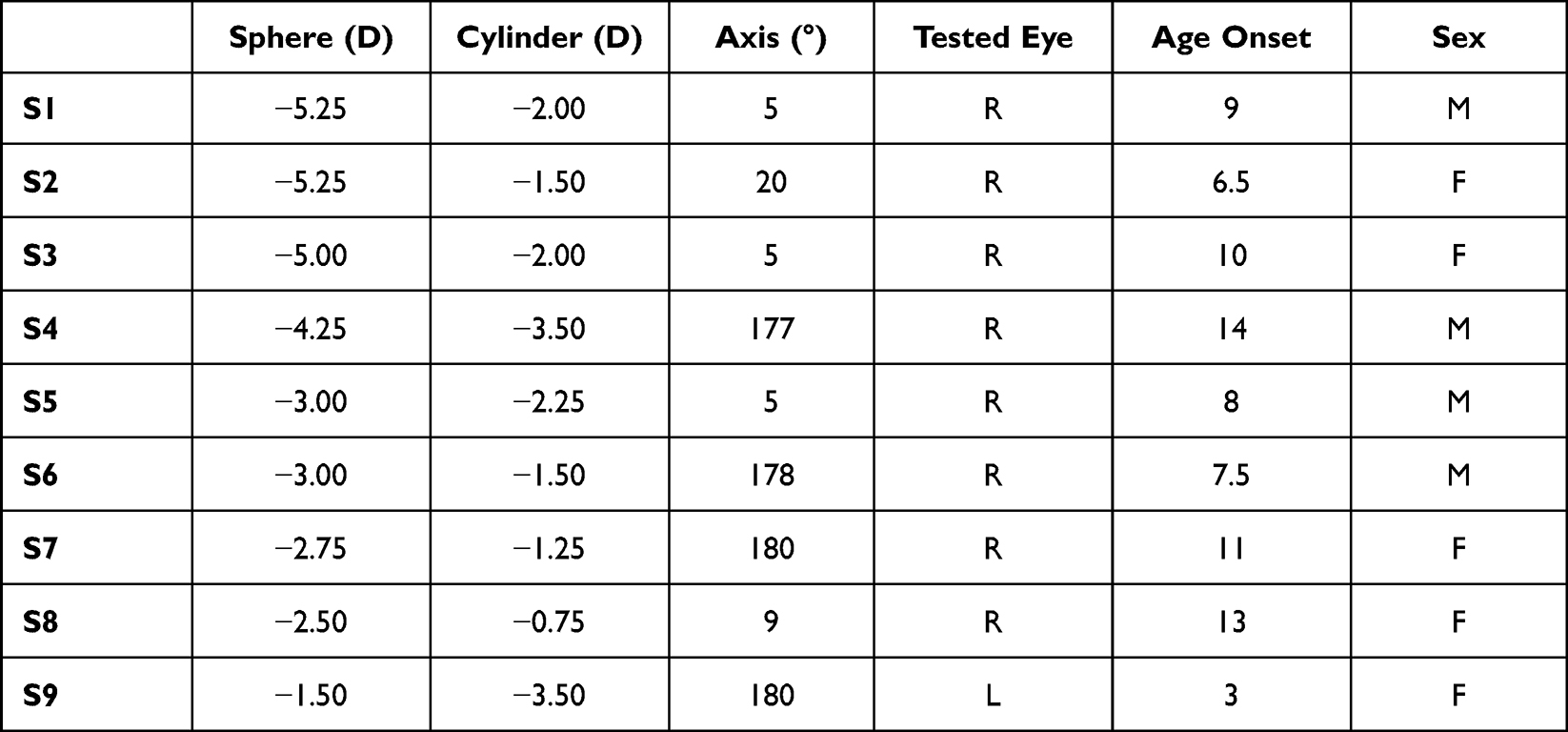 |
Table 1 Characteristics of Subjects |
Experimental Procedures
Apparatus
Stimuli were presented on a Dell LCD gamma-corrected monitor (Dell Technologies, Round Rock, TX, USA; resolution: 3840 × 2160, frame rate: 60Hz) on a grey background (25cd/m2). Throughout the test, the subject viewed the display monocularly (viewing distance: 3 meters) and with full correction, while instructed to fixate on the centre of the display. Each session lasted 4h.
Procedure
At the beginning of each session, subjects underwent a phase of training trials to familiarize themselves with the task and the instructions. Following the training, each test began with a subjective threshold adjustment of the spatial frequency of the grating oriented obliquely. This pre-adjustment was used to adjust the starting spatial frequency of the target stimuli (spatial frequency range: 20–40 cpd) on account of individual variability in performance. After pre-adjustment of an individual’s threshold, a fixation cross (depicted in Figure 1) (Length: 1°), serving as an adapter, appeared at the centre of the display. The cross adapter was used to help subjects direct their attention along the attended orientation. The width of the cross was continuously modulated (Width range: 0.02–20arcmin, Increment: 2.5arcmin, Frequency beat: 2Hz) in order to engage subjects in the attentive tracking of one of the lines of the cross.52 The cross adaptor appeared at the beginning of the test (duration: 40s) and before each target onset (duration: 4s) with a voice prompt instructing the subject to attend to one of the two lines of the central cross, the horizontal or the vertical. The long and short presentations of the cross were separated by a blank background (ISI: 1s). Using the same physical adaptor for both attentional conditions ensured that modulation of performance was attributed to attention only. As in previous studies using a similar paradigm,30,53 no neutral meridional attention was introduced, as it would have implied the change in the physical adaptor, preventing the comparison of the attention conditions. After the cross disappeared (ISI: 1s), an annulus Gabor target (Inner/Outer diameter: 1.25–5°, contrast: 50%) was briefly presented in the fovea (duration: 0.3s). The grating was filtered by a rotationally symmetric Gaussian low pass filter with a standard deviation σ=0.7° to reduce edge effects. Considering the difficulty of singling out one of the two lines, the cross was designed with vertical (grey-on-white, contrast: 100%) and horizontal (grey-on-black, contrast: 100%) lines having opposite polarity. To ensure that there was no local luminance adaptation and polarity-related bias, the size of the fixation cross was constrained such that the visual cue and the annulus target did not overlap spatially. Considering that there was a negligible effect of polarity on spatial interaction,54,55 the possibility that polarity could result in different contrast adaptation effects at the remote location of the target was also excluded.
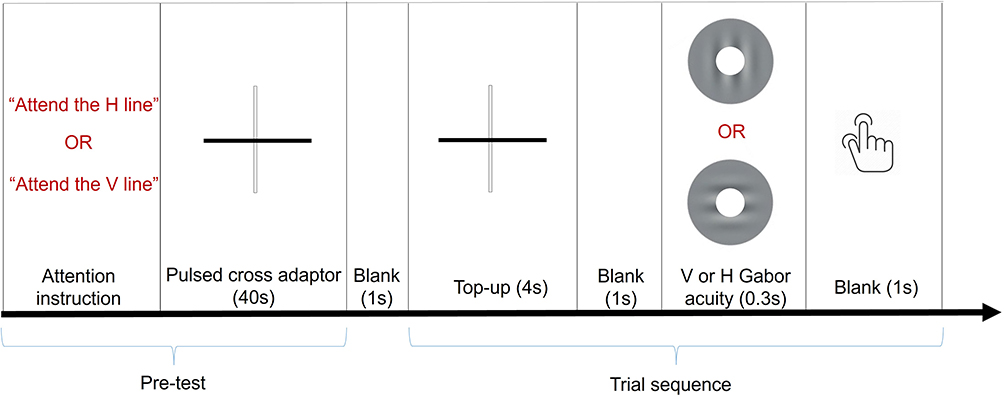 |
Figure 1 The temporal structure of the study. The sequence of the stimulation started with the presentation of a high contrast pulsed cross adaptor, used to direct the attention of the subject along the vertical or the horizontal at the beginning of the test and just before the presentation of the visual target. The visual target, an annulus Gabor either vertical (V) or horizontal (H), is used to determine resolution acuity under the meridional attention manipulation. |
Visual Conditions & Test
Each attention condition (i.e., horizontal attention and vertical attention) was alternatively tested in brief separate blocks. During each attention condition session, horizontal (H) and vertical (V) resolution acuity (RA) were tested randomly. Subjects performed a total of ten blocks of trials for each condition (horizontal vs vertical attention, horizontal vs vertical Gabor). Each block of trials was separated by a break of 5 min to prevent the decline of attention and attenuate adaptation. The subject’s task was to identify the orientation of the Gabor. Trials were congruent when the orientation of attention matched the orientation of the target and incongruent when there was a mismatch.
RA: For each block condition, the spatial frequency of the grating was controlled via a 1-up-1-down staircase method converging to the 50% threshold with a step size of 3 cpd. To determine retinal resolution, each of the measured thresholds was averaged and corrected by accounting for the horizontal and vertical magnification effects introduced by the spectacle lens correction.56
RT: In order to evaluate the effect of adaptation and attention, reaction time (RT) was measured as the duration from the target onset to the button press. To eliminate outliers, an upper and lower cut-off was applied to RTs, and values exceeding two standard deviations were removed. For each block, the average RT was calculated.
Baseline Condition
To evaluate adaptation, resolution acuity and RTs were measured in a separate test in the absence of attention instruction and adapter using the annulus grating acuity.
Analysis
RT (in milliseconds) and RA (in cycles per degree cpd) were used to determine the foveal vertical-horizontal asymmetry (fVHA), measured as the ratio between vertical (V) and horizontal (H) performance. Visual performance and its meridional/attentional modulations were regressed using Pearson correlation against the amount of sphere (Sph) and cylinder (Cyl) with the goal to find whether, and to what degree, the overall blur (defocus) and meridional blur difference (astigmatism) accounted for the attention-related modulations. Two types of attention-related modulations of performance were analyzed:
- congruency-related attention modulation (cAE), which measures the effect of attending to the meridian congruent to the target orientation as compared to the meridian incongruent to target orientation. This measure estimates the relative advantage of attending to the orientation of the target.
- the orientation-related attention modulation (oAE), which measures the effect of attending to the horizontal meridian as compared to the vertical. This measure estimates the relative advantage of attending to a specific meridian (eg vertical).
Objective refraction was used to report the regression, as no marked difference was found between objective and subjective refraction regression. All the data were reported with mean and standard error.
Results
Adaptor Effects
A significant elevation of mean RT (No adaptor: 904ms ± 32ms, with adaptor: 1142ms ± 42ms, Two-way repeated ANOVA: adaptor effect F(1,8) = 0.711, p = 0.002) accompanied by significant reduction of mean resolution acuity was observed in the presence of the remote adaptor (Figure 2, No adaptor: 62.270 cpd± 3.699 cpd, with adaptor: 46.148 cpd ± 2.640 cpd, Two-way repeated ANOVA: adaptor effect F(1,8) = 0.756, p = 0.001), confirming that performance was affected by the remote adaptor. No interaction between adaptor and target orientation was found (Two-way repeated ANOVA: RT, adaptor X orientation F(1,8) = 0.157 p = 0.257; Resolution acuity, adaptor X orientation F(1,8) = 0.002 p = 0.898) nor was there a statistical difference in foveal meridional anisotropy (RT: fVHA with adaptor: 0.9404 ±0.00791, fVHA without adaptor: 0.9403 ± 0.0762; paired t(8) = −0.011, p = 0.992; Resolution acuity, fVHA with adaptor: 0.04186 ± 04186, fVHA without adaptor: 1.0628 ± 0.08292; paired t(8) = 0.149, p = 0.885) observed for the conditions with or without the remote adapter, ruling out differential adaptation effects between the horizontal and vertical.
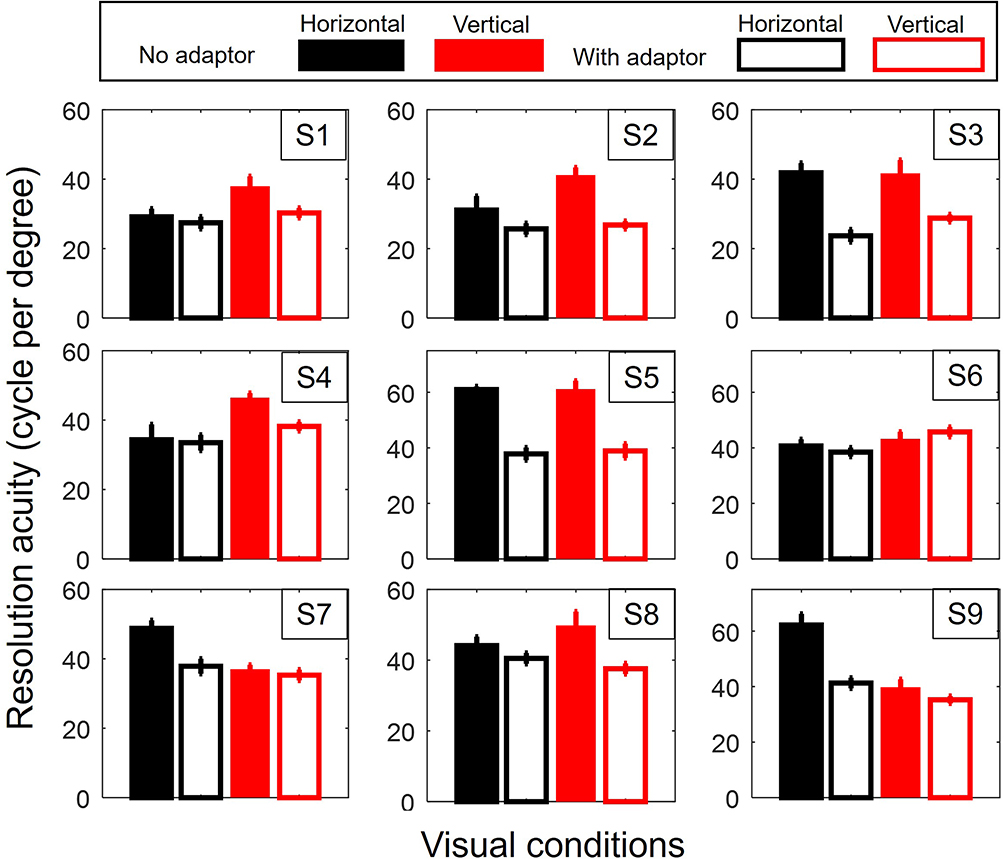 |
Figure 2 Adaptor effect. Effect of the adaptor on horizontal (H) and vertical (V) resolution acuity for each subject. In all the figures, the error bar corresponds to the standard error. |
fVHA
RT: For all subjects, mean RT was faster along the vertical (fVHA = V/H = 0.9404 ±0.00791; one sample t(8)= −7.533, p < 0.001), showing evidence of a systematic performance difference between horizontal and vertical. This confirmed the inversion of the horizontal-vertical foveal asymmetry found in myopic WTR astigmatism.46
RA: Horizontal and vertical also showed significant differences in resolution acuity in meridional difference (|V − H| = 6.703 ± 0.486, one sample t(8) = 13.794, p < 0.001), but, in contrast with RTs, the direction of fVHA varied among subjects.
fVHA and Refractive Errors
RA: When subjects attended target orientation, the vertical-horizontal asymmetry increased in line with the amount of defocus (Figure 3A, fVHA Vcongruent/Hcongruent Sph: r(9) = 0.711, p = 0.032; Cyl: r(9) = −0.253, p = 0.512), such that higher myopes had better vertical acuity than lower myopes. Interestingly, when the target was unattended, fVHA was not significantly associated with refractive errors (Sph: r(9) = −0.157, p = 0.687; Cyl: r(9) = 0.020, p = 0.960).
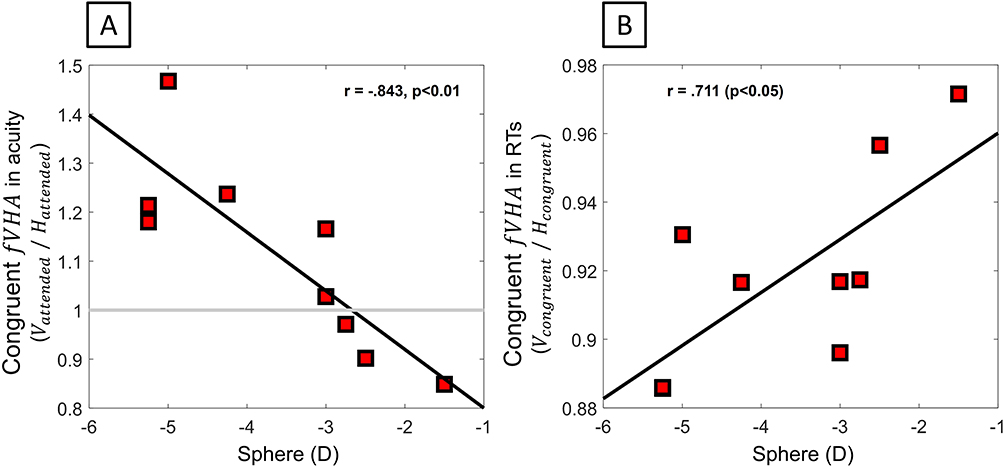 |
Figure 3 Foveal meridional anisotropy (fVHA) of performance. (fVHA) when congruently attending to the orientation of targets as a function of the amount of sphere for (A) resolution acuity and (B) RTs. As the amount of defocus increased, a relative increase in vertical performance was observed as compared to the horizontal. |
RT: RT shows the same correlation between individual congruent fVHA and the amount of defocus (Figure 3B, Sph: r(9) = −0.843, p = 0.004; Cyl: r(9) = −0.053, p = 0.892), with superior RT along the vertical as compared to the horizontal, as defocus increased. Again, when the target was unattended, fVHA was not significantly associated with refractive errors (Sph: r(9) = −0.351, p = 0.355; Cyl: r(9) = −0.023, p = 0.953). Overall, the results indicated that, in situations of enhanced processing, congruent attention tended to advantage processing on the vertical as compared to the horizontal meridian when myopia increased.
Congruence Effect on fVHA
RT: In addition, congruent attention significantly accentuated the vertical-horizontal asymmetry in RTs as compared to incongruent attention for higher myopes (Figure 4A, N = 7, Sphere < −2.50D, fVHAcongruent/fVHAincongruent = 0.932 ± 0.021; one-sample t(6) = 3.249, p = 0.017), indicating a significant impact of attention on the magnitude of vertical-horizontal asymmetry in higher myopes.
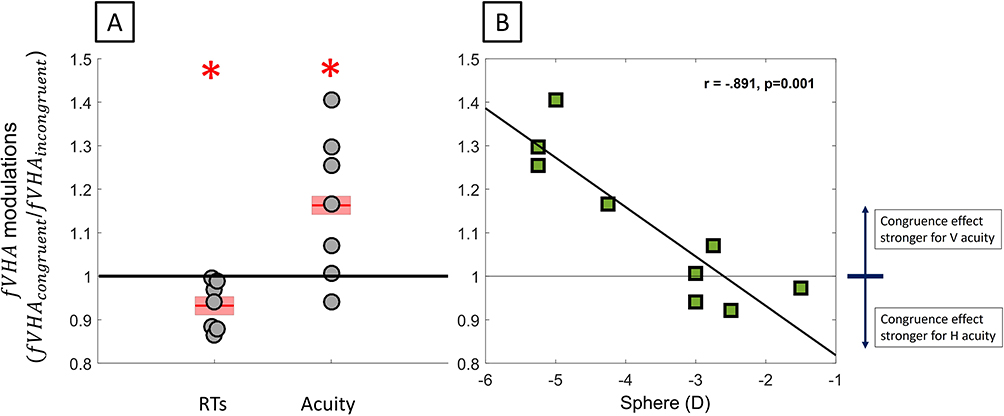 |
Figure 4 Effect of attention congruency on (fVHA). (A) Attention-related modulation of meridional asymmetry for RTs and acuity in subjects with sphere <−2.50D. (B) Attention-related modulation of meridional asymmetry in acuity as a function of sphere. *P < 0.05. |
RA: For resolution acuity, there was a strong correlation between the attention-related change in fVHA and the amount of defocus (Figure 4B, fVHAcongruent/fVHAincongruent Sph: r(9) = −0.891, p = 0.001; Cyl: r(9) = −0.082, p = 0.833), confirming the increased vertical-horizontal asymmetry for higher myopes.
Congruency Effect
RT: Globally, attending to a specific meridian systematically statistically improved RTs for that meridian (Figure 5, cAE = RTcongruent/RTincongruent = 0.957 ±0.013; one sample t(8) = −3.319, p = 0.011), confirming that attention was successfully oriented to the distinct orientation to be attended to by the subjects during the task. Considering the relationship between RT and contrast,57,58 this selective enhancement aligned with the possibility of compensatory effect of attention on meridional blur, whereby attending to a meridian resulted in improvement of blurred acuity.
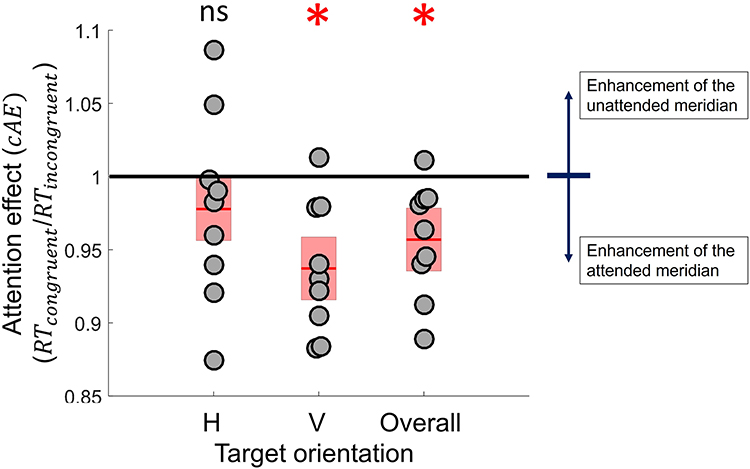 |
Figure 5 Congruency-related attentional modulation of RTs. Change in RTs was associated with switching attention from the congruent to the incongruent meridian for horizontal, vertical or both target orientations. The grey dots indicate the individual attention effect on RT values for each target meridian; the red bars indicate the mean value; the shaded areas represent one standard error. *P < 0.05, ns not significant. |
RA: For RA, a statistically significant change of the magnitude of resolution with attention congruency was observed (|Congruent-Incongruent| = 3.655 ± 0.653, one sample t(8) = 5.592, p < 0.001). While there was a slight but insignificant tendency of attention-related modulations to improve the resolution of the attended meridian, in contrast with RTs, no systematic overall improvement (5/9 subjects) of the attended meridian was found (Figure 6A, cAE = 1.018 ± 0.022, one sample t(8) = 0.800, p = 0.447), indicating there were individual differences in the visual effects of attention congruency. In contrast, a strong tendency for the strength of congruence-related attention modulation to increase as astigmatism decreased was observed (Figure 6B, |Congruent-incongruent|, Sph: r(9) = 0.004, p = 0.991; Cyl, r(9) = 0.845, p = 0.004), suggesting the possibility of a reduced meridional effect of attention switching in higher astigmats.
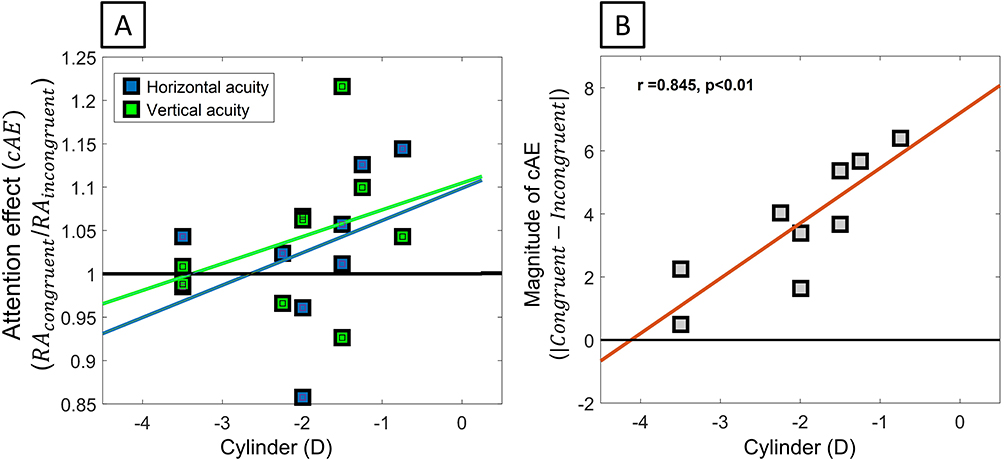 |
Figure 6 Congruency-related attentional modulation of resolution acuity. (A) Change in resolution acuity associated with switching attention from the congruent to the incongruent meridian for horizontal to vertical. (B) Magnitude of the congruence-related attention modulation as a function of astigmatism. |
Meridional Asymmetry of Attention
RT: However, consistent with a foveal asymmetry of attention strength, a statistically significant increased magnitude of attention asymmetry was observed (|cAEV − cAEH| = 0.068 ± 0.018, one sample t(8) = 3.80, p = 0.0005). The congruence-related attentional modulation was also significantly greater for vertical RT, as compared to horizontal RT for higher myopes (Figure 7A, N = 7, Sphere < −2.50D, cAEv–cAEH = −0.07 ±0.022; one-sample t(6) = 3.128, p = 0.020) in line with the greater (congruence-related) attention enhancement of the poorer vertical meridian.
 |
Figure 7 Meridional asymmetry of congruence-related attention. (A) Attention-related modulation of meridional asymmetry for RTs and acuity in subjects with sphere <−2.50D. (B) Attention-related modulation of meridional asymmetry in acuity as a function of sphere. *P < 0.05, ns not significant. |
RA: For RT, a significant magnitude of meridional asymmetry in attention effects was observed (|cAEv − cAEH| = 0.279±0.030, one sample t(8) = 9.238, p < 0.001), which significantly increased with myopia (Sph: r(9)= −0.687, p = 0.041; Cyl, r(9) = −0.072, p = 0.853). Again, as myopia increased, attention congruency significantly enhanced acuity of the poor vertical meridian as compared to horizontal acuity (Figure 7B, cAEv – cAEH Sph: r(9) = −0.873, p = 0.002; Cyl, r(9) =−0.053, p = 0.892), with a superior attention effect in vertical acuity for the higher myopes (Sphere < −4.00D).
Orientation of Attention
RT: Interestingly, when attending to the vertical, the overall RT was significantly improved for the higher myopes (Figure 8A, N = 7, Sphere < −2.50D, oAE: 1.036 ± 0.012; one-sample t(6) = 3.018, p = 0.023) and the vertical-horizontal foveal asymmetry significantly heightened, as compared with when attending to the horizontal (fVHAattend v/fVHAattend H = 0.916 ± 0.025; one-sample t(8) = −3.402, p = 0.009). This finding highlights the existence of a better orientation of attention for overall performance, which was associated with a greater meridional asymmetry.
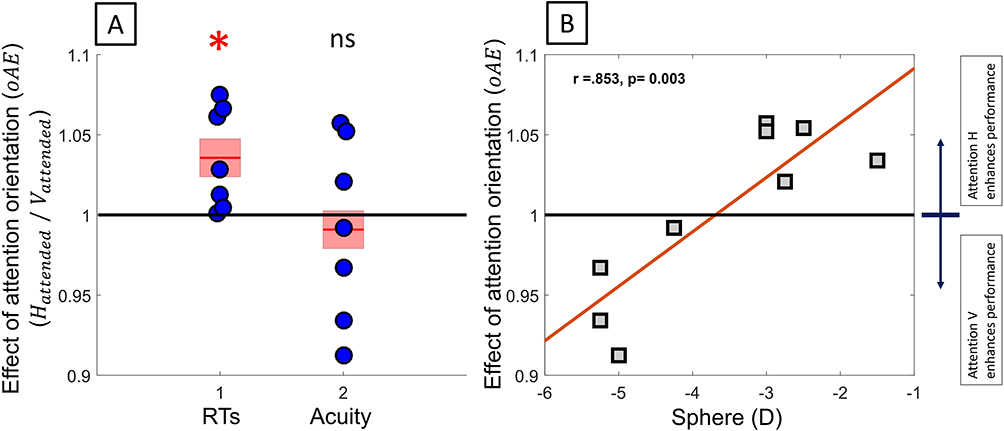 |
Figure 8 Orientation-related attentional modulations. (A) Attention-related modulation of meridional asymmetry for RTs and acuity in subject with sphere <−2.50D. (B) Orientation-related attentional modulations in overall acuity as a function of defocus. *P < 0.05, ns not significant. |
RA: A strong correlation was observed between overall orientation-related attentional modulations of acuity and the amount of defocus (Figure 8B, oAEoverall =RAAttend H/RAAttend V: Sph: r(9) = 0.853, p = 0.003; Cyl, r(9) = 0.056, p = 0.886), which confirmed an optimal orientation of attention to enhance overall acuity. Attending the poor vertical meridian tended to improve the overall resolution more for higher myopes (Figure 8B, N = 4, Sphere < −4.00D, 0.951 ± 0.018, one sample t(3) = 2.766, p = 0.070) but decreased resolution for low myopes (Figure 8B, N = 5, Sphere ≥ −3.00D, 1.044 ± 0.0007, one sample t(4) = 6.214, p = 0.003), as compared to attending to the horizontal.
Concurrent with this overall enhancement through attention orienting, the strength of attention asymmetry (Figure 9A and Figure 9, |oAEH − oAEV|: Cyl: r(9) = 0.778, p = 0.014) was shown to be significantly correlated with the amount of cylinder, but not with the amount of defocus (Sph: r(9) = −0.115, p = 0.768). Remarkably, this meridional difference in attention effect became more pronounced as astigmatism decreased, indicating reduced meridional attentional difference for low astigmats. While it suggested an effect of meridional blur difference in attention modulation, the direction of this modulation was in contradiction with the hypothesis that optical resolution difference increases attention difference and thus coinciding more with the idea of an attentional deficit, whereby the differential meridional effect of attention orienting would be reduced by astigmatism.
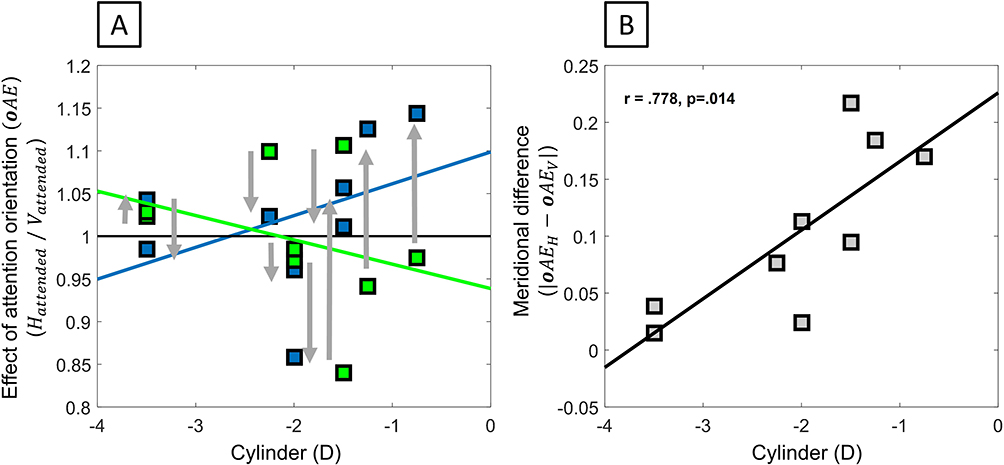 |
Figure 9 Meridional difference in orientation-related attentional modulations. (A) Orientation-related attentional modulations of vertical and horizontal acuity. (B) Magnitude of the difference in the orientation-related attentional modulations as a function of astigmatism. The arrows indicate the acuity changes in orientation-related attentional modulations between meridians. |
Discussion
Studies on foveal meridional anisotropy in corrected astigmatic subjects46,59,60 and neural compensatory effects of attention in normal eyes48–51 showed the existence of asymmetry of visual performance related to individual refractive status and attention modulation, respectively. To our knowledge, this is the first study that bridges the gap between these two separate streams of research by assessing the relationship between attention orientation and meridional performance in astigmatic myopes. In agreement with previous studies showing an interaction between attention and blur,26,27 the results demonstrated pronounced attention-related modulations of horizontal-vertical foveal asymmetry (Figures 5 and 6) affecting reaction times and resolution acuity, which points to the likely impact of attention on blurred acuity. In addition, this study revealed the first evidence that attention-related modulations are significantly correlated with individual refractive errors in astigmats (Figures 4B and 6–9B). Such correlation supports the existence of compensatory attentional changes, which could counteract the effects of refractive errors. Thus, the unresolved origin of refractive error-related changes in meridional anisotropies found in young adults46 is likely to reflect, not just changes in sensory adaptation,31 but also variations in the brain networks underlying feature-based attentional control.61–63 A change in attentional network in myopic astigmats is consistent with the recent observation in myopes of brain changes,64–66 including regions supporting attention. Given the influence of attentional networks on cognition and emotion,67–69 an alteration of voluntary attentional control with refractive errors could also have important behavioral and psychological implications,69–72 which warrants further investigations. These results may open up new perspectives for enhancing vision using attention training in myopic astigmats.
Changes in RTs indicated modulations of attention systematically improved the attended meridian (Figure 3A), providing evidence that subjects oriented their attention to the instructed orientation. Attention congruency was superior at the attended vertical meridian in the higher myopes (Figure 4A), consistent with the idea that flexible orienting of attention can take place to compensate for the meridional asymmetry of performance arising from optical or/and neural asymmetry. While selective RT enhancements did not systematically reflect on spatial performance, the results revealed that attending to a specific meridian produced a global modulation of RT and resolution across the two meridians (Figure 7B), more compatible with a compensatory effect of the overall blurred acuity through optimal orienting of attention. For higher myopes, this general enhancement was achieved when subjects attended to the stronger blurred meridian (ie, the vertical), highlighting a preferential orientation of attention.
Concurrent to this general modulation, astigmatism-related changes in the magnitude of attention asymmetry were observed (Figure 8A and Figure 8). Remarkably, the strength of attention asymmetry did not increase with astigmatism as expected if attention had differentially enhanced the meridional blurs, but resulted in the opposite effect. It follows that increased astigmatism may impair the asymmetry of attention modulation observed in normal subjects in favour of a general correction of blur through a more rigid, but also more optimal orientation of attention. Interestingly, such an attention deficit echoes previous findings showing an alteration of attention effect in myopes.34–37 Disentangling the separate effects of defocus and astigmatism on attention effects remains needed to clarify the impact of astigmatism per se on the compensatory effect of attention on neural asymmetry.
In summary, this study supports a general compensation of the overall blurred acuity through preferential orientation of attention, but not a meridional compensation of blurred acuity. It is likely that utilizing attention orientation to correct meridional blur is not a satisfactory strategy to correct meridional blurred acuity, as it would require the ability to rapidly switch attention between each meridian, an operation that may challenge the relatively slow time course of feature-based attention.73 In contrast, a general enhancement of acuity suggests a preferential orientation of attention in astigmatic myopes, which may be correlated with difference in attentional allocation and/or flexibility. A general acuity enhancement through preferential orienting may be acquired through learning during the period of uncorrected blur. How such preferential attention orienting is affected by the age of onset of astigmatism and its correction remains to be determined. Since no neutral baseline measurement was used during the test, this study cannot elucidate whether astigmatic subjects naturally oriented their attention toward the optimal attention orientation. Although our result demonstrated a consistent effect of attention on horizontal-vertical foveal asymmetry, a larger sample of participants would be needed to fully elucidate the meridional distribution of attention in astigmatic myopes, and the cost/benefit associated with its deployment. Considering some differences of attention-related modulation of RT and acuity, it is possible that the visual benefit of enhanced temporal processing could have been contaminated by individual differences in the adaptation effect. However, it is well known that RTs and acuity can be complexly related,74 and so these differences may be caused by the differential processes and pathways involved in rapid motor response and visual discrimination. Thus, whether attention can effectively correct for ocular blur or can be a biomarker of abnormal refractive error development requires more research. Future studies should address the important question of whether attention operates via selective compensatory response from ocular accommodation and/or an enhanced neuronal response.
Conclusion
The results of this study show that the orientation of attention modulates reaction time and resolution acuity and hence has the capability to compensate for the optical effects associated with episodes of uncorrected refractive errors during abnormal refractive development. Determining whether this attention compensation is restricted to the correction of defocus blur or more complex blur patterns is critical to clarify the degree of tuning of the brain to the eye’s optics.
Acknowledgments
The author thanks Prof. Chea-su Kee for his support and insightful comments. The author is also grateful to Dr. Maureen Boost for proofreading.
This study was supported by PolyU (UGC) Start-up Fund (P0039484), The Centre of Myopia Research from The Hong Kong Polytechnic University and received funding support from the InnoHK initiative and the Hong Kong Special Administrative Region Government.
Disclosure
The author reports no conflicts of interest in this work.
References
1. Blindness GB. Vision impairment collaborators on behalf of the vision loss expert group of the global burden of disease study. Causes of blindness and vision impairment in 2020 and trends over 30 years, and prevalence of avoidable blindness in relation to VISION 2020: the right to sight: an analysis for the global burden of disease study. Lancet Glob Health. 2021;9(2):e144–e60.
2. Hashemi H, Fotouhi A, Yekta A, Pakzad R, Ostadimoghaddam H, Khabazkhoob M. Global and regional estimates of prevalence of refractive errors: systematic review and meta-analysis. J Curr Ophthalmol. 2018;30(1):3–22. doi:10.1016/j.joco.2017.08.009
3. Dolgin E. The myopia boom. Nature. 2015;519(7543):276. doi:10.1038/519276a
4. Liang Y, Leung TW, Lian JT, Kee CS. Significant increase in astigmatism in children after study at home during the COVID-19 lockdown. Clin Exp Optom. 2022;106(1–9):322–330. doi:10.1080/08164622.2021.2024071
5. Wong SC, Kee CS, Leung TW. High prevalence of astigmatism in children after school suspension during the COVID-19 pandemic is associated with axial elongation. Children. 2022;9(6):919. doi:10.3390/children9060919
6. Vitale S, Sperduto RD, Ferris FL. Increased prevalence of myopia in the United States between 1971–1972 and 1999–2004. Arch Ophthalmol. 2009;127(12):1632–1639. doi:10.1001/archophthalmol.2009.303
7. Valluru G, Klawe J, Liu B, Ramulu P, Ahmad S. Characterizing astigmatism in the United States. J Cataract Refract Surg. 2022;48(5):519–527. doi:10.1097/j.jcrs.0000000000000786
8. Sankaridurg P, Tahhan N, Kandel H, et al. IMI impact of myopia. Invest Ophthalmol Vis Sci. 2021;62(5):2. doi:10.1167/iovs.62.5.2
9. Zhang J, Wu Y, Sharma B, Gupta R, Jawla S, Bullimore MA. Epidemiology and burden of astigmatism: a systematic literature review. Optom Vis Sci. 2023;100(3):218. doi:10.1097/OPX.0000000000001998
10. Fulton AB, Hansen RM, Petersen RA. The relation of myopia and astigmatism in developing eyes. Ophthalmology. 1982;89(4):298–302. doi:10.1016/S0161-6420(82)34788-0
11. Kee CS. Astigmatism and its role in emmetropization. Exp Eye Res. 2013;114:89–95. doi:10.1016/j.exer.2013.04.020
12. Read SA, Collins MJ, Carney LG. A review of astigmatism and its possible genesis. Clin Exp Optom. 2007;90(1):5–19. doi:10.1111/j.1444-0938.2007.00112.x
13. Leung TW, Lam AKC, Deng L, Kee CS. Characteristics of astigmatism as a function of age in a Hong Kong clinical population. Optom Vis Sci. 2012;89(7):984–992. doi:10.1097/OPX.0b013e31825da156
14. Wildsoet CF, Chia A, Cho P, et al. IMI–interventions for controlling myopia onset and progression report. Invest Ophthalmol Vis Sci. 2019;60(3):M106–M131. doi:10.1167/iovs.18-25958
15. Wang JC, Chun RK, Zhou YY, et al. Both the central and peripheral retina contribute to myopia development in chicks. Ophthalmic Physiol Opt. 2015;35(6):652–662. doi:10.1111/opo.12239
16. Choi KY, Yu WY, Lam CHI, et al. Childhood exposure to constricted living space: a possible environmental threat for myopia development. Ophthalmic Physiol Opt. 2017;37(5):568–575. doi:10.1111/opo.12397
17. Wang D, Chun RKM, Liu M, et al. Optical defocus rapidly changes choroidal thickness in schoolchildren. PLoS One. 2016;11(8):e0161535. doi:10.1371/journal.pone.0161535
18. Troilo D, Wallman J. The regulation of eye growth and refractive state: an experimental study of emmetropization. Vision Res. 1991;31(7–8):1237–1250. doi:10.1016/0042-6989(91)90048-A
19. McFadden SA, Wildsoet C. The effect of optic nerve section on form deprivation myopia in the Guinea pig. J Comp Neurol. 2020;528(17):2874–2887. doi:10.1002/cne.24961
20. Wallman J, Winawer J. Homeostasis of eye growth and the question of myopia. Neuron. 2004;43:447–468. doi:10.1016/j.neuron.2004.08.008
21. Troilo D, Smith EL, Nickla DL, et al. IMI–Report on experimental models of emmetropization and myopia. Invest Ophthalmol Vis Sci. 2019;60(3):M31–M88.
22. de Lestrange-Anginieur E, Ding Y. Modulations of face perception in response to a novel time-varying optical perturbation after aberration correction using adaptive optics. Eur Phys J Spec Top. 2022;231(4):713–722. doi:10.1140/epjs/s11734-021-00393-6
23. Goodhew SC, Clarke R. Contributions of parvocellular and magnocellular pathways to visual perception near the hands are not fixed, but can be dynamically altered. Psychon Bull Rev. 2016;23(1):156–162. doi:10.3758/s13423-015-0844-1
24. Van Essen DC, Anderson CH, Felleman DJ. Information processing in the primate visual system: an integrated systems perspective. Science. 1992;255(5043):419–423. doi:10.1126/science.1734518
25. Maunsell JH, Treue S. Feature-based attention in visual cortex. Trends Neurosci. 2006;29(6):317–322. doi:10.1016/j.tins.2006.04.001
26. De Lestrange-Anginieur E, Leung TW, Kee CS. Joint effect of defocus blur and spatial attention. Vision Res. 2021;185:88–97. doi:10.1016/j.visres.2021.04.002
27. De Lestrange-Anginieur E, Kee CS. Investigation of the impact of blur under mobile attentional orientation using a vision simulator. PLoS One. 2020;15(6):e0234380. doi:10.1371/journal.pone.0234380
28. De Lestrange-Anginieur EAJ, Leung TW, Kee CS. Does attention impact perceptual blur?
29. Cufflin MP, Mallen EA. Blur adaptation: clinical and refractive considerations. Clin Exp Optom. 2020;103(1):104–111. doi:10.1111/cxo.13033
30. Liu T, Hou Y. Global feature-based attention to orientation. J Vis. 2011;11(10):8. doi:10.1167/11.10.8
31. Webster MA, Georgeson MA, Webster SM. Neural adjustments to image blur. Nat Neurosci. 2002;5(9):839–840. doi:10.1038/nn906
32. Corbetta M, Akbudak E, Conturo TE, et al. A common network of functional areas for attention and eye movements. Neuron. 1998;21(4):761–773. doi:10.1016/S0896-6273(00)80593-0
33. Naber M, Alvarez GA, Nakayama K. Tracking the allocation of attention using human pupillary oscillations. Front Psychol. 2013;4:919. doi:10.3389/fpsyg.2013.00919
34. Carrasco M. Visual attention: the past 25 years. Vision Res. 2011;51(13):1484–1525. doi:10.1016/j.visres.2011.04.012
35. Saenz M, Buraĉas GT, Boynton GM. Global feature-based attention for motion and color. Vision Res. 2003;43(6):629–637. doi:10.1016/S0042-6989(02)00595-3
36. Martınez A, Di Russo F, Anllo-Vento L, Hillyard SA. Electrophysiological analysis of cortical mechanisms of selective attention to high and low spatial frequencies. Clin Neurophysiol. 2001;112(11):1980–1998. doi:10.1016/S1388-2457(01)00660-5
37. Kerber KL, Thorn F, Bex PJ, Vera-Diaz FA. Peripheral contrast sensitivity and attention in myopia. Vision Res. 2016;125:49–54. doi:10.1016/j.visres.2016.05.004
38. McKone E, Davies AA, Fernando D. Blurry means good focus: myopia and visual attention. Perception. 2008;37(11):1765–1768. doi:10.1068/p6156
39. Mascetti GG, Turatto M, Facoetti A. Four paradigms to study visual–spatial attention of myopic subjects. Brain Res Protoc. 2001;7(3):241–247. doi:10.1016/S1385-299X(01)00070-8
40. Turatto M, Facoetti A, Serra G, et al. Visuospatial attention in myopia. Cogn Brain Res. 1999;8(3):369–372. doi:10.1016/S0926-6410(99)00025-7
41. Eriksen CW, St James JD. Visual attention within and around the field of focal attention: a zoom lens model. Percept Psychophys. 1986;40(4):225–240. doi:10.3758/BF03211502
42. Leung TW, Kee CS, De Lestrange E. Corneal shape factors are correlated with ocular aberrations in myopes with or without astigmatism. Invest Ophthalmol Vis Sci. 2018;59(9):2130.
43. Leung TW, De Lestrange-anginieur EAJ, Lam HIC, Kee CS. Ocular higher-order aberrations improve the retinal image quality in astigmatic eyes.
44. De Lestrange-Anginieur E, Kee CS. Optical performance of progressive addition lenses (PALs) with astigmatic prescription. Sci Rep. 2021;11(1):1–14. doi:10.1038/s41598-021-82697-0
45. Liu T. Feature-based attention: effects and control. Curr Opin Psychol. 2019;29:187–192. doi:10.1016/j.copsyc.2019.03.013
46. Leung TW, Li RW, Kee CS. Meridional anisotropy of foveal and peripheral resolution acuity in adults with emmetropia, myopia, and astigmatism. Invest Ophthalmol Vis Sci. 2021;62(10):11. doi:10.1167/iovs.62.10.11
47. Appelle S. Perception and discrimination as a function of stimulus orientation: the” oblique effect” in man and animals. Psychol Bull. 1972;78(4):266. doi:10.1037/h0033117
48. Intriligator J, Cavanagh P. The spatial resolution of visual attention. Cogn Psychol. 2001;43(3):171–216. doi:10.1006/cogp.2001.0755
49. Fuller S, Rodriguez RZ, Carrasco M. Apparent contrast differs across the vertical Meridian: visual and attentional factors. J Vis. 2008;8(1):16. doi:10.1167/8.1.16
50. Mackeben M. Sustained focal attention and peripheral letter recognition. Spat Vis. 1999;12:51–72. doi:10.1163/156856899X00030
51. Bloem IM, Ling S. Attentional modulation interacts with orientation anisotropies in contrast perception. J Vis. 2017;17(11):6. doi:10.1167/17.11.6
52. Blaser E, Pylyshyn ZW, Holcombe AO. Tracking an object through feature space. Nature. 2000;408(6809):196–199. doi:10.1038/35041567
53. Liu T, Larsson J, Carrasco M. Feature-based attention modulates orientation-selective responses in human visual cortex. Neuron. 2007;55(2):313–323. doi:10.1016/j.neuron.2007.06.030
54. Magnussen S, Kurtenbach W. A test for contrast-polarity selectivity in the tilt aftereffect. Perception. 1979;8(5):523–528. doi:10.1068/p080523
55. Solomon JA, Sperling G, Chubb C. The lateral inhibition of perceived contrast is indifferent to on-center/off-center segregation, but specific to orientation. Vision Res. 1993;33:2671–2683. doi:10.1016/0042-6989(93)90227-N
56. Jalie M. Ophthalmic Lenses & Dispensing.
57. Parry NRA, Murray IJ, McKeefry DJ. Reaction time measures of adaptation to chromatic contrast. Vis Neurosci. 2008;25(3):405–410. doi:10.1017/S0952523808080449
58. Plainis S, Parry NR, Sapountzis P, Murray IJ. Orientation and spatial frequency selectivity following adaptation: a reaction time study. Perception. 2015;44(3):301–314. doi:10.1068/p7797
59. Mitchell DE, Wilkinson F. The effect of early astigmatism on the visual resolution of gratings. J Physiol. 1974;243(3):739–756. doi:10.1113/jphysiol.1974.sp010774
60. Vinas M, de Gracia P, Dorronsoro C, et al. Astigmatism impact on visual performance: meridional and adaptational effects. Optom Vis Sci. 2013;90(12):1430–1442. doi:10.1097/OPX.0000000000000063
61. Petersen SE, Posner MI. The attention system of the human brain: 20 years after. Annu Rev Neurosci. 2012;35:73–89. doi:10.1146/annurev-neuro-062111-150525
62. Liu T, Slotnick SD, Serences JT, Yantis S. Cortical mechanisms of feature-based attentional control. Cereb Cortex. 2003;13(12):1334–1343. doi:10.1093/cercor/bhg080
63. Hopfinger JB, Buonocore MH, Mangun GR. The neural mechanisms of top-down attentional control. Nat Neurosci. 2000;3(3):284–291. doi:10.1038/72999
64. Cheng Y, Huang X, Hu YX, et al. Comparison of intrinsic brain activity in individuals with low/moderate myopia versus high myopia revealed by the amplitude of low-frequency fluctuations. Acta Radiologica. 2020;61(4):496–507. doi:10.1177/0284185119867633
65. Zhang XW, Dai RP, Cheng GW, Zhang WH, Long Q. Altered amplitude of low-frequency fluctuations and default mode network connectivity in high myopia: a resting-state fMRI study. Int J Ophthalmol. 2020;13(10):1629. doi:10.18240/ijo.2020.10.18
66. Takeuchi H, Taki Y, Nouchi R, et al. Refractive error is associated with intracranial volume. Sci Rep. 2018;8(1):175. doi:10.1038/s41598-017-18669-0
67. Posner MI, Rothbart MK. Temperament and brain networks of attention. Philos Trans R Soc B. 2018;373(1744):20170254. doi:10.1098/rstb.2017.0254
68. Yamaguchi S, Onoda K. Interaction between emotion and attention systems. Front Neurosci. 2012;6:139. doi:10.3389/fnins.2012.00139
69. Bush G, Luu P, Posner MI. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn Sci. 2000;4(6):215–222. doi:10.1016/S1364-6613(00)01483-2
70. Lanyon RI, Giddings JW. Psychological approaches to myopia: a review. Optom Vis Sci. 1974;51(4):271–281. doi:10.1097/00006324-197404000-00006
71. Łazarczyk JB, Urban B, Konarzewska B, et al. The differences in level of trait anxiety among girls and boys aged 13–17 years with myopia and emmetropia. BMC Ophthalmol. 2016;16(1):1–7. doi:10.1186/s12886-016-0382-2
72. Dias L, Manny RE, Hyman L, Fern K; Comet Group. The relationship between self-esteem of myopic children and ocular and demographic characteristics. Optom Vis Sci. 2002;79(11):688–696. doi:10.1097/00006324-200211000-00006
73. Hopf JM, Boelmans K, Schoenfeld MA, Luck SJ, Heinze HJ. Attention to features precedes attention to locations in visual search: evidence from electromagnetic brain responses in humans. J Neurosci. 2004;24(8):1822–1832. doi:10.1523/JNEUROSCI.3564-03.2004
74. van Ede F, de Lange FP, Maris E. Attentional cues affect accuracy and reaction time via different cognitive and neural processes. J Neurosci. 2012;32(30):10408–10412. doi:10.1523/JNEUROSCI.1337-12.2012
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Evaluation of the EVO/EVO+ Sphere and Toric Visian ICL: Six Month Results from the United States Food and Drug Administration Clinical Trial
Packer M
Clinical Ophthalmology 2022, 16:1541-1553
Published Date: 21 May 2022
Photorefractive Keratectomy Enhancement (PRK) After Small-Incision Lenticule Extraction (SMILE)
Moshirfar M, Parsons MT, Chartrand NA, Lau CK, Stapley S, Bundogji N, Ronquillo YC, Hoopes PC
Clinical Ophthalmology 2022, 16:3033-3042
Published Date: 12 September 2022
A Comprehensive Retrospective Analysis of EVO/EVO+ Implantable Collamer Lens: Evaluating Refractive Outcomes in the Largest Single Center Study of ICL Patients in the United States
Albo C, Nasser T, Szynkarski DT, Nguyen N, Mueller B, Libfraind L, Parkhurst G
Clinical Ophthalmology 2024, 18:69-78
Published Date: 9 January 2024
Three-Month Outcomes of SMILE Pro with the VISUMAX 800 for Myopic Astigmatism in a Large Population
Cung HS, Tran LHT, Tran TN
Clinical Ophthalmology 2025, 19:417-425
Published Date: 7 February 2025
Prevalence of Refractive Errors Among School-Age Children and Adolescents in Saudi Arabia: A Systematic Review and Meta-Analysis
Al Khathami A, Baklola M, Alshehri AA, Alnasser LH, Alshehri RS, Salawi MA, Alwadai RH, Al Ghazwi M, Alanazi AAQ, Alshammari AM, Al-bawah N, Hafez M, Abu Melha A
Clinical Ophthalmology 2025, 19:2117-2132
Published Date: 5 July 2025