Back to Journals » Journal of Pain Research » Volume 18
LncRNA-Associated ceRNA Network in the Spinal Cord Reveals the Central Mechanism of Chronic Inflammatory Visceral Pain
Authors Li X, Wang X, Dong X, Li Z, Lu Y, Liu L, Yang G, Hong J ![]() , Yang Y
, Yang Y ![]() , Zhang C, Wu H
, Zhang C, Wu H ![]() , Zhang D
, Zhang D ![]() , Ma X
, Ma X
Received 29 May 2025
Accepted for publication 8 December 2025
Published 17 December 2025 Volume 2025:18 Pages 6853—6868
DOI https://doi.org/10.2147/JPR.S543556
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Wendy Imlach
Xiaoying Li,1,2,* Xuejun Wang,3,* Xiaoqing Dong,4,* Zhiyuan Li,5,* Yunqiong Lu,1 Li Liu,1 Guang Yang,2 Jue Hong,2 Yanting Yang,2 Cuihong Zhang,2 Huangan Wu,1,2 Dan Zhang,2 Xiaopeng Ma1,2
1Yueyang Hospital of Integrated Traditional Chinese and Western Medicine, Shanghai University of Traditional Chinese Medicine, Shanghai, People’s Republic of China; 2Laboratory of Acupuncture-Moxibustion and Immunology, Shanghai Research Institute of Acupuncture and Meridian, Shanghai, People’s Republic of China; 3Eye Institute and Department of Ophthalmology, Eye & ENT Hospital, Fudan University, Shanghai, People’s Republic of China; 4Department of Acupuncture and Moxibustion, Xi’an Traditional Chinese Medicine Hospital of Encephalopathy, Xi’an, People’s Republic of China; 5Department of Acupuncture and Moxibustion, Shenzhen Hospital, Beijing University of Chinese Medicine, Shenzhen, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Xiaopeng Ma; Dan Zhang, Email [email protected]; [email protected]
Objective: To study and analyze the varieties across long non-coding RNAs (lncRNAs), microRNAs (miRNAs), and mRNAs in spinal cord, construct the competing endogenous RNAs (ceRNAs) network, and discuss the possible central mechanism in the development of chronic inflammatory visceral pain (CIVP).
Methods: The colitis-associated CIVP rat model was prepared using 2,4,6-trinitrobenzene sulfonic acid (TNBS) enema. The abdominal withdrawal reflex (AWR), mechanical withdrawal threshold (MWT), and thermal withdrawal latency (TWL) were used to evaluate the rat’s pain behaviors. The whole transcriptome sequencing technique was adopted to analyze the differentially expressed lncRNAs, miRNAs, and mRNAs in the spinal cord. LncRNA-associated ceRNA network was constructed and identified using the ceRNA MuTATE method and the hypergeometric distribution algorithm. The sequencing result was then verified using the quantitative real-time PCR (RT-qPCR).
Results: Compared to the normal group, CIVP rat models had significantly increased AWR and decreased MWT and TWL (all P< 0.05), suggesting hyperalgesia. In the spinal cord of CIVP rats, 158 lncRNAs, 24 miRNAs, and 1216 mRNAs were found to be differentially expressed. 3 top-scored lncRNAs, 1 pain-related lncRNA, 3 downstream miRNAs, and 2 mRNAs were selected for RT-qPCR verification, and 2 lncRNAs, 3 miRNAs, and 2 mRNAs were verified (all P< 0.05).
Conclusion: LncRNA TCONS_00001477 and TCONS_00019815 may regulate Col2a1 and Hoxd10 mRNAs by competitively binding to miR-3120, miR-542-5p, and miR-450b-3p, thereby participating in colitis associated CIVP.
Keywords: colitis, visceral pain, lncRNA, ceRNA network, central mechanism
Introduction
Visceral pain can be found in numerous visceral diseases, such as inflammatory bowel disease (IBD), pancreatitis, functional dyspepsia, and irritable bowel syndrome (IBS).1,2 Inflammatory visceral pain (IVP), a form of pathological pain experienced by IBD patients, usually presents as abdominal pain.3 Prolonged IVP will hinder the patient’s quality of life (QOL), affect their social function, and aggravate the risk for mental disorders like anxiety and depression.4 Medications for IVP primarily include opioids and non-steroidal anti-inflammatory drugs (NSAIDs), yet the efficacy is rather unsatisfactory, which suggests certain complexity in the mechanism of IVP’s development and persistence.5 Therefore, it is indeed necessary to study the potential molecular mechanism of IVP.
Long non-coding RNAs (lncRNAs) are a group of non-coding RNA transcripts with over 200 nucleotides in length. A series of studies have proved the role of lncRNAs in regulating chronic pain. For instance, lncRNA MALAT1 promotes neuropathic pain progression in chronic constriction injury (CCI) rat models;6 inhibition of lncRNA DILC attenuates neuropathic pain via the SOCS3/JAK2/STAT3 pathway.7 Evidence also supports the role of microRNAs (miRNAs) as a key regulator in the onset of chronic neuropathic pain.8–10 LncRNAs can bind to miRNAs as competing endogenous RNAs (ceRNAs) to modulate the expression of target messenger RNAs (mRNAs), affecting the development of various diseases.11–13 In recent years, the ceRNA network has gone viral in clinical research and has been found closely related to chronic pathological and inflammatory pain.14–16 However, the role of the lncRNA-related ceRNA network in chronic inflammatory visceral pain (CIVP) has not been reported yet.
The spinal cord plays an important role in the formation of visceral pain associated with gastrointestinal disorders.17 The abnormal activation of spinal cells, particularly the microglia and astrocyte, ignite the neural transduction continuously via multiple inflammatory mediators and promoting central sensitization which contributes to visceral hypersensitivity.18 A study shows that the spinal LncRNA RT1-CE10 can influence chronic visceral pain by interaction with ATP1a3.19 So our research will focus on spinal RNA profiles and lncRNA-related ceRNA network regulation in CIVP. Hence, by establishing rat models of CIVP using 2,4,6-trinitrobenzene sulfonic acid (TNBS) enema, this research was to observe the expression profiles of lncRNAs, miRNAs, and mRNAs in the rat’s spinal cord using the whole transcriptome RNA sequencing (RNA-seq) technique and unveil the expression differences between normal rats and CIVP rats. Further, we also built a lncRNA-miRNA-mRNA ceRNA network and analyzed the function and pertinent pathways of the differentially expressed genes based on Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG), aiming to reveal the central mechanism in the development of CIVP and seek novel target for the treatment.
Materials and Methods
Animals
Fourteen healthy specific-pathogen-free (SPF) male Sprague-Dawley (SD) rats, weighing (150±20) g, were provided by Shanghai Jihui Experimental Animal Co., Ltd., China (SCXK(Shanghai)2017–0012). The rats were housed at (20±2) °C and 50%–70% humidity for seven days of adaptive feeding. This experiment conformed to the Guiding Opinions on Treating Animals issued by the Ministry of Science and Technology of the People’s Republic of China ((2006)398) and was approved by the Ethics Committee of Yueyang Clinical Medicine School, Shanghai University of Traditional Chinese Medicine (No. YYLAC-2020-085). The rats were randomized into a normal group (NG) or a model group (MG), each consisting of 7 rats.
CIVP Modeling
The enema solution was made by mixing 5% (W/V) TNBS (100 mg/Kg, Sigma, MO, USA) and 50% ethanol at a volume ratio of 2:1 and was administered at 3 mL/Kg to prepare the colitis related CIVP rat models for 4 consecutive weeks.20–22 At the seventh day after completing the modeling, we estimated the rat’s pain behaviors and randomly selected one rat from each group for colonic histopathological observation to identify if the model was a success.
Pain Behavior Tests
Abdominal Withdrawal Reflex (AWR)
According to AI-Chaer, et al,23 the AWR was tested by applying four levels of distending pressure in sequence to measure visceral sensitivity: 20 mmHg, 40 mmHg, 60 mmHg, and 80 mmHg. Each pressure level was tested three times at a five-minute interval, each time lasting 20s, and the average score was taken as the final result. Scoring criteria:24 no behavioral response, scored 0 points; occasional head movements at the start of the stimulation while the rat maintaining stationary, scored 1 point; slight contraction of rat abdominal muscles but without lifting, scored 2 points; strong contraction of abdominal muscles with obvious lifting but no pelvic or scrotal lifting, scored 3 points; arched abdomen with the pelvis and scrotums lifted, scored 4 points.
Mechanical Withdrawal Threshold (MWT)
The MWT was performed by stimulating the rat’s hind paw center with the von Frey filament (Stoelting, IL, USA), no longer than 4 s at a time, to see if the rat would present positive responses like lifting or licking the paw.25 If no positive responses were triggered, a higher stimulation level would be provided until the initial positive response occurred. Then, the rat would be stimulated again with a lower stimulation level. Each level of stimulation was applied five times at an interval of 30s. The stimulation gradient started from 2.0 g, to 4.0 g, 6.0 g, 8.0 g, and 15.0 g. The 50% withdrawal threshold value was calculated using the up-down method.
Thermal Withdrawal Latency (TWL)
The Hargreaves’ method26 was referred to conduct the TWL test using the BME2410A thermal stimulator (Institute of Medical Biology of Chinese Academy of Medical Sciences, Yunnan, China) to radiate the rat’s one hind sole. The rat was placed in a glass box for 15 min to adapt before the test. The TWL reading was the time from the start of the thermal radiation till the rat lifted its paw. Each rat underwent three times of tests with a three-minute break in between two tests.
Sample Preparation
After the pain behavior tests, rats were anesthetized using intraperitoneal injection of pentobarbital sodium and sacrificed via abdominal aortic blood collection. The part of the intestine from 2 cm above the anus to the end of the cecum was collected and fixed in 4% paraformaldehyde. Then, the L6-S2 segment of spinal cord was collected and kept at −80 °C.
Colonic Histopathological Observation
The colon tissues kept in 4% paraformaldehyde were treated with formaldehyde fixation, gradient ethanol and dimethylbenzene dehydration, and paraffin embedding. Afterward, the samples were sliced at 4 μm, stained with hematoxylin-eosin (HE), and observed under an optical microscope (Olympus, Tokyo, Japan) for histomorphological changes.
RNA-Seq
Each group contributed three pieces of whole spinal cord samples (L6-S2) to run RNA-seq. The total RNA was extracted first using the mirVana™ miRNA isolation kit (Ambion-1561, USA). The NanoDrop 2000 ultraviolet spectrophotometer (Thermo Fisher, MA, USA) was used to determine the purity and concentration. The Agilent 2100 TapeStation system (Agilent, CA, USA) was employed to check the quality of the total RNA; those with an RNA integrity number (RIN) within 7–10 were considered qualified samples for RNA-seq. The TruSeq Stranded Total RNA Library Prep Gold kit containing RiboZero beads (Illumina, CA, USA) was adopted to build the lncRNA and mRNA libraries, and the TruSeq Small RNA Sample Prep kit (Illumina, CA, USA) was used to construct the microRNA library. HISAT227,28 was used to align the clean reads with designated reference genomes to obtain its locus on the reference genome and the typical sequence information of the sample. We used Bowtie229 to align the clean reads and Express for quantification to obtain the FPKM (fragments per kilobase of exon model per million mapped fragments) and counts of mRNA and lncRNA to calculate their expression. The expression of miRNA was expressed as transcript per million (TPM). The raw sequences were deposited into the Sequence Read Archive (SRA) database with the BioProject accession number PRJNA905659. (https://www.ncbi.nlm.nih.gov/sra/PRJNA905659).
Screening and Cluster Analysis of Differentially Expressed Genes
The DeSeq software was used to analyze between-group differences in repetitive biological data, and the edgeR software was used to analyze non-repetitive biological data. The fold change (FC) and P-value were adopted to present the expression differences of one gene in different samples. When P<0.05 and log2FC>1.5, it was accepted that the lncRNAs and miRNAs were differentially expressed between groups; P<0.05 and log2FC>1.2 indicated that the mRNAs were differentially expressed between groups.30 Cluster 3.0 and Treeview programs were used to generate heat maps and volcano plots of the differentially expressed genes.
Construction of ceRNA Network
To learn the relationships among lncRNAs, miRNAs, and mRNAs, we first calculated the co-expressed miRNA-lncRNA pairs and miRNA-mRNA pairs and predicted the mutual regulatory relationship via the Miranda software. The threshold value of correlation analysis was set as the absolute value of correlation coefficient r ≥ 0.80 and P ≤ 0.05.31 Further, the ceRNA MuTATE method32 was used to calculate the ceRNA pair scores and the hypergeometric distribution algorithm was used to calculate the possibility of ceRNA pairs sharing certain miRNAs. Finally, the ceRNA relationship pair with high reliability was obtained. The Cytoscape software was employed to visualize the corresponding regulation networks.
Functions and Pathway Enrichments of the Target Genes
We used GO (http://www.geneontology.org) and KEGG (http://www.genome.ad.jp/kegg) to study the biological functions of pathway enrichments of the target genes in the ceRNA network and adopted STRING (https://string-db.org)33 to analyze the interactions of the target genes’ corresponding proteins.
Verification of the RNA-Seq Results
Quantitative real-time PCR (RT-qPCR) was used to verify the RNA-seq results. First, we employed Trizol (Invitrogen, CA, USA) to extract the total RNA from the spinal cord and used ND-1000 Nanodrop (Thermo Fisher, MA, USA) to detect the concentration. Then, we translated the total RNA into cDNA using a miRNA reverse transcription kit (EZBioscience, MN, USA) and amplified cDNA in the Roche LightCycler 480 (Roche, Basel, Switzerland) with the miRNA qPCR kit (EZBioscience, MN, USA); the PrimeScript™RT Master Mix kit (Perfect Real Time Kit, Takara, Shiga, Japan) was used to synthesize the cDNA of lncRNA and mRNA, and the amplification was done in the TB Green® Premix Ex Taq ™ II kit (Takara, Shiga, Japan). We adopted the 2−ΔΔCt method to analyze the relative expression of the transcripts. The primer sequence was synthesized by Shanghai Jierui Co., Ltd (Supplementary Table 1).
Statistical Analyses
Data analyses were performed using SPSS 24.0. Data that conformed to normal distribution were expressed as mean ± standard deviation (x±SD) and as [Median(P25, P75)] if not. The independent samples t-test was used to compare data that concurrently met normal distribution and homogeneity of variance; the non-parametric test was used if either condition was not met. Statistical significance was accepted when P<0.05 (α=0.05).
Results
CIVP Rat’s Pain Behavior Assessment
Under the optical microscope, we found integrated epithelial structures with uniformly distributed single-layer columnar epithelial cells, continuous intestinal mucosa, and neatly arranged glands without infiltration of inflammatory cells in the colonic tissues of NG rats (Figure 1A). In contrast, the colonic tissues of CIVP rats in the MG showed seriously damaged mucosa, loss of epithelial layer, ulceration affecting the submucosal and muscular layers, irregularly arranged or disappeared glands, submucosal thickening, vascular proliferation, and massive infiltration of monocytes and lymphocytes (Figure 1B). Compared to rats in the NG, CIVP rats showed notably increased AWR scores at 40 mmHg, 60 mmHg, and 80 mmHg distending levels (all P<0.05) and significantly decreased MWT and TWL (both P<0.01) (Figure 1C–E).
 |
Figure 1 Morphological observation of rat’s colonic tissues, AWR scores, MWT, and TWL (A) NG rat’s colonic tissue (HE staining, ×200); (B) MG rat’s colonic tissue (HE staining, ×200). Arrows indicate ulcers and inflammatory responses; (C) Comparison of rat’s AWR scores; (D) Comparison of rat’s MWT; (E) Comparison of rat’s TWL. n=6, vs NG, *P<0.05, **P<0.01. Abbreviations: NG, Normal group; MG, Model group; HE, Hematoxylin-eosin; AWR, Abdominal withdrawal reflex; MWT, Mechanical withdrawal threshold; TWL, Thermal withdrawal latency. |
Differentially Expressed lncRNAs, miRNAs, and mRNAs in CIVP Rat’s Spinal Cord
Compared to the NG, CIVP rats had 158 differentially expressed (DE) lncRNAs (DElncRNAs) (79 up-regulated and 79 down-regulated), 24 DEmiRNAs (all down-regulated), and 1216 DEmRNAs (404 up-regulated and 812 down-regulated) (Figure 2 and Supplementary Tables 2–4)).
 |
Figure 2 Differentially expressed lncRNAs, miRNAs, and mRNAs in CIVP rat’s spinal cord. (A) Heat map of lncRNAs; (B) Volcano plot of lncRNAs; (C) Heat map of miRNAs; (D) Volcano plot of miRNAs; (E) Heat map of mRNAs; (F) Volcano plot of mRNAs. On the heat map, the red color suggests a high expression level, and the blue color for a low expression level. On the volcano plot, the red spots stand for up-regulated genes, and the green spots stand for down-regulated genes. |
Abbreviations: NG, Normal group; MG, Model group.
Construction of the lncRNA-miRNA-mRNA ceRNA
The pearsonr was used to calculate the lncRNA-miRNA and miRNA-mRNA correlations, and the Miranda program was adopted to predict their bindings. Finally, 41 miRNA-lncRNA pairs and 919 miRNA-mRNA pairs were collected, and ceRNAs were analyzed. According to the rank of the MuTATE score, the top 100 mRNA-lncRNA pairs were mapped; of the 100 mRNA-lncRNA pairs, 200 mRNA-miRNA-lncRNA groups were mapped (Figure 3 and Supplementary Table 5).
 |
Figure 3 Crosstalk analysis and the establishment of ceRNA network. (A) Interaction map of the top 100 mRNA-lncRNA pairs; (B) Interaction map of 200 lncRNA-miRNA-mRNA groups. The yellow, green, and purple colors stand for lncRNAs, miRNAs, and mRNAs, respectively. |
Functional Analysis and Pathway Enrichment of mRNAs
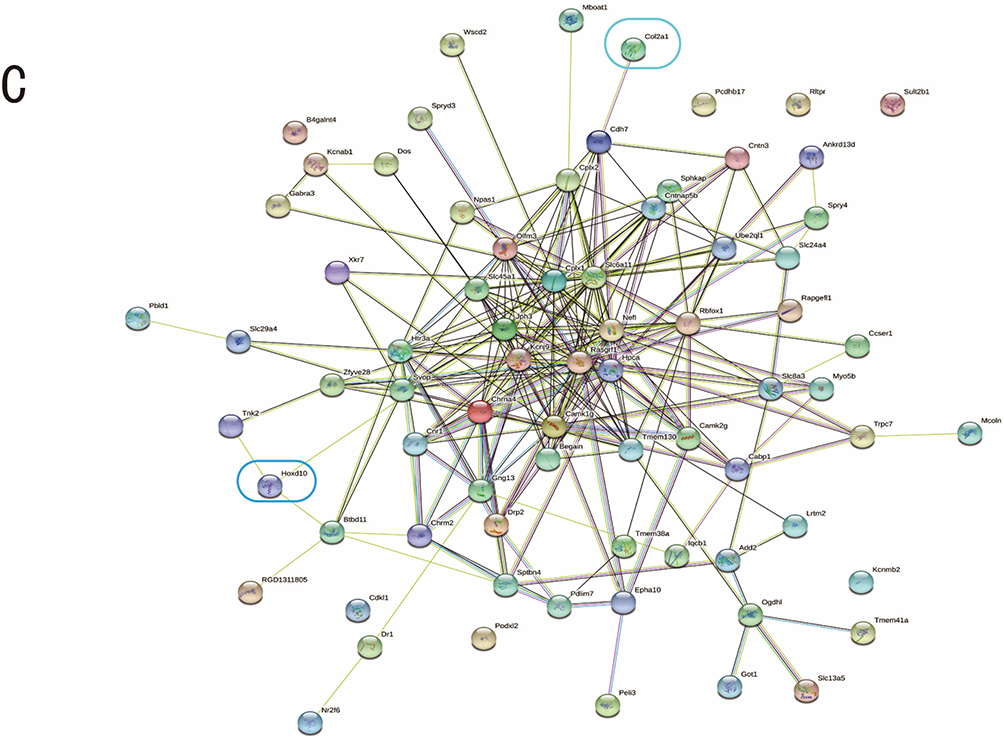
We analyzed the mRNAs in the ceRNA network using GO and KEGG, finding that the involved biological processes mainly include synaptic plasticity, ion transmembrane transport, and calmodulin-dependent protein kinase activity, and Toll signaling pathway; the involved cellular components include neuronal cell bodies, cell membranes, projection neurons, and postsynaptic membranes; the involved molecular functions include calmodulin protein binding, neuropeptide Y receptor activity, cation channel activity, and neurotransmitter receptor binding (Figure 4A). The pathways primarily included calcium signaling pathway, neuroactive legend-receptor interaction, cholinergic synapses, neural retrograde signaling, Apelin signaling pathway, and Ras signaling pathway (Figure 4B). The protein-protein interaction (PPI) analysis revealed PPIs between multiple target genes (Figure 4C).
Figure 4 Continued. Figure 4 GO, KEGG, and PPI analyses of mRNAs in the ceRNA network. (A) The top 10 GO enrichment items. The red color indicates biological processes, blue for cellular components, and green for molecular functions. (B) The top 30 KEGG enrichment pathways. The size of the dots corresponds to the number of differentially expressed genes in GO items. The enrichment P-value shrinks, and the significance grows as the dot’s color changes from purple to blue, green, and red. (C) PPI network. Abbreviations: GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; PPI, Protein-protein interaction.
RNA-Seq Verification
The three top-scored lncRNAs and one pain-related lncRNA were selected to undergo qRT-PCR verification. The results demonstrated that the MG was significantly different from the NG in comparing the expression of lncRNA-TCONS_00001477, TCONS_00019815, and TCONS_00024376, which was consistent with the RNA-seq findings. The three lncRNAs’ downstream miRNAs were also put through verification, discovering that miR3120, a commonly shared downstream miRNA of TCONS_00019815 and TCONS_00001477, and TCONS_00019815’s downstream miR 542–5p and miR 450b-3p passed verification. We also selected four lncRNA-regulated mRNAs for verification, of which, Col2a1 and Hoxd10 mRNAs showed significant differences between the two groups (all P<0.05) (Figure 5). The Miranda program was used to predict the targeting relationships of lncRNA-miRNA-mRNA sequences (Figure 6). The findings revealed that lncRNA-TCONS_00001477 could target miR3120 to regulate the expression of Col2a1 mRNA, and TCONS_00019815 could target miR3120, miR 542–5p, and miR 450b-3p to influence the expression of Col2a1 and Hoxd10 mRNAs.
 |
Figure 5 RNA-seq verification and targeting relationship prediction. (A) Comparison of the expression of lncRNATCONS_00001477; (B) Comparison of the expression of lncRNA TCONS_00019815; (C) Comparison of the expression of lncRNATCONS_00024376; (D) Comparison of the expression of miR3120; (E) Comparison of the expression of miR 542–5p; (F) Comparison of the expression of miR 450b-3p; (G) Comparison of the expression of Col2a1mRNA; (H) Comparison of the expression of Hoxd10 mRNA; (I) Prediction of the targeting relationships between miR3120 and lncRNA-TCONS_00001477; (J) Prediction of the targeting relationships between miR3120 and lncRNA-TCONS_00019815; (K) Prediction of the targeting relationships between miR 450–3p and lncRNA-TCONS_00019815; (L) Prediction of the targeting relationships between miR 542–5p and lncRNA-TCONS_00019815; (M) Prediction of the targeting relationships between miR 542–5p and Hoxd10; (N) Prediction of the targeting relationships between miR3120 and Col2a1; (O) Prediction of the targeting relationships between miR 542–5p and Col2a1; (P) Prediction of the targeting relationships between miR 450b-3p and Hoxd10. n=6; vs NG, *P<0.05, **P<0.01. Abbreviations: NG, Normal group; MG, Model group. |
 |
Figure 6 Verified lncRNA-miRNA-mRNA ceRNA network. |
Discussion
One investigation reported that 38% of IBD patients experienced chronic pain lasting over three months, mainly manifesting as abdominal pain, and a severe disease condition, poor QOL, and depressed and anxious moods could be commonly found among these patients, along with dependence on acetaminophen and opioid drugs.34 Inflammation is a significant pathological feature of the colon in IBD, so colitis-associated VP is mostly IVP and chronic. CIVP has complex pathogenesis and various risk factors, including inflammation, intestinal obstruction, psychological factors, neurobiological factors, and genetic factors.35 It has been generally accepted that the development of CIVP is a joint result of peripheral and central neurosensitization.36 The brain and spinal cord, the essential parts of the central nervous system for integrating and processing pain signals, play a key role in transmitting and regulating nociceptive information. The spinal cord can repeatedly receive the visceral sensory signals transmitted by the dorsal root ganglia, and amplify them. The persistent activation of spinal neurons is a crucial factor for the central sensitization.37,38 Therefore, the profound exploration of CIVP’s pathogenesis targeting central integration and regulation may provide an important breakthrough point for clinical diagnosis and treatment. In recent years, a large number of studies have mentioned that the ceRNA network is closely related to pathological pain, inflammatory pain, and visceral pain.14–16,39 However, there is not any paper reporting the role of lncRNA-associated ceRNA network in CIVP. The TNBS-induced colitis-related inflammatory pain model is well-recognized and stable.22 Hence, the current study established colitis-induced CIVP rat models using TNBS enema to observe the central regulation mechanism in CIVP from the perspective of lncRNA-associated ceRNA network. Meanwhile, we have also tested the pain behavioral indexes (AWR, TWL, MWT) of all rats to verify whether the CIVP model was valid because previous studies indicate that intestinal hypersensitivity has been associated with the peripheral sensitization of primary colonic sensory afferents and referred hyperalgesia in several body regions including the abdominal wall and hind paws.40,41
The lncRNA and miRNA both belong to non-coding RNAs (ncRNAs), playing extensive and crucial biological roles in the body. For instance, a variety of lncRNAs and miRNAs have been proven guilty in the development and persistence of inflammatory pain, neuralgia, and visceral pain.42–44 LncRNAs act to regulate iron channels, inflammatory pathways, neurotransmitter release, oxidative stress, and neuronal autophagy and apoptosis.45 MiRNAs may regulate kinds of pain by activating glial cells, promoting the expression of inflammatory factors, and inhibiting central and peripheral sensitization.46,47 This study thoroughly analyzed and collected DElncRNAs, DEmiRNAs, and DEmRNAs in the spinal cord of CIVP rat models, demonstrating that CIVP rats had significant colonic damage along with pain behavior changes and visceral hypersensitivity, consistent with previous reports.48,49 Furthermore, compared to the NG, CIVP rats had 158 DElncRNAs, 24 DEmiRNAs, and 1216 DEmRNAs in the spinal cord, suggesting that the differential expression of these RNAs in the spinal cord may be an important mechanism in the development of CIVP. Nevertheless, the interaction of the three parties in CIVP remains a question and should be studied.
LncRNA refers to a type of RNA with a length greater than 200 nucleotides and without protein-coding function. It is a predominant part of ncRNAs. Despite the incapability of coding proteins, lncRNAs can regulate mRNA translation or directly bind to proteins to modulate corresponding target genes and signaling pathways. Also, it can act as a miRNA sponge, absorbing miRNAs to regulate them, or compete with miRNA response elements to affect the function of target mRNAs. Therefore, besides collecting the differentially expressed RNAs, this study also built the lncRNA-related ceRNA network to discover the possible interactions among these RNAs. We adopted pearsonr to calculate the lncRNA-miRNA and miRNA-mRNA correlations and finally collected 41 miRNA-lncRNA pairs and 919 miRNA-mRNA pairs to map the ceRNA network. The MuTATE method and hypergeometric distribution algorithm were used to obtain more reliable ceRNA pairs. The target genes in the ceRNA network were also analyzed using GO and KEGG. Research indicates that abnormal molecular functions and signaling pathway activation of certain target genes are closely associated with chronic inflammatory pain, visceral hypersensitivity, and VP.50–56 The findings above demonstrate that mutual regulations exist in the lncRNA-related ceRNA network, and lncRNAs may affect the transcription activity of target genes by regulating miRNAs; the functional abnormalities of corresponding high-scored target genes in the ceRNA network are related to the pathogenesis of CIVP. This experiment explored the plausible ceRNA network that is related to the pathogenesis of CIVP and their mutual regulations from the perspective of lncRNAs in the spinal cord, which is different from the previous report.57
Finally, this study selected top-scored lncRNAs in the ceRNA network, lncRNA-TCONS_00001477 and lncRNA-TCONS_00019815, and the pain-related lncRNA-TCONS_00024376 for RT-qPCR verification. The results found that these lncRNAs had higher expression in CIVP rat’s spinal cord tissues (all P<0.05), which is consistent with the RNA-seq outcomes. Then, downstream miRNAs of the three lncRNAs were also put through verification, discovering that miR3120, the downstream miRNA of TCONS_00019815 and TCONS_00001477, and miR 542–5p and miR 450b-3p, the downstream miRNAs of TCONS_00019815, were differentially expressed (all P<0.01). Further, we selected the corresponding mRNAs Col2a1 and Hoxd10 in the ceRNA network to undergo qRT-PCR assay, and they also passed the verification. It was reported that miR-3120/Hsc70 could suppress the expression of TRPV1 in the dorsal root ganglion (DRG) to participate in forced swim stress‑induced mechanical hyperalgesia in rats in an inflammatory state.58 Regarding miR 542–5p, this miRNA can regulate cell proliferation and inflammatory responses.59 Col2a1, serving as a synthetic metabolic marker for cartilage metabolism, has been widely studied in osteoarthritis and lumbar disc diseases. Up-regulating Col2a1 in the cartilage tissues can promote cartilage repair and extracellular matrix synthesis in osteoarthritis rats, reducing knee joint pain.60–62 As a member of the Hox transcription factor family, Hoxd10 has been found to be expressed along the rostrocaudal axis neuronal regions of the lumbosacral spinal cord, and this gene is closely related to limb motor function.63,64 Other studies proved that Hoxd10 could attenuate colitis and Alzheimer’s disease via Rho/ROCK signaling pathway.65,66 In sum, Col2a1 and Hoxd10 can produce anti-inflammatory and neuroprotective effects, though more scientific evidence is required to see if they play a role in regulating colitis-related CIVP.
Indeed, this study still has some areas for improvement. For example, the experiment only adopted bioinformatic techniques to predict the crosstalk in the ceRNA network. Further verification of the corresponding genes and their interaction mechanisms is needed in the future. This may include using dual-luciferase reporter assay and RT-PCR to verify the expression correlation between lncRNAs and mRNAs; using methods like RNA immunoprecipitation to test if lncRNAs and mRNAs commonly share microRNA response elements (MREs); interfering with the expression of relevant RNAs combined with pain behavior tests to clarify their effects on CIVP in vivo; also, adopting flow cytometry, in vitro animal experiments, clinical trials, and other methods to verify the biological function of target genes. Besides, we selected the whole spinal cord segment from L6 to S2 for study this time, and we expect to conduct further research focusing on the microglia or astrocytes. The findings are derived from a TNBS-induced colitis model, so their generalizability to other forms of visceral pain (eg, irritable bowel syndrome) remains unclear.
Conclusion
This study detected and analyzed the differential expression of lncRNAs, miRNAs, and mRNAs in the spinal cord tissues of CIVP rats using the whole transcriptome sequencing technique, and preliminarily constructed the lncRNA-associated ceRNA network to explore the pathogenesis of CIVP, providing new research targets and ideas. The results indicate that spinal lncRNA TCONS¬_00001477 and TCONS¬_00019815 may regulate the expression of Col2a1 and Hoxd10 by competitively binding to miR3120, miR542-5p, and miR450b-3p and participate in the development of CIVP (Figure 7).
 |
Figure 7 Summariation of the research. This picture was drawn by BioRender scientific illustration software. Abbreviations: TNBS, 2,4,6-Trinitrobenzenesulfonic acid solution; DE, differentially expressed; AWR, Abdominal withdrawal reflex; MWT, Mechanical withdrawal threshold; TWL, Thermal withdrawal latency; GO, Gene Ontology; KEGG, Kyoto Encyclopedia of Genes and Genomes; PPI, Protein-protein interaction. |
Data Sharing Statement
The data that support the findings in the current study are available from the corresponding author D Zhang upon reasonable request.
Acknowledgment
OE Biotech Co. Ltd., Shanghai, China, performed the high-throughput sequencing and analysis.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the National Natural Science Foundation of China (No. 82104985, No. 81674073);Natural Science Foundation of Shanghai (No. 20ZR1453000, No. 23ZR1460100); Outstanding Leader Plan of Shanghai (No. 060); Natural Science Foundation of Shenzhen (No. JCYJ20210324135600002).
Disclosure
The authors declare that they have no competing interests.
References
1. Collett B. Visceral pain: the importance of pain management services. Br J Pain. 2013;7(1):6–7. doi:10.1177/2049463713480138
2. Drossman DA, Hasler WL. Rome IV-Functional GI Disorders: disorders of Gut-Brain Interaction. Gastroenterology. 2016;150(6):1257–1261. doi:10.1053/j.gastro.2016.03.035
3. Hardy PY, Fikri J, Libbrecht D, Louis E, Joris J. Pain Characteristics in Patients with Inflammatory Bowel Disease: a Monocentric Cross-Sectional Study. J Crohns Colitis. 2022;16(9):1363–1371. doi:10.1093/ecco-jcc/jjac051
4. Woolf CJ, Salter MW. Neuronal plasticity: increasing the gain in pain. Science. 2000;288(5472):1765–1769. doi:10.1126/science.288.5472.1765
5. Zielinska A, Salaga M, Wlodarczyk M, Fichna J. Focus on current and future management possibilities in inflammatory bowel disease-related chronic pain. Int J Colorectal Dis. 2019;34(2):217–227. doi:10.1007/s00384-018-3218-0
6. Wu J, Wang C, Ding H. LncRNA MALAT1 promotes neuropathic pain progression through the miR‑154‑5p/AQP9 axis in CCI rat models. Mol Med Rep. 2020;21(1):291–303. doi:10.3892/mmr.2019.10829
7. Liu Y, Feng L, Ren S, Zhang Y, Xue J. Inhibition of lncRNA DILC attenuates neuropathic pain via the SOCS3/JAK2/STAT3 pathway. Biosci Rep. 2020;40(6):2. doi:10.1042/BSR20194486
8. Li H, Huang Y, Ma C, Yu X, Zhang Z, Shen L. MiR-203 involves in neuropathic pain development and represses Rap1a expression in nerve growth factor differentiated neuronal PC12 cells. Clin J Pain. 2015;31(1):36–43. doi:10.1097/AJP.0000000000000070
9. Xia LX, Ke C, Lu JM. NEAT1 contributes to neuropathic pain development through targeting miR-381/HMGB1 axis in CCI rat models. J Cell Physiol. 2018;233(9):7103–7111. doi:10.1002/jcp.26526
10. Dou L, Lin H, Wang K, et al. Long non-coding RNA CCAT1 modulates neuropathic pain progression through sponging miR-155. Oncotarget. 2017;8(52):89949–89957. doi:10.18632/oncotarget.21192
11. Tay Y, Kats L, Salmena L, et al. Coding-independent regulation of the tumor suppressor PTEN by competing endogenous mRNAs. Cell. 2011;147(2):344–357. doi:10.1016/j.cell.2011.09.029
12. Salmena L, Poliseno L, Tay Y, Kats L, Pandolfi PP. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011;146(3):353–358. doi:10.1016/j.cell.2011.07.014
13. Chen G, Wang Z, Wang D, et al. LncRNADisease: a database for long-non-coding RNA-associated diseases. Nucleic Acids Res. 2012;41(D1):D983–6. doi:10.1093/nar/gks1099
14. Wang K, Bao JP, Zhou ZM, Mao L, Wu XT. CircRNA-Associated ceRNA Network Reveals Focal Adhesion and Metabolism Pathways in Neuropathic Pain. J Oncol. 2022;2022:7246904. doi:10.1155/2022/7246904
15. Hou X, Weng Y, Guo Q, et al. Transcriptomic analysis of long noncoding RNAs and mRNAs expression profiles in the spinal cord of bone cancer pain rats. Mol Brain. 2020;13(1):47. doi:10.1186/s13041-020-00589-2
16. Fang ZH, Liao HL, Tang QF, et al. Interactions Among Non-Coding RNAs and mRNAs in the Trigeminal Ganglion Associated with Neuropathic Pain. J Pain Res. 2022;15:2967–2988. doi:10.2147/JPR.S382692
17. Farmer AD, Aziz Q. Mechanisms of visceral pain in health and functional gastrointestinal disorders. Scand J Pain. 2014;5(2):51–60. doi:10.1016/j.sjpain.2014.01.002
18. Long JY, Wang XJ, Li XY, et al. Spinal Microglia and Astrocytes: two Key Players in Chronic Visceral Pain Pathogenesis. Neurochem Res. 2022;47(3):545–551. doi:10.1007/s11064-021-03486-9
19. Tang Y, Liu Z, Wu X, et al. The involvement of spinal lncRNA RT1-CE10 in chronic functional visceral pain. Mol Pain. 2025;21:17448069251358692. doi:10.1177/17448069251358692
20. Morris GP, Beck PL, Herridge MS, Depew WT, Szewczuk MR, Wallace JL. Hapten-induced model of chronic inflammation and ulceration in the rat colon. Gastroenterology. 1989;96(3):795–803.
21. Zhou Q, Price DD, Caudle RM, Verne GN. Visceral and somatic hypersensitivity in TNBS-induced colitis in rats. Dig Dis Sci. 2008;53(2):429–435. doi:10.1007/s10620-007-9881-6
22. Zhou Q, Price DD, Caudle RM, Verne GN. Visceral and somatic hypersensitivity in a subset of rats following TNBS-induced colitis. Pain. 2008;134(1–2):9–15. doi:10.1016/j.pain.2007.03.029
23. Al-Chaer ED, Kawasaki M, Pasricha PJ. A new model of chronic visceral hypersensitivity in adult rats induced by colon irritation during postnatal development. Gastroenterology. 2000;119(5):1276–1285. doi:10.1053/gast.2000.19576
24. Xu GY, Winston JH, Chen JD. Electroacupuncture attenuates visceral hyperalgesia and inhibits the enhanced excitability of colon specific sensory neurons in a rat model of irritable bowel syndrome. Neurogastroenterol Motil. 2009;21(12):1302–e125. doi:10.1111/j.1365-2982.2009.01354.x
25. Chaplan SR, Bach FW, Pogrel JW, Chung JM, Yaksh TL. Quantitative assessment of tactile allodynia in the rat paw. J Neurosci Methods. 1994;53(1):55–63. doi:10.1016/0165-0270(94)90144-9
26. Hargreaves K, Dubner R, Brown F, Flores C, Joris J. A new and sensitive method for measuring thermal nociception in cutaneous hyperalgesia. Pain. 1988;32(1):77–88. doi:10.1016/0304-3959(88)90026-7
27. Kim D, Langmead B, Salzberg SL. HISAT: a fast spliced aligner with low memory requirements. Nat Methods. 2015;12(4):357–360. doi:10.1038/nmeth.3317
28. Huang K, Wu L, Gao Y, et al. Transcriptome Sequencing Data Reveal LncRNA-miRNA-mRNA Regulatory Network in Calcified Aortic Valve Disease. Front Cardiovasc Med. 2022;9:886995. doi:10.3389/fcvm.2022.886995
29. Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nat Methods. 2012;9(4):357–359. doi:10.1038/nmeth.1923
30. Bonfanti C, Rossi G, Tedesco FS, et al. PW1/Peg3 expression regulates key properties that determine mesoangioblast stem cell competence. Nat Commun. 2015;6:6364. doi:10.1038/ncomms7364
31. Shen Y, Gong L, Xu F, et al. Insight into the lncRNA-mRNA Co-Expression Profile and ceRNA Network in Lipopolysaccharide-Induced Acute Lung Injury. Curr Issues Mol Biol. 2023;45(7):6170–6189. doi:10.3390/cimb45070389
32. Tay Y, Rinn J, Pandolfi PP. The multilayered complexity of ceRNA crosstalk and competition. Nature. 2014;505(7483):344–352. doi:10.1038/nature12986
33. Szklarczyk D, Gable AL, Lyon D, et al. STRING v11: protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019;47(D1):D607–D613. doi:10.1093/nar/gky1131
34. Morrison G, Van Langenberg DR, Gibson SJ, Gibson PR. Chronic pain in inflammatory bowel disease: characteristics and associations of a hospital-based cohort. Inflamm Bowel Dis. 2013;19(6):1210–1217. doi:10.1097/MIB.0b013e318280e729
35. Docherty MJ, Jones III RC. Wallace MS. Managing pain in inflammatory bowel disease. Gastroenterol Hepatol. 2011;7(9):592–601.
36. Bielefeldt K, Davis B, Binion DG. Pain and inflammatory bowel disease. Inflamm Bowel Dis. 2009;15(5):778–788. doi:10.1002/ibd.20848
37. Woolf CJ. Central sensitization: implications for the diagnosis and treatment of pain. Pain. 2011;152(3 Suppl):S2–S15. doi:10.1016/j.pain.2010.09.030
38. Li W, Zhang L, Chen S. Spinal Astrocytes in Chronic Visceral Pain. Dev Neurobiol. 2025;85(4):e22996. doi:10.1002/dneu.22996
39. Wang XJ, Li XY, Guo XC, et al. LncRNA-miRNA-mRNA Network Analysis Reveals the Potential Biomarkers in Crohn’s Disease Rats Treated with Herb-Partitioned Moxibustion. J Inflamm Res. 2022;15:1699–1716. doi:10.2147/JIR.S351672
40. Cameron DM, Brennan TJ, Gebhart GF. Hind paw incision in the rat produces long-lasting colon hypersensitivity. J Pain. 2008;9(3):246–253. doi:10.1016/j.jpain.2007.10.017
41. Lopez-Estevez S, Aguilera M, Gris G, de la Puente B, Carceller A, Martinez V. Genetic and Pharmacological Blockade of Sigma-1 Receptors Attenuates Inflammation-Associated Hypersensitivity during Acute Colitis in CD1 Mice. Biomedicines. 2023;11(10):2758. doi:10.3390/biomedicines11102758
42. Zhang C, Gao R, Zhou R, et al. The emerging power and promise of non-coding RNAs in chronic pain. Front Mol Neurosci. 2022;15:1037929. doi:10.3389/fnmol.2022.1037929
43. Irfan J, Febrianto MR, Sharma A, et al. DNA Methylation and Non-Coding RNAs during Tissue-Injury Associated Pain. Int J Mol Sci. 2022;23(2):752. doi:10.3390/ijms23020752
44. Mahurkar-Joshi S, Chang L. Epigenetic Mechanisms in Irritable Bowel Syndrome. Front Psychiatry. 2020;11:805. doi:10.3389/fpsyt.2020.00805
45. Hu C, He M, Xu Q, Tian W. Advances With Non-coding RNAs in Neuropathic Pain. Front Neurosci. 2021;15:760936. doi:10.3389/fnins.2021.760936
46. Zhang X, Zhu L, Wang X, Xia L, Zhang Y. Advances in the role and mechanism of miRNA in inflammatory pain. Biomed Pharmacother. 2023;161:114463. doi:10.1016/j.biopha.2023.114463
47. Sabina S, Panico A, Mincarone P, et al. Expression and Biological Functions of miRNAs in Chronic Pain: a Review on Human Studies. Int J Mol Sci. 2022;23(11):6016. doi:10.3390/ijms23116016
48. Staal RGW, Gandhi A, Zhou H, et al. Inhibition of P2X7 receptors by Lu AF27139 diminishes colonic hypersensitivity and CNS prostanoid levels in a rat model of visceral pain. Purinergic Sig. 2022;18(4):499–514. doi:10.1007/s11302-022-09892-0
49. Kogure Y, Kanda H, Wang S, et al. Daikenchuto attenuates visceral pain and suppresses eosinophil infiltration in inflammatory bowel disease in murine models. JGH Open. 2020;4(6):1146–1154. doi:10.1002/jgh3.12410
50. Qiu C, Yang LD, Yu W, et al. Paeonol ameliorates CFA-induced inflammatory pain by inhibiting HMGB1/TLR4/NF-kappaB p65 pathway. Metab Brain Dis. 2021;36(2):273–283. doi:10.1007/s11011-020-00645-9
51. Senol SP, Temiz-Resitoglu M, Guden DS, Sari AN, Sahan-Firat S, Tunctan B. Suppression of TLR4/MyD88/TAK1/NF-kappaB/COX-2 Signaling Pathway in the Central Nervous System by Bexarotene, a Selective RXR Agonist, Prevents Hyperalgesia in the Lipopolysaccharide-Induced Pain Mouse Model. Neurochem Res. 2021;46(3):624–637. doi:10.1007/s11064-020-03197-7
52. Hockley JR, Gonzalez-Cano R, McMurray S, et al. Visceral and somatic pain modalities reveal Na(V) 1.7-independent visceral nociceptive pathways. J Physiol. 2017;595(8):2661–2679. doi:10.1113/JP272837
53. Li YC, Tian YQ, Wu YY, et al. Upregulation of Spinal ASIC1 and NKCC1 Expression Contributes to Chronic Visceral Pain in Rats. Front Mol Neurosci. 2020;13:611179. doi:10.3389/fnmol.2020.611179
54. Wouters MM, Balemans D, Van Wanrooy S, et al. Histamine Receptor H1-Mediated Sensitization of TRPV1 Mediates Visceral Hypersensitivity and Symptoms in Patients With Irritable Bowel Syndrome. Gastroenterology. 2016;150(4):875–87e9. doi:10.1053/j.gastro.2015.12.034
55. Tsubota M, Matsui K, Nakano M, et al. Essential role of Ca(v)3.2 T-type calcium channels in butyrate-induced colonic pain and nociceptor hypersensitivity in mice. Eur J Pharmacol. 2020;887:173576. doi:10.1016/j.ejphar.2020.173576
56. Chen W, Guo S, Wang S. MicroRNA-16 Alleviates Inflammatory Pain by Targeting Ras-Related Protein 23 (RAB23) and Inhibiting p38 MAPK Activation. Med Sci Monit. 2016;22:3894–3901. doi:10.12659/msm.897580
57. Zhang D, Dong X, Li X, et al. Moxibustion ameliorates chronic inflammatory visceral pain via spinal circRNA-miRNA-mRNA networks: a central mechanism study. Mol Brain. 2024;17(1):23. doi:10.1186/s13041-024-01093-7
58. Xu S, Liu S, Yang J, et al. miR‑3120/Hsc70 participates in forced swim stress‑induced mechanical hyperalgesia in rats in an inflammatory state. Mol Med Rep. 2024;29(1):1. doi:10.3892/mmr.2023.13126
59. Dong W, Wang S, Qian W, Li S, Wang P. Cedrol alleviates the apoptosis and inflammatory response of IL-1beta-treated chondrocytes by promoting miR-542-5p expression. Vitro Cell Dev Biol Anim. 2021;57(10):962–972. doi:10.1007/s11626-021-00620-3
60. Cheng F, Hu H, Sun K, Yan F, Geng Y. miR-455-3p enhances chondrocytes apoptosis and inflammation by targeting COL2A1 in the in vitro osteoarthritis model. Biosci Biotechnol Biochem. 2020;84(4):695–702. doi:10.1080/09168451.2019.1690974
61. Wang Q, Chen Y, Shen X, Chen J, Li Y. Intra-Articular Injection of miR-29a-3p of BMSCs Promotes Cartilage Self-Repairing and Alleviates Pain in the Rat Osteoarthritis. Tissue Eng Regen Med. 2021;18(6):1045–1055. doi:10.1007/s13770-021-00384-7
62. He L, He T, Xing J, et al. Bone marrow mesenchymal stem cell-derived exosomes protect cartilage damage and relieve knee osteoarthritis pain in a rat model of osteoarthritis. Stem Cell Res Ther. 2020;11(1):276. doi:10.1186/s13287-020-01781-w
63. Lance-Jones C, Omelchenko N, Bailis A, Lynch S, Sharma K. Hoxd10 induction and regionalization in the developing lumbosacral spinal cord. Development. 2001;128(12):2255–2268. doi:10.1242/dev.128.12.2255
64. Misra M, Shah V, Carpenter E, McCaffery P, Lance-Jones C. Restricted patterns of Hoxd10 and Hoxd11 set segmental differences in motoneuron subtype complement in the lumbosacral spinal cord. Dev Biol. 2009;330(1):54–72. doi:10.1016/j.ydbio.2009.03.009
65. Ruan Z, Li Y, He R, Li X. Inhibition of microRNA-10b-5p up-regulates HOXD10 to attenuate Alzheimer’s disease in rats via the Rho/ROCK signalling pathway. J Drug Target. 2021;29(5):531–540. doi:10.1080/1061186X.2020.1864739
66. Xu J, Lin N. HOXD10 regulates intestinal permeability and inhibits inflammation of dextran sulfate sodium-induced ulcerative colitis through the inactivation of the Rho/ROCK/MMPs axis. Open Med. 2024;19(1):20230844. doi:10.1515/med-2023-0844
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Identification of Pivotal ceRNA Networks Associated with Stanford-A Aortic Dissection via Integrated Bioinformatics Analysis
Hu Y, Liu Z, Qin Y, Wu N, Yang T, Cheng X, Wang C, Wang X
International Journal of General Medicine 2025, 18:1509-1527
Published Date: 17 March 2025