Back to Journals » Infection and Drug Resistance » Volume 13
Higher Prevalence of Multi-Antimicrobial Resistant Bacteroides spp. Strains Isolated at a Tertiary Teaching Hospital in China
Authors Wang Y, Han Y, Shen H, Lv Y, Zheng W, Wang J
Received 16 January 2020
Accepted for publication 16 May 2020
Published 27 May 2020 Volume 2020:13 Pages 1537—1546
DOI https://doi.org/10.2147/IDR.S246318
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Eric Nulens
Yanyan Wang, Yanqiu Han, Huimin Shen, Yingying Lv, Wenqi Zheng, Junrui Wang
Department of Clinical Laboratory, Affiliated Hospital of Inner Mongolian Medical University, Hohhot 010050, People’s Republic of China
Correspondence: Junrui Wang
Department of Clinical Laboratory, Affiliated Hospital of Inner Mongolian Medical University, Hohhot 010050, People’s Republic of China
Tel +86 13347104892
Email [email protected]
Purpose: The study investigates the molecular epidemiology of multi-drug resistant (MDR) Bacteroides spp. isolates and the clinical characteristics of the patients.
Materials and Methods: Bacteroides spp. clinical strains were identified through MALDI-TOF MS and VITEK-2 anaerobes and corynebacterium (ANC) cards. A broth microdilution method was employed to detect the antimicrobial sensitivities of Bacteroides spp. isolates. PCR was used to detect the resistance genes, including cfxA, cepA, cfiA, ermF, nim, as well as the upstream insertion sequence (IS) element of the cfiA gene. The effects of broad-spectrum efflux pump inhibitors (EPIs) on the minimal inhibitory concentration (MICs) of cefoxitin, moxifloxacin, and imipenem for MDR Bacteroides spp. were investigated.
Results: The total resistance rates of 115 Bacteroides spp. isolates to cefoxitin, moxifloxacin, clindamycin, metronidazole, imipenem and meropenem were 4.3%, 16.5%, 80.0%, 5.2%, 13.9% and 13.9%, respectively. The positive rates of carbapenem resistance gene cfiA were 38.9% and 8.6% for B. fragilis and non-B. fragilis isolates, respectively. The isolation rate of MDR isolates reached up to 18.26% (21/115), and the isolation rate among the gastrointestinal cancer patients was significantly higher when compared to the non-gastrointestinal cancer patients (52.38%/26.08%, P = 0.006). Furthermore, MDR isolates were more likely to be isolated from the patients exposed to cephalosporins 3 months before Bacteroides spp. isolation (76.19%/31.52%, P = 0.000).
Conclusion: The overall resistance rates of Bacteroides spp. isolates against multiple antimicrobials were at a high level, especially for B. fragilis. The CfiA gene carrying rate among B. fragilis isolates was as high as 38.9%, and its mediated carbapenem resistance was the major resistance mechanism for B. fragilis. The findings of this study imply that the real resistance tendency of Bacteroides spp. may be underestimated and need to be given more attention.
Keywords: anaerobe, Bacteroides fragilis, carbapenem resistance, clinical characteristics
Introduction
Bacteroides spp. is a common anaerobe that occupies the intestines of humans. It is also the most common anaerobe recovered from various infections, such as intra-abdominal infection, foot ulcer and bloodstream infection.1–3 In recent years, MDR Bacteroides fragilis (B. fragilis) has emerged in several areas around the world.4–6 However, its resistance capability has been significantly varied among different geographical locations.7
In treating an infection of Bacteroides spp., metronidazole, clindamycin and β-lactam antimicrobials are routinely prescribed. Based on relevant literature, the resistance of Bacteroides spp. isolates to these kinds of antimicrobials were related to some specific resistance genes. The nim gene was found to be closely correlated with metronidazole,8,9 while the presence of the cfxA gene strongly correlated with cefoxitin resistance.10 Macrolide-Lincosamide-Streptogramin B resistant determinants, such as erm genes, are widely distributed among Bacteroides spp. isolates and the ermF gene is responsible for B. fragilis resistance to clindamycin.10,11 The existence of imipenem resistant Bacteroides spp. isolates were first reported over three decades ago.12 As reported in recent studies, the primary mechanism of carbapenem resistance of B. fragilis was the production of metallo-β-lactamase that is encoded by the cfiA gene;13 the specific upstream insertion sequence (IS) element is required for its expression and has been confirmed by some subsequent investigations.14 However, owing to a lack of popularization of anaerobic cell culture in Chinese hospitals, not as much is known regarding the actual resistance tendency and resistance mechanism of Bacteroides spp. clinical isolates.
In this study, we aim to detect the resistance features and possible resistance mechanism(s) of Bacteroides spp. clinical isolates collected at a 3000-bed tertiary teaching hospital in China.
Materials and Methods
Anaerobes Culture, Isolation and Identification
Between March 2017 and February 2019, all of the anaerobic samples were cultured under anaerobic conditions. The anaerobe isolates were collected from clinical samples of patients from the Affiliated hospital of Inner Mongolian Medical University and identified with both Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry (microTyper MS, Tianrui, China) and VITEK-2 Compact automated microbiology system (BioMérieux, France) with anaerobes and corynebacterium (ANC) cards (BioMérieux, France). The duplicated isolates found in individuals were deleted.
Clinical Characteristics of the Patients Enrolled in This Study
A total of 113 patients were enrolled in this study. The patients were mainly middle-aged and older adults, with a median age of 49 years (range 2 to 82), and male patients accounted for 60.18% (68/113), females accounted for 39.82% (45/113). Intra-abdominal and pelvic infections accounted for the most substantial proportion (92.04%, 104/113), followed by bacteremia (4.42%, 5/113), skin and soft tissue (2.65%, 3/113) and pulmonary infection (0.88%, 1/113).
Antimicrobial Susceptibility Test
The minimal inhibitory concentration (MIC) values of 11 antibiotics against all of the isolates was determined using a broth microdilution method under anaerobic conditions. This was done according to the recommendation of the Clinical and Laboratory Standard Institute (CLSI)15 and the European Committee on Antimicrobial Susceptibility Testing (EUCAST).16 The antibiotics were as follows: ampicillin/sulbactam, amoxicillin/clavulanic acid, imipenem, cefoxitin, meropenem, piperacillin, moxifloxacin, piperacillin/tazobactam, metronidazole, clindamycin, and tigecycline. The following concentrations of the antibiotics were tested: ampicillin/Sulbactam (0.25/0.125–32/16µg/mL), amoxicillin/clavulanic acid (0.125/0.06–16/8µg/mL), imipenem (0.25–32µg/mL), cefoxitin (0.5–64µg/mL), meropenem (0.25–32µg/mL), clindamycin (0.125–16µg/mL), piperacillin (0.5–128µg/mL), metronidazole (0.25–32µg/mL), moxifloxacin (0.06–8µg/mL), piperacillin/tazobactam (0.5/4–64/4µg/mL), and tigecycline (0.25–32µg/mL).
After 48 hours incubation at 35°C under anaerobic condition, the antimicrobial MIC values were evaluated based on the breakpoints of CLSI and EUCAST,16 as seen in Table 1. B. fragilis ATCC 25285 was used as the control strain. Production of metallo-β-lactamases (MBLs) was detected by an EDTA inhibition assay for any isolates that had reduced susceptibility or resistance to carbapenem antibiotics.
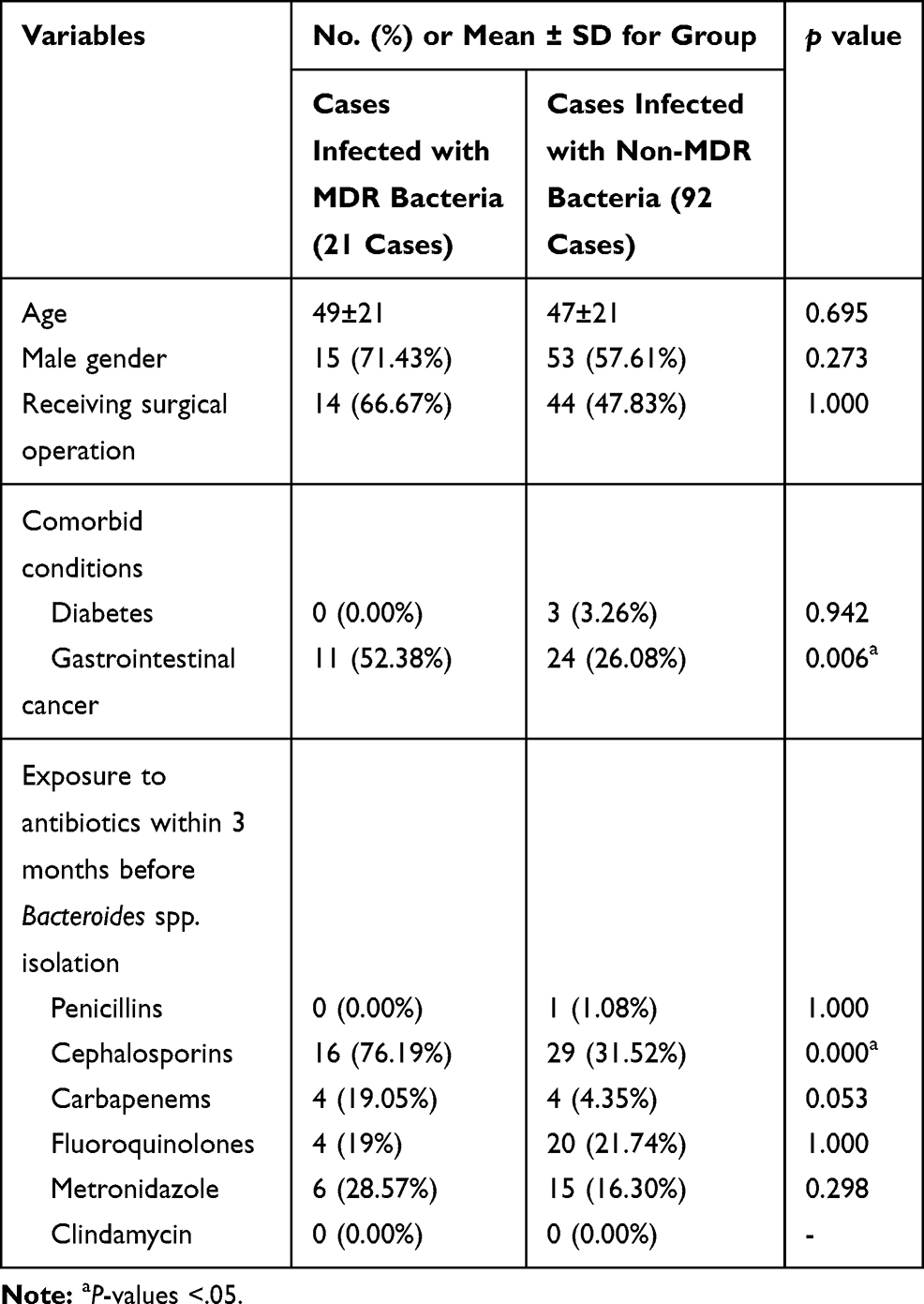 |
Table 1 Demographic and Clinical Characteristics of 113 Inpatients Included in the Study |
Detection of Resistance Genes
DNA of the isolates was extracted using a bacterial DNA extraction kit (Qiagen Ltd, Germany). The most common antibiotic resistance genes (cepA, cfxA, cfiA, ermF, nim)4,8 and the upstream insertion sequence (IS)14 of the cfiA gene was amplified. Then, the PCR products were analyzed with 1.2% agarose gel electrophoresis, and the DNA amplicon of the PCR was purified using the QIAquick Gel Extraction Kit (Qiagen Ltd, Germany). The IS elements were further verified by PCR using well-known primers17 and then sequenced.
Efflux Pump Inhibition Assay
Broad-spectrum efflux pump inhibitors (EPIs), carbonyl cyanide m-chlorophenylhydrazone (CCCP) and verapamil (Sigma, St Louis, MO, US) were selected to test the effects of EPIs on the MICs of cefoxitin, moxifloxacin, and imipenem for MDR Bacteroides spp. In this study, multi-drug resistant (MDR) isolates were defined as the ones resistant to at least three types of antimicrobials, including penicillins (ampicillin/sulbactam, amoxicillin/clavulanic acid, piperacillin, and piperacillin/tazobactam), cephalosporins (cefoxitin), carbapenems (imipenem and meropenem), clindamycin, nitroimidazoles (metronidazole) and fluoroquinolones. CCCP or verapamil was added to brucella blood agar plates to obtain final concentrations of 25μg/mL and 100μg/mL, respectively.18 Antimicrobials tested in this study were added to the plates in gradient mode beforehand. Then, the plates were inoculated with the test isolates at the required concentration and incubated anaerobically at 35°C for 48 h, and the MIC of each plate was measured.
Statistical Analysis
Two-side Student’s t-tests, Chi-square tests and Fisher’s exact tests were used to perform univariate analyses. SPSS software version 23.0 (SPSS, Chicago, IL, USA) was used for the analysis. P-values of <0.05 were considered statistically significant.
Results
Clinical Characteristics of the Patients Infected with MDR Isolates
A total of 21 MDR Bacteroidesspp. isolates were collected in this study. The isolation rate among the gastrointestinal cancer patients was significantly higher compared to the non-gastrointestinal cancer patients (52.38%/26.08%, P = 0.006). Furthermore, MDR isolates were more likely to be isolated from the patients who were exposed to cephalosporins within 3 months before Bacteroides spp. isolation (76.19%/31.52%, P = 0.000), as shown in Table 1.
Antimicrobial Susceptibility Test results
Among 115 Bacteroides spp. isolates, B. fragilis accounted for 69.6% (80/115), followed by Bacteroides thetaiotaomicron (17.4%, 20/115), Bacteroides ovatus (3.5%, 4/115), Bacteroides uniformis (2.6%, 3/115), Bacteroides vulgatus (2.6%, 3/115), Bacteroides novobacterioides (2.6%, 3/115), and Parabacteroides distasonis (1.7%, 2/115). The total resistance rates of 115 Bacteroides spp. isolates to ampicillin/sulbactam, amoxicillin/clavulanic, cefoxitin, piperacillin, piperacillin/tazobactam, moxifloxacin, clindamycin, metronidazole, imipenem and meropenem were 22.6%, 19.6%,4.3%, 27.8%, 8.7%, 16.5%, 80.0%, 5.2%, 13.9% and 13.9%, respectively.
When comparing against non-B. fragilis isolates, B. fragilis isolates showed a higher level of resistance to the majority of the antimicrobials used in this study, as shown in Table 2. The resistance rates of 80 B. fragilis isolates to both imipenem and meropenem were 18.8% (15/80). A total of 26 Bacteroides spp. isolates had reduced susceptibility or were resistant to imipenem or meropenem, 16 isolates were found to be MBL producers, including 15 isolates of B. fragilis and one isolate of B. thetaiotaomicron, respectively. Moreover, MIC90 values of B. fragilis and non-B. fragilis isolates against tigecycline were 4 µg/mL and 2 µg/mL, respectively.
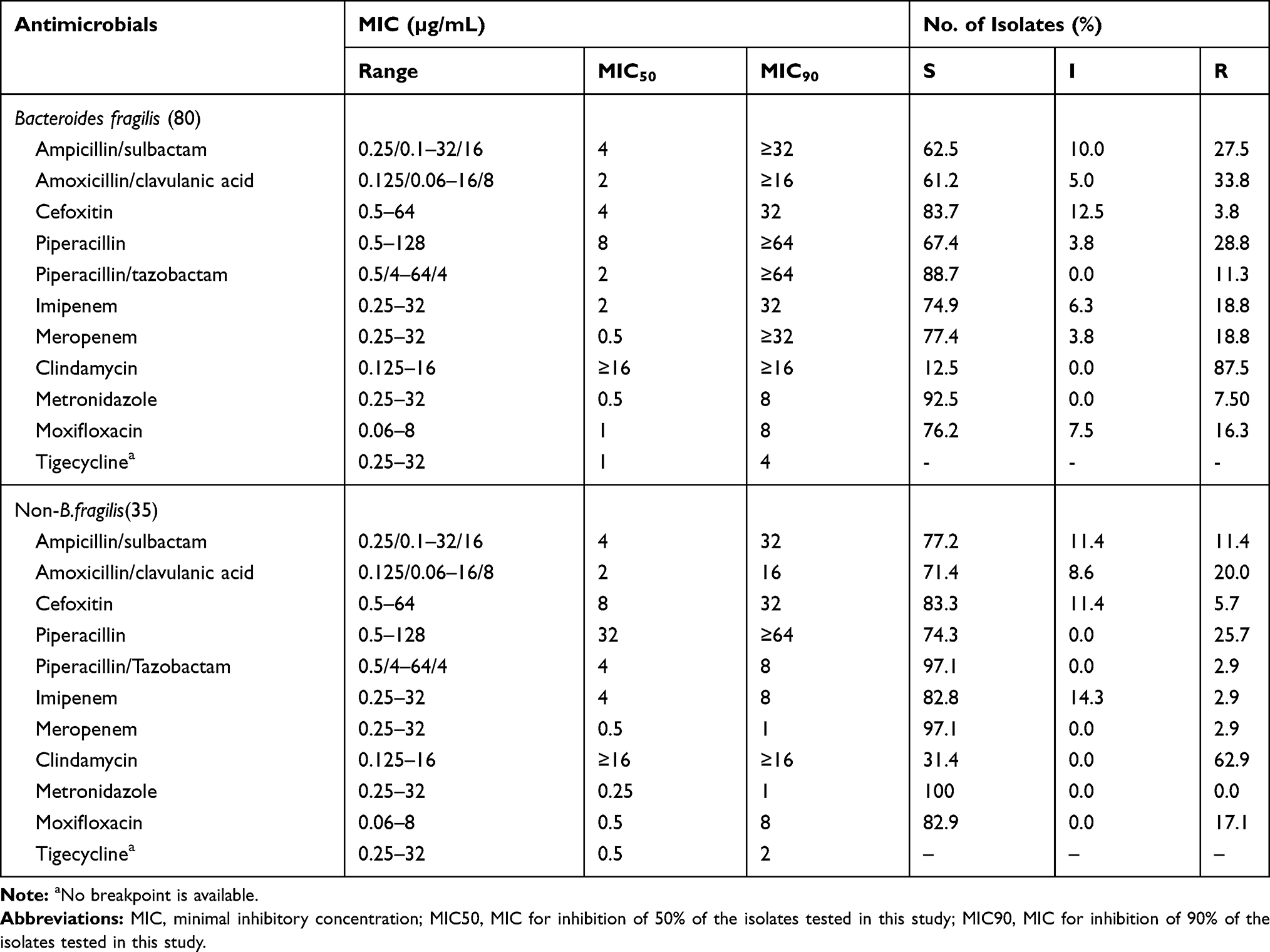 |
Table 2 In vitro Activities of 11 Antimicrobial Agents Against 115 Bacteroides spp. Isolates |
Distribution of Resistance Genes
Among 115 Bacteroides spp. isolates, the positive rates of resistance genes cfiA, cfxA, cepA, ermF and nim were 29.6% (34/115), 32.2% (37/115), 65.2% (75/115), 89.6% (103/115) and 30.4% (35/115), respectively. The positive rates of the above resistance genes of B. fragilis isolates were all significantly higher than those of non-B. fragilis isolates, as shown in Figure 1. A total of 31 B. fragilis and four non-B.fragilis isolates were positive when testing for the nim gene; however, only 6 B. fragilis isolates showed resistance to metronidazole.
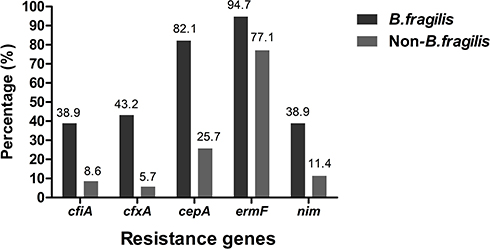 |
Figure 1 Prevalence of resistance genes among B. fragilis and non-B. fragilis isolates. |
For 15 imipenem resistant B. fragilis isolates, tests for the cfiA gene were all positive, and 13 isolates harbored the IS element, which belonged to IS1187 (11 isolates), IS942 (1 isolate) and IS1169 (1 isolate), as shown in Table 3. Only 1 non-B. fragilis isolate was found to be carbapenem-resistant, which was positive when testing for the cfiA gene; however, no IS element was detected (Figure 2). Among the 13 isolates with both a cfiA gene and an IS element, the BF-18 isolate had a significantly longer IS element (3290 bp) than any other IS elements of the other 12 isolates. Moreover, the IS sequence was an inverted repeat segment. Nonetheless, there were also two inverted repeats in another typical isolate (BF-15) that belonged to IS1187. The positional relations between the cfiA gene and the IS sequence within BF-18 and BF-15 isolates are shown in Figure 3.
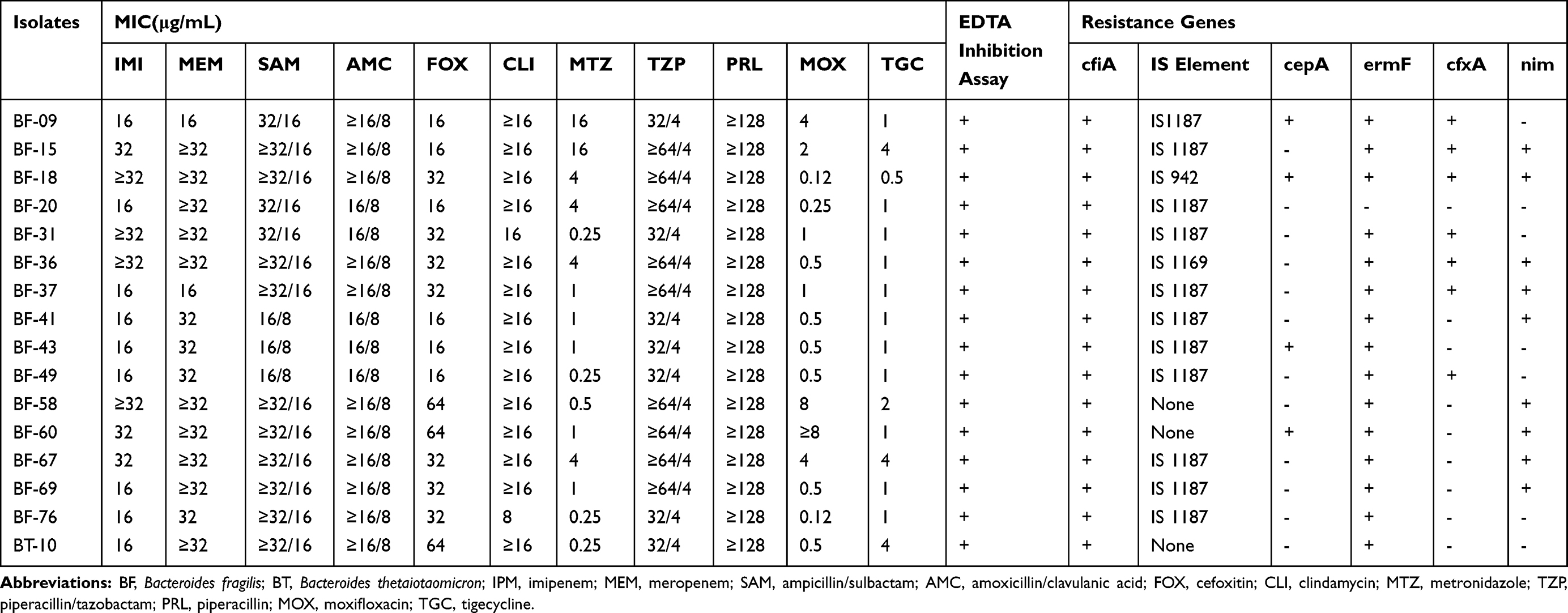 |
Table 3 Characteristics of 16 Carbapanem Resistant Bacteroidesspp. Isolates |
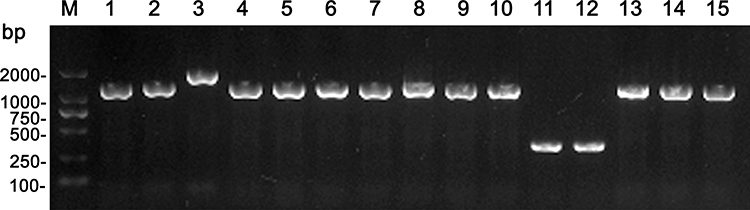 |
Figure 2 PCR results of cfiA and IS element of 15 carbapenem resistant B. fragilis isolates. Notes: 1, BF-09; 2, BF-15; 3, BF-18; 4, BF-20; 5, BF-31; 6, BF-36; 7, BF-37; 8, BF-41; 9, BF-43; 10, BF-49; 11, BF-58; 12, BF-60; 13, BF-67; 14, BF-69; 15, BF-76 |
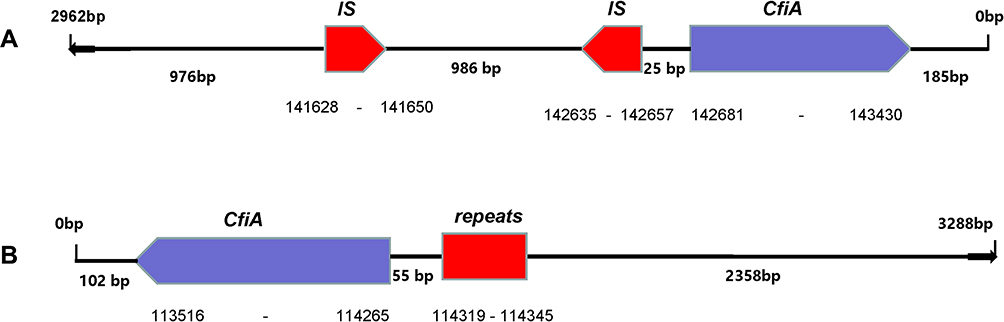 |
Figure 3 IS element analysis of BF-15 and BF-18 isolates. Notes: (A) BF-15 isolate; (B) BF-18 isolate. |
Effects of EPIs on MICs of Antimicrobials Tested
Two kinds of EPIs, CCCP and verapamil, were selected in this study as they can inhibit the RND-family and non-RND-family efflux pumps, respectively. A total of 21 MDR isolates with six different resistance patterns (a-h) were selected in this study. The antimicrobial MIC values of 10 Bacteroides spp. isolates (47.62%%, 10/21) showed a significant decrease by 4- to 64-fold, whereas the remaining isolates were not affected by EPIs. The MICs of four carbapenem-resistant isolates decreased significantly when EPIs were added. The same inhibition effects of CCCP and verapamil were also observed (Table 4). For three cefoxitin resistant B.fragilis isolates (BF-10, BF-58 and BF-60), neither CCCP nor verapamil showed an inhibitory effect on their MIC values. When 25 μg/mL of CCCP was added, the MICs of cefoxitin, moxifloxacin and imipenem for one B. fragilis isolate (BF-17) decreased simultaneously, as shown in Table 4. However, the isolate was not resistant to cefoxitin, moxifloxacin or imipenem. The MICs of moxifloxacin for four isolates decreased significantly when EPIs were added in eight moxifloxacin resistant isolates enrolled in this study. Lastly, the inhibition effects of CCCP and verapamil were not consistent among different isolates.
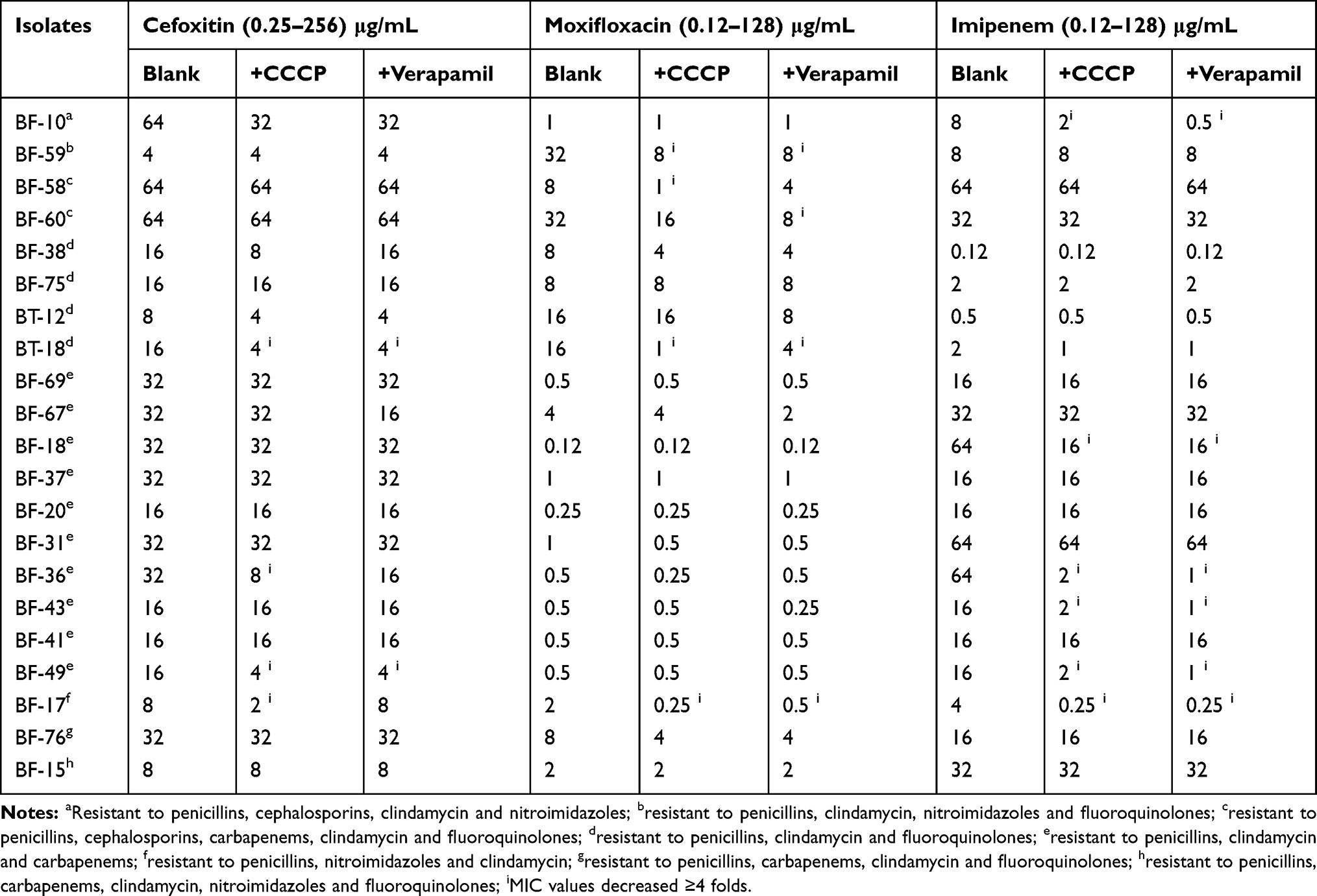 |
Table 4 Effects of EPIs on MICs of Cefoxitin, Moxifloxacin and Imipenem |
Discussion
In recent years, reports of resistance features in anaerobes have increased. However, only a few reports have come from China. In this study, 115 Bacteroides spp. clinical isolates were isolated from a 3000-bed tertiary teaching hospital in China that showed higher resistance rates to multiple commonly used antibiotics, including several β-lactams and metronidazole. The resistance rates of Bacteroides spp. isolates to carbapenem and metronidazole were at a higher level, and the isolates were mainly B. fragilis. It is well known that the mechanism of carbapenem resistance within B. fragilis is mainly mediated by the cfiA gene, and the upstream IS element is also required. In this study, the positive rate of the cfiA gene of carbapenem-resistant B. fragilis was as high as 38.9%, which is higher than other recently published reports.10,18,19 Notably, B. fragilis isolates with a cfiA gene can easily be converted to resistant genes by the effects of its upstream IS element,20 which implies that a larger group of patients may act as a potential reservoir of carbapenem-resistant B. fragilis producers who are colonized or infected by B. fragilis with a silent cfiA gene. Since the B. fragilis isolates tested in this study were mainly from intra-abdominal samples, intestinal screening of cfiA positive B. fragilis within a specific group of people appears to be a beneficial strategy for better understanding the actual distribution of the cfiA gene. So far, only a few reports have shown that an IS-less activation mechanism was found to exist in B. fragilis isolates with elevated imipenem MICs or imipenem resistant isolates.17 In this study, two B. fragilis isolates and one non-B. fragilis isolate resistant to carbapenem was found to be cfiA positive; however, the IS element was deficient. The potential non-IS resistance mechanism needs to be further investigated. In addition, a longer IS element (3290 bp) was detected in a carbapenem-resistant B. fragilis isolate, which was significantly different from other IS elements observed in this study. For this isolate, the MIC value of imipenem and meropenem were both higher than 32 μg/mL, which may possess a unique regulating mechanism in mediating carbapenem resistance. Furthermore, one B. fragilis isolate with an elevated imipenem MIC showed a reduction in their imipenem MIC value after exposure to the EPIs, CCCP and verapamil. Interestingly, the cfiA gene was not detected in this isolate. Moreover, the cfiA mediated resistance mechanism and the overexpression of the efflux pump may be another important mechanism in mediating carbapenem resistance in B. fragilis, especially for those with lower resistance to carbapenem.
Apart from carbapenem antibiotics, the resistance rates of B. fragilis isolates to several other β-lactam antibiotics were all higher than those of non-B. fragilis isolates, including ampicillin/sulbactam, amoxicillin/clavulanic acid, piperacillin and piperacillin/tazobactam. Consistent with a resistance phenotype, the prevalence of cepA and cfxA genes among B. fragilis isolates were significantly higher than those of non-B. fragilis isolates. Veloo et al5 reported that cfiA and cepA genes were collectively responsible for amoxicillin resistance, yet none of the B. fragilis isolates harbored both genes in this study. However, the resistance of amoxicillin was not detected, and a total of 20 B. fragilis isolates harbored both cfiA and cepA in this study. Notably, nine non-B. fragilis isolates harbored cepA, which was considerably different from what has been reported in previous reports.5,21,22 Kierzkowska et al10 reported that in B. fragilis, phenotypic cefoxitin resistance was strongly correlated with cfxA gene expression. However, only two B. fragilis and two non-B. fragilis isolates were resistant to cefoxitin, and none of the isolates harbored the cfxA gene in this study. Owing to the limitation of the number of isolates, the correlation between cefoxitin resistance and expression of cfxA gene cannot be confirmed.
The resistance rate of B. fragilis to metronidazole in this study was 7.5%, which is notably higher than previous reports.5–7,10,23,24 No metronidazole resistant non-B. fragilis isolate was detected either. To date, the mechanism of metronidazole resistance is still not well-known, and it is reported that nim genes closely correlate with metronidazole resistance.8,9,25 The positive rate of nim gene detection in this study was 38.9% and 11.4% for B. fragilis and non-B. fragilis isolates, respectively. However, the correlation between metronidazole resistance and expression of the nim gene was not observed. For six metronidazole resistant B. fragilis isolates, only three isolates were positive with the nim gene. The primers selected to amplify nim genes was a pair of universal primers, which could simultaneously detect nim A, nimB, nim C and nim D genes. Despite the sequence divergence of the nim genes, the primers were used for epidemiological investigations of nim genes in some previous studies.4,10 So, the nim genes reported in this study could not be assigned to a specific type of nim gene.
In this study, the total resistance rate of B. fragilis isolates against clindamycin was as high as 87.5%, which is significantly higher than that reported by Fernández-Canigia et al6 and Justesen et al.26 It is well known that erm (A-F) is widely distributed in Bacteroides spp. strains and are responsible for macrolide-lincosamide-streptogramin B resistance in B. fragilis.27 In the present study, the positive rate of the ermF gene (94.7%) was comparable with the phenotypic resistance rate (87.5%), which suggests that the ermF gene may be mainly responsible for B. fragilis resistance in this study.
The resistance rate to moxifloxacin was 16.5% and is significantly lower than that observed in another study from China.7 However, there is no apparent difference between B. fragilis isolates and non-B. fragilis isolates. It is reported that fluoroquinolone resistance may correlate with multiple mechanisms, including the mutation of the fluoroquinolone resistance determining region of the genes of gyrase and/or topoisomerase IV, as well as increased efflux.10,28 In this study, the effects of overexpression of the efflux pump on MIC values of moxifloxacin was observed in 50% (4/8) of the isolates investigated in this study, including 3 B. fragilis and 1 B. thetaiotaomicron isolates.
Moreover, both RND-family and non-RND-family EPIs showed significant inhibition effects on the MIC of the B. thetaiotaomicron isolate. The inhibition effects of non-RND-family efflux pump inhibitor on fluoroquinolone resistance of B. thetaiotaomicron has been confirmed in a previous study,28 and a multi-drug and toxic compound extrusion (MATE)-type efflux system has been well described.28 The specific features of the efflux pump system in this isolate needs to be further investigated.
For tigecycline, the MIC50 and MIC90 for both B. fragilis and non-B. fragilis isolates in this study were all higher than those reported in a previous study.6 Since there is no available breakpoint for tigecycline, it has not been feasible to assess its clinical significance, until now. In addition, the antimicrobial susceptibility testing method used in this study was a broth microdilution method; the results need to be further verified with the agar dilution method in a subsequent investigation.
During this study, a total of 21 multi-drug-resistant isolates were isolated from 21 different patients, 20 (95.24%, 20/21) of which were successfully cured with suitable antimicrobial combinations that were based on antimicrobial susceptibility test results; 19 patients among them were also co-infected with aerobic bacteria. One male patient who was 78 years old and had rectal cancer died during treatment; they had also received surgical intervention before B. fragilis isolation. Moreover, this patient developed bacteremia during the process of anti-infection treatment. Significantly, three patients aged 2, 4 and 13 years old, received surgical intervention for the treatment of acute appendicitis or hepatic abscess, and the MDR isolates should have originated from themselves. Notably, it is a stark reminder that resistance tendency of anaerobes among children may be underestimated or not well recognized, and universal monitoring resistance characteristics of anaerobic bacteria among specific groups of people should be given more attention.
Conclusion
The overall resistance rates of B. fragilis clinical isolates to several commonly used antimicrobials were at a high level, especially for carbapenem, and the dominant resistance mechanism to carbapenem was mediated by the cfiA gene. In addition, the resistance rate of Bacteroides spp. to metronidazole is at a high level in this study, and the actual resistance rate should be investigated with a larger quantity of isolates in the future. The actual resistance tendency of anaerobes may be underestimated in some geographical locations. The MDR B. fragilis isolates were more likely to be isolated from patients who had a history of exposure to cephalosporins at some point over a previous 3-month period. Moreover, it should be noted that the emergence of MDR B. fragilis among child patients implies that antimicrobial usage should be more strictly restricted. Furthermore, universal screening for the resistance determinants of B. fragilis should also be performed among specific groups of people in the districts without any strict regulations against antibiotic prescription.
Ethics Approval and Consent to Participate
This study is exempt from formal ethical approval and informed consent according to the local ethical guidelines, since the isolates identification and in vitro antimicrobial susceptibility test were routinely performed for normal clinical microbiological samples and the data analysis was analyzed anonymously.
Acknowledgments
This study is supported by Program for Young Talents of Science and Technology in Universities of Inner Mongolia Autonomous Region (NJYT-17-B06), Research Project of Affiliated Hospital of Inner Mongolia Medical University (NYFY YB042) and Major Project of Affiliated Hospital of Inner Mongolia Medical University (NYFY ZD 012). The two programs (NJYT-17-B06 and NYFY YB042) provide research expenditure for purchasing reagents and experimental consumables for gene detection related experiments, and the program (NYFY ZD 012) provide expenditure for whole genome sequencing of the two B. fragilis isolates.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Goldstein EJ, Citron DM, Vreni Merriam C, Warren Y, Tyrrell KL. Comparative in vitro activities of ertapenem (MK-0826) against 1,001 anaerobes isolated from human intra-abdominal infections. Antimicrob Agents Chemother. 2000;44(9):2389–2394. doi:10.1128/AAC.44.9.2389-2394.2000
2. Tan TY, Ng LS, Kwang LL, Rao S, Eng LC. Clinical characteristics and antimicrobial susceptibilities of anaerobic bacteremia in an acute care hospital. Anaerobe. 2017;43:69–74. doi:10.1016/j.anaerobe.2016.11.009
3. Yunoki T, Matsumura Y, Yamamoto M, et al. Genetic identification and antimicrobial susceptibility of clinically isolated anaerobic bacteria: a prospective multicenter surveillance study in Japan. Anaerobe. 2017;48:215–223. doi:10.1016/j.anaerobe.2017.09.003
4. Sarvari KP, Soki J, Kristof K, et al. Molecular characterisation of multidrug-resistant Bacteroides isolates from Hungarian clinical samples. J Glob Antimicrob Resist. 2018;13:65–69. doi:10.1016/j.jgar.2017.10.020
5. Veloo ACM, Baas WH, Haan FJ, Coco J, Rossen JW. Prevalence of antimicrobial resistance genes in Bacteroides spp. and Prevotella spp. Dutch clinical isolates. Clin Microbiol Infect. 2019;25(9):
6. Fernandez-Canigia L, Litterio M, Legaria MC, et al. First national survey of antibiotic susceptibility of the Bacteroides fragilis group: emerging resistance to carbapenems in Argentina. Antimicrob Agents Chemother. 2012;56(3):1309–1314. doi:10.1128/AAC.05622-11
7. Gao Q, Wu S, Xu T, Zhao X, Huang H, Hu F. Emergence of carbapenem resistance in Bacteroides fragilis in China. Int J Antimicrob Agents. 2019;53(6):859–863. doi:10.1016/j.ijantimicag.2019.02.017
8. Trinh S, Reysset G. Detection by PCR of the nim genes encoding 5-nitroimidazole resistance in Bacteroides spp. J Clin Microbiol. 1996;34(9):2078–2084. doi:10.1128/JCM.34.9.2078-2084.1996
9. Vishwanath S, Shenoy PA, Chawla K. Antimicrobial resistance profile and nim gene detection among Bacteroides fragilis group isolates in a University Hospital in South India. J Glob Infect Dis. 2019;11(2):59–62. doi:10.4103/jgid.jgid_116_18
10. Kierzkowska M, Majewska A, Szymanek-Majchrzak K, Sawicka-Grzelak A, Mlynarczyk A, Mlynarczyk G. The presence of antibiotic resistance genes and bft genes as well as antibiotic susceptibility testing of Bacteroides fragilis strains isolated from inpatients of the Infant Jesus Teaching Hospital, Warsaw during 2007–2012. Anaerobe. 2019;56:109–115. doi:10.1016/j.anaerobe.2019.03.003
11. Pumbwe L, Wareham DW, Aduse-Opoku J, Brazier JS, Wexler HM. Genetic analysis of mechanisms of multidrug resistance in a clinical isolate of Bacteroides fragilis. Clin Microbiol Infect. 2007;13(2):183–189. doi:10.1111/j.1469-0691.2006.01620.x
12. Cuchural GJ
13. Thompson JS, Malamy MH. Sequencing the gene for an imipenem-cefoxitin-hydrolyzing enzyme (CfiA) from Bacteroides fragilis TAL2480 reveals strong similarity between CfiA and Bacillus cereus beta-lactamase II. J Bacteriol. 1990;172(5):2584–2593. doi:10.1128/JB.172.5.2584-2593.1990
14. Kato N, Yamazoe K, Han CG, Ohtsubo E. New insertion sequence elements in the upstream region of cfiA in imipenem-resistant Bacteroides fragilis strains. Antimicrob Agents Chemother. 2003;47(3):979–985. doi:10.1128/AAC.47.3.979-985.2003
15. Clinical and laboratory standards institute. Methods for Antimicrobial Susceptibility Testing of Anaerobic Bacteria: CLSI Guidelines M100. Vol. 32.
16. Breakpoint tables for interpretation of MIC’s and zone diameters. Version 8.0. European committee on antimicrobial susceptibility testing; 2018.
17. Soki J, Fodor E, Hecht DW, et al. Molecular characterization of imipenem-resistant, cfiA-positive Bacteroides fragilis isolates from the USA, Hungary and Kuwait. J Med Microbiol. 2004;53(Pt 5):413–419. doi:10.1099/jmm.0.05452-0
18. Ferlov-Schwensen SA, Sydenham TV, Hansen KCM, Hoegh SV, Justesen US. Prevalence of antimicrobial resistance and the cfiA resistance gene in Danish Bacteroides fragilis group isolates since 1973. Int J Antimicrob Agents. 2017;50(4):552–556. doi:10.1016/j.ijantimicag.2017.05.007
19. Kouhsari E, Mohammadzadeh N, Kashanizadeh MG, et al. Antimicrobial resistance, prevalence of resistance genes, and molecular characterization in intestinal Bacteroides fragilis group isolates. APMIS. 2019;127(6):454–461. doi:10.1111/apm.12943
20. Podglajen I, Breuil J, Bordon F, Gutmann L, Collatz E. A silent carbapenemase gene in strains of Bacteroides fragilis can be expressed after a one-step mutation. FEMS Microbiol Lett. 1992;70(1):21–29. doi:10.1111/j.1574-6968.1992.tb05178.x
21. Garcia N, Gutierrez G, Lorenzo M, Garcia JE, Piriz S, Quesada A. Genetic determinants for cfxA expression in Bacteroides strains isolated from human infections. J Antimicrob Chemother. 2008;62(5):942–947. doi:10.1093/jac/dkn347
22. Tran CM, Tanaka K, Watanabe K. PCR-based detection of resistance genes in anaerobic bacteria isolated from intra-abdominal infections. J Infect Chemother. 2013;19(2):279–290. doi:10.1007/s10156-012-0532-2
23. Hansen KCM, Schwensen SAF, Henriksen DP, Justesen US, Sydenham TV. Antimicrobial resistance in the Bacteroides fragilis group in faecal samples from patients receiving broad-spectrum antibiotics. Anaerobe. 2017;47:79–85. doi:10.1016/j.anaerobe.2017.04.013
24. Yim J, Lee Y, Kim M, et al. Antimicrobial susceptibility of clinical isolates of Bacteroides fragilis group organisms recovered from 2009 to 2012 in a Korean hospital. Ann Lab Med. 2015;35(1):94–98. doi:10.3343/alm.2015.35.1.94
25. Sydenham TV, Soki J, Hasman H, Wang M, Justesen US. Identification of antimicrobial resistance genes in multidrug-resistant clinical Bacteroides fragilis isolates by whole genome shotgun sequencing. Anaerobe. 2015;31:59–64. doi:10.1016/j.anaerobe.2014.10.009
26. Justesen US, Hansen F, Ostergaard C, et al. High rates of reduced susceptibility in the Bacteroides fragilis group isolated from blood cultures–the first national survey in Denmark. Int J Antimicrob Agents. 2013;42(2):188–190. doi:10.1016/j.ijantimicag.2013.04.025
27. Chung WO, Werckenthin C, Schwarz S, Roberts MC. Host range of the ermF rRNA methylase gene in bacteria of human and animal origin. J Antimicrob Chemother. 1999;43(1):5–14. doi:10.1093/jac/43.1.5
28. Miyamae S, Ueda O, Yoshimura F, Hwang J, Tanaka Y, Nikaido H. A MATE family multidrug efflux transporter pumps out fluoroquinolones in Bacteroides thetaiotaomicron. Antimicrob Agents Chemother. 2001;45(12):3341–3346. doi:10.1128/AAC.45.12.3341-3346.2001
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.