")
Back to Journals » OncoTargets and Therapy » Volume 14
Identification of HOXD10 as a Marker of Poor Prognosis in Glioblastoma Multiforme
Authors Li Y, Ma K , Xie Q, Zhang X, Zhang X , Chen K , Kong L, Qian R
Received 3 September 2021
Accepted for publication 11 October 2021
Published 27 October 2021 Volume 2021:14 Pages 5183—5195
DOI https://doi.org/10.2147/OTT.S336225
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Dr Alberto Bongiovanni
Yanxin Li,1 Ke Ma,2 Qi Xie,3 Xianwei Zhang,3 Xiulei Zhang,4 Kui Chen,1 Lingfei Kong,3 Rongjun Qian1
1Department of Neurosurgery, Henan Provincial People’s Hospital, People’s Hospital of Zhengzhou University, People’s Hospital of Henan University, Zhengzhou, Henan, 450003, People’s Republic of China; 2Department of Medical Oncology, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, Henan, 450052, People’s Republic of China; 3Department of Pathology, Henan Provincial People’s Hospital, People’s Hospital of Zhengzhou University, People’s Hospital of Henan University, Zhengzhou, Henan, 450003, People’s Republic of China; 4Department of Microbiome Laboratory, Henan Provincial People’s Hospital, People’s Hospital of Zhengzhou University, People’s Hospital of Henan University, Zhengzhou, Henan, 450003, People’s Republic of China
Correspondence: Rongjun Qian
Department of Neurosurgery, Henan Provincial People’s Hospital, People’s Hospital of Zhengzhou University, People’s Hospital of Henan University, No. 7, Wei Wu Road, Zhengzhou, Henan, 450003, People’s Republic of China
Tel +86 15037138200
Email [email protected]
Lingfei Kong
Department of Pathology, Henan Provincial People’s Hospital, People’s Hospital of Zhengzhou University, People’s Hospital of Henan University, No. 7, Wei Wu Road, Zhengzhou, Henan, 450003, People’s Republic of China
Tel +86 18538298216
Email [email protected]
Purpose: HOXD10 is a tumor modulator that can either be a tumor-suppressor or a tumor-promoting gene. However, the role of HOXD10 in glioblastoma multiforme (GBM) remains unclear.
Methods: Immunohistochemistry (IHC) was applied to detect protein expression of HOXD10 in GBM and normal brain tissue patients. Clinicopathological characteristics with GBM were recorded, and a Kaplan–Meier curve was plotted. Additionally, the mRNA expression of HOXD10 and its effect on prognosis were analyzed using the online tool GEPIA and the Cancer Genome Atlas (TCGA), Chinese Glioma Genome Atlas (CGGA), and the Gene Expression Omnibus (GEO) databases. Based on the mRNA expression of HOXD10, GBM patients from TCGA database were divided into low- and high-HOXD10 expression groups to analyze the Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways, and construct a lncRNA-miRNA-mRNA network and a protein–protein interaction (PPI) network.
Results: The mRNA expression of HOXD10 was up-regulated in GBM according to GEPIA, while the protein expression of HOXD10 in GBM was down-regulated according to IHC analysis of samples from patients collected from our hospital. Correlation analysis showed that HOXD10 expression was significantly related to IDH1 status. Univariate analysis revealed that low HOXD10 expression, complete surgical resection, postoperative radiotherapy, postoperative temozolomide chemotherapy and IDH1 mutation were all beneficial prognostic factors. Further multivariate analysis revealed that only complete surgical resection and postoperative radiotherapy were independent prognostic factors. GO and KEGG enrichment analyses indicated that HOXD10 expression is mainly involved in cytokine-cytokine receptor interactions. In the ceRNA network, 89 nodes, containing 45 mRNAs, 39 miRNAs and five lncRNAs associated with prognosis were involved. The PPI network revealed a tight interaction between HOXD10 and HOXD8, HOXD9, HOXD11, HOXD13 and HOXB3.
Conclusion: Based on our experimental data, although HOXD10 expression is low in GBM compared with normal brain tissue, GBM patients with high HOXD10 expression have a worse prognosis. HOXD10 may play different or even opposite roles in different stages of GBM occurrence and development. For patients with GBM, HOXD10 may be a valid predictor of prognosis.
Keywords: HOXD10, GBM, prognosis, ceRNA, protein-protein interaction
Introduction
Glioblastoma multiforme (GBM) is the most common and aggressive primary intracranial tumor in adults, with a high fatality rate. The median survival time of GBM is only 14–15 months, with a 10% probability of 5-year survival.1 Improving the survival time of GBM patients remains a difficult problem.
HOX genes encode proteins that function as critical master regulatory transcription factors during embryogenesis and also play key roles in the development of cancers.2 In humans, 39 HOX genes are organized into four clusters (HOX-A, HOX-B, HOX-C, and HOX-D) on different chromosomes. At present, the expression of HOX family genes in a variety of tumors has been reported, such as lung cancer,3–5 breast cancer,6–8 ovarian cancer,9–12 prostate cancer,13–16 colorectal carcinoma17–20 and glioma.21–23
Although deregulated HOX gene expression was originally associated with oncogenic activities, it is now apparent that HOX genes might be lost as well as gained in cancer, and they may function as tumor suppressors as well as tumor promoters.24 The aberrant expression of specific HOX genes tends to differ based on the tissue type and tumor site.25 HOXD10, a tumor suppressor gene, is expressed at a low level in most tumors compared with normal tissues, such as in cholangiocellular carcinoma,26 gastric carcinogenesis,27 prostate cancer28 and hepatocellular carcinoma.29 However, the expression of HOXD10 in glioma is limited and controversial, especially in GBM. Fattah et al analyzed the expression profile of HOX family genes in neoplastic and non-neoplastic astrocytes using both cell lines and human brain tissues. That study showed that HOXD10 was silent or expressed at very low levels in nonneoplastic astrocytes, while they were over-expressed in malignant astrocytic cell lines and GBM specimens.30 Additionally, Buccoliero et al used RT-PCR to examine the relative expression of HOX-D genes in a group of 14 pediatric low-grade gliomas, and the results reveled that HOXD10 expression was found in four (29%) out of 14 astrocytoma samples, whereas it was not found in any non-neoplastic brain tissues.31 In Chinese studies, HOXD10 is highly expressed in normal brain tissues and expressed at a low level in human glioma tissues, and it is negatively correlated with the malignant degree and prognosis of glioma.32,33
Considering the inconsistency of the abovementioned research results and the small sample sizes included in these studies, with the aim to further clarify the significance of HOXD10 in GBM, we analyzed the mRNA expression of HOXD10 and its effect on prognosis using data derived from online databases such as GEPIA and The Cancer Genome Atlas (TCGA), the Chinese Glioma Genome Atlas (CGGA), and the Gene Expression Omnibus (GEO) (GSE74187 and GSE4412). Importantly, we examined the protein expression of HOXD10 in 71 GBM patients collected from our hospital, and their overall survival time (OS) and progression-free survival time (PFS) were analyzed. Additionally, based on the mRNA expression of HOXD10, GBM patients from TCGA were divided into a low-HOXD10-expression group and a high-HOXD10-expression group to analyze gene ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways, and construct a long non-coding RNA (lncRNA)-microRNA(miRNA)-mRNA network as well as a protein-protein interaction (PPI) network.
Materials and Methods
Databases
GEPIA is a newly developed interactive web server for analyzing the RNA sequencing expression data of tumors and normal samples from TCGA and the Genotype Tissue Expression (GTEx) projects using a standard processing pipeline, which was used to compare the mRNA expression of HOXD10 in GBM and normal tissues.
The mRNA-seq data in count format of the TCGA-GBM cohort was obtained from the TCGA dataset (https://portal.gdc.cancer.gov/). mRNA microarray data from 301 glioma patients and mRNA sequencing data from 693 glioma patients were obtained from the CGGA data portal (http://www.cgga.org.cn/). Patients histologically classified with primary GBM were screened for subsequent analyses. mRNA microarray data from GSE74187 and GSE4412 were obtained from GEO (https://www.ncbi.nlm.nih.gov/geo/).
At the same time, survival information matching the above datasets was obtained, and patients with a survival time greater than 30 days were used for survival analysis.
Clinicopathological Data Collection and Analysis
Clinical data such as age, gender and pathological results were retrospectively collected from 71 patients who underwent surgical resection and were pathologically confirmed with GBM from January 2016 to December 2018 at the Henan Provincial People’s Hospital. The patients included 43 males and 28 females aged between 10 and 81 years. The tumors of the patients were staged according to the 2016 WHO classification system. Survival time was calculated from the date of diagnosis to the end of follow-up. The patients were followed up until death or until the end date of the study (31 January 2021). The median follow-up was 11.4 months. The study was approved by the Ethics Committees of the Henan Provincial People’s Hospital and the ethical approval numbers was 2020-107. All the patients selected for our study were fully informed about our experiment protocols and signed an informed consent to participate in this study. The study was conducted in accordance with the Declaration of Helsinki.
Immunohistochemistry
All specimen tissues were formalin fixed, paraffin embedded and sectioned into 4-µm serial sections. Additionally, 15 normal brain tissue samples were assessed as the control group. The immunohistochemistry was performed according to our previously published article.34 Briefly, after antigen retrieval (using a microwave at high power for 8 min, followed by middle-low power for 7 min in EDTA solution at pH 9.0), the sections were treated to quench endogenous peroxidase activity. Rabbit serum was used for blocking. The slides were then stained with HOXD10 antibody at 1:200 (ab85698, Abcam, Cambridge, UK) overnight at 4°C and secondary antibodies were incubated for 1 h at room temperature. After adding substrate and hematoxylin staining, the slides were covered and observed by microscope. The proportion of stained cells in each field was assessed as follows: 0% = 0; 1–24% = 1; 25–49% = 2; 50–74% = 3; and 75–100% = 4. The intensity of overall staining was graded as follows: no staining = 0; weak staining = 1; moderate staining = 2; and strong staining = 3. The overall immunostaining score for each specimen was calculated by multiplying the percentage score by the intensity score.35 A final score of 0–3 indicated negative expression and 4–12 indicated positive expression.
Differentially Expressed Gene Analysis and Functional Enrichment Analysis
With the aim to understand the role of HOXD10 in GBM, we performed the functional enrichment analysis. Differentially expressed genes (DEGs) were identified with the “DEseq2 package” in R software. Fold change > 1.0 and adjusted P < 0.05 were set as the cutoffs. A volcano plot of the DEGs was drawn using the “ggplot2” package. Then, we transformed the gene symbols into Entrez IDs via the “org.Hs.eg.db” package and performed pathway enrichment analysis for the DEGs based on the GO database and KEGG by using the “cluster Profiler”, “ggplot2” and “enrich plot” packages. The P-values were adjusted by the False Discovery Rate (FDR) method and an FDR < 0.25 was considered significantly enriched pathways.
Construction of a lncRNA-miRNA-mRNA (ceRNA) Network
The “survival” package was used to identify six lncRNAs that are related to the prognosis of GBM. Relevant lncRNAs-target data (miRNAs) were obtained from miRCode. Relevant miRNA-target data (mRNAs) were obtained from miRTarBase. Then, a lncRNA-miRNA-mRNA network was constructed according to the hypothesis of ceRNA.36 In total, we obtained 89 nodes (45 mRNAs, 39 miRNAs and 5 lncRNAs) and 227 edges to construct the lncRNA-miRNA-mRNA network. Nodes and edge files were exported and visualization was performed with Cytoscape 3.6.1.
PPI Network Analysis
The interaction of 138 HOXD10 correlated differentially expressed mRNAs (DEmRNAs) were analyzed using the online tool STRING (https://www.string-db.org/). A PPI network was constructed by setting medium confidence at 0.400 and excluding irrelevant genes. The network was exported and visualized using Cytoscape 3.6.1.
Statistical Analysis
Differences between groups were analyzed using the χ2 test or Student’s t-test depending on the data type. Kaplan-Meier plots and Log rank tests were used for survival analysis. Multivariate analyses were based on the Cox proportional hazards regression model. All statistical analyses were performed in RStudio (version 3.6.8), and we considered a P-value < 0.05 to be statistically significant.
Results
High mRNA Expression of HOXD10 in GBM Predicted Poor Prognosis
The mRNA expression of HOXD10 in GBM and normal tissues was analyzed using GEPIA. This analysis revealed that the HOXD10 expression was much higher in GBM compared to normal tissues (Figure 1A). Based on the database analysis of TCGA, CGGA, GSE74187 and GSE4412, high HOXD10 expression was associated with shorter survival time than those with low HOXD10 expression in GBM (Figure 1B–F).
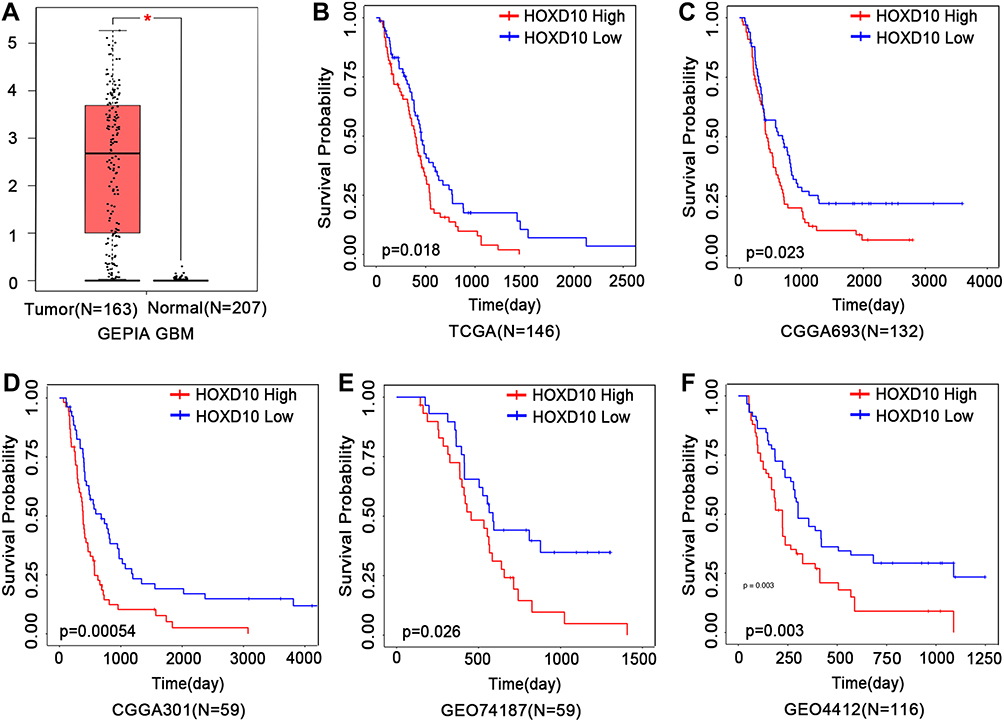 |
Figure 1 (A) The mRNA expression of HOXD10 is significantly upregulated in GBM compared with normal brain tissues. *P < 0.05. (B–F) High mRNA expression of HOXD10 in GBM predicted poor prognosis based on database analyses. (B), TCGA database; (C and D), CGGA database; (E and F), GEO database. Abbreviations: GBM, glioblastoma multiforme; TCGA, The Cancer Genome Atlas; CGGA, Chinese Glioma Genome Atlas; GEO, Gene Expression Omnibus. |
Immunohistochemical analysis of HOXD10 protein expression and the association with clinicopathological characteristics.
Then, we collected and followed up 71 GBM patients and 15 normal brain tissues from the Henan Provincial People’s Hospital. IHC was used to examine the protein expression of HOXD10 in GBM. Immunostaining of HOXD10 was observed in the nucleus of cancer cells (Figure 2A–D), and the positive staining rate in GBM was 26.80% (19/71), while the positive rate in normal brain tissues was 73.3% (11/15). The expression of HOXD10 was much higher in normal brain tissue than that in GBM samples (χ2 = 11.825, P = 0.001). The association between HOXD10 expression and the clinicopathological characteristics of GBM patients are listed in Table 1. The results revealed that the expression of HOXD10 was significantly associated with the status of IDH1.
 |
Table 1 Associations Between HOXD10 Protein Expression and GBM Patient Clinicopathological Characteristics |
 |
Figure 2 Representative immunohistochemical staining of HOXD10 in GBM and normal brain tissues. (A), HOXD10 was negative in normal brain tissue (X400). (B), HOXD10 was positive in normal brain tissue (X400). (C), HOXD10 was negative in GBM (X400). (D), HOXD10 was positive in GBM (X400). Abbreviation: GBM, glioblastoma multiforme. |
Univariate and Multivariate Survival Analyses
Survival analysis showed that complete surgical resection, postoperative radiotherapy, postoperative temozolomide (TMZ) chemotherapy, IDH1 mutation, and low HOXD10 expression were beneficial prognostic factors (Figure 3A–E). Further multivariate Cox regression analysis revealed that complete surgical resection and postoperative radiotherapy were independent prognostic factors (Table 2). Furthermore, GBM patients who received complete surgical resection, postoperative radiation therapy and had only one lobe involved at the initial treatment had a longer median PFS (mPFS) (Figure 3F–H), and these three factors were independent predictive factors (Table 3).
 |
Table 2 Multivariate Analysis of Prognosis for GBM (N=71) |
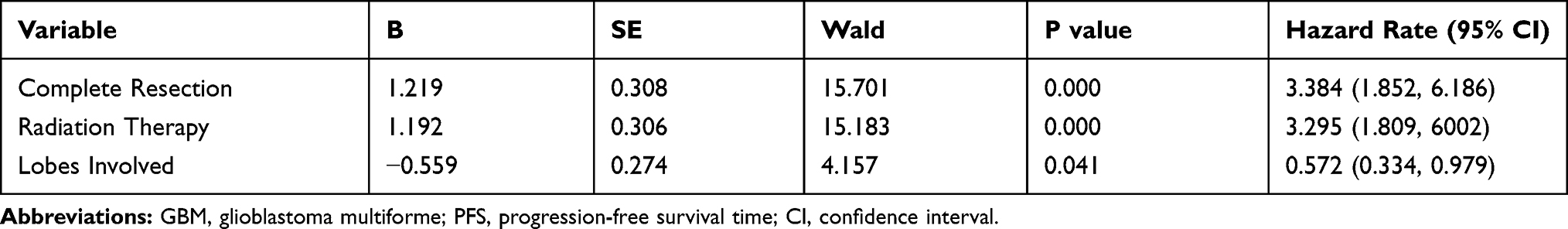 |
Table 3 Multivariate Analysis of PFS for GBM (N=71) |
 |
Figure 3 Univariate and multivariate survival analyses. (A), Complete surgical resection can improve the survival time of GBM patients. (B), Postoperative TMZ chemotherapy can improve the survival time of GBM patients. (C), Postoperative radiation therapy can improve the survival time of GBM patients. (D), GBM patients with IDH1 mutation had a longer survival time. (E), GBM patients with high HOXD10 protein expression had a shorter survival time. (F), Complete surgical resection can improve the PFS of GBM patients. (G), Tumors that invade multiple lobes were more likely to recur after surgery. (H), Postoperative radiation therapy can prolong the PFS of GBM patients. Abbreviations: GBM, glioblastoma multiforme; TMZ, Temozolomide; mOS, median overall survival time; mPFS, median progression-free survival time. |
DEG Profiles and Functional Enrichment Analysis
To understand the biological functions related to the DEGs, we divided GBM patients from TCGA database into low and high expression groups based on the median value of HOXD10 expression. A total of 138 DEGs were identified (Table S1; Figure 4A). GO contains biological process (BP), cellular components (CC) and molecular function (MF). There was no item enriched in CC process and the enriched items in BP and MF were shown in Figure 4B. KEGG enrichment analyses showed that the DEGs based on HOXD10 expression were mainly involved in cytokine-cytokine receptor interactions, viral protein interactions with cytokines and cytokine receptors and neuroactive ligand-receptor interaction signaling pathways (Figure 4C).
 |
Figure 4 Differential analysis of gene expression profiles in high- and low-HOXD10 expression groups and pathway enrichment analysis. (A), Volcano plot showing the differentially expressed genes. (B), Bubble chart of the results of GO enrichment analysis. (C), Bar plot of the results of KEGG enrichment analysis. Abbreviations: FDR, false discovery rate; BP, biological process; MF, molecular function. |
ceRNA Network
The “survival” package was used to identify six survival-related lncRNAs (Figure 5A–F). LINC01539 was removed because there was no suitable miRNA match. Based on the expression level of each lncRNA, the ceRNA network was divided into a lncRNA high expression group and a lncRNA low expression group. In the up-regulated ceRNA network, there were four lncRNAs, 37 miRNAs and 42 mRNAs (Figure 6A). In the down-regulated ceRNA network, there was one lncRNA, three miRNAs and three mRNAs (Figure 6B).
 |
Figure 5 Survival analysis of differentially expressed lncRNAs. (A–D and F), High expression of HOTAIR, HOXC-AS3, HOXC12-AS, LINC01305 and LINC01539 were poor prognostic factors. (E), High expression of MIR663AHG was good prognostic factor. |
 |
Figure 6 Construction of a ceRNA network. (A), The ceRNA network was constructed based on the up-regulation of lncRNAs screened by HOXD10 expression level and related to the survival of GBM patients. (B), The ceRNA network was constructed based on down-regulated lncRNAs screened by HOXD10 expression level and related to the survival of GBM patients. Note: Yellow, mRNA; Green, miRNA; Red, LncRNA. |
PPI Network
To construct the PPI network, 138 HOXD10-correlated DEmRNAs were analyzed, and 71 genes were included in the network after the elimination of disconnected nodes. The network is shown in Figure 7. HOXD10 was closely related to other HOX family members, including HOXD8, HOXD9, HOXD11, HOXD13 and HOXB3.
 |
Figure 7 Protein–protein interaction (PPI) network analysis. After eliminating disconnected nodes, 71 genes were involved in the PPI. The network was visualized with Cytoscape 3.6.1. |
Discussion
Previous studies on HOXD10 in GBM are limited, and most have focused on gliomas of all grades. In the present study, to our knowledge, this is the first examination of HOXD10 expression specifically in GBM. We found that the protein expression of HOXD10 in GBM was much lower than that in normal brain tissues, contrary to the results of the online tool GEPIA. Sample size may go some way to explaining the difference. But, more importantly, we found it interesting that in the small number of studies by Chinese authors, the expression of HOXD10 in normal brain tissues was much higher than that in human glioma tissues.32,33 While studies from the non-Chinese scholars, HOXD10 expression in glioma was slightly higher than that in non-neoplastic brain.30,31 Whether racial difference is one reason for the inconsistency of our data and the online tool GEPIA remains to be further verified. Lastly, whether HOXD10 differs in the expression levels of RNA and protein remains to be further studied.
Both our own data and database-derived data suggest that GBM patients with high HOXD10 expression have significantly worse OS. However, a previous study of 40 glioma samples showed that high expression of HOXD10 was a factor that predicted better prognosis.33 In addition to small sample sizes, the previous studies included not only GBM, but also other grades of glioma.
In fact, HOXD10 is recognized as a tumor suppressor and is known to be down-regulated in a variety of cancer types,26–29,32 such as colon carcinomas,37 early-stage head and neck squamous cell carcinoma (HNSCC)38 and oral squamous cell carcinoma,39 in which HOXD10 is up-regulated. In colon carcinomas, up-regulation of HOXD10 contributes to the crypt colonic stem cell overpopulation that drives colorectal cancer (CRC) development37 and in HNSCC and cervical cancer, HOXD10 over-expression resulted in increased proliferation ability.38,40 Based on our data analysis, although HOXD10 expression was low in GBM compared with normal brain tissue, patients with high HOXD10 expression have a worse prognosis. The underlying mechanism could be like that of TGF-β, a well-known bifunctional regulator that can either inhibit or stimulate cell proliferation.41 Typically, this cellular effect accompanies an induction of mesodermal cell fate and inhibition of neural cell differentiation. However, during pathological conditions, these defined effects of TGF-β can be reversed; for example, the growth inhibitory effect is replaced with its tumor promoting ability.42 Therefore, based on unique features with respect to its expression and function, HOXD10 is best described as a “tumor modulator” rather than an oncogene or tumor suppressor gene. HOXD10 may play different or even opposite roles in different stages of GBM occurrence and development. Further larger samples and deeper exploration need to be performed.
IDH1 mutation as a marker of prognosis has been widely recognized and it is regarded as a therapeutic target,43 which was further confirmed in this study. Correlation analysis showed that HOXD10 expression was associated the status of IDH1, although the specific regulatory mechanism remains unclear.
GO enrichment analyses revealed the DEGs based on the expression of HOXD10 from TCGA database mainly acted as the DNA-binding transcription activators and involved in the particular tissue or organ development. In accordance with the previous studies that HOX proteins can act as transcription factors to enhance or repress gene expression during normal embryonic development.2 HOX proteins are integral to normal temporospatial limb and organ development along the anterior-posterior axis.44 KEGG analysis showed that the signaling pathways mainly involved in the cytokine-cytokine receptor interaction, which indicated that HOXD10 may regulate the inflammatory response and immune function in GBM.
In the ceRNA network, we found that HOTAIR and LINC01305 were enriched in a number of nodes. HOTAIR is one of the most extensively studied lncRNAs found to be dysregulated in human cancer.45 Tan et al used qRT-PCR to measure the HOTAIR expression in serum from 43 GBM and 40 control samples, and the results revealed that HOTAIR expression was significantly correlated with high grade brain tumors and that serum HOTAIR can be used as a novel prognostic and diagnostic biomarker for GBM.46 Previous research also indicated that HOTAIR promoted malignant progression and poor prognosis in glioma patients and exhibited pro-oncogenic activity.47 LINC01305 has been widely explored in cervical cancer and is involved in the epithelial-mesenchymal transition by regulating the PI3K/AKT signaling pathway.48,49 In the present study, the database analysis showed that LINC01305 was over-expressed and predicted poor prognosis in GBM. However, its clinical significance and prognostic value needed to be further verified in our tissue samples. Through gene enrichment and PPI network analysis of HOXD10-related genes, we found that HOXD10 is mainly involved in the signal transduction of cytokine interactions, and there was mutual regulation between HOXD10 and multiple cytokines, such as CCL20 and CXCL6. HOXD10 has been found to be involved in tumor proliferation, invasion or angiogenesis through multiple signaling pathways, such as the RHOC/AKT/MAPK and ERK signaling pathways.26,29,38,40,50,51 Whether HOXD10 interacts with cytokines to regulate the GBM tumor microenvironment is a future research direction.
Conclusion
Based on our experimental data, although HOXD10 expression is low in GBM compared with normal brain tissues, GBM patients with high HOXD10 expression have a worse prognosis. HOXD10 may play different or even opposite roles in different stages of GBM occurrence and development. For patients with GBM, HOXD10 may be a valid predictor of prognosis.
Acknowledgments
The present study was supported by grants from the National Natural Science Foundation of China (No. U1904160). We thank International Science Editing (http://www.internationalscienceediting.com) for editing this manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Gallego O. Nonsurgical treatment of recurrent glioblastoma. Curr Oncol. 2015;22(4):e273–e281. doi:10.3747/co.22.2436
2. Shah N, Sukumar S. The Hox genes and their roles in oncogenesis. Nat Rev Cancer. 2010;10(5):361–371. doi:10.1038/nrc2826
3. Abe M, Hamada J, Takahashi O, et al. Disordered expression of HOX genes in human non-small cell lung cancer. Oncol Rep. 2006;15(4):797–802.
4. Plowright L, Harrington KJ, Pandha HS, Morgan R. HOX transcription factors are potential therapeutic targets in non-small-cell lung cancer (targeting HOX genes in lung cancer). Br J Cancer. 2009;100(3):470–475. doi:10.1038/sj.bjc.6604857
5. Bodey B, Bodey B, Groger AM, Siegel SE, Kaiser HE. Immunocytochemical detection of homeobox B3, B4, and C6 gene product expression in lung carcinomas. Anticancer Res. 2000;20(4):2711–2716.
6. Hur H, Lee JY, Yun HJ, Park BW, Kim MH. Analysis of HOX gene expression patterns in human breast cancer. Mol Biotechnol. 2014;56(1):64–71. doi:10.1007/s12033-013-9682-4
7. Makiyama K, Hamada J, Takada M, et al. Aberrant expression of HOX genes in human invasive breast carcinoma. Oncol Rep. 2005;13(4):673–679.
8. Shaoqiang C, Yue Z, Yang L, et al. Expression of HOXD3 correlates with shorter survival in patients with invasive breast cancer. Clin Exp Metastasis. 2013;30(2):155–163. doi:10.1007/s10585-012-9524-y
9. Cheng W, Liu J, Yoshida H, Rosen D, Naora H. Lineage infidelity of epithelial ovarian cancers is controlled by HOX genes that specify regional identity in the reproductive tract. Nat Med. 2005;11(5):531–537. doi:10.1038/nm1230
10. Naora H, Montz FJ, Chai CY, Roden RB. Aberrant expression of homeobox gene HOXA7 is associated with mullerian-like differentiation of epithelial ovarian tumors and the generation of a specific autologous antibody response. Proc Natl Acad Sci U S A. 2001;98(26):15209–15214. doi:10.1073/pnas.011503998
11. Yamashita T, Tazawa S, Yawei Z, et al. Suppression of invasive characteristics by antisense introduction of overexpressed HOX genes in ovarian cancer cells. Int J Oncol. 2006;28(4):931–938.
12. Ota T, Klausen C, Salamanca MC, Woo HL, Leung PC, Auersperg N. Expression and function of HOXA genes in normal and neoplastic ovarian epithelial cells. Differentiation. 2009;77(2):162–171. doi:10.1016/j.diff.2008.09.018
13. Miller GJ, Miller HL, van Bokhoven A, et al. Aberrant HOXC expression accompanies the malignant phenotype in human prostate. Cancer Res. 2003;63(18):5879–5888.
14. Waltregny D, Alami Y, Clausse N, de Leval J, Castronovo V. Overexpression of the homeobox gene HOXC8 in human prostate cancer correlates with loss of tumor differentiation. Prostate. 2002;50(3):162–169. doi:10.1002/pros.10045
15. Chen J, Zhu S, Jiang N, Shang Z, Quan C, Niu Y. HoxB3 promotes prostate cancer cell progression by transactivating CDCA3. Cancer Lett. 2013;330(2):217–224. doi:10.1016/j.canlet.2012.11.051
16. Javed S, Langley SE. Importance of HOX genes in normal prostate gland formation, prostate cancer development and its early detection. BJU Int. 2014;113(4):535–540. doi:10.1111/bju.12269
17. Kanai M, Hamada J, Takada M, et al. Aberrant expressions of HOX genes in colorectal and hepatocellular carcinomas. Oncol Rep. 2010;23(3):843–851.
18. Vider BZ, Zimber A, Hirsch D, et al. Human colorectal carcinogenesis is associated with deregulation of homeobox gene expression. Biochem Biophys Res Commun. 1997;232(3):742–748. doi:10.1006/bbrc.1997.6364
19. Liao WT, Jiang D, Yuan J, et al. HOXB7 as a prognostic factor and mediator of colorectal cancer progression. Clin Cancer Res. 2011;17(11):3569–3578. doi:10.1158/1078-0432.CCR-10-2533
20. Jung C, Kim RS, Zhang H, et al. HOXB13 is downregulated in colorectal cancer to confer TCF4-mediated transactivation. Br J Cancer. 2005;92(12):2233–2239. doi:10.1038/sj.bjc.6602631
21. Costa BM, Smith JS, Chen Y, et al. Reversing HOXA9 oncogene activation by PI3K inhibition: epigenetic mechanism and prognostic significance in human glioblastoma. Cancer Res. 2010;70(2):453–462. doi:10.1158/0008-5472.CAN-09-2189
22. Murat A, Migliavacca E, Gorlia T, et al. Stem cell-related “self-renewal” signature and high epidermal growth factor receptor expression associated with resistance to concomitant chemoradiotherapy in glioblastoma. J Clin Oncol. 2008;26(18):3015–3024. doi:10.1200/JCO.2007.15.7164
23. Tabuse M, Ohta S, Ohashi Y, et al. Functional analysis of HOXD9 in human gliomas and glioma cancer stem cells. Mol Cancer. 2011;10:60. doi:10.1186/1476-4598-10-60
24. Abate-Shen C. Deregulated homeobox gene expression in cancer: cause or consequence? Nat Rev Cancer. 2002;2(10):777–785. doi:10.1038/nrc907
25. Joo MK, Park JJ, Chun HJ. Impact of homeobox genes in gastrointestinal cancer. World J Gastroenterol. 2016;22(37):8247–8256. doi:10.3748/wjg.v22.i37.8247
26. Yang H, Zhou J, Mi J, et al. HOXD10 acts as a tumor-suppressive factor via inhibition of the RHOC/AKT/MAPK pathway in human cholangiocellular carcinoma. Oncol Rep. 2015;34(4):1681–1691. doi:10.3892/or.2015.4194
27. Wang L, Chen S, Xue M, et al. Homeobox D10 gene, a candidate tumor suppressor, is downregulated through promoter hypermethylation and associated with gastric carcinogenesis. Mol Med. 2012;18:389–400. doi:10.2119/molmed.2011.00172
28. Mo RJ, Lu JM, Wan YP, et al. Decreased HoxD10 expression promotes a proliferative and aggressive phenotype in prostate cancer. Curr Mol Med. 2017;17(1):70–78. doi:10.2174/1566524017666170220104920
29. Guo Y, Peng Y, Gao D, et al. Silencing HOXD10 by promoter region hypermethylation activates ERK signaling in hepatocellular carcinoma. Clin Epigenetics. 2017;9:116. doi:10.1186/s13148-017-0412-9
30. Abdel-Fattah R, Xiao A, Bomgardner D, Pease CS, Lopes MB, Hussaini IM. Differential expression of HOX genes in neoplastic and non-neoplastic human astrocytes. J Pathol. 2006;209(1):15–24. doi:10.1002/path.1939
31. Buccoliero AM, Castiglione F, Rossi Degl’Innocenti D, et al. Hox-D genes expression in pediatric low-grade gliomas: real-time-PCR study. Cell Mol Neurobiol. 2009;29(1):1–6. doi:10.1007/s10571-008-9282-1
32. Li M, Wang J, Sun G, et al. Expression and clinical significance of HOXD10 gene in gliomas. Jiangsu Med J. 2011;37(10):1154–1155.
33. Li XB, Xia Y, Nie L, Jin H. Expression and clinical significance of HOXD10 and SENP6 in brain glioma. J Chengdu Med Coll. 2020;15(4):495–498.
34. Fan Y, Ma K, Wang C, et al. Prognostic value of PD-L1 and PD-1 expression in pulmonary neuroendocrine tumors. Onco Targets Ther. 2016;9:6075–6082. doi:10.2147/OTT.S115054
35. Detre S, Saclani Jotti G, Dowsett M. A “quickscore” method for immunohistochemical semiquantitation: validation for oestrogen receptor in breast carcinomas. J Clin Pathol. 1995;48(9):876–878. doi:10.1136/jcp.48.9.876
36. Zhou M, Hu L, Zhang Z, Wu N, Sun J, Su J. Recurrence-associated long non-coding RNA signature for determining the risk of recurrence in patients with colon cancer. Mol Ther Nucleic Acids. 2018;12:518–529. doi:10.1016/j.omtn.2018.06.007
37. Bhatlekar S, Addya S, Salunek M, et al. Identification of a developmental gene expression signature, including HOX genes, for the normal human colonic crypt stem cell niche: overexpression of the signature parallels stem cell overpopulation during colon tumorigenesis. Stem Cells Dev. 2014;23(2):167–179. doi:10.1089/scd.2013.0039
38. Hakami F, Darda L, Stafford P, Woll P, Lambert DW, Hunter KD. The roles of HOXD10 in the development and progression of head and neck squamous cell carcinoma (HNSCC). Br J Cancer. 2014;111(4):807–816. doi:10.1038/bjc.2014.372
39. Rodini CO, Xavier FC, Paiva KB, et al. Homeobox gene expression profile indicates HOXA5 as a candidate prognostic marker in oral squamous cell carcinoma. Int J Oncol. 2012;40(4):1180–1188. doi:10.3892/ijo.2011.1321
40. Sharma S, Mandal P, Sadhukhan T, et al. Bridging links between long noncoding RNA HOTAIR and HPV oncoprotein E7 in cervical cancer pathogenesis. Sci Rep. 2015;5:11724. doi:10.1038/srep11724
41. Morikawa M, Derynck R, Miyazono K. TGF-beta and the TGF-beta family: context-dependent roles in cell and tissue physiology. Cold Spring Harb Perspect Biol. 2016;8(5):a021873. doi:10.1101/cshperspect.a021873
42. Golestaneh N, Mishra B. TGF-beta, neuronal stem cells and glioblastoma. Oncogene. 2005;24(37):5722–5730. doi:10.1038/sj.onc.1208925
43. Turkalp Z, Karamchandani J, Das S. IDH mutation in glioma: new insights and promises for the future. JAMA Neurol. 2014;71(10):1319–1325. doi:10.1001/jamaneurol.2014.1205
44. Gehring WJ, Hiromi Y. Homeotic genes and the homeobox. Annu Rev Genet. 1986;20:147–173. doi:10.1146/annurev.ge.20.120186.001051
45. Yao Y, Li J, Wang L. Large intervening non-coding RNA HOTAIR is an indicator of poor prognosis and a therapeutic target in human cancers. Int J Mol Sci. 2014;15(10):18985–18999. doi:10.3390/ijms151018985
46. Tan SK, Pastori C, Penas C, et al. Serum long noncoding RNA HOTAIR as a novel diagnostic and prognostic biomarker in glioblastoma multiforme. Mol Cancer. 2018;17(1):74. doi:10.1186/s12943-018-0822-0
47. Zhang JX, Han L, Bao ZS, et al. HOTAIR, a cell cycle-associated long noncoding RNA and a strong predictor of survival, is preferentially expressed in classical and mesenchymal glioma. Neuro Oncol. 2013;15(12):1595–1603. doi:10.1093/neuonc/not131
48. Yan SP, Chu DX, Qiu HF, et al. LncRNA LINC01305 silencing inhibits cell epithelial-mesenchymal transition in cervical cancer by inhibiting TNXB-mediated PI3K/Akt signalling pathway. J Cell Mol Med. 2019;23(4):2656–2666. doi:10.1111/jcmm.14161
49. Xu Y, Zhu H, Ma H, Yuan L, Hu Q, Yang L. LINC01305 inhibits malignant progression of cervical cancer via miR-129-5p/Sox4 axis. Am J Transl Res. 2020;12(11):7581–7592.
50. Myers C, Charboneau A, Cheung I, Hanks D, Boudreau N. Sustained expression of homeobox D10 inhibits angiogenesis. Am J Pathol. 2002;161(6):2099–2109. doi:10.1016/S0002-9440(10)64488-4
51. Yuan YH, Wang HY, Lai Y, et al. Epigenetic inactivation of HOXD10 is associated with human colon cancer via inhibiting the RHOC/AKT/MAPK signaling pathway. Cell Commun Signal. 2019;17(1):9. doi:10.1186/s12964-018-0316-0
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.