Back to Journals » Journal of Inflammation Research » Volume 19
Macrophage Hypoxia Signaling Pathways and Their Roles in Sepsis
Authors Yuan M ![]() , Tian L
, Tian L ![]() , Xu X
, Xu X
Received 26 August 2025
Accepted for publication 24 January 2026
Published 10 February 2026 Volume 2026:19 562619
DOI https://doi.org/10.2147/JIR.S562619
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 6
Editor who approved publication: Dr Anh Ngo
Mu Yuan,1,2 Lixing Tian,1,2,* Xiang Xu1– 3,*
1Department of Stem Cell and Regenerative Medicine, National Key Laboratory of Trauma and Chemical Poisoning, Army Characteristic Medical Center, Army Medical University, Yuzhong District, Chongqing, 400042, People’s Republic of China; 2Central Laboratory, State Key Laboratory of Trauma, Burn, and Combined Injury, Army Characteristic Medical Center, Army Medical University, Yuzhong District, Chongqing, 400042, People’s Republic of China; 3Yunnan Key Laboratory of Stem Cell and Regenerative Medicine, Science and Technology Achievement Incubation Center, Kunming Medical University, Chengguan District, Kunming, 650500, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiang Xu, Email [email protected] Lixing Tian, Email [email protected]
Abstract: Sepsis is a severe systemic inflammatory response syndrome, and macrophages play a crucial role in its pathogenesis. In recent years, more and more attention has been paid to the molecular signaling pathways of hypoxia, especially hypoxia-inducible factor 1-alpha (HIF-1α) and its related metabolic reprogramming, which have shown their importance in regulating the inflammatory response and functional state of macrophages. At present, although a large number of studies have explored the functional changes of macrophages in sepsis, there are still many unsolved mysteries about the specific mechanism and clinical significance of the effect of hypoxia signaling pathway on macrophage function and disease progression. In this review, the metabolic pathways regulated by HIF-1α, the activation of inflammasomes, the reprogramming of cell functions, and the effects of hypoxia-related signaling pathways on macrophage function and the pathological process of sepsis were summarized. Combined with the latest single-cell transcriptomics data and molecular mechanism research, we systematically explore the potential therapeutic targets and intervention strategies, which provide theoretical basis and research direction for the precise treatment of sepsis.
Keywords: sepsis, macrophages, hypoxia, HIF-1α, metabolic reprogramming, inflammasome, therapeutic target
Introduction
Sepsis is a severe systemic inflammatory response syndrome caused by infection, which is characterized by multiple organ dysfunction. Its complex immunopathological mechanism leads to a high clinical mortality.1 The occurrence of sepsis is closely related to the host’s immune response, especially in the early stage of infection, there will be a strong cytokine storm, and often accompanied by immunosuppression in the later stage. The loss of control of this process will directly lead to the death of patients with sepsis.2 Studies have shown that the pathogenesis of sepsis involves a variety of immune cells, including macrophages, which play a central role in the immune response to sepsis.3,4
As an important immune cell of the body, macrophages have a high degree of heterogeneity and plasticity, and can regulate their functions according to the different microenvironment. In the process of sepsis, macrophages are not only involved in the clearance of pathogens but also regulate inflammatory response and maintain immune homeostasis by secreting a variety of cytokines. The polarization of macrophages (such as M1 and M2) plays an important role in the process and outcome of sepsis. M1 macrophages mainly promote inflammation, while M2 macrophages are related to anti-inflammation and tissue repair.5,6 Hypoxic microenvironment is particularly important in sepsis. Many studies have shown that hypoxia-inducible factor-1alpha (HIF-1α), as a key regulator of hypoxia signaling, plays an important role in the regulation of macrophage function. HIF-1α is not only involved in the regulation of metabolic reprogramming of macrophages but also affects their polarization state and inflammatory response. For example, the expression of HIF-1α can promote the polarization of macrophages to M1 type and enhance their inflammatory response ability. In hypoxic environment, HIF-1α can also affect the function of macrophages by regulating metabolic pathways such as glycolysis and fatty acid oxidation.7,8 The role of sepsis hypoxia on macrophage function and polarization and its impact on disease prognosis are controversial, and their research may also provide better therapeutic targets for targeted therapy of sepsis.
In the research of sepsis, to explore the mechanism of macrophage hypoxia signaling pathway not only can help us better understand the pathogenesis mechanism of sepsis, and provides an important theoretical basis for new therapeutic strategy, but also will help us to improve the prognosis of sepsis patients from the metabolism and function of macrophages. The studies in this area are gradually approaching deepening, which constantly sheds light on the influence of hypoxia to macrophage function, and the significance of its function in sepsis, opening a new direction for its clinical application in the future.
Effects of Hypoxic Microenvironment on Macrophages in Sepsis
Mechanism of Hypoxia in Sepsis-Associated Tissues
Sepsis is a complex systemic inflammatory response, which is often accompanied by microcirculation disorders and tissue hypoperfusion, leading to tissue hypoxia.9 In patients with sepsis, the damage of microcirculation is the main cause of tissue hypoperfusion.10 Sepsis can cause microvascular dysfunction, resulting in rheological changes of red blood cells, and then lead to tissue hypoxia and multiple organ dysfunction.11 The microcirculation system of patients with sepsis is often characterized by vasodilatation and hemodynamic instability, which will affect the transport and utilization of oxygen.10,12
In the early stages of sepsis, although cardiac output may increase, the function of microcirculation does not improve. Instead, microvascular obstruction and flow stagnation may occur, which will lead to tissue hypoperfusion and hypoxia.13 For example, one study found that in septic patients, microvascular perfusion was markedly reduced, resulting in inadequate oxygen supply.9 In addition, the inflammatory response in microcirculation and changes in blood composition such as erythrocyte rheology can also aggravate the degree of tissue hypoxia, further leading to organ damage and functional failure.11,14
To overcome this limitation, researchers are seeking new therapeutic strategies, including for example the bloodflow modulators that will likely improve microcirculation and tissue oxygenation. For instance, specific nanoparticles can alleviate sepsis-related hypoperfusion and hypoxia, through enhancing microcirculation perfusion and tissue oxygenation.15 Similarly, the recent progress in microcirculation monitoring enables us to gain insights and to address microcirculation impairment and tissue hypoxia in septic patients in a new way. Early in patients with septic shock, microcirculation monitoring of sublingual microcirculation has demonstrated dysfunction of this system linked to hyperlactatemia and metabolic acidosis but monitoring may help the clinicians detect these changes, albeit in the limited evidence available, in part, because of methodological protocols and of the limited possibilities offered by semi-quantitative analysis methods.16 The development of new methods furthered the efficiency of monitoring microcirculation. A hyperspectral imaging (HSI) along with machine learning may be able to noninvasively monitor skin microcirculation, differentiate healthy and septic patients, and give a self-learned and standardized evaluation, for instance.17
Metabolic disorder caused by sepsis is one of the important factors causing tissue hypoxia. In the state of sepsis, the body’s metabolic demand increases significantly, especially the oxygen demand of immune cells. In this case, although oxygen supply may remain normal in some respects, tissues still experience a state of hypoxia because metabolic demands exceed the capacity to supply.15 Studies have shown that the hypermetabolic state caused by sepsis is closely related to the activation of HIF-1α, which regulates the metabolic adaptation of cells under hypoxic conditions and promotes the occurrence of anaerobic metabolism, which in turn further aggravates intracellular hypoxia (Figure 1).18
 |
Figure 1 Mechanisms of hypoxia formation in sepsis-associated tissues. Sepsis can cause microvascular dysfunction, resulting in rheological changes of red blood cells, and then lead to tissue hypoxia and multiple organ dysfunction. The microcirculation system of patients often shows vasodilation and hemodynamic instability, which will affect the transport and utilization of oxygen. In the early stage of the disease, although cardiac output may increase, the function of microcirculation does not improve. On the other side, microvascular obstruction and flow stagnation may occur, which will lead to tissue hypoperfusion and hypoxia. Metabolic disorder is one of the important factors causing tissue hypoxia in sepsis. In the disease state, the body’s metabolic demand increases significantly, especially the oxygen demand of immune cells. In this condition although oxygen supply may remain normal in some respects, the tissue still experiences a hypoxic state because metabolic demand exceeds the ability to supply it. Created using Figdraw (https://www.figdraw.com/static/index.html#/). |
Another key factor is increased oxidative stress. In patients with sepsis, oxidative stress causes cell damage by generating excessive reactive oxygen species (ROS), affecting the metabolic function of cells.19 This metabolic disorder will not only lead to energy metabolism disorders but also affect the oxygen utilization efficiency of cells and further aggravate tissue hypoxia. For example, inhibition of certain metabolic pathways has been shown to ameliorate hypoxia and reduce organ damage in a mouse model of sepsis.20
The interaction between sepsis-induced metabolic disorders and oxidative stress aggravates tissue hypoxia. The understanding of this process is essential for the development of new therapeutic strategies, especially in the intervention of sepsis-related metabolic disorders and hypoxia state.
Regulation of Macrophage Function and Metabolism by Hypoxia
Under hypoxia, the metabolism and function of macrophages change significantly. These changes are mainly manifested in the dual characteristics of enhanced glycolysis and impaired mitochondrial function. A mouse study showed that hypoxia can lead to the decrease of macrophage proliferation and phagocytosis, accompanied by transcriptome reprogramming and metabolic remodeling. Specifically, under hypoxic conditions, the glycolytic pathway of macrophages was significantly activated. The increased levels of fructose 1, 6-diphosphate, gluconate 6-phosphate and ribose 5-phosphate, the key intermediates of the pentose phosphate pathway, indicate that this pathway is also an important metabolic regulation pathway for macrophages to adapt to hypoxia. Although the enhanced glycolysis provides much needed energy for macrophages, it has also been found that the function of mitochondria is inhibited under hypoxia, which may lead to reduced efficiency of oxidative phosphorylation and further affect cell survival and function.21
HIF-1α plays a key role in this process. In a mouse model constructed with LPS, HIF-1α not only regulates the metabolism of macrophages but also plays an important role in macrophage polarization and inflammatory response.22 Specifically, HIF-1α can promote the polarization of macrophages to M1 type, thereby enhancing their pro-inflammatory function, activating related metabolic pathways and improving cell adaptation to hypoxia.8,22 In addition, mouse cell models suggest that hypoxia also promoted the proportion of M2 macrophages, indicating its regulatory role in anti-inflammatory response.21 This dual polarization enables macrophages to flexibically respond to different physiological demands in hypoxic environment, thus playing an important role in inflammation and repair processes.
Hypoxia in sepsis significantly affects macrophage function and immunometabolic reprogramming by regulating HIF and its downstream metabolic pathways. In animal models of sepsis, hypoxia activates HIF-1α in macrophages, induces the expression of key glycolytic enzymes (such as HK2 and PFKFB3), promotes glycolysis and inhibits mitochondrial oxidative phosphorylation (OXPHOS), thereby enhancing the proinflammatory phenotype (M1 type).7,23,24 Up-regulation of HIF-1α also promotes succinate accumulation and superoxide anion generation, further amplifying the inflammatory response.24,25 Animal experimental studies show that targeted inhibition of SPHK1/S1PR3 axis can block the HIF-1α pathway, reduce the activity of glycolytic enzymes and inhibit M1 polarization, thereby alleviating organ damage in sepsis.23 The increased activity of succinate dehydrogenase (SDH) leads to the accumulation of succinate, activates HIF-1α and promotes the release of inflammatory factors such as IL-1β. Chicoric acid (CA) reduces HIF-1α expression and NLRP3 inflammasome activation by inhibiting SDH and reducing succinate accumulation.25 Human cell model samples demonstrated that the expression of miR-210 is increased in hypoxic environment, which impairs mitochondrial respiration by down-regulating the iron-sulfur cluster assembly enzyme ISCU, forcing macrophages to turn to glycolysis, and exacerbating the inflammatory storm of sepsis.26 A study in mice has shown that histone methyltransferase SETD2 inhibits HIF-1α transcription by catalyse H3K36me3 modification. Down-regulation of SETD2 in sepsis leads to overexpression of HIF-1α, which promotes M1 polarization and glycolysis, and aggravates acute lung injury.24 A study in mice has demonstrated that the acetylation of α-tubulin mediated by the acetyltransferase KAT2A promotes NLRP3 inflammasome assembly. CA can block this pathway by inhibiting KAT2A to alleviate myocardial injury.26 A human-based study has shown that M1 macrophages rely on glycolysis to produce ATP and inflammatory medium (such as IL-6, TNF alpha), while M2 type by fatty acid oxidation for anti-inflammatory function.27 Animal studies have also shown that hypoxia disrupts the M1/M2 balance through metabolic switching and amplifies the immune disorder in sepsis.23,28 Tissue hypoxia caused by microcirculation disorders promotes thrombosis and neutrophil extracellular traps formation (NETosis), forming a vicious cycle of “hypoxia-HIF activation-inflammation-hypoxia”.29–33 Sepsis-induced hypoxia remodels macrophage metabolism through HIF-1α/glycolytic axis, succinate accumulation, and epigenetic modifications, driving pro-inflammatory polarization and organ damage (Figure 2). Targeting the above pathways (such as SPHK1 inhibitors and CA) may become a new strategy for the treatment of sepsis.
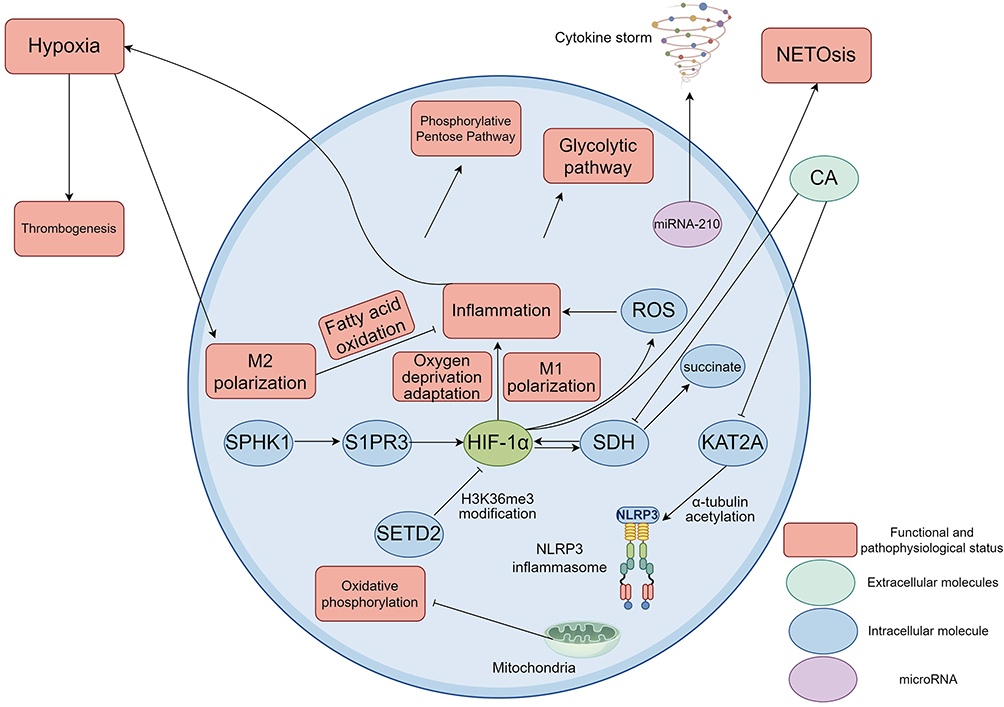 |
Figure 2 A series of changes occur in the function and metabolism of macrophages under hypoxic conditions. Hypoxia activates HIF-1α, which in turn promotes the polarization of macrophages to M1 type, enhances their pro-inflammatory function, and helps cells adapt to hypoxic conditions. Interestingly, hypoxia also increased the proportion of M2-type macrophages. Up-regulation of HIF-1α would promote glycolysis and inhibit OXPHOS of mitochondria, thereby reinforcing the proinflammatory phenotype (M1). In addition, HIF-1α also leads to succinate accumulation and superoxide anion production, further amplifying the inflammatory response. By targeting inhibition of the SPHK1/S1PR3 axis, the HIF-1α pathway can be blocked, the activity of glycolytic enzymes can be reduced, and M1 polarization can be inhibited. On the other hand, increased SDH activity causes succinate accumulation, activates HIF-1α, and promotes the release of inflammatory factors such as IL-1β. CA reduced HIF-1α expression and NLRP3 inflammasome activation by inhibiting SDH and reducing succinate accumulation. In hypoxic environment, miR-210 is up-regulated, which down-regulates the iron-sulfur cluster assembase ISCU, impairs mitochondrial respiration, and forces macrophages to turn to glycolysis for energy supply, thereby exacerbating the inflammatory storm in sepsis. Studies have shown that histone methyltransferase SETD2 inhibits HIF-1α transcription by catalyse H3K36me3 modification. Down-regulation of SETD2 in sepsis leads to overexpression of HIF-1α, which promotes M1 polarization and glycolysis, and aggravates acute lung injury. The acetylation of α-tubulin mediated by the acetyltransferase KAT2A promotes NLRP3 inflammasome assembly. CA can block this pathway by inhibiting KAT2A to alleviate myocardial injury. M1 macrophages rely on glycolysis to produce ATP and inflammatory mediators (such as IL-6 and TNF-α), while M2 macrophages maintain anti-inflammatory function through fatty acid oxidation. Hypoxia disrupts the M1/M2 balance through metabolic switching and amplifies the immune disorder in sepsis. Tissue hypoxia caused by microcirculation disorders promotes thrombosis and NETosis, forming a vicious cycle of “hypoxia-HIF activation-inflammation-hypoxia”. Created using Figdraw (https://www.figdraw.com/static/index.html#/). Abbreviations: HIF-1α, Hypoxia-inducible factor 1 alpha; OXPHOS, Oxidative phosphorylation; SPHK1, Sphingosine kinase 1; S1PR3, Sphingosine 1 phosphate receptor 3; SDH, Succinate dehydrogenase; IL-1β, Interleukin-1 beta; CA, Chicoric acid; NLRP3, NOD-like receptor family pyrin domain containing 3; miRNA-210, microRNA-210; ISCU, Iron-sulfur cluster assembly enzyme; SETD2, SET domain-containing protein 2; KAT2A, K-acetyltransferase 2A; ATP, Adenosine triphosphate; IL-6, Interleukin-6; TNF-α, Tumor necrosis factor-α; NETosis, Neutrophil extracellular trap formation. |
Through the in-depth study of macrophage metabolism under hypoxic conditions, we can better understand its functional changes under various pathological conditions and provide new ideas for future treatment. Especially in diseases such as sepsis, the intervention of macrophage metabolic reprogramming may become an important direction for treatment, which can effectively improve the function of macrophages and the prognosis of patients.34
Role of Hypoxia in Septic Macrophages in Sepsis
In sepsis, the hypoxic state of macrophages plays a key role in disease progression. It mainly regulates the metabolic reprogramming of macrophages, inflammatory response and immune function through the HIF-1α pathway, thus affecting the severity and outcome of sepsis.
Macrophage hypoxia can promote inflammation and aggravation of sepsis through HIF-1α. When sepsis occurs, infection or injury leads to local tissue hypoxia. Macrophages enter a hypoxic state and up-regulate the expression of HIF-1α.8,35 Human clinical samples and animal studies have shown that HIF-1α, as a major transcription factor, drives glycolytic metabolic reprogramming and increases the expression of key glycolytic enzymes such as PFKM (6-phosphofructokinase muscle) and PFKP.35,36 This helps the macrophages to maintain their energy requirements, but also enhances the proinflammatory phenotype, promoting the release of inflammatory cytokines such as IL-1β, IL-6, TNF-α, and IL-27.8,35,37 PFKM expression was significantly upregulated in monocytes from patients with sepsis, which can exacerbate the inflammatory storm; Inhibition of HIF-1α by recombinant thrombomodulin (rTM) reduced PFKM expression and proinflammatory cytokine levels in vitro. In addition, macrophage hypoxia also promotes PFKM expression through methyltransferase-like 3 (METTL3)-mediated m6A modification, further amplifying glycolysis and inflammatory cycles.35 In classically activated macrophages, hypoxia signals enhance the accumulation of methylglyoxal (MG), which acts in concert with HIF-1α to drive the production of TNF and IL-1β, thereby worsening the systemic inflammatory response in sepsis.35 Results from human cell models suggested that the polarization of macrophages is changed in hypoxic condition. Studies have shown that hypoxia can promote the polarization of macrophages to M2 type, thereby reducing the release of pro-inflammatory cytokines and changing the inflammatory microenvironment. This process may be regulated through the p38 signaling pathway, but not solely dependent on hypoxia inducible factor (HIF).38 In addition, experiments with mouse cells showed that hypoxia also promotes the accumulation of macrophages at sites of chronic inflammation by enhancing their proliferative response to CSF-1 and their survival response to TNF.39 Hypoxia also has an important effect on the metabolic remodeling of macrophages. Studies have shown that the glycolytic pathway of macrophages is activated under hypoxia, accompanied by dynamic changes in mRNA transcription and degradation.40 This metabolic remodeling not only supports the survival of macrophages in hypoxic environment but also may affect the course of sepsis by regulating inflammatory response.21 The synergistic effect of hypoxia and inflammatory signals further aggravates the functional changes of macrophages. Experiments with primary human cells showed that hypoxia and inflammatory signals jointly induce transcriptome turnover in macrophages and accelerate mRNA degradation, thereby regulating the expression of inflammation-related genes.40 Mouse cell and model experiments showed that this synergistic effect may be achieved by regulating specific signaling pathways, such as activation of NF-κB and HIF family members.41 Another study confirmed that a mouse model of sepsis showed that HIF-1α stabilization led to increased glycolysis and M1 polarization of macrophages (proinflammatory phenotype), exacerbating organ damage.40
Macrophage hypoxia can intensify sepsis-induced organ dysfunction. Hypoxia of macrophage is not only aggravating inflammation but is also directly causing organ dysfunction. For example, in sepsis-induced acute lung injury (ALI) and acute kidney injury (AKI), macrophage hypoxia contributes to tissue injury and fibrosis via HIF-1α activation. For instance, renal hyperemia mouse models have demonstrated that hypoxia in conjunction with septic factors, such as the Toll-like receptor (TLR) 2 pathway, enhances tubular nephrotoxicity and tubular fibrosis through up-regulation of high mobility group box 1 (HMGB1) and TLR2 expression.42 Likewise, stimulation of hypoxia-related signaling pathways in mouse models of ALI caused the release of proinflammatory factors from macrophages, aggravated lung barrier damage and apoptosis, and HIF-1α knockdown (such as aloin A treatment) significantly ameliorated these injuries.43 What is more, studies in both mouse cells and mouse models stimulated by LPS indicate that Kupffer cells enhance the glycolysis, while forming excessive ROS and inflammatory factors in a state of hypoxic signaling pathway activation, and then stimulate sepsis-related liver dysfunction. Extracellular Vesicles (EVs) derived from MSCs extracellular inhibit the glycolysis via down-regulation of HIF-1α, reduce the inflammation as well as liver function (Figure 3).44
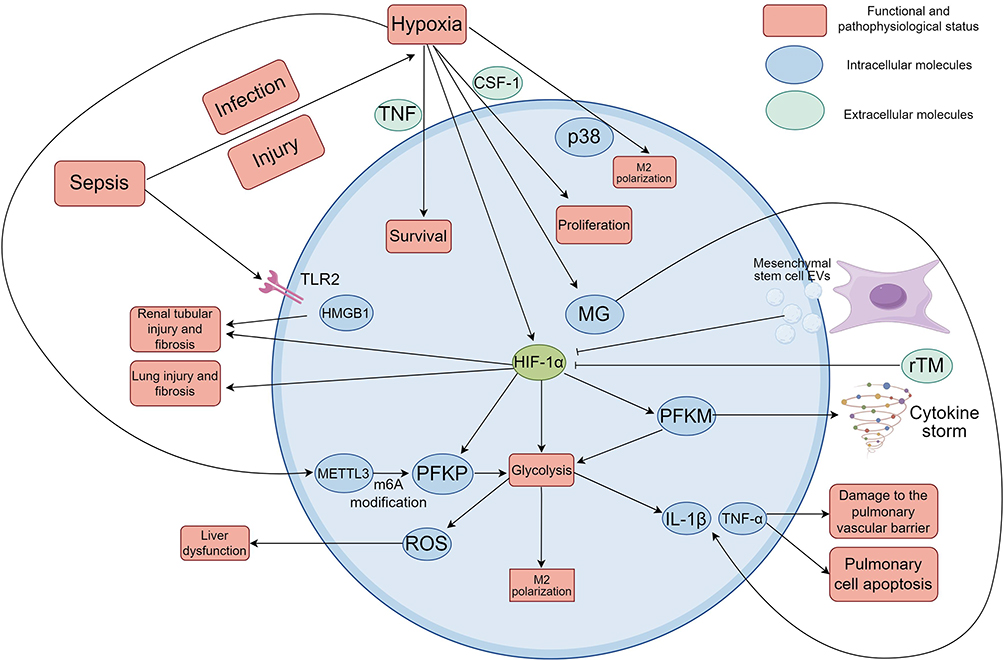 |
Figure 3 Action mechanism of macrophage hypoxia in sepsis. Sepsis resulted from infection or injury will cause local hypoxia of tissue and the macrophage is hypoxic under the action of local hypoxia. HIF-1α was upregulated by macrophage at same time. HIF-1α could drive the glycolytic metabolic reprogramming and upregulate key glycolytic enzyme like PFKM and PFKP. This maintains the energy supply of macrophages while also boosting their proinflammatory phenotype to induce the release of inflammatory cytokines such as TNF-α and IL-1β. Expression of PFKM was strongly up-regulated in patients’ monocytes, which exacerbated the inflammatory storm. rTM can block HIF-1α and reduce expression of PFKM and level of pro-inflammatory factors.Macrophage hypoxia also increases PFKM expression by METTL3-mediated m6A methylation to further boost glycolysis and inflammatory cycles. Hypoxia signaling increased MG accumulation which interacts with HIF-1α to activate the TNF and IL-1β production to exacerbate the systemic inflammatory responses in sepsis. Hypoxia can induce macrophages polarization from M1 type to M2 type, which is able to restrain pro-inflammatory cytokine released and modulate the inflammatory microenvironment. The mechanism may controlled by p38 signal pathway, but not by HIF. Hypoxia also enhanced the macrophages’ proliferative response to CSF-1 and survived response to TNF. HIF-1α stabilization increases the glycolysis and macrophages polarize to M1 type and exacerbates tissue damage. Macrophage hypoxia induces tissue injury and fibrosis in sepsis-associated ALI and AKI via HIF-1αactivation. Sepsis-induced hypoxia triggers activation of TLR2 pathway causing renal tubular injury and fibrosis with an increase in expression of HMGB1and TLR2. In ALI model, hypoxia causes macrophages to induce pro-inflammatory factors inducing lung barrier destruction and cell apoptosis. In hypoxia, the macrophage glycolysis is promoted and the excessive reactive ROS and inflammatory factors are generated, resulting in the induction of liver dysfunction of sepsis. MSC-EVs suppress the glycolysis, inflammation and hepatoprotection by repression of HIF-1α. Created using Figdraw (https://www.figdraw.com/static/index.html#/). Abbreviations: HIF-1α, Hypoxia-inducible factor 1 alpha; PFKM, Phosphofructokinase, Muscle; PFKP, Phosphofructokinase, Platelet; TNF-α, Tumor necrosis factor-α; IL-1β, Interleukin-1 beta; rTM, Recombinant thrombomodulin; METTL3, Methyltransferase-like 3; CSF-1, Colony-stimulating factor 1; MG, Methylglyoxal; ALI, Acute lung injury; AKI, Acute kidney injury; TLR2, Toll like receptor 2; HMGB1, High mobility group box-1 protein; ROS, Reactive oxygen species; MSC, Mesenchymal stem cell; EVs, Extracellular vesicles. |
However, some studies have found that hypoxia and related signaling pathways in sepsis not only promote disease progression but also slowdown the process of the disease. Studies using human cells as well as mouse cells and models have shown that transforming growth factor-β (TGF-β) can induce enhanced glycolysis of macrophages and inhibit the production of pro-inflammatory cytokines, showing a unique anti-inflammatory phenotype. This metabolic reprogramming depends on the mTOR-c-MYC signaling pathway and is accompanied by epigenetic modification changes such as Smad3 activation, which ultimately improves sepsis survival.45 Of note, the similarity between TGF-β-induced metabolic states and hypoxic responses suggests that hypoxic signaling may exert protective effects through similar mechanisms.15 It has been shown that treatment with the hydroxylase inhibitor dimethyloxallyl glycine (DMOG) mimics hypoxic conditions by promoting the polarization of peritoneal macrophages to the M2 (anti-inflammatory phenotype) and upregulating IL-10 production by B1 cells, both of which work together. It markedly reduces levels of proinflammatory cytokines such as TNF-α, leading to improved survival in a mouse model of endotoxic shock.46 In conclusion, hypoxia may play different roles on macrophages in sepsis, which can cause both pro-inflammatory and anti-inflammatory effects of macrophages, but hypoxia under inflammatory conditions causes a comprehensive effect involving multiple signaling pathways. Although most studies support that hypoxia mainly causes pro-inflammatory effects of macrophages, its mechanism still needs to be further studied in order to provide the correct targets for treatment.
Intervention of the macrophage hypoxia pathway has been proved to be a potential target for the treatment of sepsis. Various molecules relieve symptoms by inhibiting HIF-1α or its related factors. Metabolic regulatory molecules such as exogenous acetate can inhibit HIF-1α-mediated glycolysis and promote anti-inflammatory effects by increasing acetyl-coa. Antioxidants such as mitoquinol (MitoQ) can reduce the hypoxic injury of ethanol-exposed macrophages after scavenging ROS, restore the HIF-1α-PFKP axis, and improve phagocytosis and animal survival.36 Studies in mouse cells have shown that natural compounds such as quercetin can reduce macrophage inflammation and apoptosis by inhibiting HIF-1 and NF-κB signaling pathways.44 Cichoric acid inhibits HIF-1α-activated glycolysis by restoring the NAD+/NADH ratio and inactivates NLRP3 inflammasome.47 These interventions were shown to be effective in improving survival and reducing organ damage.36,43,48
In sepsis, although a few studies have shown that the hypoxia state of macrophages plays a protective role, most of the current studies have shown that it mainly drives metabolic reprogramming and inflammation amplification through the HIF-1α axis to aggravate disease progression. Up-regulation of HIF-1α enhances the expression of glycolytic enzymes and the release of proinflammatory cytokines, leading to tissue damage. The literature suggests that inhibition of this pathway can protect organ function and improve prognosis.35,43,48 This suggests that macrophage hypoxia is a key link in the treatment of sepsis, and further research on the mechanism is needed to develop new therapeutic targets with significant effects in the future.
The Regulatory Mechanism of HIF-1α in Septic Macrophages
Expression Regulation and Stability Mechanism of HIF-1α
HIF-1α is a core transcription factor in the response of cells to hypoxia. Its expression and stability play an important role in metabolic adaptation, angiogenesis and tumor growth. The expression of HIF-1α is regulated by a variety of metabolites and signaling pathways. Among them, succinate, as an important metabolic intermediate, can enhance the stability of HIF-1α by inhibiting its hydroxylation, thereby promoting its transcriptional activity and enhancing the adaptability of cells to hypoxia. A study has shown that succinate affects the expression of downstream target genes such as VEGF (vascular endothelial growth factor) by activating the transcription of HIF-1α, which plays an important role in promoting tumor angiogenesis and growth, the study was based on analysis of human tumor samples.49
Another critical process that regulates the stability of HIF-1α is the intracellular oxygen sensing. Normal oxygen concentration in normal cells, HIF-1α is identified by the VHL (Von Hippel-Lindau protein), then HIF-1α is hydroxylated and recognized by ubiquitination protein marks and degraded. In human cells and animal models, it was found that under hypoxia, the hydroxylation process of HIF-1α was inhibited and then accumulated into the nucleus and regulated the expression of corresponding genes. Moreover, the stability and activity of HIF-1α in the cell are closely correlated with intracellular oxygen levels through the PI3K/Akt signaling pathway and mTOR signaling pathway.50,51 The above signaling cascades’ activation stimulate the increase of HIF-1α expression and transcriptional activity, a very important part in the metabolic reprograming and cell survival. Thus, cells control the stability and activity of HIF-1α to meet the metabolic demands of cells in response to environmental perturbations through complex signal transduction.
In certain studies, the human intestinal epithelial cells and animal macrophage models, it was shown that the stability of HIF-1α also had to do with other post-transcriptional modifications, such as acetylation and SUMOylation. These modifications not only influence the stability of HIF-1α but also regulate its interaction with other proteins, thus influencing its function. For example, an acetylation of HIF-1 α can increase its stability, while a SUMOylation may diminish its transcriptional activity.52,53 These regulatory mechanisms may also open up new way to interpret HIF-1αfunctioning during various physiologic and pathologic conditions.
In sepsis (such as sepsis induced by bacterial infection), HIF-1α of macrophages plays a central role in disease progression by regulating inflammatory responses and metabolic pathways.54 Mouse macrophages and animal experiments have shown that the pathological microenvironment of sepsis (such as hypoxia and inflammatory stimulation) triggers the expression of HIF-1α in macrophages, affecting its stability and downstream effects, which in turn leads to inflammatory storm and organ damage.55 This mechanism involves a variety of transcription and translation level regulation, enzymatic modification, and the interaction of signaling pathways.
In mouse cells and animal models of sepsis, Gram-negative bacteria toxins (especially lipopolysaccharide (LPS)) can also induce up-regulation of HIF-1α mRNA and protein in macrophage through the Toll-like receptor 4 (TLR4) pathway, reduce the mRNA level of prohydroxylase and this will stimulate HIF-1α accumulation that induces the expression of proinflammatory factors (eg TNF-α, IL-6).54 Likewise, in mouse cells and in mouse models of sepsis, proinflammatory Gram-positive bacterial toxins (such as lipopolysaccharide) are able to directly promote HIF-1α mRNA transcription, and stimulate this transcription by downregulating the anti-inflammatory transcription factor Kruppel-like transcription factor 2 (KLF2).56 Interleukin-1β (IL-1β) independent inflammation in sepsis microenvironment, pro-inflammatory cytokines can upregulate HIF-1a protein synthesis and nuclear translocation under normoxia by inducing translation of HIF-1a and without impacting on mRNA expression or its hydroxylation status to modulate the inflammatory response of macrophages at infection. Furthermore, in human ovarian cancer cell line OVCAR-3, IL-1β can synergistically with hypoxia further induce up-regulation HIF-1α expression and reinforce its downstream target gene transcription such as adrenomedullin.57 Tissue hypoxia in sepsis is an important trigger which can strongly upregulate the expression of HIF-1a in macrophages through hypoxiastress. HIF-1α mRNA and protein level under hypoxia conditions are up-regulated, and p38 MAPK signaling pathway (phosphorylated MK2) are up-regulated, and it also leads to the up-regulation of inflammatory cytokines (TNF-α) expression. This pathway is shown to be a clinical feature of sepsis in a mouse model that shows hypothermia.58
IL-1β, the most pro-inflammatory factor released in inflammatory microenvironment of the infection could facilitate HIF-1α protein synthesis and nuclear accumulation in normoxia, via its stimulatory effect on translation, independent of HIF-1α mRNA expression and hydroxylation level, to propel macrophage’s inflammatory behavior during an infection. Furthermore, IL-1β synergically with hypoxia can also further promote HIF-1α expression and up-regulate the transcriptions of their downstream target genes, adrenomedullin.57 Hypoxia which is a common event of sepsis is a major activator that can up regulate the expression of HIF-1α of macrophages by a strong hypoxia stress. Under hypoxia condition, mRNA and protein of HIF-1α are elevated, and p38 MAPK signaling pathway (eg, phosphorylated MK2) is activated, and up-regulate inflammatory cytokines (eg, TNF-α) expression which are correlated with clinical symptoms of hypothermia in mouse model of sepsis.58
The stability of HIF-1α is mainly determined by its hydroxylation and degradation pathways. Under normoxia, HIF-1α is easily hydroxylated and ubiquitylated by prolyl hydroxylase (PHD), but sepsis can block this process. For example, IL-1β stabilizes the structure of HIF-1α protein by accelerating its translation (but not its hydroxylation), promoting its accumulation under normoxia.57 Similarly, hypoxia itself reduces hydroxylation and maintains HIF-1α stabilization by inhibiting PHD activity.58 Human GEO database samples and animal cells and models suggest that activating transcription factor 4 (ATF4) is up-regulated in sepsis and can directly bind to the HIF-1α promoter region and inhibit its ubiquitination modification, thereby stabilizing the HIF-1α protein. This promotes glycolysis through mammalian target of rapamycin (mTOR)-dependent pathways (such as up-regulation of hexokinase II) and enhances the expression of inflammatory factors. ATF4 deficiency will reduce the stability of HIF-1α and reduce the inflammation in sepsis.59 SET domain-containing protein 2 (SETD2) catalyzes the trimethylation of histone H3 lysine 36 (H3K36me3), binds to the HIF-1α gene promoter and inhibits its expression, thereby indirectly reducing the stability of HIF-1α. SETD2 deficiency can enhance HIF-1α accumulation, aggravate macrophage M1 polarization and glycolysis, and promote organ damage in sepsis-induced acute lung injury mouse model.24 KLF2 restricts the stability of HIF-1α by down-regulating its mRNA expression in sepsis induced by Gram-positive bacteria. Loss of KLF2 enhances HIF-1α stability and inflammatory response.56 Short-chain fatty acids, such as acetate, increase histone acetylation and reduce HIF-1α stabilization by activating acetyl-coa synthetase 2 in sepsis. This reduces inflammation by inhibiting HIF-1α-dependent glycolysis, but high doses of acetate exposure transiently stabilize HIF-1α.48 In addition, glycolytic intermediates such as methylglyoxal accumulate in response to HIF-1α, further stabilizing their protein and promoting inflammation, and deletion of HIF-1α inhibits this process.8
Overall, septic macrophage’s HIF-1α expression is primarily regulated by TLR4, inflammatory cytokines and hypoxia, whose stability is also primarily regulated by ubiquitination, histone modification and metabolic pathways, all of which can cause inflammation, and these pathways are directly associated with sepsis’ pathological outcome, and their inhibition as HIF-1α did show protective effect in the model.54,55 The insight to these processes may serve as a guide for immunotherapy of sepsis.
HIF-1α-Mediated Gene Expression and Its Function
Studies have found that HIF-1α, a transcription factor, acts as a regulator of related gene expression of inflammation response and metabolism in hypoxia in a mouse model study. Studies have found that in oxygen deficiency, HIF-1α in cytoplasm is stabilized and is transferred into nucleus and combines with the corresponding gene promoters so as to make it express pro-inflammation factors, including IL-6, VEGF and MMP-2.60,61 Besides promoting secretion of the inflammatory mediators, the activation of HIF-1α enhances the capacity of cells to endure environmental stresses. For instance, during liver fibrosis, the up-regulation of HIF-1 α expression is also correlated with the increased levels of IL-6 secretion, resulting in increased activation of the hepatic stellate cells, aggravating inflammatory response and fibrosis.
In addition, HIF-1α also affects the metabolic state of cells by regulating the expression of glycolysis-related enzymes. In many human cancer cells and animal models, HIF-1α is able to activate genes such as glucose transporter (GLUT1), hexokinase (HK2) and lactate dehydrogenase (LDHA), thereby promoting anaerobic glycolysis under hypoxic conditions to meet the energy demand of rapid cell proliferation.60,61 These metabolic alterations not only support the growth of tumors but also may affect the function of other tissues, suggesting the multiple roles of HIF-1α in pathophysiological states.
The activation of HIF-1α is closely related to the activation of the NLRP3 inflammasome. Studies based on mice and mouse myoblast cells have shown that HIF-1α can promote the expression of NLRP3 under hypoxic conditions, thereby enhancing the assembly and activation of the inflammasome, leading to the release of inflammatory cytokines, which plays a key role in the inflammatory response of a variety of diseases. For example, in ischemic diseases, HIF-1α promotes the activation of NLRP3 inflammasome by regulating the expression of SUMO1, leading to increased inflammation and cell death.62,63 This mechanism is not only reflected in cardiovascular diseases but also related to the development of a variety of malignant tumors, suggesting that HIF-1α and its downstream signaling pathways may serve as potential targets for therapeutic intervention. Furthermore, HIF-1α might also contribute to the function of NLRP3 inflammasome by regulating some other genes involving in inflammatory response. For instance, the upregulation of HIF-1α was correlated with the secretion of proinflammatory cytokines including IL-1β which would increase the activation of NLRP3 inflammasome that was found in many chronic inflammatory diseases and suggesting that HIF-1α played a crucial role in regulating inflammation.60,63,64
PFKM is significantly up-regulated in monocytes from patients with sepsis, and rTM reduces its expression by inhibiting the HIF-1α/METTL3 axis, thereby inhibiting glycolysis and inflammation in cells.35 Studies in mouse cells and animal models of sepsis have shown that SPHK1/S1PR3 axis activates HIF-1α-dependent glycolytic enzymes and promotes the transformation of macrophages to a pro-inflammatory phenotype.23 Chicoric acid (CA) inhibits succinate dehydrogenase (SDH)-mediated succinate accumulation, blocks HIF-1α stability, and reduces glycolysis and inflammation.47 Acetic acid reduces HIF-1α expression by inhibiting its expression,48 whereas MDM2 in human triple-negative breast cancer cells enhances HIF-1α production by activating the iNOS-NO-HIF-1α axis.65 HIF-1α mediates the accumulation of methylglyoxal (MG), a byproduct of glycolysis, in macrophages, which promotes the expression of TNF-α and IL-1β.8 HIF-1α drives the transformation of macrophages to M1 phenotype. Animal experimental studies show that Laminarin enhanced the pro-inflammatory function of macrophages through HIF-1α pathway, which was reversed by PF-543, an SPHK1 inhibitor.23 Murine macrophages and model studies have shown that HIF-1α-dependent glycolysis provides energy and substrates for pro-inflammatory cytokine synthesis. FGF21 attenuates sepsis-induced liver injury by blocking this cycle by degrading HIF-1α through autophagy.66 Acetic acid increases the level of acetyl-coa, inhibits the acetylation of NF-κB p65, and indirectly inhibits inflammation through HIF-1α.48 Aralia saponins A (ARA) can inhibit the activation of macrophages by restoring PHD2/HIF-1α signaling axis and improve sepsis-induced lung injury.43 Human and mouse macrophage and mouse model studies have shown that MDM2 degrades the E3 ligase SPSB2, stabilizes iNOS and activates HIF-1α. Knockout of MDM2 reduces obesity-related inflammation but worsens sepsis.65
Interaction Between Metabolic Reprogramming and Hypoxic Signaling in Septic Macrophages
Molecular Mechanisms of Metabolic Reprogramming of Macrophages Under Hypoxic Conditions
The influence of hypoxia signaling of macrophages metabolic reprogramming of sepsis is noteworthy. HIF-1α plays a key role in the metabolic reprogramming of macrophages. Studies have shown that HIF-1α promotes the inflammatory response of macrophages by regulating the glycolytic pathway. In sepsis models, the stabilization of HIF-1α is closely related to the metabolic reprogramming of macrophages, which can be achieved by inhibiting oxidative phosphorylation and increasing glycolysis.15 In addition, mouse models of hepatic ischemia-reperfusion and monocyte-derived macrophages showed that HIF-1α also interacts with other signaling pathways, such as STING signaling pathway, to further regulate macrophage function and inflammatory response.67
Macrophages affect function under hypoxic conditions through metabolic reprogramming. Studies have found that under acute hypoxia, the proliferation and phagocytosis of macrophages are inhibited, and their transcriptome and metabolome are significantly changed. Such changes include the activation of pro-inflammatory responses as well as the enhancement of anti-inflammatory mechanisms, manifested as an increase in M2 macrophages and an increase in anti-inflammatory metabolites such as itaconic acid.21 However, it has been shown that individual hypoxia tolerance alone can affect the basal state of macrophages. Macrophages from hypoxia-sensitive rats showed a higher pro-inflammatory tendency in the non-activated state, and their LPS-activated expression of HIF-1α, TNF-α, IL-1β and CCR7 (markers of M1 type) showed a stronger M1 polarization potential.68 In addition, under combined stress of glucose-oxygen deprivation, human monocytes activate the NLRP3 inflammasome by inhibiting the HMGCR pathway, leading to increased release of IL-1β, a classic M1-associated cytokine, which also supports the possibility that the hypoxic environment may drive a proinflammatory response.69 However, under sepsis conditions, hypoxia promotes the polarization of macrophages to M1 type (proinflammatory), which is mainly achieved by enhancing inflammatory signaling pathways and metabolic reprogramming. Studies have shown that in lipopolysaccharide (LPS) -induced acute lung injury or sepsis and other inflammatory models, the polarization of M1 macrophages can be enhanced through hypoxia-related pathway HIF-1α/PKM2 axis and aggravation of inflammation.70 The result suggests that hypoxia may be biased to drive M1 polarization in the context of inflammation. Hypoxia is common in sepsis, due to inadequate oxygen delivery or increased metabolic demand, which activates transcription factors such as hypoxia-inducible factor-1α (HIF-1α) to regulate macrophage function. For example, murine macrophages and model studies have shown that hypoxia has been shown to enhance lipopolysaccharide (LPS)-induced p38 MAPK activation, which in turn increases the expression and secretion of tumor necrosis factor-alpha (TNF-alpha), a hallmark cytokine of M1-type macrophages, which promotes a proinflammatory response.58 Second, hypoxia drives M1 polarization by modulating metabolic pathways. p38 MAPK and MAPK phosphatase-1 (MKP-1) play a key role in the regulation of glycolysis through the bifunctional glycolysis regulator PFKFB3 in mouse cells and mouse animal models of sepsis. This metabolic reprogramming further reinforces the proinflammatory phenotype.71 In addition, using in vivo mouse models and in vitro cultured murine macrophages, hypoxia has been shown to affect the function of nuclear receptors such as estrogen-related receptor α (ERRα). ERRα deficiency increases Toll-like receptor (TLR)-induced inflammation and impairs mitochondrial respiration while enhancing glycolysis, which mimics the metabolic profile of M1-type macrophages and promotes inflammatory polarization.72 These findings suggest that macrophages under hypoxic conditions adapt to the hypoxic environment through metabolic and transcriptional reprogramming, thereby playing an important role in sepsis. Hypoxia can cause the polarization of macrophages to M2 type, but current studies have shown that hypoxia signaling pathway can promote the polarization of macrophages to M1 type in sepsis, play a pro-inflammatory role, and promote the progress of the disease. Further studies are still needed to explore the mechanism, which may be of great significance for targeted therapy of diseases.
The hypoxia signaling pathway also further affects the function of macrophages by interacting with other signaling pathways. For example, research shows that HIF-1 α interaction with PI3K/AKT/CREB pathways can adjust the polarization state of the macrophages, which affects its function in sepsis (Figure 4). This complex signaling network not only reveals the adaptive mechanism of macrophages under hypoxic conditions but also provides new ideas for the treatment of sepsis.73 As the mechanism of defending against the infection and tissue damage during sepsis pathological process, the up-regulation of pro-protective genes plays an important role for the body. Antimicrobial peptides, as a crucial member of innate immune, can exert rapid antibacterial effect and anti-growth effect on the pathogenic microorganisms. Mouse cell model research demonstrates that transcriptional regulation of antimicrobial peptides through various signalling pathways including hypoxia inducing factor (HIF) signalling pathway is significant.74 The signalling pathways get activated in case of Inflammation and inducement of antimicrobial peptide synthesis. This proves to be significant in enhancing the immune system in case of the host. In addition to that, the activation of tissue repair genes are also of similar importance. Gene up-regulation of factors that assist in healing of injured tissues will reverse damage to tissues and improve prognosis, with the help of fibronectin and matrix metalloproteinases.75 Natural compounds Kaempferol were found to mitigate sepsis-induced ARDS by modulating HIF-1, NF-κB and PI3K-AKT pathways and upregulation of antimicrobial and tissue repair genes.76
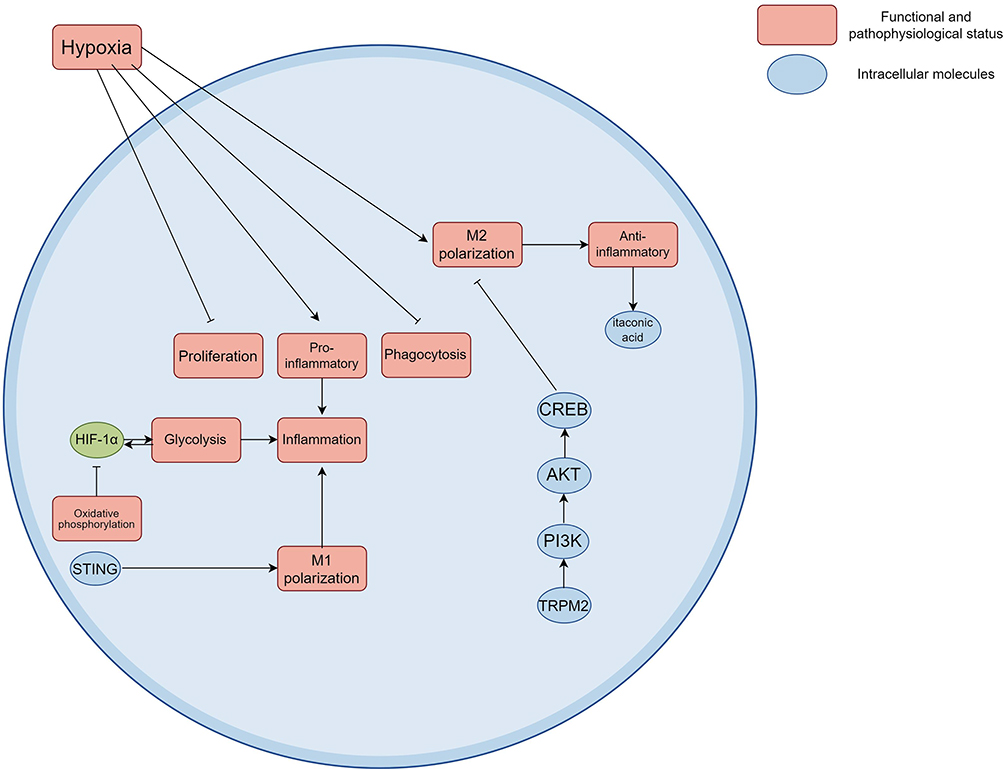 |
Figure 4 Molecular mechanisms underlying the functional reprogramming of macrophages under hypoxic conditions. Under the condition of acute hypoxia, proliferation and phagocytosis of macrophage is restrained, the transcriptome and metabolic group changed significantly at the same time, these changes include promoting the activation of the inflammatory response and inflammatory mechanisms of enhanced, characterized by increased type M2 macrophages and anti-inflammatory metabolites itaconic acid increased. In sepsis model, the stabilization of HIF-1α is closely related to the metabolic reprogramming of macrophages, which can be achieved by inhibiting oxidative phosphorylation and increasing glycolysis. The interaction between HIF-1α and PI3K/AKT/CREB pathway can regulate the polarization state of macrophages. Created using Figdraw (https://www.figdraw.com/static/index.html#/). Abbreviations: HIF-1α, Hypoxia-inducible factor 1 alpha; STING, Stimulator of interferon genes; PI3K, Phosphatidylinositol 3-kinase; AKT, Protein Kinase B; CREB, cAMP response element bound protein; NF-κB, Nuclear factor kappa-B; ARDS, Acute respiratory distress syndrome. |
Hypoxia signaling pathway plays a key role in inflammatory reaction and immune action of macrophages by regulating macrophage metabolism reprogramming during sepsis, which will be a better way for us to clarify the pathophysiological mechanism of sepsis as well as to lay a theoretical foundation for new therapeutic strategy development. More in-depth investigations on the very molecular level of such signaling pathways is the field of future studies, aiming to give an adequate clinical target for the treatment of sepsis.
Metabolic Mechanisms and Functions of Succinate Dehydrogenase (SDH) and Succinate
Succinate dehydrogenase (SDH), as a key enzyme in the tricarboxylic acid cycle (TCA cycle), not only plays an important role in energy metabolism but also plays an indispensable role in metabolic reprogramming and signal transduction of cells. A comprehensive study of gene knockout cell models, human patient-derived primary cells, and mouse disease models has shown that the enhanced activity of SDH leads to the accumulation of succinate, which is closely related to the occurrence of a variety of diseases. For example, defects in SDH often lead to abnormal accumulation of succinate in cells, which triggers abnormal cellular metabolism and eventually leads to the occurrence and development of tumors.77 The accumulation of succinate not only affects energy metabolism but also exacerbates oxidative stress in cells by inducing the production of ROS. The increase of ROS is closely related to cell damage, inflammatory response and tumor progression. Succinate further promotes the formation of tumor microenvironment and the migration of tumor cells through its receptor SUCNR1 (succinate receptor 1)-mediated signal transduction pathway.78,79 Therefore, the enhanced SDH activity and the accumulation of succinate are not only the result of metabolic reprogramming but also one of the response mechanisms of cells to internal and external stresses.
Metabolic reprogramming refers to the process by which cells change their metabolic pathways to adapt to survival in a specific environment or condition. In pathological conditions such as sepsis, the activation of inflammatory response is often accompanied by metabolic reprogramming. Succinate is an intermediate product of TCA cycle, and its intracellular concentration is closely related to the intensity of inflammatory response. A study based on human cells adipose-derived mesenchymal stem cells has shown that succinate can promote the secretion of pro-inflammatory cytokines by activating the SUCNR1 receptor, thereby aggravating the inflammatory response.80 The succinate build-up in the metabolic reprogramming process can alter redox state of cells, induce cells to get energy from glycolysis way and inhibit the activity of mitochondrial oxidative phosphorylation. As the cell’s energy metabolism was affected by redox state in hypoxia, cells’ adaptation to survive in hypoxic condition was enhanced, which was the result of stabilization of HIF-1α. This further suggests that succinate and its metabolites are involved in regulation of cells immune response and metabolic status, which brings new ideas and potential targets for disease of sepsis and related diseases.
Potential Role of microRNAs in the Regulation of Hypoxia Signaling
MicroRNA (miRNA), as a kind of important non-coding small RNA molecules, plays a key role in the biological processes of cells, especially in the cellular response to hypoxia. Studies have shown that hypoxia can induce the expression of specific miRNAs, thereby affecting the function and metabolic reprogramming of macrophages. For example, miR-210 is widely recognized as one of the most important miRNAs under hypoxia. It is upregulated in various cell types and involved in the regulation of cell survival, proliferation and metabolic pathways.81 By regulating the expression of hypoxia-inducible factor HIF-1α, miR-210 can affect multiple signaling pathways related to energy metabolism, thereby regulating the physiological state and function of cells. Specifically, miR-210 promoted the metabolic reprogramming of macrophages by repressing its target genes, orienting them toward glycolytic metabolism rather than mitochondrial oxidative phosphorylation under hypoxic conditions.26 This metabolic reprogramming not only contributes to the survival of macrophages in hypoxic environment but also enhances their adaptability to inflammatory response. Under hypoxic conditions, upregulation of miR-210 is able to inhibit pathways related to mitochondrial respiration and energy production, thereby promoting cell survival and functional maintenance (Table 1). This regulatory mechanism is of great significance for understanding the biological characteristics of macrophages under hypoxic conditions and its role in diseases such as sepsis. In addition, the expression of miR-210 is associated with a variety of disease states, such as up-regulation in pulmonary hypertension and some tumors, showing its potential regulatory role in pathological states. In these cases, miR-210 not only serves as a marker of hypoxia response but also may serve as a potential therapeutic target.81,82 By regulating the expression of miR-210, it is expected to improve the prognosis of hypoxia-related diseases and provide new therapeutic strategies.
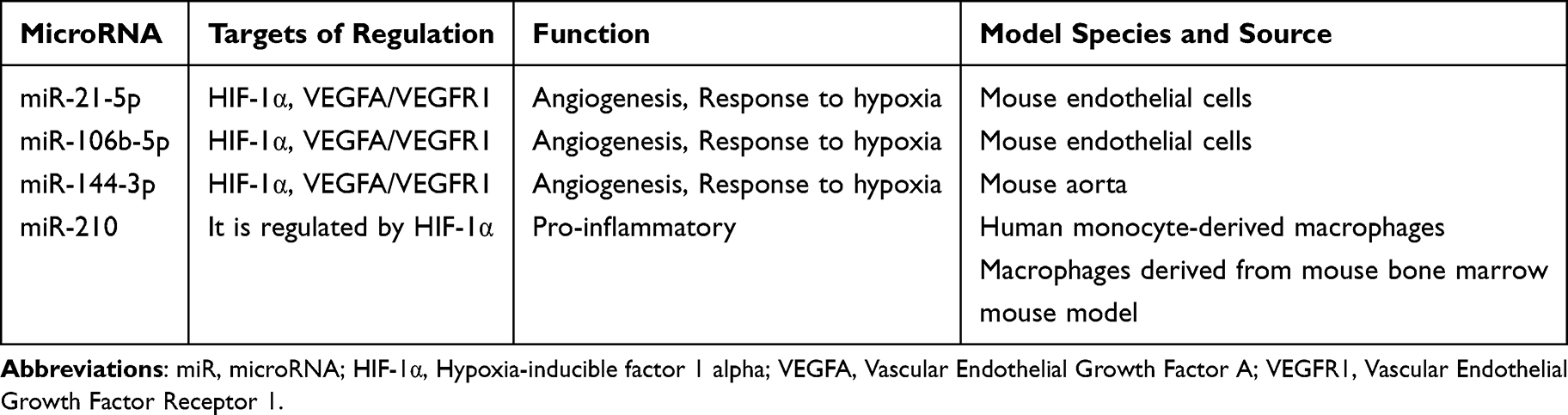 |
Table 1 Potential Role of microRNA in the Regulation of Hypoxic Signaling in Septic Macrophages |
In the inflammatory environment of sepsis (including monocytes/macrophages), specific miRNAs (such as miR-21-5p, miR-106b-5p, miR-144-3p) regulate the HIF-1α signaling pathway and its mediated physiological processes (such as hypoxia adaptation, angiogenesis) by directly targeting the HIF-1α gene or its downstream key effector molecules (such as VEGFA/VEGFR1). Changes in the expression of these miRNAs constitute an important post-transcriptional regulatory mechanism in the pathophysiology of sepsis, suggesting that they can be used as new targets to study and intervene the role of HIF-1α signaling in the dysfunction of macrophages in sepsis.83–85 It has been suggested that the delivery of miRNA inhibitors/mimics by macrophage membrane nanoparticles (MM NPs) is a new therapeutic strategy.86 In general, miRNA plays an important role in the regulation of hypoxic signaling, especially in the hypoxic adaptation, inflammatory state and metabolic reprogramming of macrophages. Future research should continue to explore the mechanism of miRNA in different pathophysiological conditions and its clinical application potential, in order to provide new directions for the treatment of sepsis and other hypoxia-related diseases.
Activation and Regulation of NLRP3 Inflammasome in Septic Macrophages
The Assembly Mechanism of the NLRP3 Inflammasome
The assembly process of NLRP3 inflammasome is a complex cell signal transduction mechanism, which is regulated by many factors. Murine macrophage cells and animal models of sepsis have shown that KAT2A (also known as Gcn5), as a histone acetyltransferase, plays an important role in regulating the assembly of NLRP3. Studies have shown that KAT2A regulates the activation and assembly of NLRP3 inflammasome by acetylating α-tubulin, thereby affecting the stability of microtubules and intracellular signal transmission. Specifically, KAT2A-mediated acetylation of α-tubulin enhances the self-aggregation of NLRP3 and promotes its binding with other inflammasome components such as ASC and caspase-1. This process is a key step in the activation of NLRP3 inflammasome, as only the formation of sufficient aggregates within the cell can effectively initiate the subsequent inflammatory response. Under the condition of septic myocardial injury, the overexpression of KAT2A leads to the increase of α-tubulin acetylation, which in turn promotes the activation of NLRP3 and its downstream signaling pathways. This mechanism is particularly important in a variety of pathological conditions, such as in sepsis and other inflammatory diseases, rise in KAT2A cells may be an adaptive adjustment of environmental stress reaction, adding to the inflammatory reaction. Overall, KAT2A, by adjusting the alpha microtubule protein acetylation, affects NLRP3 inflammatory corpuscle assembly and its subsequent inflammation, which to understand the inflammatory corpuscle regulatory mechanism provides a new perspective.25,47 Hypoxia signaling plays an important role in promoting the activation of NLRP3 inflammasome. Microglial and macrophage studies in mice have shown that hypoxic environment can induce a series of responses in cells, which ultimately lead to the assembly and activation of NLRP3 inflammasome. Under hypoxic conditions, intracellular metabolic changes can lead to the activation of some key molecular signaling pathways that enhance NLRP3 aggregation and activation. Hypoxia affects NLRP3 activation through a variety of mechanisms. Among them, HIF is one of the main regulatory factors, and its expression is significantly increased under hypoxic conditions. HIF can directly regulate the expression of genes related to inflammatory response, thereby promoting the assembly of NLRP3 inflammasome. Specifically, up-regulation of HIF-1α is able to promote the loss of intracellular K+, which is one of the important signals for the activation of NLRP3 inflammasome. In addition, hypoxia can promote intracellular calcium ion flow, enhance the interaction of NLRP3 and NEK7, further promote inflammation of the small body assembly. It has also been found that the increase of reactive oxygen species (ROS) in hypoxic environment also promotes the activation of NLRP3. ROS can not only directly promote NLRP3 accumulation but also indirectly enhance inflammatory response by affecting other intracellular signaling pathways, such as the NF-κB pathway. Therefore, the presence of hypoxia signal not only promotes the activation of NLRP3 inflammasome but also may further aggravate the degree of inflammatory response by affecting the metabolic state of cells, which provides a new understanding of NLRP3 regulation in pathological states such as sepsis.87,88
Contribution of Inflammasome Activation to the Pathology of Sepsis
The activation of inflammasome plays an active role in the pathogenic mechanism of the sepsis, and especially in the release of proinflammatory cytokines. NLRP3 inflammasome as a main inflammasome is reported to be activated by bacterial components like lipopolysaccharide (LPS) in the sepsis, triggering a series of inflammatory responses. Mouse models and in vitro macrophage studies have demonstrated that NLRP3 activation causes the caspase-1 activation with subsequent to pro-inflammatory cytokine maturation and release; IL-1β and interleukin-18 (IL-18) are centrally involved in the acute pro-inflammatory response process in sepsis.89
Under the background of sepsis, over-activated NLRP3 inflammasomes can trigger local inflammatory response not only but also systemic inflammatory response, leading to multiple organ dysfunction. Research revealed that activation of NLRP3 inflammasome is related with the release of cytokines like IL-1β and IL-18, which would on the one hand trigger the inflammatory response by improving the body’s immune function, on the other hand cause tissue damage and cellular death. In mouse model of sepsis, suppression of NLRP3 could greatly decrease IL-1β and IL-18 and inhibit the damage of inflammation and tissues caused by sepsis.90,91 In addition, studies in CLP mice have shown that NLRP3 inflammasome also interacts with other cellular signaling pathways, such as TRAF6-NF-κB signaling pathway, which further promotes the release of pro-inflammatory factors.92 Through the regulation of NLRP3 inflammasome, researchers hope to develop new therapeutic strategies to reduce the inflammatory response and improve the prognosis of sepsis.93 Therefore, in-depth exploration of the mechanism of NLRP3 inflammasome in sepsis can not only help us understand the pathophysiology of sepsis but also provide a new target for potential clinical treatment.
Another important factor in the sepsis pathologic process is apoptosis, and inflammasome activation is an important part. Activation of the inflammasome, NLRP3, contributes both to release of pro-inflammatory cytokines including IL-1β and IL-18 and to directly induced cell death, pyroptosis. Pyroptosis, an inflammatory cell death process characterized by rupturing cell membrane, contents release, was first suggested to be very important to sepsis.89
In mouse model of sepsis, the activation of NLRP3 inflammasome is confirmed to closely associated with the occurrence of a large number of tissue damages, for instance, in the animal cell and model experiments, excessive activation of NLRP3 may lead to damage of the lung and kidney organs, and by these damages they may evoke ALI and AKI. Researchers have found that after suppressing the activation of NLRP3 inflammasome, cell apoptosis and tissues damage were greatly reduced, and they suggested the NLRP3 inflammasome plays an important role in apoptosis and tissues damage of sepsis.90,94
In addition, studies have shown that there is a complex interplay between apoptosis and pyroptosis. In mouse and cellular models of sepsis-induced liver injury, apoptosis may be promoted by the inflammatory response, and the activation of inflammasomes may exacerbate tissue damage by promoting apoptosis. Therefore, the regulation of inflammasome can not only reduce the release of pro-inflammatory factors but also reduce the pathological effects of sepsis by regulating the survival state of cells, providing new ideas for the treatment of patients with sepsis.95
The activation of inflammasomes significantly affects the progression and prognosis of sepsis through the release of proinflammatory cytokines, cell apoptosis and tissue damage in the pathological process of sepsis, which may become a potential therapeutic target.
Effect of Metabolic Reprogramming on NLRP3 Inflammasome Activation
In pathological condition such as sepsis and acute lung injury, macrophages undergo widespread metabolic rewiring, manifested largely by upregulated glycolysis and the suppression of oxidative phosphorylation. This metabolic switch appears to be a principal driver of NLRP3 inflammasome activation. Studies have demonstrated that activation of the NLRP3 inflammasome involves ATP and specific metabolites that are critical in cellular energy metabolism, for instance, increased amounts of glycolytic products such as lactate have been demonstrated to be able to facilitate NLRP3 assembly and activation.96 Furthermore, alteration in glycolytic enzymes’ activity, eg, inhibitions of pyruvate dehydrogenase kinase (PDHK), can considerably downregulate NLRP3 activation, suggesting a close relation between the finely tuned metabolic pathways and inflammasome activation.97 During macrophages encounter pathogen or other inducers, intracellular energy state and metabolite levels are altered rapidly. Such alterations can provide energy supply for the inflammatory response, and they also induce the NLRP3 inflammasome assembly by mediating metabolic signaling pathway.
For instance, in another paper, PDHK inhibitor markedly dampened NLRP3 inflammasome activity, thereby indicating a major part of PDHK in metabolic reprogramming and inflammasome activation.97 Also, overactive glycolytic activity does more than just supplying energy, but additionally promotes inflammation through inflammasome formation, and release proinflammatory cytokine.98 Accordingly, modulations of cellular energy metabolism are critical factors in NLRP3 inflammasome assembly/activation, providing proof for a crosstalk between metabolism reprogramming and immuneresponse.
Metabolic reprogramming not only affects the energy metabolism of cells but also regulates the inflammatory response through metabolites. NLRP3 inflammasome activation is closely related to the accumulation of a variety of metabolites, such as lactate and other metabolic intermediates. These products are not only the products of cellular metabolism but also involved in the regulation of inflammatory response as signaling molecules. Studies have found that the accumulation of lactic acid can promote the activation of NLRP3 inflammasome, which in turn triggers the production of inflammatory factors such as IL-1β.99 In addition, studies have shown that the increase of metabolites such as succinate can promote the activation of NLRP3 by activating HIF-1α, which further emphasizes the importance of metabolites in regulating the inflammatory response (Figure 5).25
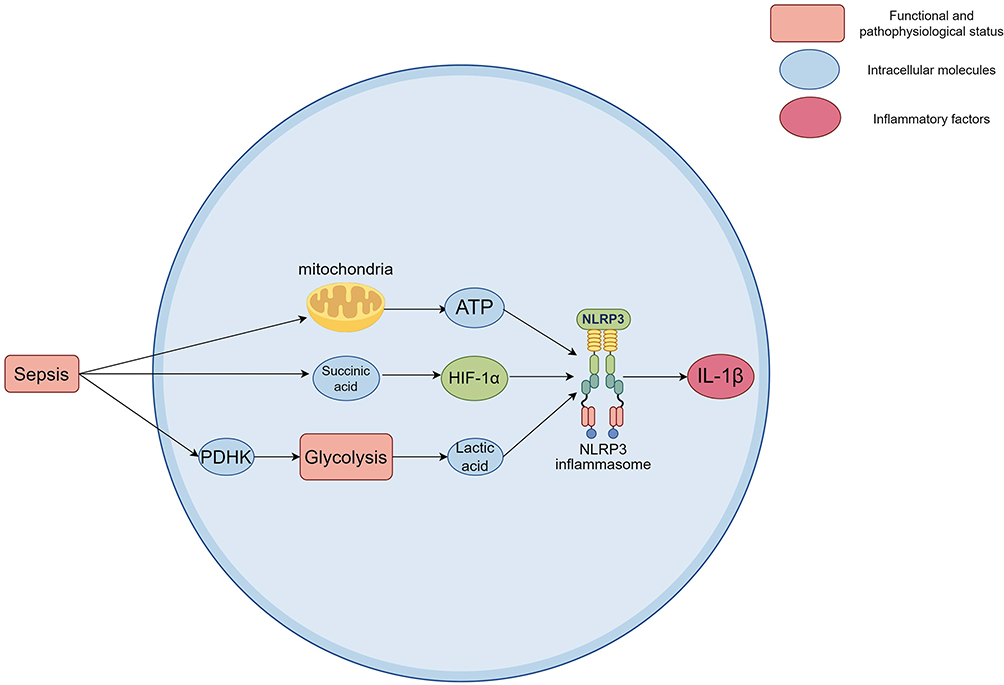 |
Figure 5 Effect of metabolic reprogramming on the activation of the NLRP3 inflammasome. The activation of NLRP3 inflammasome requires a large amount of ATP and the participation of specific metabolites. Increased succinate can promote NLRP3 activation by activating HIF-1α. Increased glycolysis product lactate has been shown to promote NLRP3 assembly and activation. Inhibition of glycolytic PDHK significantly reduces the activation of NLRP3 inflammasome. The accumulation of lactate can promote the activation of NLRP3 inflammasome, which in turn leads to the production of inflammatory factors such as IL-1β. Created using Figdraw (https://www.figdraw.com/static/index.html#/). Abbreviations: NLRP3, NOD-like receptor family pyrin domain containing 3; ATP, Adenosine triphosphate; HIF-1α, Hypoxia-inducible factor 1 alpha; PDHK, pyruvate dehydrogenase kinase; IL-1β, Interleukin-1 beta. |
Meanwhile, there have been numerous studies to explore how we could utilize such metabolites as therapeutic targets. For instance, means to target metabolic reprogramming by eg, administration of anti-inflammatory drugs or inhibition of modulating metabolic pathways, can suppress activation of the NLRP3 inflammasome thereby reducing the connected inflammatory response.100 Such approach of metabolites as signaling molecules reveals the possibility to make use of metabolic reprogramming for clinical management of inflammatory disease. Further exploration of such an interaction mechanism between metabolites and inflammasomes will be of theoretical benefit and applied guidance for new antiinflammatory therapy development.
Therapeutic Targets and Intervention Strategies of Molecular Signaling Pathways in Hypoxia
Application of Hypoxia Metabolic Enzyme Inhibitors in Septic Macrophages
Hypoxia related metabolic enzyme inhibitors in septic macrophages is a hot topic for the purpose of improving the therapeutic effect of sepsis by controlling the metabolism of macrophages. In this way, the related literature of this issue also provides abundant theoretical support. Experiments in humans and animals have shown that macrophages have important roles in sepsis and their excessive activity leads to an excessive release of inflammatory factors, aggravating the pathophysiology. An endogenous metabolite, itaconate, is also proved to have anti-inflammatory effects, where it can block the stimulator of interferon genes (STING) pathway. Yet the hyperactivity glycolytic pathway still keeps the hyperactivated status of the macrophage. A nano particle (LDO) of itaconic acid derivative combined with glycolysis inhibitor thus can effectively reprogram the macrophage to recover the normal balanced polarised status. This approach was proved to be beneficial for aminig cytokine storm, mitigating acute lung injury and dramatically improving survival in a sepsis model.101 Further, studies in mouse models of sepsis-induced acute kidney injury and human renal proximal tubular epithelial cells stimulated by hypoxia combined with LPS have shown that xanthine oxidase (XO) aggravates oxidative stress and inflammation through ROS production in sepsis-associated acute kidney injury, and XO inhibition can protect renal function by improving renal hypoxia. In particular, inhibition by the XO-specific inhibitor febuxstat or inhibition of XO gene expression by shRNA not only alleviated serum urea nitrogen, creatinine, and inflammatory factors, but also alleviated the degree of histological damage, reduced ROS production and macrophage infiltration, thereby significantly enhancing sepsis-induced AKI prognosis (Figure 6). This implies that XO inhibitors may provide a promising therapeutic approach for alleviating cellular hypoxia and reducing inflammation.102 Using hypoxia-related metabolic enzyme inhibitors on septic macrophage will benefit metabolic pathway and inflammatory reaction at certain level, and indicates that these hypotheses are actually playing a very important role in regulating metabolic pathway and controlling inflammatory response, both providing new evidence for the pathological mechanism of sepsis and having a guiding role in clinical treatment of sepsis with novel strategies. The safety and efficacy of such metabolic modulators in clinical practice could be further tested in future research, allowing us to provide better treatments to sepsis patients.
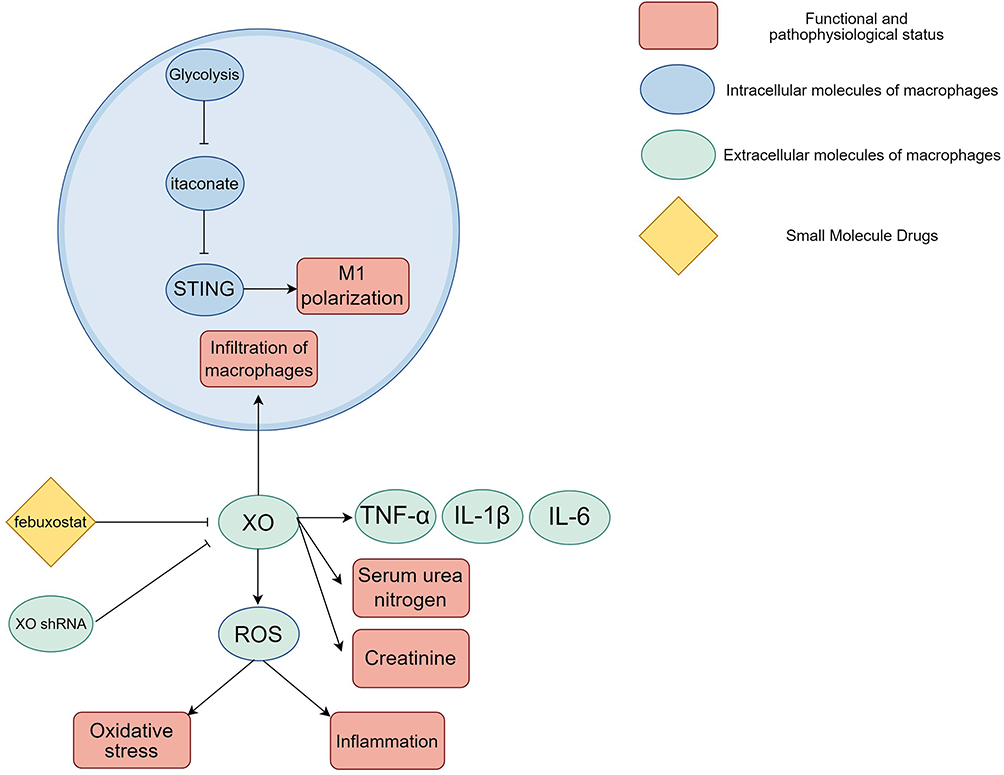 |
Figure 6 Application of hypoxia metabolic enzyme inhibitors in septic macrophages. Itaconate, an endogenous metabolite, has shown anti-inflammatory potential by inhibiting the STING pathway, and its efficacy is limited by an overactive glycolytic pathway that maintains macrophage hyperactivation. XO aggravates oxidative stress and inflammation by producing ROS in sepsis-associated acute kidney injury. Inhibition of XO can protect renal function by improving renal hypoxia. XO specific inhibitor febuxostat or down-regulation of XO gene expression by shRNA significantly improves the prognosis of sepsis-induced acute kidney injury. The levels of serum urea nitrogen, creatinine and inflammatory factors were reduced, and histological damage was also ameliorated, ROS production and macrophage infiltration were reduced. Created using Figdraw (https://www.figdraw.com/static/index.html#/). Abbreviations: STING, Stimulator of interferon genes; XO, Xanthine oxidase; ROS, Reactive oxygen species, shRNA, Short hairpin RNA. |
Regulation Strategies of HIF-1α and Downstream Signaling Pathways
HIF-1α is a key transcription factor involved in the regulation of cellular responses to hypoxia and inflammation. Therefore, the research and development of related drugs to inhibit or activate HIF-1α has received extensive attention. A recent study based on human breast cancer cell models has shown that regulating the expression and activity of HIF-1α can achieve positive effects in the treatment of a variety of diseases. For example, in tumor therapy, the overexpression of HIF-1α is closely related to tumor progression, metastasis and drug resistance. Therefore, inhibitors targeting HIF-1α are considered to be one of the effective strategies for the treatment of tumors. The study has shown that molecular compound plumbagin can effectively inhibit the expression of HIF-1α, thereby reducing the survival rate and the tolerance of tumor cells to treatment.103 In addition, a study based on human gastric cancer cells and animal subcutaneous tumor models has shown that HIF-1α activity can also be reduced by inhibiting the PI3K/Akt/mTOR pathway, which shows good prospects in inhibiting tumor cell proliferation and migration.103,104
The activation of HIF-1α has also been proposed as an important target in chronic inflammatory and autoimmune diseases. In diseases such as rheumatoid arthritis, animal models and cellular experimental studies suggest that the interaction of GRK2 and HIF-1α is thought to be a key mechanism in regulating inflammatory responses, and drugs targeting this pathway may help relieve symptoms.105 Studies in mouse sepsis models, BEAS-2B (human lung epithelial cells) and human umbilical vein endothelial cells (HUVEC) have shown that low molecular weight heparin (LMWH), for example, has shown the potential to reduce inflammatory responses by inhibiting HIF-1α activity.106 These studies suggest that the direction of drug development includes not only direct inhibition of HIF-1α expression but also modulation of its downstream signaling pathways to achieve therapeutic effects.
The regulation of inflammasome activity is significant for the pathogenesis of various diseases. As an important transcriptional factor, HIF-1α participates in regulating the activity of many inflammasomes, mainly in hypoxia and inflammatory situations. For example, research on cell of mouse and animal models of sepsis indicates that the activation of HIF-1α leads to assembly of the NLRP3 inflammasome resulting in discharge of inflammatory factors such as IL-1β and will have significant roles in sepsis and other inflammatory diseases.107 Investigations of this mechanism to control activation of inflammasome by regulation of activity of HIF-1α are being studied to restrict inflammatory response.
In a mouse model of sepsis (induced by cecal ligation and puncture), upregulation of Hmgcs2 promoted the activation of Src, which subsequently activated PI3K/Akt signaling pathway, a common upstream pathway of HIF-1α.108,109 Studies in mouse models of sepsis and in vitro in human cells have shown that the activation resulted in the polarization of macrophages to a protective M2 phenotype, as indicated by an increased proportion of IL-10-positive M2 macrophages, and a reduction in macrophage-mediated inflammatory injury and cardiomyocyte apoptosis.110 In patients with sepsis, free-radical production and lipid peroxidation may exacerbate dysregulation of signaling pathways. This study suggests that antioxidative interventions (eg, polyunsaturated fatty acid supplementation) to mitigate disorders in lipid metabolism may indirectly affect HIF-1α activity, which is known to be regulated by free radicals.111
HIF-1α and downstream signalling molecules can be modulated to represent novel therapeutic targets with particular emphasis on pathogenesis control of inflammation and treating the inflammatory disease conditions, such as sepsis. Future research will aid the understanding of drugs, and small molecule based therapy targeting HIF-1α to act in a clinical setting to efficiently treat disease associated with inflammasome activation.
Future Research Directions and Challenges
Systems Biology of Hypoxic Signaling in Septic Macrophages
The occurrence of sepsis is closely related to the functional changes of macrophages, and macrophages will undergo metabolic reprogramming in the hypoxic environment, which has an important impact on the immune response to sepsis. Recent studies have shown that the metabolic pathway of macrophages plays a key role in sepsis, especially the dynamic regulatory network with HIF-1α and related metabolites. The integrated analysis of multi-omics provides a new perspective to understand this complex biological process. For example, it has been found that under septic conditions, macrophages adapt to hypoxia by enhancing glycolysis and reducing mitochondrial respiration.112 In addition, up-regulation of HIF-1α not only promotes glycolysis of macrophages but also affects their cellular function and inflammatory response.26 The construction of dynamic regulatory networks allowed us to identify key genes and signaling pathways, thereby providing potential targets for future therapeutic strategies.
Reprogramming of the cellular metabolism becomes particularly relevant from this point of view, given that, besides influencing the polarization state of the macrophage itself (eg, in switching between M1 and M2 phenotypes), it is also directly linked with the cytokines which they generate. For instance, when in hypoxic conditions, macrophages will resort a metabolic pathway based mainly in glycolysis, down-regulating mitochondrial function, and will cause overwhelming secretion of inflammatory factors, which is an important pathologic mechanism of sepsis.35 Hence, through integrating transcriptomics, metabolomics and proteomics data, we can have a better understanding of the metabolic change of septic macrophages in hypoxic conditions and the effect on immune response.
Transcriptome analysis based on a human clinical database reveals a central role of hypoxia-inducible factor-1α (HIF-1α) signaling pathway, which is continuously up-regulated in sepsis. Studies based on human monocyte sepsis models have shown that impaired monocyte inflammation and antigen presentation by inhibiting IKK phosphorylation leads to immunosuppression.107,113 This research result integrates human clinical databases on sepsis, bioinformatics analysis from public databases, experiments with mouse models, and mechanism studies of in vitro cell models. Through metabolomics studies, it has further revealed that the hypoxia-glycolysis-lactic acid (HGL) axis drives immunometabolic reprogramming. In this process, changes in the expression of genes such as HK3 and PGK1 affect the functional heterogeneity of neutrophils through lactic acid modification.114 Animal models and animal cell studies demonstrated that the regulation of HIF-1α on downstream targets such as VEGF-A was verified by proteomics and confirmed its association with oxidative stress and mitochondrial damage.107,115 Pharmacological models provide direct evidence for targeted therapies: Tempol, a superoxide dismutase mimetic, alleviates medullary hypoxia with renal-arterial infusion in early sepsis but is ineffective in late-stage models, highlighting the effect of timing on targeting strategies.116 Anti-drag polymers (DRPs) have demonstrated the therapeutic potential of rheological regulation in a mouse model of sepsis by reducing microthrombosis and tissue hypoxia by improving microcirculation hemodynamics.13 Multi-omics integration strategies, such as transcriptome screening of HIF-1α target genes, metabolome profiling of HGL networks, and proteomics validation of pathway proteins, combined with pharmacological validation, not only will they clarify the central mechanism of HIF-1α-HGL axis and so forth in sepsis hypoxia, but also provide a theoretical basis for personalized treatment, such as molecular subtyping based immunoregulation.114 However, the current research still needs to deepen the integration and clinical translation of multi-omics data to optimize the precision and timeliness of targeted therapy.
In the pathological process of sepsis, macrophages are not only affected by their own hypoxia state but also regulated by the surrounding microenvironment. The interaction between macrophages and other immune cells plays a crucial role in sepsis. For example, studies have shown that macrophages regulate systemic inflammatory response and immune tolerance in sepsis by interacting with other cells such as T cells, B cells and fibroblasts.117 The communication between these cells not only affects the activation state of macrophages but also changes their metabolic pathways, which further affects the progression of sepsis.
In addition, factors such as cytokines, metabolites and oxygen concentration in the microenvironment can also significantly affect the function of macrophages. For example, HIF-1α plays a key role in cytokine expression and metabolic reprogramming.8 In sepsis models, up-regulation of HIF-1α expression is closely associated with macrophage polarization, especially in M1 macrophages, whose enhanced glycolytic capacity is directly related to the production of inflammatory factors. By regulating these factors in the microenvironment, it can provide new ideas for the treatment of sepsis. For example, small molecule drugs can be used to target HIF-1α or related metabolic pathways, thereby improving macrophage function and reducing inflammatory response.24
Systems biology study on macrophage hypoxia signaling in sepsis has illustrated complex role of macrophages in sepsis from multi-omics integration analysis and deep exploration of cell-to-cell interaction, which not only presents a new view to our immunology insight on sepsis but also offers a potential target for therapy in the future.
Clinical Translation and Precision Treatment Strategies
Despite the central role of the hypoxia-inducible factor HIF-1α signaling pathway in the immunometabolic reprogramming of macrophages in sepsis,35,47 its clinical translation still faces multiple limitations.
The published intervention strategies, which target this pathway, have the following problems: Not target-specific: HIF-1α is generally engaged in various cellular physiological processes, and targeting it systematically may lead to off-target.15,118 Spatial-temporal constraints of the metabolic targeting: Studies in cellular and animal experimental models in mice have shown that the macrophage glycolytic reprogramming is dynamic and spatially dependent. In the temporal dimension, PF-543, an inhibitor of SPHK1/S1PR3 axis, inhibited glycolytic signals such as HIF-1α, HK2, and PFKFB3 at the early stage of sepsis, and dynamically balanced macrophage polarization.23 Animal model experiments and in vitro cell experiments showed that alveolar type II epithelial cells deliver exosomal lncRNA Rmrp to alveolar macrophages to inhibit PFKFB3 expression and glycolysis, leading to immune tolerance, which explains the organ-specific mechanism of secondary pneumonia after sepsis.119 Limitations of the transformation model: It is mainly dependent on the LPS or CLP animal model and lacks diversity of the human sepsis.120 Single-cell sequencing indicated that macrophages in sepsis are heterogeneous, but their current targeting strategies have not fully recognized diversity.121 Complexity of regulatory mechanism: LPS stimulation can promote HIF-1α expression via p38 MAPK, while lead to TNF-α secretion enhancement; SB203580, p38 inhibitor, did not inhibit TNF-α expression under hypoxia. This indicates that hypoxia could enhance the role of HIF-1α in other ways. In this study, we employed the mouse macrophage cell RAW264.7 and the mouse model of LPS-induced endotoxemia.58 Lack of clinical verification: Although the regulatory role of HIF-1α in septic macrophages has been confirmed in the literature, the disadvantage lies in the fact that the above literature is all animal and cellular experiments, and there are no clinical experiments that can be used for clinical observations of its effectiveness and safety. In the future, clinical trials should be selected for studies.
Tissue-specific delivery systems, such as macrophage-targeting nanoparticles, have been developed to reduce the risk of systemic inhibition of HIF-1α122. Combined with single-cell multi-omics technology (such as scRNA-seq9), the immuno-typing of patients with sepsis can be performed to develop precise intervention strategies based on macrophage metabolic phenotype. Exploring the upstream regulatory nodes of HIF-1α (such as PHD210) rather than directly targeting HIF-1α itself may provide a more controllable therapeutic window. Promoting organoid and human primary macrophage model studies to verify the dose-response relationship of compounds (such as acetate 5 and MitoQ8) in human cells; Biomarkdriven clinical trials are designed to integrate HIF-1α activity (such as PET imaging), serum metabolites (MG2) and patient stratification to achieve true precision treatment.
Conclusion
Severe critical illness and pathogenesis of sepsis are still under investigation. Macrophage as the significant element of immune system is closely related to the pathogenesis and process in the occurrence of sepsis. The results of this review show that macrophage hypoxia is mainly involved in the progress of sepsis, but its mechanism of promoting macrophages to M2 polarization and anti-inflammatory protection still needs to be further studied to find more precise targets, so as to play a role in comprehensive treatment. Hypoxia molecular signaling pathway in macrophage, especially HIF-1α-mediated metabolic reprogramming and inflammation regulation are considered as one main breakthrough to reveal the pathogenesis mechanism of sepsis. HIF-1α not only participates in the macrophage energy metabolism regulation, but it also has an impact on the inflammatory macrophage response, which consequently influences the sepsis clinical presentation. Metabolic enzymes including SDH and its product succinate and these endogenous signaling pathways not only provide support for the survival and function of macrophages but also may become new targets for intervention in the process of sepsis. Meanwhile, microRNA and exosome-mediated signaling mechanisms brought new clues of macrophage regulation. Future research should focus on using multi-omics combined with machine learning technology to reveal the complexity and diversity of macrophage hypoxia regulation, in order to provide more effective strategies for the prevention and treatment of sepsis.
Data Sharing Statement
Data sharing is not applicable to this article as no data were created or analysed in this study.
Author Contributions
All authors took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work. M.Y. was mainly responsible for the conceptualization, investigation, writing – original draft and writing-review and editing of the manuscript; L.T. contributed primarily to the conceptualization, writing – review and editing and funding acquisition of the manuscript. X.X. mainly contributed to the conceptualization, supervision, writing – review and editing and project administration of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China No.82402548.
Disclosure
The authors state that they have no conflicts of interest.
References
1. Ding H, Yang P, Zhang XH, et al. Efficacy of pretreatment with lycium barbarum polysaccharide in various doses in influencing splenic immunity and prognosis of sepsis in rats. Evid Based Complement Alternat Med. 2022;2022:9508603. doi:10.1155/2022/9508603
2. Liu YC, Shou ST, Chai YF. Immune checkpoints in sepsis: new hopes and challenges. Int Rev Immunol. 2022;41:207–24. doi:10.1080/08830185.2021.1884247
3. Ma C, Liu H, Yang S, et al. The emerging roles and therapeutic potential of B cells in sepsis. Front Pharmacol. 2022;13:1034667. doi:10.3389/fphar.2022.1034667
4. van der Poll T, Shankar-Hari M, Wiersinga WJ, van der Poll T. The immunology of sepsis. Immunity. 2021;54:2450–2464. doi:10.1016/j.immuni.2021.10.012
5. Hou FF, Mi JH, Wang Q, et al. Macrophage polarization in sepsis: emerging role and clinical application prospect. Int Immunopharmacol. 2025;144:113715. doi:10.1016/j.intimp.2024.113715
6. Chen X, Liu Y, Gao Y, et al. The roles of macrophage polarization in the host immune response to sepsis. Int Immunopharmacol. 2021;96:107791. doi:10.1016/j.intimp.2021.107791
7. Song C, Xu J, Gao C, et al. Nanomaterials targeting macrophages in sepsis: a promising approach for sepsis management. Front Immunol. 2022;13:1026173. doi:10.3389/fimmu.2022.1026173
8. Prantner D, Vogel SN. Intracellular methylglyoxal accumulation in classically activated mouse macrophages is mediated by HIF-1α. J Leukoc Biol. 2025;117.
9. Peng X, Zheng F, Zhu B, et al. Research progress of diagnostic and therapeutic value of carbon dioxide-derived indicators in patients with sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue. 2024;36:435–440. doi:10.3760/cma.j.cn121430-20240122-00074
10. Hawiger J, Veach RA, Zienkiewicz J. New paradigms in sepsis: from prevention to protection of failing microcirculation. J Thromb Haemost. 2015;13:1743–1756. doi:10.1111/jth.13061
11. Pantos C, Apostolaki V, Kokkinos L, et al. Acute triiodothyronine treatment and red blood cell sedimentation rate (ESR) in critically ill COVID-19 patients: a novel association? Clin Hemorheol Microcirc. 2021;79:485–488. doi:10.3233/CH-211215
12. Orbegozo D, Su F, Santacruz C, et al. Effects of different crystalloid solutions on hemodynamics, peripheral perfusion, and the microcirculation in experimental abdominal sepsis. Anesthesiology. 2016;125:744–754. doi:10.1097/ALN.0000000000001273
13. DE B, OA B, Trofimov AO, et al. Alleviation of post-sepsis ischaemia by drag-reducing polymers. Adv Exp Med Biol. 2024;1463:203–207. doi:10.1007/978-3-031-67458-7_34
14. Østergaard L. SARS CoV-2 related microvascular damage and symptoms during and after COVID-19: consequences of capillary transit-time changes, tissue hypoxia and inflammation. Physiol Rep. 2021;9e14726.
15. Vanderhaeghen T, Vandewalle J, Libert C. Hypoxia-inducible factors in metabolic reprogramming during sepsis. FEBS J. 2020;287:1478–1495. doi:10.1111/febs.15222
16. Sharawy N, Mahrous R, Whynot S, et al. Clinical relevance of early sublingual microcirculation monitoring in septic shock patients. Clin Hemorheol Microcirc. 2018;68:347–359. doi:10.3233/CH-170244
17. Kohnke J, Pattberg K, Nensa F, et al. A proof of concept for microcirculation monitoring using machine learning based hyperspectral imaging in critically ill patients: a monocentric observational study. Crit Care. 2024;28:230. doi:10.1186/s13054-024-05023-w
18. Ruan H, Li YZ, Zhang Q, et al. Identification and clinical validation of hypoxia-inducible factor 1α protein as the potential biomarker in patients with sepsis. Shock. 2023;59:855–863. doi:10.1097/SHK.0000000000002122
19. Jensen IJ, McGonagill PW, Berton RR, et al. Prolonged reactive oxygen species production following septic insult. Immunohorizons. 2021;5:477–488. doi:10.4049/immunohorizons.2100027
20. Yajnik V, Maarouf R. Sepsis and the microcirculation: the impact on outcomes. Curr Opin Anaesthesiol. 2022;35:230–235. doi:10.1097/ACO.0000000000001098
21. Sun B, Long Y, Xu G, et al. Acute hypoxia modulate macrophage phenotype accompanied with transcriptome re-programming and metabolic re-modeling. Front Immunol. 2025;16:1534009. doi:10.3389/fimmu.2025.1534009
22. Qiu B, Yuan P, Du X, et al. Hypoxia inducible factor-1α is an important regulator of macrophage biology. Heliyon. 2023;9.
23. Wang D, Bi X, Zhao L, et al. Targeting SphK1/S1PR3 axis ameliorates sepsis-induced multiple organ injury via orchestration of macrophage polarization and glycolysis. Biochim Biophys Acta Mol Cell Res. 2025;1872:119877. doi:10.1016/j.bbamcr.2024.119877
24. Meng Y, Kong KW, Chang YQ, et al. Histone methyltransferase SETD2 inhibits M1 macrophage polarization and glycolysis by suppressing HIF-1α in sepsis-induced acute lung injury. Med Microbiol Immunol. 2023;212:369–379. doi:10.1007/s00430-023-00778-5
25. Sun HJ, Zheng GL, Wang ZC, et al. Chicoric acid ameliorates sepsis-induced cardiomyopathy via regulating macrophage metabolism reprogramming. Phytomedicine. 2024;123:155175. doi:10.1016/j.phymed.2023.155175
26. Virga F, Cappellesso F, Stijlemans B, et al. Macrophage miR-210 induction and metabolic reprogramming in response to pathogen interaction boost life-threatening inflammation. Sci Adv. 2021;7.
27. Castoldi A, Naffah DSC, Câmara NO, et al. The macrophage switch in obesity development. Front Immunol. 2015;6:637. doi:10.3389/fimmu.2015.00637
28. Na YR, Je S, Seok SH. Metabolic features of macrophages in inflammatory diseases and cancer. Cancer Lett. 2018;413:46–58. doi:10.1016/j.canlet.2017.10.044
29. McInturff AM, Cody MJ, Elliott EA, et al. Mammalian target of rapamycin regulates neutrophil extracellular trap formation via induction of hypoxia-inducible factor 1 α. Blood. 2012;120:3118–3125. doi:10.1182/blood-2012-01-405993
30. von Brühl ML, Stark K, Steinhart A, et al. Monocytes, neutrophils, and platelets cooperate to initiate and propagate venous thrombosis in mice in vivo. J Exp Med. 2012;209:819–835. doi:10.1084/jem.20112322
31. Eltzschig HK, Carmeliet P. Hypoxia and inflammation. N Engl J Med. 2011;364:656–665. doi:10.1056/NEJMra0910283
32. Middleton EA, He XY, Denorme F, et al. Neutrophil extracellular traps contribute to immunothrombosis in COVID-19 acute respiratory distress syndrome. Blood. 2020;136:1169–1179. doi:10.1182/blood.2020007008
33. Levi M, van der Poll T. Coagulation and sepsis. Thromb Res. 2017;149:38–44. doi:10.1016/j.thromres.2016.11.007
34. Emam M, Tabatabaei S, Sargolzaei M, et al. Response to oxidative burst-induced hypoxia is associated with macrophage inflammatory profiles as revealed by cellular genome-wide association. Front Immunol. 2021;12:688503. doi:10.3389/fimmu.2021.688503
35. Yao C, Zhu H, Ji B, et al. rTM reprograms macrophages via the HIF-1α/METTL3/PFKM axis to protect mice against sepsis. Cell Mol Life Sci. 2024;81:456. doi:10.1007/s00018-024-05489-5
36. Pant BD, Ahuja A, Roychowdhury S, et al. Mitoquinol improves phagocytosis and glycolysis in ethanol-exposed macrophages via HIF-1α-PFKP axis. J Immunol. 2025;214:1754–1772. doi:10.1093/jimmun/vkaf078
37. Evans CE. Hypoxia and HIF activation as a possible link between sepsis and thrombosis. Thromb J. 2019;17:16. doi:10.1186/s12959-019-0205-9
38. Ke X, Chen C, Song Y, et al. Hypoxia modifies the polarization of macrophages and their inflammatory microenvironment, and inhibits malignant behavior in cancer cells. Oncol Lett. 2019;18:5871–5878. doi:10.3892/ol.2019.10956
39. Hamilton JA, Lacey DC, Turner A, et al. Hypoxia enhances the proliferative response of macrophages to CSF-1 and their pro-survival response to TNF. PLoS One. 2012;7e45853.
40. Courvan E, Parker RR. Hypoxia and inflammation induce synergistic transcriptome turnover in macrophages. Cell Rep. 2024;43:114452. doi:10.1016/j.celrep.2024.114452
41. Riboldi E, Porta C, Morlacchi S, et al. Hypoxia-mediated regulation of macrophage functions in pathophysiology. Int Immunol. 2013;25:67–75. doi:10.1093/intimm/dxs110
42. Nakamura I, Umehara M, Yagi-Tomita A, et al. Toll like receptor 2 mediated exacerbation of sepsis associated acute kidney injury by renal congestion in mice. Sci Rep. 2025;15:22081. doi:10.1038/s41598-025-05878-1
43. Ma J, Wang J, Wang J, et al. Araloside A alleviates sepsis-induced acute lung injury via PHD2/HIF-1α in macrophages. Phytomedicine. 2024;135:156089. doi:10.1016/j.phymed.2024.156089
44. Pei L, Jin Z, Wang J, et al. MSCs-derived extracellular vesicles alleviate sepsis-associated liver dysfunction by inhibiting macrophage glycolysis-mediated inflammatory response. Int Immunopharmacol. 2024;128:111575. doi:10.1016/j.intimp.2024.111575
45. Xiao X, Ye M, Xu Y, et al. Role of hypoxia-inducible factor in immunometabolism of sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue. 2020;32:382–384. doi:10.3760/cma.j.cn121430-20200226-00238
46. Hams E, Saunders SP, Cummins EP, et al. The hydroxylase inhibitor dimethyloxallyl glycine attenuates endotoxic shock via alternative activation of macrophages and IL-10 production by B1 cells. Shock. 2011;36:295–302. doi:10.1097/SHK.0b013e318225ad7e
47. Zhu XX, Zheng GL, Lu QB, et al. Cichoric acid ameliorates sepsis-induced acute kidney injury by inhibiting M1 macrophage polarization. Eur J Pharmacol. 2024;976:176696. doi:10.1016/j.ejphar.2024.176696
48. Li N, Gong Y, Zhu Y, et al. Exogenous acetate attenuates inflammatory responses through HIF-1α-dependent glycolysis regulation in macrophage. Cell Mol Life Sci. 2024;82:21. doi:10.1007/s00018-024-05521-8
49. Wang YM, Gu ML, Ji F. Succinate dehydrogenase-deficient gastrointestinal stromal tumors. World J Gastroenterol. 2015;21:2303–2314. doi:10.3748/wjg.v21.i8.2303
50. Abdi SM, Shiri A, Aghapour M, et al. Normoxic HIF-1α stabilization caused by local inflammatory factors and its consequences in human coronary artery endothelial cells. Cells. 2022;12:11. doi:10.3390/cells12010011
51. Thomas C, Leleu D, Masson D. Cholesterol and HIF-1α: dangerous liaisons in atherosclerosis. Front Immunol. 2022;13:868958. doi:10.3389/fimmu.2022.868958
52. Malkov MI, Flood D, Taylor CT. SUMOylation indirectly suppresses activity of the HIF-1α pathway in intestinal epithelial cells. J Biol Chem. 2023;299:105280. doi:10.1016/j.jbc.2023.105280
53. Chen Q, Cui K, Zhao Z, et al. LPS stimulation stabilizes HIF-1α by enhancing HIF-1α acetylation via the PARP1-SIRT1 and ACLY-Tip60 pathways in macrophages. FASEB J. 2022;36e22418.
54. Peyssonnaux C, Cejudo-Martin P, Doedens A, et al. Cutting edge: essential role of hypoxia inducible factor-1alpha in development of lipopolysaccharide-induced sepsis. J Immunol. 2007;178:7516–7519. doi:10.4049/jimmunol.178.12.7516
55. Zinkernagel AS, Johnson RS, Nizet V. Hypoxia inducible factor (HIF) function in innate immunity and infection. J Mol Med. 2007;85:1339–1346. doi:10.1007/s00109-007-0282-2
56. Mahabeleshwar GH, Qureshi MA, Takami Y, et al. A myeloid hypoxia-inducible factor 1α-Krüppel-like factor 2 pathway regulates gram-positive endotoxin-mediated sepsis. J Biol Chem. 2012;287:1448–1457. doi:10.1074/jbc.M111.312702
57. Frede S, Freitag P, Otto T, et al. The proinflammatory cytokine interleukin 1beta and hypoxia cooperatively induce the expression of adrenomedullin in ovarian carcinoma cells through hypoxia inducible factor 1 activation. Cancer Res. 2005;65:4690–4697. doi:10.1158/0008-5472.CAN-04-3877
58. FQ L, Liu Y, Lui VC, et al. Hypoxia modulates lipopolysaccharide induced TNF-alpha expression in murine macrophages. Exp Cell Res. 2008;314:1327–1336. doi:10.1016/j.yexcr.2008.01.007
59. Liu T, Wen Z, Shao L, et al. ATF4 knockdown in macrophage impairs glycolysis and mediates immune tolerance by targeting HK2 and HIF-1α ubiquitination in sepsis. Clin Immunol. 2023;254:109698. doi:10.1016/j.clim.2023.109698
60. Woods PS, Cetin-Atalay R, Meliton AY, et al. HIF-1 regulates mitochondrial function in bone marrow-derived macrophages but not in tissue-resident alveolar macrophages. Sci Rep. 2025;15:11574. doi:10.1038/s41598-025-95962-3
61. Smolarz B, Łukasiewicz H, Samulak D, et al. Hypoxia-induced factor-1α and its role in endometrial cancer. Anticancer Res. 2024;44:3697–3712. doi:10.21873/anticanres.17195
62. Wang Y, Wu W, Xu Y, et al. Ncl liquid-liquid phase separation and SUMOylation mediate the stabilization of HIF-1α expression and promote pyroptosis in ischemic hindlimb. Biochim Biophys Acta Mol Basis Dis. 2025;1871(4):167706. doi:10.1016/j.bbadis.2025.167706
63. Lee SY, Kim SJ, Park KH, et al. Differential but complementary roles of HIF-1α and HIF-2α in the regulation of bone homeostasis. Commun Biol. 2024;7:892. doi:10.1038/s42003-024-06581-z
64. Yang HL, Lin PY, Vadivalagan C, et al. Coenzyme Q(0) defeats NLRP3-mediated inflammation, EMT/metastasis, and Warburg effects by inhibiting HIF-1α expression in human triple-negative breast cancer cells. Arch Toxicol. 2023;97:1047–1068. doi:10.1007/s00204-023-03456-w
65. Wu KKL, Xu X, Wu M, et al. MDM2 induces pro-inflammatory and glycolytic responses in M1 macrophages by integrating iNOS-nitric oxide and HIF-1α pathways in mice. Nat Commun. 2024;1(15):8624. doi:10.1038/s41467-024-53006-w
66. Zhu J, Jin Z, Wang J, et al. FGF21 ameliorates septic liver injury by restraining proinflammatory macrophages activation through the autophagy/HIF-1α axis. J Adv Res. 2025;69:477–494. doi:10.1016/j.jare.2024.04.004
67. Gomes M, Guimarães ES, Marinho FV, et al. STING regulates metabolic reprogramming in macrophages via HIF-1α during Brucella infection. PLoS Pathog. 2021;17e1009597.
68. Dzhalilova D, Kosyreva A, Lokhonina A, et al. Molecular and phenotypic distinctions of macrophages in tolerant and susceptible to hypoxia rats. PeerJ. 2023;11e16052.
69. Raulien N, Friedrich K, Strobel S, et al. Glucose-oxygen deprivation constrains HMGCR function and Rac1 prenylation and activates the NLRP3 inflammasome in human monocytes. Sci Signal. 2024;
70. He S, Fan C, Ji Y, et al. SENP3 facilitates M1 macrophage polarization via the HIF-1α/PKM2 axis in lipopolysaccharide-induced acute lung injury. Innate Immun. 2023;29:25–34. doi:10.1177/17534259231166212
71. Mager CE, Mormol JM, Shelton ED, et al. p38 MAPK and MKP-1 control the glycolytic program via the bifunctional glycolysis regulator PFKFB3 during sepsis. J Biol Chem. 2023;299:103043. doi:10.1016/j.jbc.2023.103043
72. Yuk JM, Kim TS, SY K, et al. Orphan nuclear receptor errα controls macrophage metabolic signaling and a20 expression to negatively regulate tlr-induced inflammation. Immunity. 2015;43:80–91. doi:10.1016/j.immuni.2015.07.003
73. Shen Y, Dhar R, Liu J, et al. TRPM2 deficiency contributes to M2b macrophage polarization via the PI3K/AKT/CREB pathway in murine sepsis. Innate Immun. 2025;31:569151471. doi:10.1177/17534259251343377
74. Murdoch C, Muthana M, Lewis C E. Hypoxia regulates macrophage functions in inflammation. J Immunol, 2005;175(10), 6257–63. doi:10.4049/jimmunol.175.10.6257
75. Johnstone K F, Herzberg M C. Antimicrobial peptides: Defending the mucosal epithelial barrier. Front Oral Health. 2022;3:958480. doi:10.3389/froh.2022.958480
76. Ding W, Huang C, Chen J, et al. Exploring the molecular mechanism by which kaempferol attenuates sepsis-related acute respiratory distress syndrome based on network pharmacology and experimental verification. Curr Comput Aided Drug Des. 2025;21:166–178. doi:10.2174/0115734099295805240126043059
77. Sun DS, Chang HH. Emerging role of the itaconate-mediated rescue of cellular metabolic stress. Tzu Chi Med J. 2022;34:134–138. doi:10.4103/tcmj.tcmj_79_21
78. Wang W, Feng N, Shi Q, et al. Succinate accumulation accelerates oxidative stress to promote pulmonary epithelial cell apoptosis during lung ischemia-reperfusion injury. J Cell Mol Med. 2025;29;e70645.
79. She L, Xu D, Wang Z, et al. Curcumin inhibits hepatic stellate cell activation via suppression of succinate-associated HIF-1α induction. Mol Cell Endocrinol. 2018;476:129–138. doi:10.1016/j.mce.2018.05.002
80. Wang B, Wang X, Guo M, et al. Succinate reduces biological activity and mitochondrial function of human adipose-derived stem cells. Cell Cycle. 2025:1–13.
81. Akgün AE, Yaylalı YT, Seçme M, et al. Kynurenine-PARP-1 link mediated by microRNA 210 may be dysregulated in pulmonary hypertension. Anatol J Cardiol. 2022;26:388–393. doi:10.5152/AnatolJCardiol.2021.861
82. Valencia-Cervantes J, Sierra-Vargas MP. Regulation of Cancer-Associated miRNAs Expression under Hypoxic Conditions. Anal Cell Pathol. 2024;2024:5523283. doi:10.1155/2024/5523283
83. Schmidt B, Roessler C, Schumann J. Septic-Induced microRNA expression modulations are linked to angiogenesis, vasomotion, and hypoxia-induced processes. Adv Exp Med Biol. 2018;1072:227–231.
84. Li K, Wu L, Zhu L, et al. The role of herb-partitioned moxibustion in the angiogenesis of colitis-associated cancer in rats. J Inflamm Res. 2025;18:6623–6639. doi:10.2147/JIR.S518214
85. Maeno H, Ono T, Dhar DK, et al. Expression of hypoxia inducible factor-1alpha during liver regeneration induced by partial hepatectomy in rats. Liver Int. 2005;25:1002–1009. doi:10.1111/j.1478-3231.2005.01144.x
86. Wang G, Ma X, Huang W, et al. Macrophage biomimetic nanoparticle-targeted functional extracellular vesicle micro-RNAs revealed via multiomics analysis alleviate sepsis-induced acute lung injury. J Nanobiotechnology. 2024;22:362. doi:10.1186/s12951-024-02597-z
87. Du H, Li CH, Gao RB, et al. Inhibition of the interaction between microglial adenosine 2A receptor and NLRP3 inflammasome attenuates neuroinflammation posttraumatic brain injury. CNS Neurosci Ther. 2024;30;e14408.
88. Moretti J, Jia B, Hutchins Z, et al. Caspase-11 interaction with NLRP3 potentiates the noncanonical activation of the NLRP3 inflammasome. Nat Immunol. 2022;23:705–717. doi:10.1038/s41590-022-01192-4
89. Zhao Y, Wu Y, Qu L, et al. NOD3 reduces sepsis-induced acute lung injury by regulating the activation of NLRP3 inflammasome and the polarization of alveolar macrophages. Inflammation. 2024;48:2395–2406. doi:10.1007/s10753-024-02197-x
90. Wang X, Wu FP, Huang YR, et al. Matrine suppresses NLRP3 inflammasome activation via regulating PTPN2/JNK/SREBP2 pathway in sepsis. Phytomedicine. 2023;109:154574. doi:10.1016/j.phymed.2022.154574
91. He W, Dong H, Wu C, et al. The role of NLRP3 inflammasome in sepsis: a potential therapeutic target. Int Immunopharmacol. 2023;115:109697. doi:10.1016/j.intimp.2023.109697
92. Zhong Z, Wang H, Liu X, et al. Inhibition of AKR1C3 attenuates sepsis-induced acute liver injury by blocking the TRAF6/NF-κB pathway and NLRP3 inflammasome activation. Int Immunopharmacol. 2025;168:115859. doi:10.1016/j.intimp.2025.115859
93. Li X, Zeng Q, Yao R, et al. Rapamycin mitigates organ damage by autophagy-mediated NLRP3 inflammasome inactivation in sepsis. Histol Histopathol. 2024;39:1167–1177. doi:10.14670/HH-18-706
94. Wu R, Wang N, Comish PB, et al. Inflammasome-dependent coagulation activation in sepsis. Front Immunol. 2021;12:641750. doi:10.3389/fimmu.2021.641750
95. Cao L, Wen M, Hu Z, et al. Homeodomain-interacting protein kinase 2 regulates NLRP3 inflammasome activation through endoplasmic reticulum stress in septic liver injury. J Int Med Res. 2023;51:655733096. doi:10.1177/03000605231173272
96. Luo R, Li X, Wang D. Reprogramming macrophage metabolism and its effect on NLRP3 inflammasome activation in sepsis. Front Mol Biosci. 2022;9:917818. doi:10.3389/fmolb.2022.917818
97. Meyers AK, Wang Z, Han W, et al. Pyruvate dehydrogenase kinase supports macrophage NLRP3 inflammasome activation during acute inflammation. Cell Rep. 2023;42:111941. doi:10.1016/j.celrep.2022.111941
98. Luo L, Zhuang X, Fu L, et al. The role of the interplay between macrophage glycolytic reprogramming and NLRP3 inflammasome activation in acute lung injury/acute respiratory distress syndrome. Clin Transl Med. 2024;14;e70098.
99. Lin HC, Chen YJ, Wei YH, et al. Lactic Acid Fermentation Is Required for NLRP3 Inflammasome Activation. Front Immunol. 2021;12:630380. doi:10.3389/fimmu.2021.630380
100. Yu Q, Guo M, Zeng W, et al. Interactions between NLRP3 inflammasome and glycolysis in macrophages: new insights into chronic inflammation pathogenesis. Immun Inflamm Dis. 2022;10;e581.
101. Zhuang J, Hai Y, Lu X, et al. A self-assembled metabolic regulator reprograms macrophages to combat cytokine storm and boost sepsis immunotherapy. Research. 2025;8:663. doi:10.34133/research.0663
102. Wang TT, Du YW, Wang W, et al. Inhibition of xanthine oxidase protects against sepsis-induced acute kidney injury by ameliorating renal hypoxia. Oxid Med Cell Longev. 2022;2022:4326695. doi:10.1155/2022/4326695
103. Jampasri S, Reabroi S, Tungmunnithum D, et al. Plumbagin suppresses breast cancer progression by downregulating hif-1α expression via a PI3K/Akt/mTOR independent pathway under hypoxic condition. Molecules. 2022;28:27. doi:10.3390/molecules28010027
104. Lang SA, Gaumann A, Koehl GE, et al. Mammalian target of rapamycin is activated in human gastric cancer and serves as a target for therapy in an experimental model. Int, J, Cancer. 2007;120:1803–1810. doi:10.1002/ijc.22442
105. Hong Z, Tie Q, Zhang L. Targeted inhibition of the GRK2/HIF-1α pathway is an effective strategy to alleviate synovial hypoxia and inflammation. Int Immunopharmacol. 2022;113:109271. doi:10.1016/j.intimp.2022.109271
106. Zhao S, Gao J, Li J, et al. pd-l1 regulates inflammation in lps-induced lung epithelial cells and vascular endothelial cells by interacting with the hif-1α signaling pathway. Inflammation. 2021;44:1969–1981. doi:10.1007/s10753-021-01474-3
107. Ruan H, Zhang Q, Zhang YP, et al. Unraveling the role of HIF-1α in sepsis: from pathophysiology to potential therapeutics-a narrative review. Crit Care. 2024;28:100. doi:10.1186/s13054-024-04885-4
108. Zeng L, Zhou HY, Tang NN, et al. Wortmannin influences hypoxia-inducible factor-1 alpha expression and glycolysis in esophageal carcinoma cells. World J Gastroenterol. 2016;22:4868–4880. doi:10.3748/wjg.v22.i20.4868
109. Zhong H, Chiles K, Feldser D, et al. Modulation of hypoxia-inducible factor 1alpha expression by the epidermal growth factor/phosphatidylinositol 3-kinase/PTEN/AKT/FRAP pathway in human prostate cancer cells: implications for tumor angiogenesis and therapeutics. Cancer Res. 2000;60:1541–1545.
110. Zou XZ, Hao JF, Hou MX. Hmgcs2 regulates M2 polarization of macrophages to repair myocardial injury induced by sepsis. Aging. 2023;15:7794–7810. doi:10.18632/aging.204944
111. Prabha PS, Das UN, Ramesh G, et al. Free radical generation, lipid peroxidation and essential fatty acids in patients with septicemia. Prostaglandins Leukot Essent Fatty Acids. 1991;42:61–65. doi:10.1016/0952-3278(91)90068-G
112. Xiao X, Zhou L, Xu Y, et al. Advances in the changes of macrophage metabolism and the regulation of immunometabolism during sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue. 2020;32:249–252. doi:10.3760/cma.j.cn121430-20191021-00049
113. Zhao S, Li H, Luo W, et al. Whole transcription analysis identified the regulation of hypoxia-inducible factors in monocytes with immune suppression: implications for clinical outcomes. Shock. 2025;63:541–551. doi:10.1097/SHK.0000000000002479
114. Wang H, Yang R, Chen N, et al. Heterogeneity of neutrophils and immunological function in neonatal sepsis: analysis of molecular subtypes based on hypoxia-glycolysis-lactylation. Mediators Inflamm. 2025;2025:5790261. doi:10.1155/mi/5790261
115. May CN, Ow CP, Pustovit RV, et al. Reversal of cerebral ischaemia and hypoxia and of sickness behaviour by megadose sodium ascorbate in ovine Gram-negative sepsis. Br J Anaesth. 2024;133:316–325. doi:10.1016/j.bja.2024.04.058
116. Peiris R, Trask-Marino A, Jufar A, et al. Effects of tempol on renal medullary tissue hypoxia in an ovine model of Gram-negative septic acute kidney injury. Exp Physiol. 2025. doi:10.1113/EP092396
117. Larson DF, Horak K. Macrophage migration inhibitory factor: controller of systemic inflammation. Crit Care. 2006;10:138. doi:10.1186/cc4899
118. Corcoran SE, O’Neill LA. HIF1α and metabolic reprogramming in inflammation. J Clin Invest. 2016;126:3699–3707. doi:10.1172/JCI84431
119. Liu C, Xuan W, Cao S, et al. Type II alveolar epithelial cells promote sepsis-induced immunosuppression in alveolar macrophages via exosomal lncrna rmrp release. Adv Sci. 2025;e376.
120. Rahimi A, Soudi S, Vakilian S, et al. Bacteriophage M13 modulates the sepsis-related inflammatory responses and organ damage in a clp model. Shock. 2023;59:493–504. doi:10.1097/SHK.0000000000002076
121. Teh YC, Chooi MY, Liu D, et al. Transitional premonocytes emerge in the periphery for host defense against bacterial infections. Sci Adv. 2022;8:eabj4641. doi:10.1126/sciadv.abj4641
122. Li Y, Li P, Wang W, Zhang Y, Li S, Li D, Lin Q. Nanoparticle-based drug delivery systems targeting inflammatory immune mechanisms in acute myocardial infarction: current advances and perspectives. Front Cardiovasc Med. 2025;12:1657300. doi:10.3389/fcvm.2025.1657300
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Extracellular Histones Activate Endothelial NLRP3 Inflammasome and are Associated with a Severe Sepsis Phenotype

Beltrán-García J, Osca-Verdegal R, Pérez-Cremades D, Novella S, Hermenegildo C, Pallardó FV, García-Giménez JL
Journal of Inflammation Research 2022, 15:4217-4238
Published Date: 25 July 2022
CD146+ Umbilical Cord Mesenchymal Stem Cells Exhibit High Immunomodulatory Activity and Therapeutic Efficacy in Septic Mice
Zhang L, Zhang X, Liu Y, Zhang W, Wu CT, Wang L
Journal of Inflammation Research 2023, 16:579-594
Published Date: 11 February 2023
BAP31 Promotes Adhesion Between Endothelial Cells and Macrophages Through the NF-κB Signaling Pathway in Sepsis
He J, Jing D, Zhao S, Duan M
Journal of Inflammation Research 2024, 17:1267-1279
Published Date: 26 February 2024
The Values of PDK1 and LDH Levels in Patients with Sepsis and Septic Shock: A Prospective Observational Study
Liu J, Zhou G, Tong Z, Wang X, Liu D
Journal of Inflammation Research 2024, 17:6815-6826
Published Date: 30 September 2024
Lung Tissue Extracellular Vesicles-Mediated Delivery of miR-128-3p as a Novel Mechanism of Acute Lung Inflammation
Deng W, Zhu X, Li H, Hu P, Qian K, Liu F
International Journal of Nanomedicine 2025, 20:4831-4848
Published Date: 15 April 2025