")
Back to Journals » Infection and Drug Resistance » Volume 11
Inhibitory effects of fluoroquinolone antibiotics on Babesia divergens and Babesia microti, blood parasites of veterinary and zoonotic importance
Authors Rizk MA , AbouLaila M , El-Sayed SAE , Guswanto A, Yokoyama N, Igarashi I
Received 12 December 2017
Accepted for publication 26 April 2018
Published 28 September 2018 Volume 2018:11 Pages 1605—1615
DOI https://doi.org/10.2147/IDR.S159519
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Eric Nulens
Mohamed Abdo Rizk,1,2 Mahmoud AbouLaila,3 Shimaa Abd El-Salam El-Sayed,1,4 Azirwan Guswanto,1,5 Naoaki Yokoyama,1 Ikuo Igarashi1
1National Research Center for Protozoan Diseases, Obihiro University of Agriculture and Veterinary Medicine, Inada‑Cho, Obihiro, Hokkaido, Japan; 2Department of Internal Medicine and Infectious Diseases, Faculty of Veterinary Medicine, Mansoura University, Mansoura, Egypt; 3Department of Parasitology and Pathology, Faculty of Veterinary Medicine, Damanhour University, Damanhour, ElBehera, Egypt; 4Department of Biochemistry and Chemistry of Nutrition, Faculty of Veterinary Medicine, Mansoura University, Mansoura, Egypt; 5Veterinary Services Section, Disease Investigation Center, Subang, West Java, Indonesia
Aim: This study aimed to evaluate the inhibitory effects of fluoroquinolone antibiotics, including enrofloxacin, enoxacin, trovafloxacin, norfloxacin, and ofloxacin, on the in vitro and in vivo growth of Babesia divergens and Babesia microti parasites, respectively.
Materials and methods: The in vitro and in vivo inhibitory effects of fluoroquinolone antibiotics against B. divergens and B. microti, respectively were evaluated using fluorescence-based assay. Additionally, combination therapies of highly effective fluoroquinolone antibiotics (enrofloxacin, enoxacin, and trovafloxacin) with diminazene aceturate, luteolin, or pyronaridine tetraphosphate were tested on the in vitro cultures of B. divergens.
Results: Enrofloxacin, trovafloxacin, and enoxacin were the most effective fluoroquinolones against the in vitro growth of B. divergens, followed by norfloxacin and ofloxacin. Furthermore, a combination of enoxacin or trovafloxacin with either diminazene aceturate, luteolin, or pyronaridine tetraphosphate significantly enhanced the inhibitory effect on the growth of B. divergens in in vitro cultures. In mice infected by B. microti, enoxacin and diminazene aceturate combination therapy exhibited a potential antibabesial effect.
Conclusion: These results suggest that safe and cheap fluoroquinolone, such as enoxacin, might be used for the treatment of clinical cases caused by Babesia spp. in animals or humans.
Keywords: Babesia, fluoroquinolones, in vitro, in vivo
Introduction
Babesia is a protozoan parasite transmitted by ticks; it is a major pathogen that infects the erythrocytes of a wide range of economically valuable animals as well as human erythrocytes.1 Babesiosis is a severe, rapidly fatal disease typified by fever, hemolytic anemia, and hemoglobinuria that causes severe economic losses in the livestock industry.1,2 Babesia microti and Babesia divergens are blood parasites that infect rodents and cattle, respectively, and have zoonotic importance.1 B. microti is the main etiologic agent recognized for babesiosis in humans in the USA and B. divergens is a serious cause of babesiosis in Europe.3
Recently, antibabesial drugs commonly used in veterinary medicine have shown problems regarding parasite resistance and toxicity to the host.4 At the same time, many of the most effective drugs against B. divergens have been withdrawn from the market due to safety or residue problems.3 Moreover, therapeutic failures have been reported in some severe cases of human babesiosis after the administration of clindamycin, azithromycin, quinine, and tetracycline.3 Therefore, there is an urgent need to develop more effective drugs against Babesia parasites that have both veterinary and zoonotic importance. Fluoroquinolones are bactericidal drugs that have the ability to interfere with the replication of an organism’s DNA by inhibiting pathogen DNA gyrases and topoisomerases.5 Following this pattern, two gene-encoding gyrases (A and B) are present within apicomplexan parasites, with the remarkable exception of Cryptosporidium sp., and are driven to their essential plastid-like organelles, the apicoplasts.6 In fact, apicomplexan apicoplast organelles play a critical role in cell survival in the later stages of parasite evolution.6 Taken together, there is no ortholog to the apicoplast DNA gyrase of apicomplexan parasites in mammals;6,7 thus, this enzyme provides an attractive target for drug development. Additionally, DNA topoisomerases (Top) are important enzymes required to produce a more relaxed DNA and thus play a critical role in DNA packaging.8 Therefore, in the present study, fluoroquinolone antibiotics, including enrofloxacin, enoxacin, trovafloxacin, norfloxacin, and ofloxacin, were selected to evaluate their inhibitory effects against the growth of B. divergens in vitro and B. microti in vivo.
Materials and methods
Ethical approval
All experimental protocols in this study were approved by the Animal Care and Use Committee, Obihiro University of Agriculture and Veterinary Medicine (approval no. 27-65). All experiments were conducted in accordance with the Fundamental Guidelines for Proper Conduct of Animal Experiment and Related Activities in Academic Research Institutions under the jurisdiction of the Ministry of Education, Culture, Sports, Science and Technology, Japan.
Parasites
A German bovine strain of B. divergens9 was used for the in vitro experiment and cultivated in purified bovine red blood cells (RBCs) using a microaerophilic, stationary-phase culture system.9 The Munich strain of B. microti10 was used for the in vivo experiment.
Chemical reagents
SYBR Green I (SGI) nucleic acid stain (10,000×; Lonza, Rockland, ME, USA) was stored at –20°C and thawed before use. A lysis buffer consisting of Tris (130 mM; pH 7.5), EDTA (10 mM), saponin (0.016%; W/V), and TritonX-100 (1.6%; V/V) was prepared in advance and stored at 4°C. Diminazene aceturate (DA; Novartis International AG, Basel, Switzerland), tetracycline hydrochloride, and chloroquine diphosphate (both drugs from Sigma-Aldrich Co., St Louis, MO, USA) were used as positive control drugs. Enrofloxacin (Figure 1A), enoxacin (Figure 1B), trovafloxacin (Figure 1C), norfloxacin (Figure 1D), ofloxacin (Figure 1E), luteolin, or pyronaridine tetraphosphate (PYR) (all drugs from Sigma-Aldrich Co.) was prepared as a 100 mM stock solution and stored at -20°C until use.
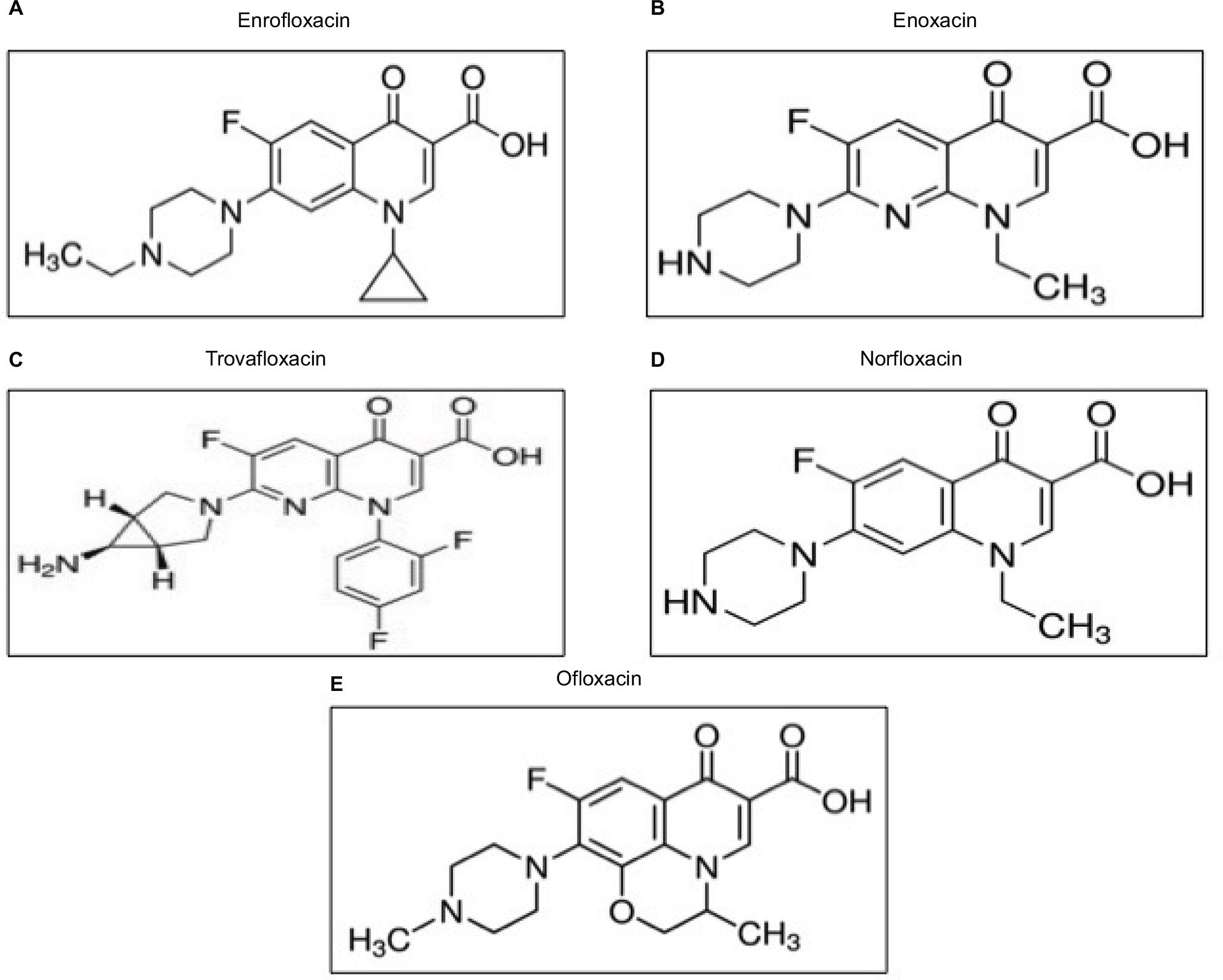 | Figure 1 Chemical structures of fluoroquinolone antibiotics used in this study: (A) enrofloxacin, (B) enoxacin, (C) trovafloxacin, (D) norfloxacin, and (E) ofloxacin. |
Evaluation of fluoroquinolone antibiotics against B. divergens in vitro
The in vitro Babesia fluorescence assay (BFA) was performed as previously described by Rizk et al.9 Briefly, Roswell Park Memorial Institute (RPMI) 1640 medium (Sigma-Aldrich Co.) alone or mixed with the indicated concentrations, 0.25, 0.5, 1, 5, and 10 µM for DA or 0.5, 10, 50, 100, and 400 µM for other fluoroquinolone antibiotics (enrofloxacin, enoxacin, trovafloxacin, norfloxacin, and ofloxacin), was used for the cultivation of parasitized RBCs (pRBCs) in double 96-well plates (Nunc, Roskilde, Denmark) at 5% hematocrit (HCT) for 4 days in triplicate wells for each concentration of the drug. On the fourth day of culture, a lysis buffer containing 2×SGI was added to each well of the first 96-well plate with gentle mixing. For a viability test, on the fourth day of treatment, 3.5 µL of the culture of untreated pRBCs, treated pRBCs, and uninfected RBCs on the second plate was removed and replaced with parasite-free bovine RBCs (3.5 µL) and suspended in 100 µL of a fresh growth medium without drugs. Then the plate was incubated at 37°C for the next 4 days. Next, the fluorescence values were determined using a fluorescence plate reader (Fluoroskan Ascent; Thermo Electron Informatics, Philadelphia, PA, USA) at 485 nm and 518 nm excitation and emission wavelengths, respectively. Each experiment was repeated three times.
In vitro drug combination test
Combination therapies of highly effective fluoroquinolone antibiotics (enrofloxacin, enoxacin, and trovafloxacin) with DA, luteolin, or PYR were tested on the in vitro cultures of B. divergens. Fluoroquinolone antibiotics/DA, luteolin, or PYR combinations (M1, M2, M3, and M4, respectively) were prepared as previously described11,12 with some modifications. Combinations were based on the calculated IC50 values obtained from the in vitro BFA (Table S1). Three separate trials were performed, consisting of triplicate experiments for each drug combination over a period of 4 days using 5% HCT. On the fourth day of culture, the fluorescence values were calculated after adding a lysis buffer containing 2×SGI to each well on the 96-well plate.
Chemotherapeutic evaluation of fluoroquinolone antibiotics in mice
Fluoroquinolone antibiotics were administered in an in vivo BFA for B. microti in BALB/c mice (CLEA, Tokyo, Japan), following a method described by Rizk et al.10 Thirty-five 8-week-old female mice were kept under specific pathogen-free conditions and divided equally into seven groups. Mice in the first six groups were inoculated intraperitoneally with 1×107 B. microti-infected RBCs, while the mice in the seventh group remained without infection and were used as a blank control. When the infected mice showed ~1% parasitemia, mice in the experimental groups received daily injections of the specific drugs for 5 days. All administered drugs were dissolved in double-distilled water (DDW), and DDW was administered subcutaneously to the mice in the first group as a placebo control.10 DA was administered to the second group subcutaneously at a dose rate of 25 mg⋅kg-1 as a reference drug control. Mice in the third, fourth, and fifth groups received daily subcutaneous injections of nontoxic doses15 of enoxacin (100 mg⋅kg-1), norfloxacin (150 mg⋅kg-1), and ofloxacin (700 mg⋅kg-1), respectively. Mice in the sixth group were treated with a combination therapy consisting of 50 mg⋅kg-1 enoxacin and 10 mg⋅kg-1 DA. Both drugs were administered subcutaneously in the same inoculation period. The emitted fluorescence signals were used as an indicator of the parasitemia percentages in all mice and monitored every 48 hours until 20 days post-inoculation or the cessation of parasitemia.
Role of fluoroquinolone antibiotics in the regression of anemia associated with Babesia
HCT values, hemoglobin levels, RBC counts, mean corpuscular volume, mean corpuscular hemoglobin, and red blood cell distribution width were used as indicators of the development of anemia in mice treated with fluoroquinolone antibiotics. Ten microliters of venous tail blood samples was collected from all mice as previously described. In all groups, the hematological parameters were monitored every 96 hours using the Celltac α MEK-6450 automatic hematology analyzer (Nihon Kohden Corporation, Tokyo, Japan).
Polymerase chain reaction (PCR) detection of B. microti in mice treated with DA and enoxacin combination therapy
Nested PCR assays targeting the B. microti small subunit rRNA (ss-rRNA) gene were used to detect parasite DNA in blood and tissue (spleen, heart, lung, and kidney) samples collected from mice treated with an enoxacin/DA combination, DA alone, and DDW (positive control) on day 20 post-infection (pi). DNA was extracted from the tissues collected from treated mice using a NucleoSpin tissue kit (Macherey-Nagel, Düren, Germany). DNA extracted from the blood of B. microti-infected mice treated with DDW (positive control) using a QIAamp DNA Blood Mini Kit (Qiagen, Tokyo, Japan) served as a positive control. PCR cycling was performed as previously described by Rizk et al.10
Statistical analysis
The differences in the fluorescence values of the in vitro cultures and among groups for the in vivo studies were analyzed with a statistical software program (GraphPad Prism version 5.0 for Windows; GraphPad Software, Inc., San Diego, CA, USA), using an independent Student’s t-test and one-way analysis of variance (ANOVA). A P-value of <0.05 was considered statistically significant. The regrowth of the parasite in the viability test was determined based on the statistically significant differences between drug-treated and positive control groups.9
Results
In vitro inhibitory effect against B. divergens
The in vitro growth of B. divergens was significantly inhibited (P < 0.05) by 500 nM and 50, 10, 50, and 400 µM treatments of enrofloxacin, enoxacin, trovafloxacin, norfloxacin, and ofloxacin, respectively (Figure 2). Likewise, 500 nM treatments with DA or chloroquine diphosphate significantly inhibited (P < 0.05) the in vitro growth of B. divergens (data not shown). Enrofloxacin, trovafloxacin, and enoxacin exhibited the highest activity against the in vitro growth of B. divergens, followed by norfloxacin and ofloxacin (Table 1). Subsequent viability tests showed that there was no regrowth of the parasite with 50 µM enrofloxacin and trovafloxacin (Table 2). Additionally, treatment of the B. divergens in vitro culture with 100 µM enoxacin or tetracycline hydrochloride prevented parasite regrowth (Table 2). Similarly, 400 µM norfloxacin and ofloxacin prevented the in vitro regrowth of B. divergens (Table 2). On the other hand, 0.25 µM DA and 0.5 µM chloroquine diphosphate were sufficient to inhibit the regrowth of B. divergens (Table 2).
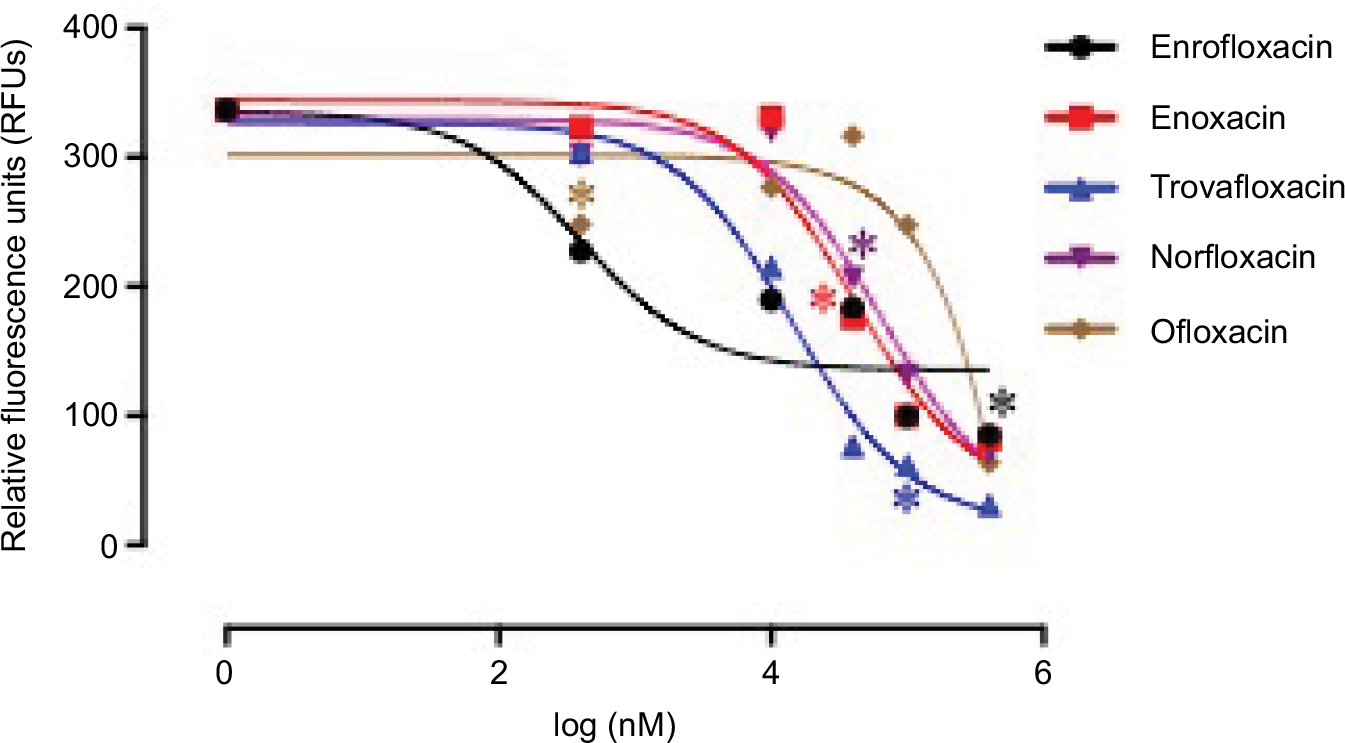 | Figure 2 Correlation between relative fluorescence units (RFUs) and the log concentrations of fluoroquinolone antibiotics (nM) in Babesia divergens on the fourth day of treatment. Each value represents the mean of triplicate wells after subtraction of the background fluorescence for non-parasitized red blood cells. |
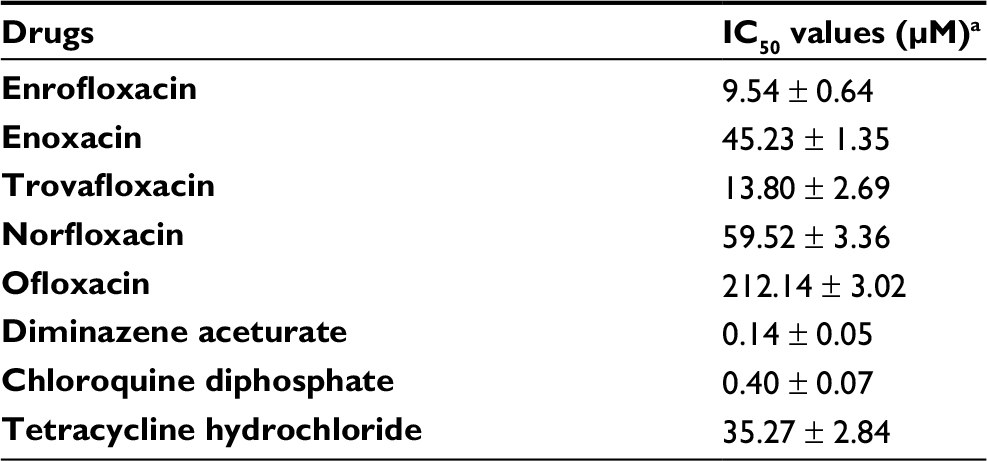 | Table 1 IC50 values of fluoroquinolone antibiotic drugs evaluated for Babesia divergens parasite Notes: aIC50 values for each drug were calculated on the fourth day of the in vitro culture using a Babesia fluorescence assay (BFA) in three separate experiments. Each drug concentration was made in triplicate in each experiment, and the final obtained IC50 values were the mean ± SD of values obtained from three separate experiments. |
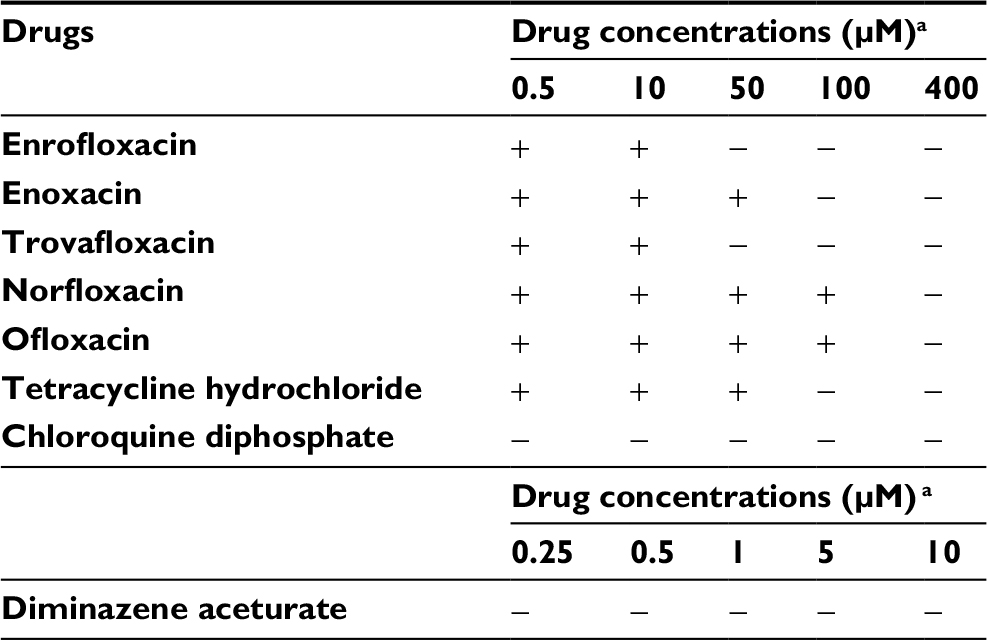 | Table 2 Viability test results of fluoroquinolone antibiotic drugs evaluated for Babesia divergens parasite Note: aEach value was calculated using a Babesia fluorescence assay (BFA) in three separate experiments. Each concentration of the drug was made in triplicate in each experiment. +, viable; −, dead. |
Drug combination test
Fluoroquinolone antibiotic drugs with potential in vitro antibabesial effects were selected to examine their inhibitory effect when used in combination with DA, luteolin, and PYR. The combined application of fluoroquinolone antibiotics/DA revealed an additive effect in all used combinations, except with the combination M1 for trovafloxacin and DA where IC50 value was three-fourth the IC50 values of both drugs, that is, a synergetic effect was observed (Table 3). Similarly, treatment of the B. divergens parasite with combined therapies consisting of trovafloxacin and PYR significantly enhanced the growth inhibition with a synergetic effect, even at the M2 concentration that consisted of three-fourth the IC50 of trovafloxacin and one-half the IC50 of PYR (Table 3). The other combinations of either fluoroquinolone antibiotics/PYR or fluoroquinolone antibiotics/luteolin exhibited additive or indifferent drug interactions (Tables 3 and S2).
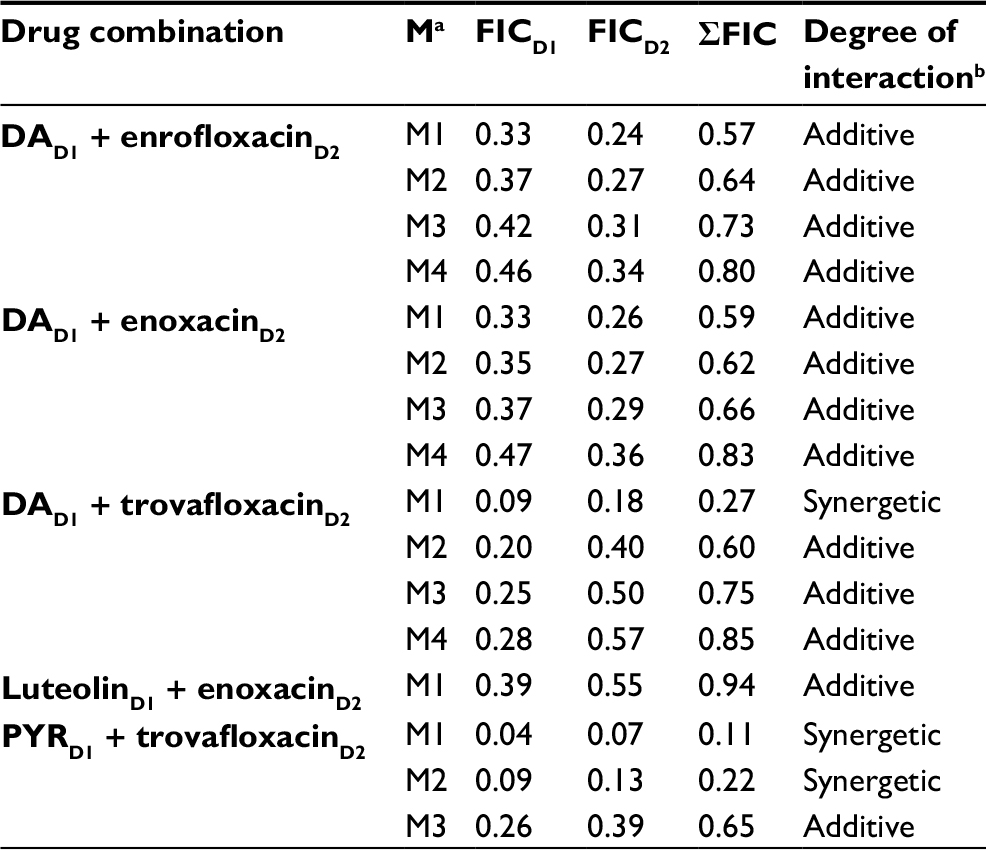 | Table 3 Synergetic and additive interactions of diminazene aceturate (DA), luteolin, and pyronaridine tetraphosphate (PYR) in combination with enrofloxacin, enoxacin, or trovafloxacin on Babesia divergens parasite on the fourth day of treatment Notes: aM1–M4 refer to the combinations of fluoroquinolone antibiotic drugs combined with different antibabesial drugs; bthe degree of drug interaction was determined based on the following fractional inhibitory concentration (FIC) index: ≤0.5 (synergetic), >0.5–1 (additive), >1 to <2 (indifferent), and ≥2 (antagonistic). FICD1 refers to the fractional inhibitory concentration of DA, luteolin, or PYR. FICD2 refers to the fractional inhibitory concentration of fluoroquinolone. |
In vivo effect of fluoroquinolone antibiotics on B. microti infection
To further validate fluoroquinolone antibiotics as antibabesial drugs, an in vivo study was performed using B. microti in a mouse model in two separate trials. Fluorescence values were significantly inhibited (P < 0.05) in mice treated with DA combined with enoxacin from days 4 to 16 pi as compared to those of the controls (Figure 3), while treatment with either enoxacin alone (100 mg⋅kg-1) or norfloxacin (150 mg⋅kg-1) produced significant inhibition of fluorescence values (P < 0.05) from days 4 to 12 pi as compared to those of the controls (Figure 3). The fluorescence values obtained from mice treated with 700 mg⋅kg-1 ofloxacin were significantly inhibited (P < 0.05) from 6 to 16 days pi as compared to the control mice (Figure 3).
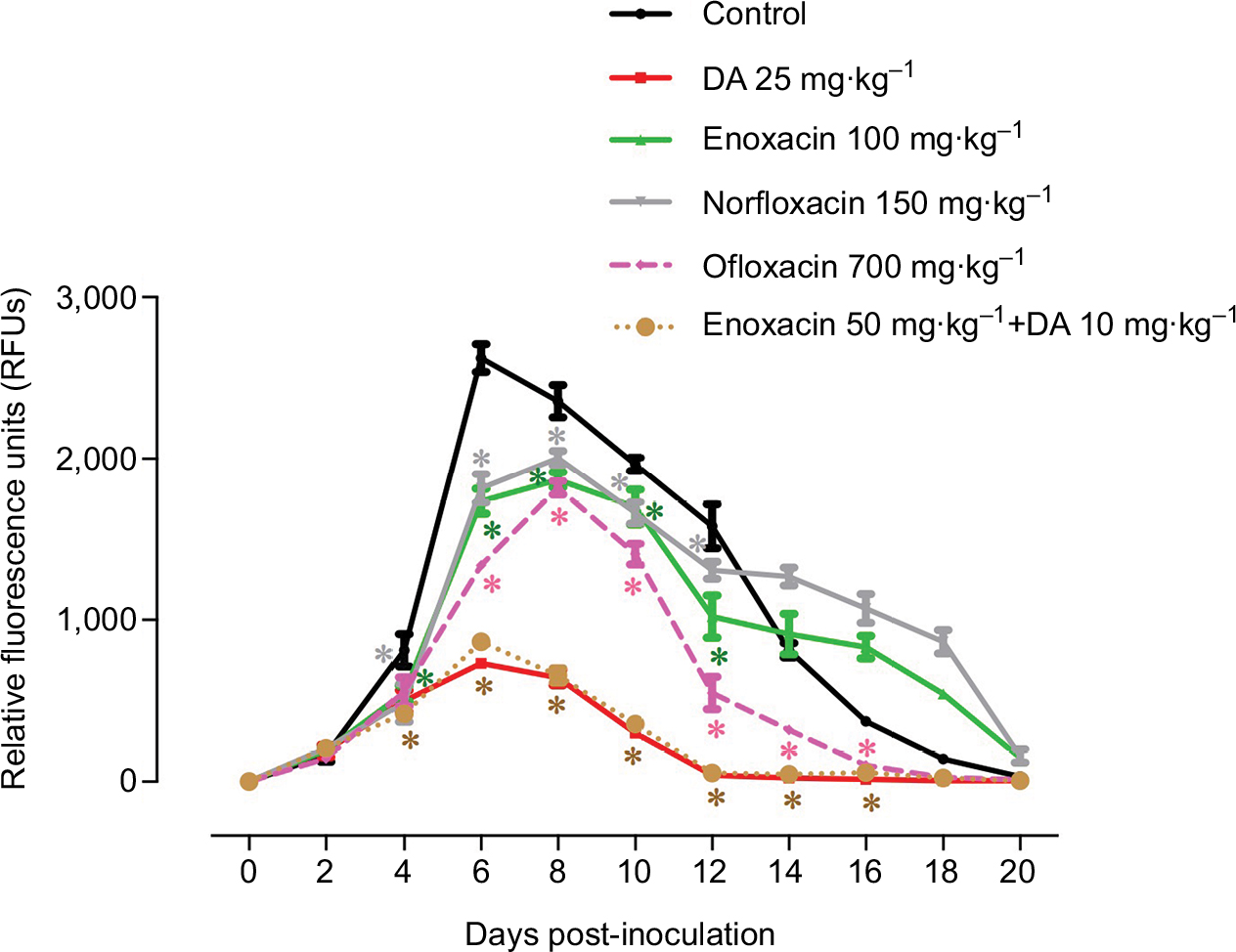 | Figure 3 Inhibitory effect of fluoroquinolone antibiotics, diminazene aceturate (DA), and the combination of enoxacin and DA on the growth of Babesia microti. Each value represents the mean ± SD of five mice per experimental group. Asterisks indicate significant differences (*P < 0.05) from day 4 to day 16 post-inoculation between the fluoroquinolone antibiotic-treated and control groups. |
Control mice treated with DDW and mice treated with either DA alone or DA combined with enoxacin showed peak fluorescence signals at day 6 pi, whereas other treated mice exhibited peak fluorescence signals at day 8 pi (Figure 3). Notably, mice treated with a combined formula (enoxacin/DA) exhibited significantly inhibited fluorescence values similar to those obtained from mice treated with 25 mg⋅kg-1 DA (Figure 3). These results indicate that a combination formula from DA and enoxacin may be a promising treatment for Babesia spp. that infect humans or animals.
Role of fluoroquinolone antibiotics in the regression of anemia in treated mice
The role of fluoroquinolone antibiotics in mouse recovery from anemia was evaluated by monitoring changes in hematological variables. The mice treated with combination therapy consisting of enoxacin and DA exhibited quick recovery from hemolytic and macrocytic anemia associated with babesiosis (Figures 4 and 5). These findings confirm the potential antibabesial effect of this combination therapy.
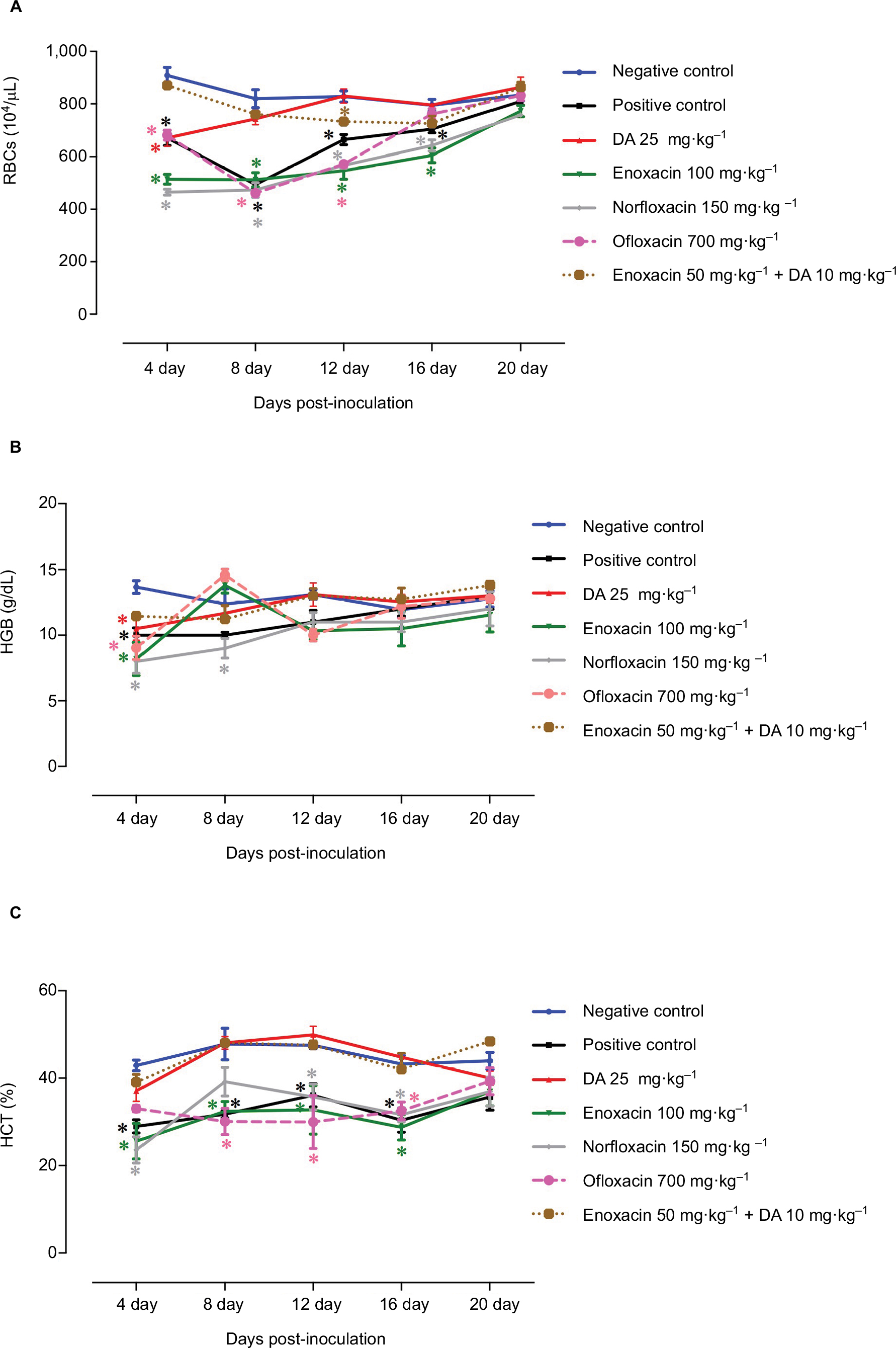 | Figure 4 Anemia monitoring in mice treated with fluoroquinolone antibiotics: (A) RBCs, (B) hemoglobin (HGB), and (C) hematocrit (HCT). Each value represents the mean ± SD of five mice per experimental group. Asterisks indicate a significant difference (*P < 0.05) between the treated or infected mice and the uninfected mice. Abbreviation: DA, diminazene aceturate. |
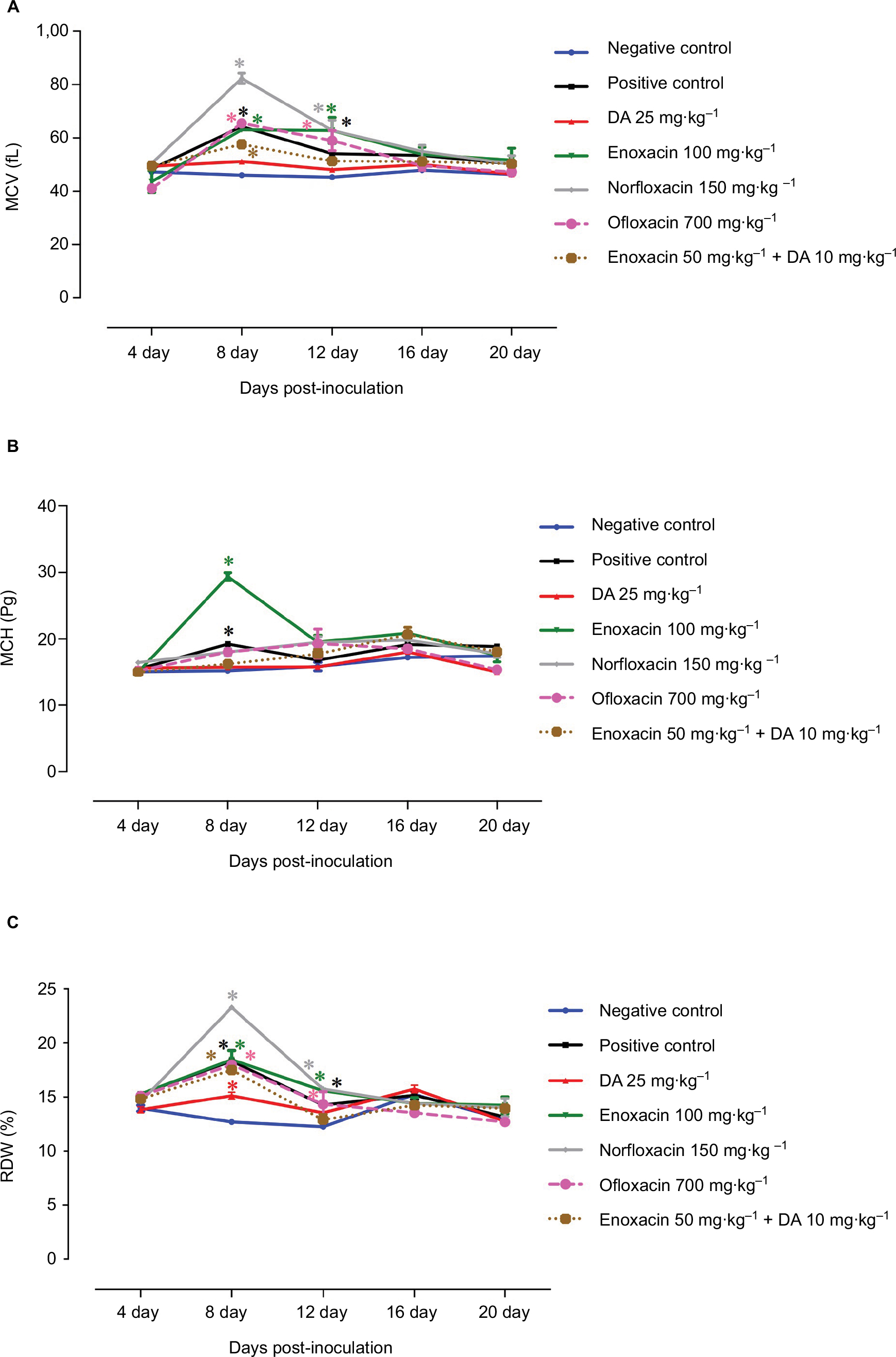 | Figure 5 Changes in mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), and red blood cell distribution width (RDW) in mice treated with fluoroquinolone antibiotics: (A) MCV, (B) MCH, and (C) RDW. Each value represents the mean ± SD of five mice per experimental group. Asterisks indicate a significant difference (*P < 0.05) between the treated or infected mice and the uninfected mice. Abbreviation: DA, diminazene aceturate. |
Detection of B. microti nucleic acid remnants in the different organs of treated mice
A nested PCR was applied to DNA samples extracted from the different organs of treated mice on day 20 pi. PCR amplification detected the B. microti ss-rRNA gene in the spleen, heart, lungs, and kidneys of mice treated with DDW or DA (Figure 6). Interestingly, parasite nucleic acid was not detected in the lungs of mice treated with an enoxacin and DA combination, while the B. microti ss-rRNA gene was detected in other organs (Figure 6).
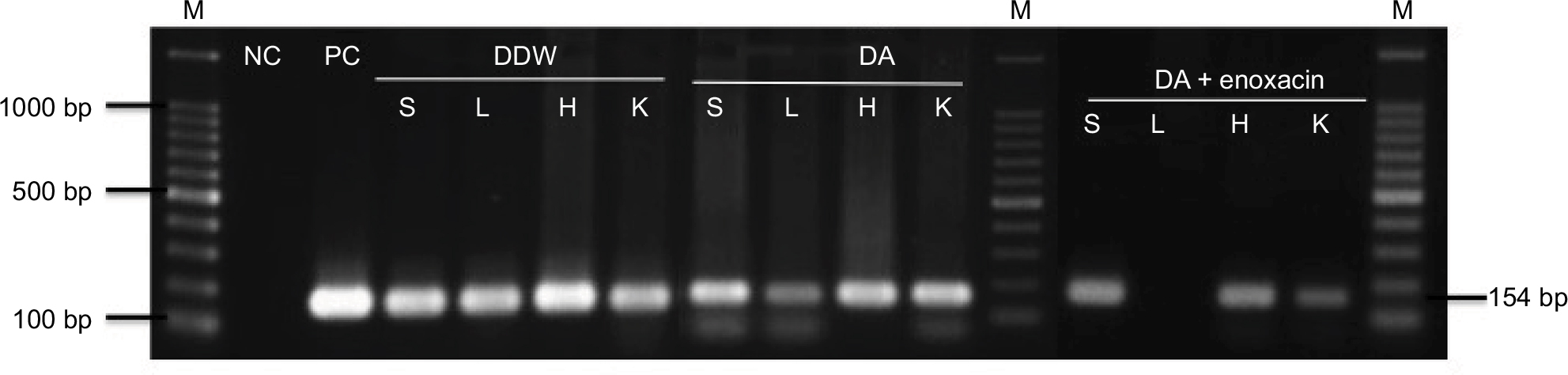 | Figure 6 PCR of the ss-rRNA gene in different organs of Babesia microti-infected mice treated with DDW (positive control), 25 mg⋅kg–1 diminazene aceturate (DA), and DA (10 mg⋅kg–1) combined with enoxacin (50 mg⋅kg–1). Note: M indicates a 100 bp DNA ladder. Abbreviations: NC, negative control; PC, positive control; DDW, double-distilled water; S, spleen; L, lung; H, heart; K, kidney; DA, diminazene aceturate; PCR, polymerase chain reaction. |
Discussion
Fluoroquinolones were proven to have antimalarial activity in vitro13 and in vivo.14 Considering the close biological similarities between Plasmodium and Babesia parasites, there is a robust motive for studying the chemotherapeutic potential of fluoroquinolones for their inhibitory effect on babesiosis in vitro and in vivo.
In the current study, the IC50 value of trovafloxacin for B. divergens was lower than those obtained against chloroquine-sensitive (3D7) and chloroquine-resistant (NF54-R) strains of Plasmodium falciparum in vitro.15 On the other hand, the norfloxacin IC50 value for B. divergens was higher than those obtained against chloroquine-sensitive (3D7) and chloroquine-resistant (NF54-R) strains of P. falciparum in vitro.15 The enoxacin and ofloxacin IC50 values for B. divergens were lower than those obtained against a chloroquine-sensitive (3D7) strain of P. falciparum in vitro.15 However, both drugs exhibited IC50 values for B. divergens higher than those obtained against a chloroquine-resistant (NF54-R) strain of P. falciparum in vitro.15 The IC50 of enoxacin for B. divergens was higher than those previously calculated against Babesia bovis, Babesia bigemina, Theileria equi, and Babesia caballi,16 which might be attributed to the enoxacin resistance of the B. divergens strain (Germany strain) used. Enrofloxacin, enoxacin, trovafloxacin, and norfloxacin exhibited lower IC50 values for B. divergens than those obtained with N-acetyl-l-cysteine.17 On the other hand, fluoroquinolone antibiotics exhibited higher IC50 values for B. divergens than those obtained with PYR, luteolin, and actinonin.9
The in vivo evaluation of fluoroquinolone against B. microti in mice demonstrated that 100 mg⋅kg-1 enoxacin, 150 mg⋅kg-1 norfloxacin, and 700 mg⋅kg-1 ofloxacin caused 34%, 31%, and 49% inhibition rates, respectively, at day 6 pi. Eight days pi, the inhibition rates caused by 100 mg⋅kg-1 enoxacin, 150 mg kg-1 norfloxacin, and 700 mg⋅kg-1 ofloxacin were 21%, 15%, and 23%, respectively. These results are lower than the 77.5% inhibition for 500 mg⋅kg-1 thiostrepton, 68.5% for 500 mg⋅kg-1 clindamycin, and 58.3% and 72.4% for 30 and 100 mg⋅kg-1 allicin, respectively.11,18
Enoxacin is a broad-spectrum antibacterial agent used to treat a wide variety of infections, including gastroenteritis, respiratory tract infections,19 and urinary tract infections.20 The drug is also characterized by its rapid and thorough absorption from the gastrointestinal tract after administration, with wide distribution throughout the body.20,21 Of note, the tissue concentrations of enoxacin usually exceed its serum concentrations, and the serum-elimination half-life of this antibacterial agent with normal renal function is ~6 hours,21 a fact that ensures the continuous effect of the drug over time. Therefore, combination therapy consisting of low doses of enoxacin and a commonly used antibabesial agent such as DA was tested in mice infected by the B. microti parasite. The results revealed that an enoxacin/DA combination seems to be more effective than fluoroquinolones alone, even though treatment with reduced doses of both chemicals was administered. A combination of 50 mg⋅kg–1 enoxacin and 10 mg⋅kg-1 DA resulted in 67% inhibition at day 6, as compared with 72% inhibition in the presence of a standard dose (25 mg⋅kg-1) of DA at day 6 pi. However, at day 8 pi, the combined therapy caused a 72% inhibition rate, whereas 25 mg⋅kg-1 DA caused a 72.5% inhibition rate. These results are higher than the 70% inhibition rates for clindamycin combined with the natural product quinine22 and the 56.35% and 53.25% inhibition rates for 85 mg⋅kg-1 PYR combined with 10 mg⋅kg-1 DA.10
Mice treated either with DA alone or with lower doses of an enoxacin and DA combination exhibited quick recovery from this anemia, indicating the potential role of this combination therapy in recovery from the hemolytic macrocytic anemia caused by babesiosis. Moreover, insignificant emitted fluorescence signals were obtained from enoxacin/DA-treated mice on day 20 pi, and the B. microti ss-rRNA gene was not detected in the lungs of mice treated with the enoxacin/DA combination. However, a remnant of the parasite’s nucleic acid was detected in the lungs of mice treated with DA alone, indicating the promising antibabesial effect of such a combination. Nevertheless, parasite DNA was detected in other organs examined via PCR. The short period of the experiment (20 days) may explain the presence of DNA remnants in mouse organs. Therefore, further studies are required to evaluate the inhibitory effect of enoxacin in combination with other antibabesial drugs such as clofazimine,23 PYR, or luteolin9,10,16 against the growth of Babesia and Theileria parasites for more prolonged periods of infection. Furthermore, future studies are warranted to explore why the DA and enoxacin combination therapy cleared the infection from the lungs of B. microti-infected mice, while the parasite DNA was still detected in the other organs of infected mice using a PCR assay.
Currently, the treatment of human babesiosis depends on the administration of combination therapies, such as azithromycin with atovaquone or clindamycin with quinine. However, a failure to recover is reported in some severe cases with the development of drug-resistant parasites.3 Therefore, the development of novel drugs and a combination formula for treating the infection is required. Safe and cheap fluoroquinolones, such as enoxacin, might be an alternative chemotherapeutic agent against human babesiosis caused by B. microti or B. divergens.
Limitations
The limitations of this study should be mentioned. In the present study, the inhibitory effects of selected fluoroquinolones against the in vitro growth of Babesia parasites were examined when used in combination with DA, luteolin, and PYR. The synergetic or antagonistic relationships between these antibabesial drugs and either fluoroquinolones (norfloxacin and ofloxacin) or lower doses of currently approved antibabesial drugs in humans, such as azithromycin, were not evaluated against the growth of B. microti in mice. Future studies are warranted to evaluate the potential of these combinations for B. microti- and/or B. rodhaini-infected mice. Successful application of this combination could reduce the potential side effect of currently used antibabesials, such as the cardiotoxicity of azithromycin.
Conclusion
In this study, the inhibitory effects of fluoroquinolones have been demonstrated against B. divergens in vitro and against B. microti in mice. The results revealed that enoxacin, enrofloxacin, and trovafloxacin exhibited the highest activity against the in vitro growth of B. divergens, while enoxacin/DA combination therapy exhibited a potential antibabesial effect in B. microti-infected mice. Such findings suggest that an enoxacin/DA combination formula might be useful for the treatment of infection caused by Babesia spp.
Data sharing statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Acknowledgments
This study was supported by the Ministry of Education, Culture, Sports, Science and Technology of Japan and Medicines for Malaria Venture (MMV) Challenge Grant.
Author contributions
Conceived and designed the experiments: MAR, MA, SAE-SE-S, II. Performed the experiments: MAR, MA, SAE-SE-S. Analyzed the data: MAR, AG. Contributed reagents/materials/analysis tools: MAR, NY, II. Wrote the manuscript: MAR, II. All authors contributed toward data analysis, drafting and critically revising the paper and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
Uilenberg G. Babesia – a historical overview. Vet Parasitol. 2006;31:3–10. | ||
Homer JM, Aguilar-Delfin I, Telford SR, et al. Babesiosis. Clin Microbiol Rev. 2000;13:451–469. | ||
Zintl A, Mulcahy G, Skerrett HE, et al. Babesia divergens, a bovine blood parasite of veterinary and zoonotic importance. Clin Microbiol Rev. 2003;16:622–636. | ||
Mosqueda J, Olvera-Ramirez A, Aguilar-Tipacamu G, et al. Current advances in detection and treatment of babesiosis. Curr Med Chem. 2012;19:1504–1518. | ||
Drlica K. Mechanism of fluoroquinolone action. Curr Opin Microbiol. 1999;2:504–508. | ||
McFadden GI, Roos DS. Apicomplexan plastids as drug targets. Trends Microbiol. 1999;7:328–333. | ||
Morrison DA. Evolution of the Apicomplexa. Where are we now? Trends Parasitol. 2009;25:375–382. | ||
García-Estrada C, Prada CF, Fernández-Rubio C, et al. DNA topoisomerases in apicomplexan parasites: promising targets for drug discovery. Proc Biol Sci. 2010;277:1777–1787. | ||
Rizk MA, El-Sayed SA, AbouLaila M, et al. Large-scale drug screening against Babesia divergens parasite using a fluorescence-based high-throughput screening assay. Vet Parasitol. 2016;227:93–97. | ||
Rizk MA, El-Sayed SAE, AbouLaila M, et al. Performance and consistency of a fluorescence-based high-throughput screening assay for use in Babesia drug screening in mice. Sci Rep. 2017;7:12774. | ||
Salama AA, AbouLaila M, Terkawi MA, et al. Inhibitory effect of allicin on the growth of Babesia and Theileria equi parasites. Parasitol Res. 2014;113:275–283. | ||
Suleman N, Kalhapure RS, Mocktar C, et al. Silver salts of carboxylic acid terminated generation 1 poly (propyl ether imine) (PETIM) dendron and dendrimers as antimicrobial agents against S. aureus and MRSA. RSC Advances. 2015;5:34967–34978. | ||
Hamzah J, Skinner-Adams T, Davis TME. In vitro antimalarial activity of trovafloxacin, a fourth-generation fluoroquinolone. Acta Trop. 2000;74:39–42. | ||
Watt G, Shanks GD, Einstein MD, et al. Ciprofloxacin treatment of drug-resistant falciparum malaria. J Infect Dis. 1991;164:602–604. | ||
Mahmoudi N, Ciceron L, Franetich JF, et al. In vitro activities of 25 quinolones and fluoroquinolones against liver and blood stage Plasmodium spp. Antimicrob Agents Chemother. 2003;47:2636–2639. | ||
Rizk MA, El-Sayed SA, Terkawi MA, et al. Optimization of a fluorescence-based assay for large-scale drug screening against Babesia and Theileria parasites. PLoS One. 2015;10:e0125276. | ||
Rizk MA, El-Sayed SAE, AbouLaila M, et al. Evaluation of the inhibitory effect of N-acetyl-L-cysteine on Babesia and Theileria parasites. Exp Parasitol. 2017;179:43–48. | ||
AbouLaila M, Munkhjargal T, Sivakumar T, et al. Apicoplast-targeting antibacterials inhibit the growth of Babesia parasites. Antimicrob Agents Chemother. 2012;56:3196–3206. | ||
Wise R, Lockley R, Dent J, et al. Pharmacokinetics and tissue penetration of enoxacin. Antimicrob Agents Chemother. 1984;26:17–19. | ||
Rafalsky V, Andreeva I, Rjabkova E, et al. Quinolones for uncomplicated acute cystitis in women. Cochrane Database Syst Rev. 2006;3:CD003597. | ||
Wise R, Lister D, McNulty CA, et al. The comparative pharmacokinetics and tissue penetration of four quinolones including intravenously administered enoxacin. Infection. 1986;14:196–202. | ||
Marley SE, Eberhard ML, Steurer FJ, et al. Evaluation of selected antiprotozoal drugs in the Babesia microti hamster model. Antimicrob Agents Chemother. 1997;41:91–94. | ||
Tuvshintulga B, AbouLaila M, Davaasuren B, et al. Clofazimine inhibits the growth of Babesia and Theileria parasites in vitro and in vivo. Antimicrob Agents Chemother. 2016;60:2739–2746. |
Supplementary materials
 | Table S1 Concentrations of fluoroquinolone antibiotic drugs combined with diminazene aceturate, luteolin, and pyronaridine tetraphosphate applied to the cultures of Babesia divergens parasite Note: aM1–M4 refer to the combinations of fluoroquinolone antibiotic drugs combined with different antibabesial drugs; bcombinations were based on the calculated IC50 values obtained from the in vitro fluorescence-based assay. |
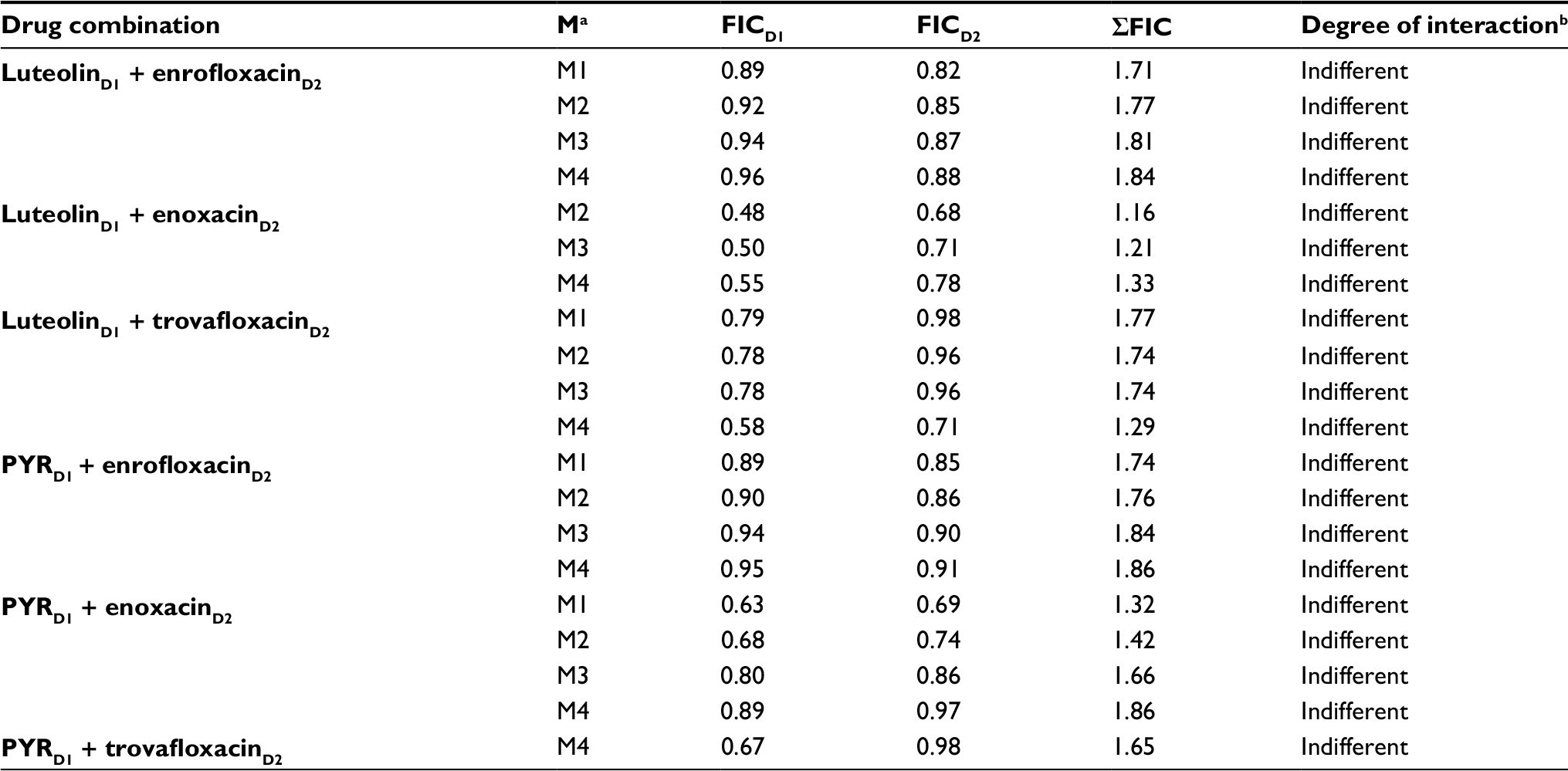 | Table S2 Indifferent interaction of diminazene aceturate (DA), luteolin, and pyronaridine tetraphosphate (PYR) in combination with enrofloxacin, enoxacin, or trovafloxacin on Babesia divergens parasite on the fourth day of treatment Notes: aM1–M4 refer to the combinations of fluoroquinolone antibiotic drugs combined with different antibabesial drugs; bthe degree of drug interaction was determined based on the following fractional inhibitory concentration (FIC) index: ≤0.5 (synergism), >0.5–1 (additive), >1 to <2 (indifferent), and ≥2 (antagonism). FICD1 refers to the fractional inhibitory concentration of DA, luteolin, or PYR. FICD2 refers to the fractional inhibitory concentration of fluoroquinolone drug. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.