Back to Journals » OncoTargets and Therapy » Volume 16
High BCR::ABL1 Expression Defines CD34+ Cells with Significant Alterations in Signal Transduction, Short-Proliferative Potential and Self-Renewal Ability
Authors Massimino M, Stella S ![]() , Tirrò E, Pennisi MS, Stagno F, Vitale SR
, Tirrò E, Pennisi MS, Stagno F, Vitale SR ![]() , Romano C
, Romano C ![]() , Tomarchio C, Parrinello NL, Manzella L, Di Raimondo F, Vigneri P
, Tomarchio C, Parrinello NL, Manzella L, Di Raimondo F, Vigneri P
Received 6 April 2023
Accepted for publication 12 July 2023
Published 3 October 2023 Volume 2023:16 Pages 803—816
DOI https://doi.org/10.2147/OTT.S413825
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Gaetano Romano
Michele Massimino,1,2,* Stefania Stella,2,3,* Elena Tirrò,2,3 Maria Stella Pennisi,2,3 Fabio Stagno,4 Silvia Rita Vitale,2,3 Chiara Romano,2,5 Cristina Tomarchio,2,3 Nunziatina Laura Parrinello,4 Livia Manzella,2,3 Francesco Di Raimondo,4 Paolo Vigneri2,3,6
1Department of Surgery and Medical-Surgical Specialties, University of Catania, Catania, Italy; 2Center of Experimental Oncology and Hematology, A.O.U. Policlinico “G. Rodolico-S. Marco”, Catania, Italy; 3Department of Clinical and Experimental Medicine, University of Catania, Catania, Italy; 4Division of Hematology, A.O.U. Policlinico “G. Rodolico-S. Marco”, Catania, Italy; 5Department of Medical, Surgical Sciences and Advanced Technologies “G.F. Ingrassia”, Anatomic Pathology, University of Catania, Catania, Italy; 6Humanitas Istituto Clinico Catanese, University Oncology Department, Catania, Italy
*These authors contributed equally to this work
Correspondence: Michele Massimino, A.U.O. Policlinico G. Rodolico S. Marco, Via Santa Sofia 78, Catania, 95123, Italy, Tel +39-95-3781952, Fax +39-95-3781949, Email [email protected]
Purpose: Chronic Myeloid Leukemia (CML) is a clonal disorder of the hematopoietic stem cell caused by expression of the BCR::ABL1 oncoprotein. High BCR::ABL1 levels have been associated to proliferative advantage of leukemic cells, blast crisis progression and tyrosine kinase inhibitors (TKIs) inefficacy. We have previously shown that high BCR::ABL1/GUSIS transcripts measured at diagnosis are associated with inferior responses to standard dose Imatinib (IM). However, the mechanisms underlying the higher rates of disease progression and development of TKIs resistance dependent on elevated BCR::ABL1 levels remain unclear.
Methods: Leukemic cells were collected from CML patients showing, at diagnosis, high or low BCR::ABL1/GUSIS. BCR::ABL1 expression levels were measured using real-time PCR. Short-term culture and long-term culture-initiating cells assays were employed to investigate the role of BCR::ABL1 gene-expression levels on proliferation, clonogenicity, signal transduction, TKIs responsiveness and self-renewal ability. Cell division was performed by carboxyfluorescein-succinimidyl ester (CFSE) assay.
Results: We found that BCR::ABL1 oncogene expression levels correlate in both PMNs and CD34+ cells. Furthermore, high oncogene levels increased both proliferation and anti-apoptotic signaling via ERK and AKT phosphorylation. Moreover, high BCR::ABL1 expression reduced the clonogenicity of leukemic CD34+ cells and increased their sensitivity to high doses IM but not to those of dasatinib. Furthermore, we observed that high BCR::ABL1 levels are associated with a reduced self-renewal of primitive leukemic cells and, also, that these cells showed comparable TKIs responsiveness with cells expressing lower BCR::ABL1 levels. Interestingly, we found a direct correlation between high BCR::ABL1 levels and reduced number of quiescent leukemic cells caused by increasing their cycling.
Conclusion: Higher BCR::ABL1 levels improving the proliferation, anti-apoptotic signaling and reducing self-renewal properties cause an increased expansion of leukemic clone.
Keywords: BCR:ABL1, self-renewal, CD34, TKIs, LTC-IC
Introduction
Chronic Myeloid Leukemia (CML) is a myeloproliferative disorder characterized by the expansion of a BCR::ABL1-driven leukemic clone displaying oncogene-dependent constitutive activation of several intracellular down-stream signals.1,2
The availability of first imatinib mesylate (IM), second-dasatinib, nilotinib, bosutinib (DAS, NIL, BOS) and third-generation ponatinib (PON) tyrosine kinase inhibitors (TKIs) has generated unprecedented rates of hematological, cytogenetic and molecular responses,3–5 that enable a number of patients to be considered for drug discontinuation achieving a durable treatment-free remission (TFR).6–8 Despite these unprecedented results, some CML patients will eventually fail their assigned TKI and, a smaller number of individuals, will progress to the deadly acute phase of the disease. While it is well established that BCR::ABL1 transforms CD34+ cells, however, discordant data are available concerning the correlation between high BCR::ABL1 transcripts, TKIs resistance and disease evolution towards blast crisis.9–14 We have previously reported the first interim results of the SCREEN (Sicily and Calabria CML REgional ENterprise) trial demonstrating that CML patients expressing high BCR::ABL1 at diagnosis achieve lower rates of transformation-free, event-free and failure-free survival compared to subjects with low BCR::ABL1.15 These findings suggest that individuals affected by the same disease, caused by the same genetic alteration, display different responses according to the expression level of their oncogenic driver.
Here, we investigate the role of high or low BCR::ABL1 expression in modulating the signal transduction, proliferation and self-renewal properties of primary leukemic progenitors collected, at diagnosis, from chronic-phase (CP) CML patients.
Materials and Methods
Subjects
Bone marrow (BM) and peripheral blood (PB) samples were collected from 26 patients with newly diagnosed chronic phase-CML (CP-CML). Patients were followed in the Division of Hematology of the A.O.U. Policlinico - “G. Rodolico – S. Marco”, Catania, Italy. BM samples were also collected from 20 healthy human donors (HDs) to isolate mesenchymal stem cells (MSCs) used for long-term culture-initiating cells (LTC-ICs) assays.
Patients and healthy donors signed an informed consent releasing anonymously their samples for research purposes in accordance with the Declaration of Helsinki. The study was conducted according to a protocol approved by Local Ethics Committee Catania 1, A.O.U. Policlinico-Vittorio Emanuele, identification code #34/2013/VE.
Isolation of Bone Marrow Mononuclear Cells and Mesenchymal Stem Cells
BM mononuclear cells (BM-MNCs) derived from HDs or CML patients at diagnosis were isolated by Ficoll-Paque Premium (GE Healthcare) density-gradient centrifugation according to the manufacturer’s protocol.
Immediately after isolation, fresh CML bone marrow MNCs were cryopreserved in freezing medium consisting of 10% dimethyl sulfoxide (DMSO, Sigma) in Fetal Bovine Serum (FBS) (Euroclone), while MNCs derived from HDs were directly plated as previously reported16 to obtain the mesenchymal stem cells (MSCs) used as feeder for LTC-IC assays.
Isolation of Polymorphonuclear Cells
For the recovery of polymorphonuclear (PMNs) cells, peripheral blood (PB) collected from CML patients was subjected to red cell lysis with ammonium chloride solution (0.8% NH4Cl, 0.1mM EDTA in water buffered with KHCO3 to achieve a final pH of 7.4) and frozen at −80°C in RLT buffer (Qiagen) for RNA extraction. PMNs derived from CML patients expressing low or high BCR::ABL1/GUSIS transcript were designed as PMNsBAL and PMNsBAH, respectively.
Isolation and Expansion of Human Hematopoietic CD34+ Progenitors
CD34+ progenitors were isolated from cryopreserved MNCs rapidly thawed at 37°C in a water bath and then resuspended in 50 mL of cool Iscove’s Modified Dulbecco’s Medium (IMDM) (Sigma-Aldrich) with 10% FBS. Pelleted cells were incubated with 100 µg/mL DNaseI (Stem Cell Technologies) in Hank’s balanced salt solution (HBSS) for 15 min at room temperature. CD34+ progenitors were then isolated by positive magnetic selection using the CD34 MicroBead kit (Miltenyi Biotec) as previously described.17 Overall purity of the final cell population was >90% as verified by flow cytometer analysis using anti-CD34-PE and anti-CD45-FITC antibodies (both Beckman Coulter).
Progenitor cells were grown in StemSpan Serum-Free Expansion medium (SFEM) supplemented, where indicated, with StemSpan CC100 Cytokine Cocktail at the following final concentrations of recombinant human (rh) cytokines: rh-Flt-3 ligand: 100 ng/mL; rh-stem cell factor: 100 ng/mL; rh interleukin-3, 20 ng/mL and rh interleukin-6, 20 ng/mL (all from Stem Cell Technologies). Cells were incubated at 37°C in a humidified atmosphere with 5% CO2.
CD34+ cells derived from patients expressing low or high BCR::ABL1/GUSIS transcript were identified as CD34+BAL and CD34+BAH.
RNA Extraction, cDNA Synthesis and Quantification of BCR::ABL1 Transcripts
For PMNs, RNA was extracted from 1×107 cells lysed in RLT buffer provided in the RNeasy mini kit (Qiagen) according to the manufacturer’s instructions. For progenitor cells, RNA was obtained from 5×105 freshly isolated CD34+ cells lysed in RLT using the RNeasy micro kit (Qiagen). One µg of total RNA from PMNs and CD34+ progenitors was reverse transcribed using M-MLV (Thermofisher Scientific) in the presence of random hexamers. BCR::ABL1 mRNA copy numbers in PMNs and CD34+ cells were quantified using the Applied Biosystem 7500 Real-Time PCR. Beta-glucuronidase (GUSB) mRNA was employed as the reference gene.18
Drug Treatments
CD34+ cells were exposed to 5 µM IM or 150 nM DAS for 24 hours. 10 mM stock solutions of IM in PBS (phosphate buffer saline) (Sigma-Aldrich) and DAS in DMSO were stored at −20°C. IM was obtained from Novartis, while DAS was a gift of Bristol-Myers Squibb and used according to the concentrations equivalent to achievable plasma levels.19 IM and DAS have been selected as they differently inhibit the ABL kinase activity when compared with Nilotinib which present a comparable pharmacophore with IM.20,21 The third-generation TKI, PON, was excluded because its use is confined to patients with resistance or intolerance of at least 2 prior TKI or showing T315I mutation22 and both populations were not included in this study.
Western Blotting and Immunoblotting
CD34+ cells were seeded at a concentration of 3x105/mL, cultured with cytokines and left untreated or exposed to IM or DAS for 24 hours. 1×105 cells were then lysed in 100 µL Laemmli buffer [62.5 mM Tris-HCl (pH 6.8), 2% w/v SDS, 10% glycerol, 50 mM dithiothreitol (DTT), 0.01% w/v bromophenol blue], sonicated, denatured and stored at −80°C. 30 µL of each protein lysate was loaded onto an SDS-PAGE gel, transferred to nitrocellulose membranes and, according to the antibody used, blocked with 5% nonfat dry milk or with 5% bovine serum albumin (BSA) (Sigma-Aldrich) in Tris-Buffered Saline with 0.1% Tween 20 (Sigma-Aldrich).
Primary antibodies used were: polyclonal anti-phospho-ABL1 (Tyr245), anti-phospho-AKT (Ser473), anti-AKT, anti-phospho-CRKL (Tyr207), anti-phospho-ERK1/2 (Thr202/Tyr204) and monoclonal anti-CRKL (clone 32H4) (all from Cell Signaling); anti-ABL1 (Santa Cruz Biotechnology); monoclonal anti-Actin (Sigma-Aldrich). After incubation with primary antibodies, appropriate horseradish peroxidase conjugated secondary antibodies (Amersham Biosciences) were added and proteins were then detected using the enhanced chemiluminescence (ECL) reagent Star (Euroclone) or WesternSure PREMIUM (Li-COR). Chemiluminescent images were digitally captured on the c-Digit blot scanner and a densitometric analysis was performed using the Image Studio Digits software version 5.2.5 (all from Li-COR).
Colony-Forming Unit Assays
500 CD34+ cells, untreated or exposed to drug treatment, were seeded in Methocult H4435 (StemCell Technologies) methylcellulose medium. Both total and differentiated myeloid colonies were counted under an optical microscope (Olympus IX71) after 14 days of culture. Pictures showing the human hematopoietic colonies have been acquired by Axio Observer Inverted Microscope (Zeiss).
Long Term Culture-Initiating Cells (LTC-ICs) Assays
LTC-IC frequency was calculated by Limiting Dilution Analysis (LDA). 1.5×104 MSCs were established as feeder layers in 96-well plates and blocked for 24 hours with 2 µg/mL Mytomicin C (Sigma Aldrich).23 CP-CML CD34+ progenitors were pretreated for 24 hours with IM or DAS and then plated on feeder cells using 40 replicates of two-fold cell dilutions (from 125 to 1000 initial test cells) in long-term culture medium (MyeloCult H5100 from StemCell Technologies) for 5 weeks with weekly half-medium changes. After 5 weeks, cells were overlaid with methylcellulose (Methocult H4435, StemCell Technologies) supplemented with conditioned medium derived from 5637 cells.24 Colonies were counted under the microscope after two additional weeks of growth, LTC-IC frequency was calculated using the L-Calc software (StemCell Technologies).
To measure LTC-IC-derived CFUs, we performed LTC-IC assays by bulk analysis with 5×104 CD34+ test cells cultivated on 445×103 MSCs cells treated as described above in a 35 mm dish. After 5 weeks, adherent and non-adherent cells were collected and 5×104 hematopoietic cells were resuspended in methylcellulose (H4435) as previously described.25,26 LTC-IC-derived CFUs were expressed as the number of clonogenic progenitors obtained after 15 days of methylcellulose culture (number of colonies multiplied by the total number of hematopoietic cells counted after 5 weeks of culture) divided by the number of initial test cells seeded on fibroblasts.25,26
CFSE Proliferation Assays
7x104 CD34+ progenitors were labeled with 10 μM CellTrace carboxy-fluorescein diacetate succinimidyl diester (CFSE; LifeTech) according to the manufacturer's instructions and then incubated in SFEM medium for 4 days without cytokines to avoid impairing BCR::ABL1-dependent proliferation. Cells were also cultured, in the same condition, for 24 hours in the presence of 0.1 μg/mL colcemid (Colchicine-Gibco) to establish CFSEmax (undivided cell population). Blocked and unblocked cells were stained with CD34-PECy7, CD38-PE-Cy5.5, CD45RA-ECD (all from Beckman Coulter), CD71-PE (Miltenyi) and analyzed by flow cytometer analysis. Data obtained for each experiment were analyzed using FCS Express 6 Plus (DeNovo software) set on the basis of the fluorescence profile obtained from cells blocked by colcemid. A representative gating strategy is reported in Supplemental Figure 1.
Statistical Analyses
Statistical analyses were performed using GraphPad Prism 5.0a (GraphPad Software Inc). Unpaired, single-tail t-tests with 95% confidence intervals and ANOVAs were used.
Results
Patient Selection
We have previously shown that individuals failing standard dose IM express higher BCR::ABL1/GUSIS levels (median 20.39%) in their PMN cells compared to patients achieving an optimal response (median 11.97%).15 We therefore defined BCR::ABL1/GUSIS values ≤12% as BCR::ABL1 low (BAL) while transcripts ≥20% were considered BCR::ABL1 high (BAH). We collected both PB and BM specimens from 26 chronic-phase CML patients at diagnosis and stratified them according to their BCR::ABL1/GUSIS levels in BAL (n=11) and BAH (n=15). Patient characteristics are summarized in Table 1.
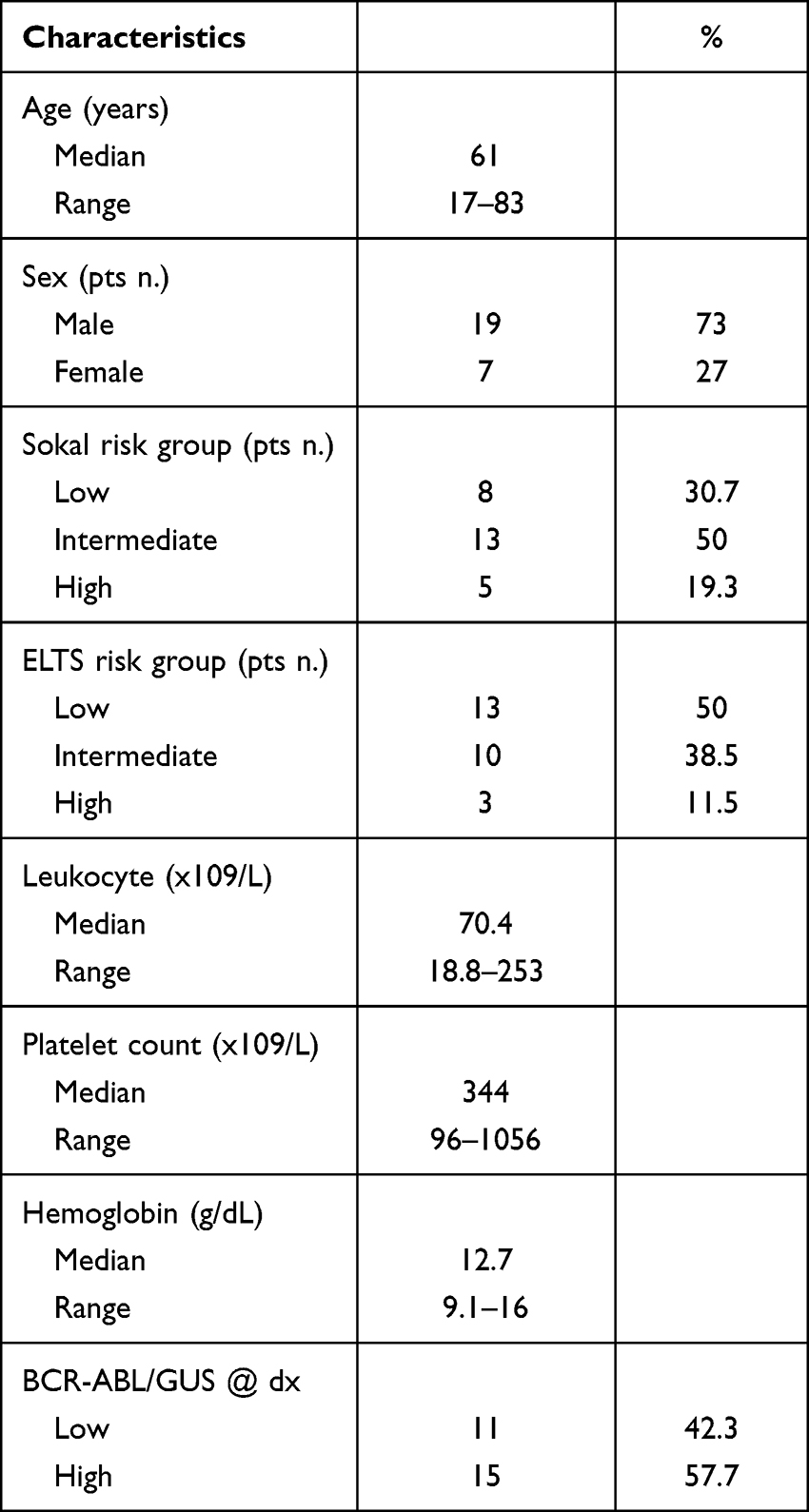 |
Table 1 Patient Characteristics (N = 26) |
BCR::ABL1 Expression in PMN Cells Correlates with BCR::ABL1 Levels in CD34+ Progenitors
We initially compared the BCR::ABL1 mRNA levels detected in PB PMNs and BM CD34+ progenitors and, as previously reported,27,28 found that CML progenitors displayed higher BCR::ABL1 mRNA levels than circulating PMNs (median CD34+ 54.56 vs PMNs 29.46) (Figure 1A).
 |
Figure 1 Correspondence between BCR::ABL1/GUSIS levels in PMNs mRNA and protein expression in CD34+ cells. (A) Scatter plot showing the median value with range of absolute copies of BCR::ABL1 mRNA in PMNs (n=26) or CD34+ cells (n=26). (B and C) Scatter plots indicate the median with range of BCR::ABL1 mRNA copies ((B) left and right panel) or BCR::ABL1/GUS% ((C) left and right panel) in PMNs and CD34+ cells derived from patients expressing low (PMNsBAL, CD34+BAL, n=11) or high (PMNsBAH, CD34+BAH, n=15) BCR::ABL1/GUSIS at diagnosis. (D) Histograms represent the median percentage value of BCR::ABL1 (left panel) or ABL1 (right panel) protein levels in CML patients expressing low BCR::ABL1/GUSIS set arbitrary at 100% (CD34+BAL, n=10) compared to those showing high BCR::ABL1/GUSIS (CD34+BAH, n=14). Bars indicate the standard deviation obtained from average value obtained from the entire cohort of patients. Unpaired t-test was used for statistical significance. |
We subsequently stratified PMNs and CD34+ cells according to the BCR::ABL1/GUSIS transcripts expressed by the 26 patients at diagnosis and measured absolute BCR::ABL1 mRNA copy numbers in each sample. We found that leukemic PMNs and CD34+ cells from patients displaying low oncogene levels (PMNsBAL and CD34+BAL) presented significantly lower BCR::ABL1 mRNA than the corresponding cells (PMNsBAH and CD34+BAH) from subjects expressing higher oncogene levels (PMNsBAL 17.68 vs 48.87 for PMNsBAH; CD34+BAL 46.57 vs 78.43 for CD34+BAH; Figure 1B, left and right panel). As the international scale conversion factor is used for diagnosis and monitoring of CML patients it is calculated on WBC (White Blood Cells), and it is not applicable for BCR::ABL1 mRNA quantification on CD34+ cells. Therefore, we calculated median oncogenic transcripts expressed as percentage of the BCR::ABL1/GUS ratio (BCR::ABL1/GUS%), thus normalizing BCR::ABL1 levels to the GUSB reference gene. We found a median value of 28.08% for PMNsBAL vs 87.82% for PMNsBAH, and 44.9% vs 93.8% when comparing CD34+BAL with CD34+BAH (Figure 1C, left and right panel).
We next evaluated the correlation between BCR::ABL1 mRNA levels and oncogenic protein quantity in CD34+BAL and CD34+BAH. To this end, we performed immunoblot experiments arbitrarily setting at 100% the densitometric average value of the BCR::ABL1 protein detected in CD34+BAL. Compared to these cells, we observed a 1.82-fold BCR::ABL1 increase in CD34+BAH (Figure 1D, left panel), that was confirmed when we measured the ABL1 protein levels used as a reference control as previously reported14,29 (Figure 1D, right panel).
High Oncogenic Transcripts Increase BCR::ABL1 Auto-Phosphorylation, and Differently Modulate Down-Stream Signaling in CD34+ Cells
We next performed immunoblot experiments to establish if the different BCR::ABL1 protein levels detected in CD34+BAH affect the oncoprotein’s auto-phoshporylation and differently modulate down-stream signaling (Supplemental Figure 2). Performing densitometric analysis, we found that BCR::ABL1 auto-phosphorylation was lower in CD34+BAL than in CD34+BAH (CD34+BAL 19.4% vs CD34+BAH 41.1%). We also found that IM and DAS abrogated BCR::ABL1 phosphorylation regardless of the amount of total protein detected in each cell population (Figure 2A).24,30 When we analyzed CRKL phosphorylation (pCRKL) in CD34+BAL and CD34+BAH we detected no significant differences in untreated cells (median CD34+BAL 26% vs CD34+BAH 21.4%). Moreover, both IM and DAS reduced pCRKL independent on BCR::ABL1 protein amount (fold reduction CD34+BAL 3.1 vs 3.3 in CD34+BAH) (Figure 2B).
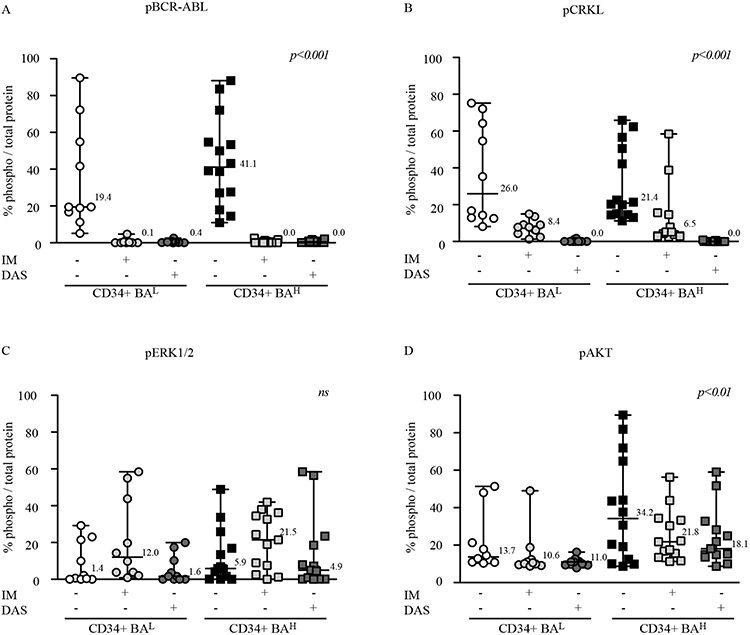 |
Figure 2 High BCR::ABL1 protein expression leads to an increase of autophosphorylation of AKT and ERK activation but show comparable responsiveness to IM or DAS treatment. (A–D) Scatter plots report the densitometric analysis expressed as median percentage value of the indicated phosphoprotein pBCR::ABL1 (A), pCRKL (B), pERK1/2 (C) and pAKT (D), normalized by each total protein previously normalized for actin. Cell lysates were obtained from CML patients expressing low (CD34+BAL, n=10) or high (CD34+BAH, n=14) BCR::ABL1/GUSIS. Cells were untreated or, dependent on their number, exposed to IM or DAS treatment for 24hrs before protein lysate preparation. Anova One-way was used to calculate statistical significance. |
Among the downstream pathways activated by BCR::ABL1, ERK plays a pivotal role in leukemogenesis.31 Although the observed effects on ERK modulation were not statistically significant, densitometric analysis revealed higher ERK phosphorylation in untreated CD34+BAH when compared with CD34+BAL (median value CD34+BAH 5.95% vs 1.4% for CD34+BAL). Exposure to IM and DAS generated very different results as the former drug increased ERK phosphorylation in both CD34+ populations while the latter compound marginally modified basal ERK phosphorylation levels (Figure 2C). The increase in phosphorylated ERK detected after IM treatment was expected as it has been previously reported that this TKI causes a paradox rise in phosphorylated ERK via an SRC-dependent mechanism.24 We also investigated AKT phosphorylation and observed that, in untreated CD34+ cells, higher BCR::ABL1 was associated with significantly improved AKT phosphorylation (median value 34.2% for CD34+BAH vs 13.7% in CD34+BAL). When we exposed the cells to different TKIs, we found that both IM and DAS reduced but did not abrogate the AKT phosphorylation, effect which was more prominent in CD34+BAH due to their higher levels of basal AKT phosphorylation. These data suggest that cells expressing high BCR::ABL1 levels show a higher AKT phosphorylation than cells with lower BCR::ABL1 amount even under TKIs exposure (Figure 2D).
High BCR::ABL1 Expression Affects the Clonogenicity of CD34+ Cells
CFU assay is a short-term culture evaluating the ability of a single cell to generate a colony measuring functionally its competence to duplicate itself overtime. We evaluated the impact of high or low BCR::ABL1 protein levels on this property, called clonogenicity, on committed CD34+ cells isolated from CML patients expressing low or high BCR::ABL1 levels. To this end, we performed CFU assays before and after exposure to IM or DAS observing that higher BCR::ABL1 levels generated significantly lower CFU than low levels (median value 170.5 for CD34+BAH vs 222 in CD34+BAL) and, also, that IM and DAS showed the same efficacy in both cell populations (Figure 3A). We subsequently enumerated erythroid burst-forming units (CFU-E/BFU-E) and granulocyte-monocyte units (GM-U) detecting that CD34+BAH generated less CFU-E/BFU-E (median value 116.5) compared to CD34+BAL (median value 167) (Figure 3B). On the contrary, a comparable number of GM-U between CD34+BAH (median value 45.5) and CD34+BAL (median value 59) was observed. Finally, IM and DAS reduced equally the number of colonies for both BFU-E and CFU-GEM colonies independent on BCR::ABL1 levels (Figure 3C). Representative pictures illustrating the colonies grown in semisolid methylcellulose medium are shown in Figure 3D.
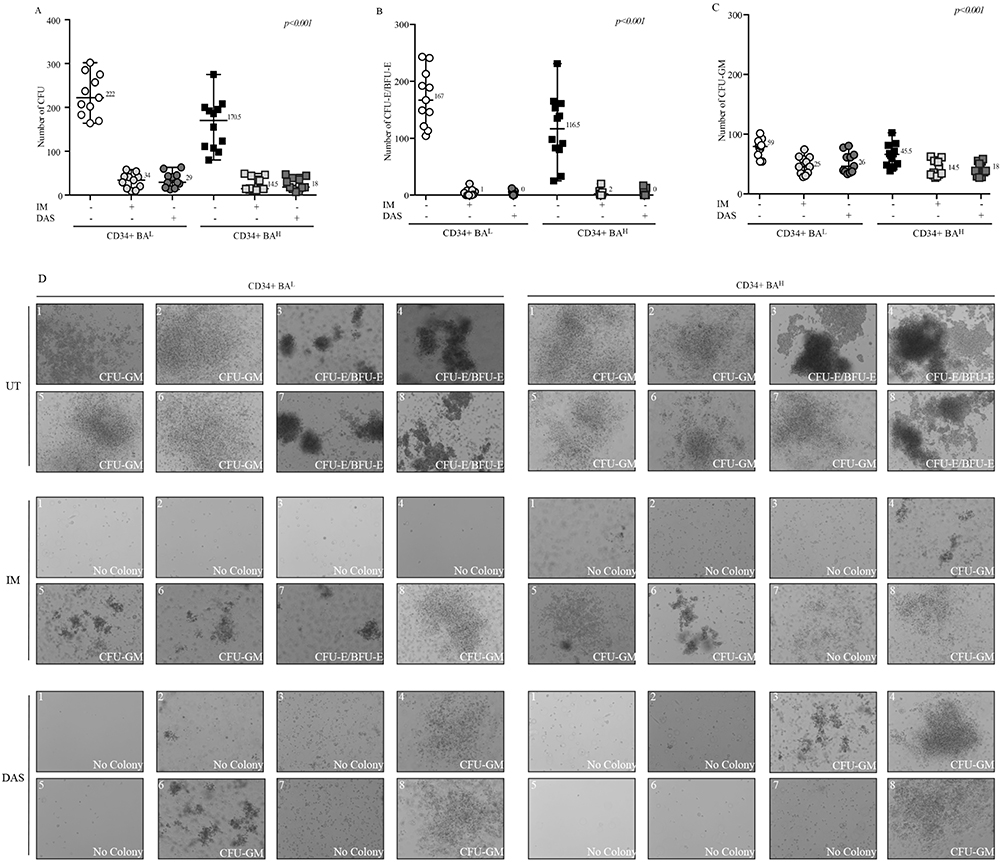 |
Figure 3 High BCR-ABL levels reduce the CD34+ clonogenicity and affect their responsiveness to TKIs. (A–C) Scatter plots show number of total CFU (A), CFU-E/BFU-E (B) and CFU-GM (C) obtained from short-term culture of CD34+ cells collected from CML patients expressing different BCR::ABL1/GUSIS levels (CD34+BAL, n=11) (CD34+BAH, n=12). Anova One-way was used to calculate statistical significance. (D) Pictures displaying colonies grown in methylcellulose medium. Each 35 mm dish was subject to acquisition of 8 representative sections by Axio Observer Inverted Microscope.Abbreviations: UT, untreated; IM, Imatinib; DAS, Dasatinib. |
High BCR::ABL1 Levels Reduce Self-Renewal Properties of Primitive Leukemic Cells
The group of Connie Eaves has clearly established that primitive CML cells exhaust their self-renewal capacity more rapidly than healthy hematopoietic progenitors.32 To investigate if high BCR::ABL1 levels modifies this biological feature of primitive leukemic progenitors, we performed LTC-IC assays by LDA using both CD34+BAH and CD34+BAL before and after exposure to IM or DAS. We observed that high BCR::ABL1 expression was associated with lower LTC-IC frequencies (median value in CD34+BAH 1:1073 vs 1:724 for CD34+BAL). However, the exposure to IM or DAS significantly yet equally reduced LTC-IC frequencies in the two cell populations (IM fold reduction 1.47 for CD34+BAH vs 1.31 in CD34+BAL; DAS fold reduction 1.53 for CD34+BAH vs 2.2 in CD34+BAL) (Figure 4A).
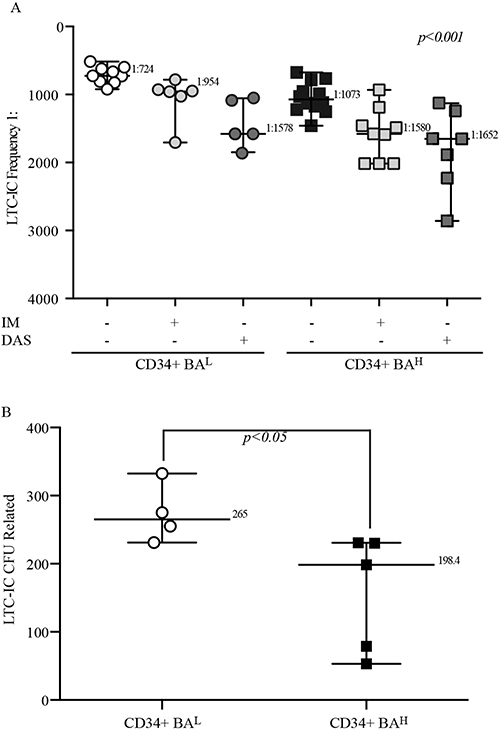 |
Figure 4 High BCR::ABL1 levels reduce the self-renewal properties of primitive leukemic cells but not affect their sensitivity to IM or DAS exposure. (A) The scatter plot report the LTC-IC frequency data of primitive leukemic cells derived from CML patients expressing low (CD34+BAL, n=9) or high (CD34+BAH, n=12) BCR::ABL1/GUSIS untreated or, dependent on their number, exposed to IM or DAS for 24hrs before the LTC-IC assay. Anova One-way was used to calculate statistical significance. (B) scatter graph shows the number of LTC-IC-derived CFU of primitive leukemic cells obtained from CML patient with different BCR::ABL1/GUSIS (CD34+BAL, n=4) (CD34+BAH, n=5). Unpaired t-test was used for statistical significance. For all experiments the number reported for each data set indicates the median value. |
To investigate if the reduced LTC-IC frequency shown by CD34+BAH was dependent on their disability to maintain their self-renewal properties, we measured the LTC-IC-derived CFUs to analyze their ability to perpetuate the myeloid progenies formation overtime after LTC-IC assay. Interestingly, we found that the survived cells after LTC-IC showing higher oncogenic transcripts displayed a significantly lower ability for generation LTC-IC-derived colony-forming units when compared to those expressing low BCR::ABL1 levels (median CFUs 198,4 for CD34+BAH vs 265 in CD34+BAL) (Figure 4B). Overall, these data support the implication that higher oncogene levels reduce the self-renewal of leukemic primitive cells.
High BCR::ABL1 Favors Cycling of Both Committed CD34+ Progenitors and More Quiescent Primitive Leukemic Cells
As published data indicate that BCR::ABL1 increases the proliferation of committed CD34+ leukemic progenitors,28,33,34 we wanted to investigate how high or low oncogenic transcripts would differently modulate the proliferation of committed CML progenitors. We therefore stained CD34+ cells expressing either high or low BCR::ABL1 levels with CFSE and found that – compared to cells blocked with colchicine – CD34+BAL and CD34+BAH showed a similar number of proliferating cells (86.14% average value for CD34+BAH vs 87.7% for CD34+BAL) (Figure 5A). However, when we analyzed the number of committed progenitors in each generation, we observed that CD34+BAH cycling cells were more frequent in the second, third and fourth generation if compared to CD34+BAL that maintained a greater number of cells either in the undivided population (average value D34+BAH 6.78% vs 9.5% in CD34+BAL) or in the first generation (average value 8.9% in CD34+BAH vs 20.6% in CD34+BAL) (Figure 5B).
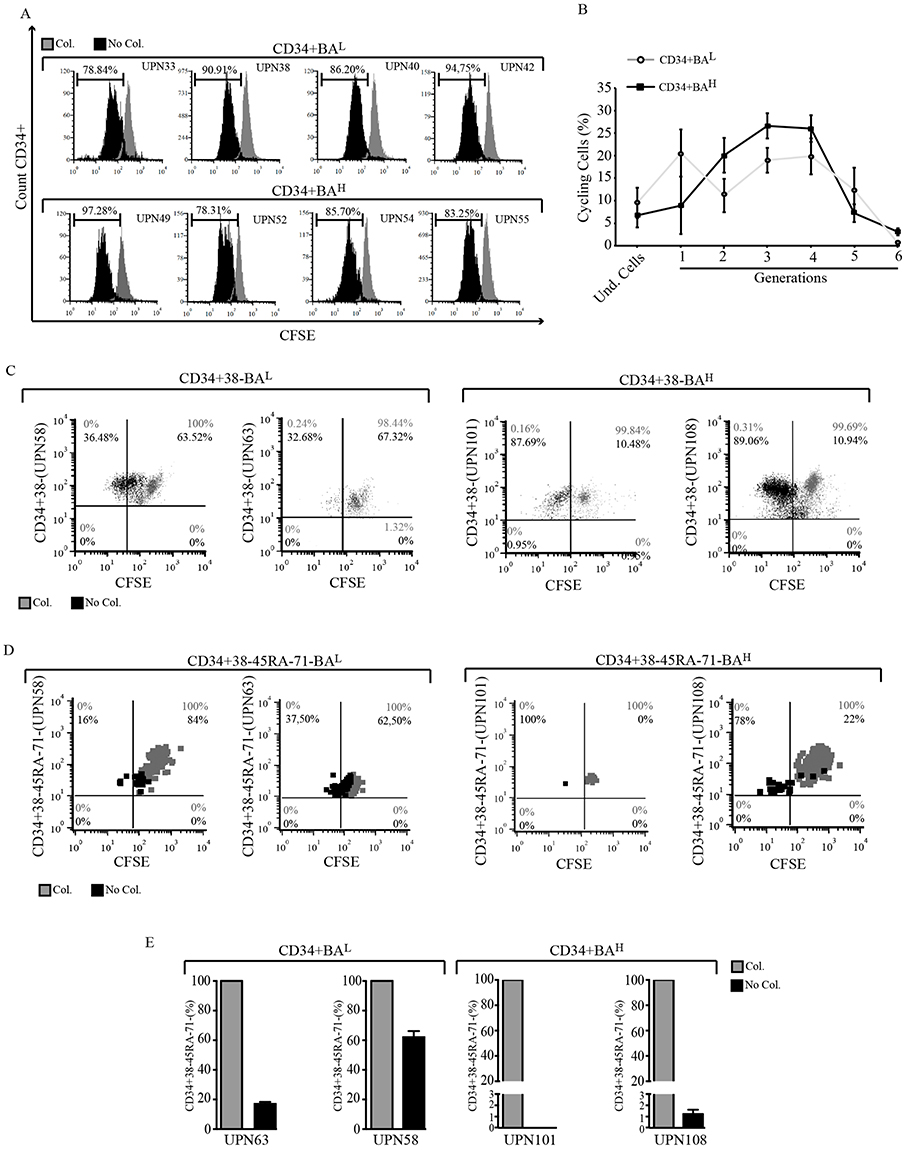 |
Figure 5 High BCR::ABL1 levels increase cell division of committed CD34+ progenitors and of quiescent leukemic. (A) Overlay histograms show the CFSE intensity of CD34+ cells (black histograms) derived from patients expressing low [UPN (Under Progression Number) 33, 38, 40 and 42] or high (UPN 49, 52, 54 and 55) BCR::ABL1/GUSIS. Gray histograms represent the same cells blocked by colcemid. The horizontal bar indicates the percentage of proliferating cells. (B) Curves indicate the average of the percentage of cycling cells within each generation during 4 days of culture. Bars indicate the standard deviation derived from average value obtained from entire cohort of patients showing low (CD34+BAL, n=4) or high (CD34+BAH, n=4) BCR::ABL1/GUSIS. (C and D) Dot plots derived by cytofluorimetric analysis show the CFSE intensity of CD34+CD38- (C) or CD34+CD38-CD45RA-CD71- (D) cultivated for 5 days (black dots) and compared to those blocked by colcemid for 24 hrs (gray dots). The number in each quadrant indicates the percentage of no cycling (upper right) or cycling (upper left) cells. (E) Histograms indicate the percentage of CD34+CD38-CD45RA-CD71- cells remaining after 4 days of culture (black column) compared to cells blocked by colchicine set arbitrary at 100% (gray column). For all experiments the cells were collected from CML patients (UPN) expressing low (CD34+BAL, n=2) or high (CD34+BAH, n=2) BCR::ABL1/GUSIS. |
To better understand the BCR::ABL1-dependent mechanisms leading to self-renewal reduction in progenitors expressing high BCR::ABL1 transcripts we repeated the CFSE experiment, staining both quiescent CD34+CD38- cells and the more quiescent phenotype CD34+CD38-CD45RA-CD71-.35 Compared to cells blocked by colchicine, we found that high BCR::ABL1 expression increased the proliferating fraction of CD34+CD38- progenitors (average UPN101/108 = 88.38% for BAH vs average UPN58/63 = 34.6% in BAL) at the expense of the non-dividing cell fraction (BAH average UPN101/108 = 10.71% vs UPN58/63 = 65.42% in BAL) (Figure 5C). Interestingly, we found similar results when we studied the more quiescent phenotype CD34+CD38-CD45RA-CD71- with a higher percentage of proliferating cells (average BAH UPN101/108 = 89% vs UPN58/63 = 26.75% in BAL), a lower number of non-dividing cells (average BAH UPN101/108 = 11% vs UPN58/63 = 73.25% for BAL) (Figure 5D), and a strong reduction of the population maintaining the CD34+CD38-CD45RA-CD71- phenotype (Figure 5E).
Discussion
In the present study, we investigated the biological consequences of opposite (ie high or low) endogenous BCR::ABL1 levels on the signal transduction, proliferation rates and clonogenic potential of committed leukemic progenitors. We also examined whether elevated BCR::ABL1 expression affected the self-renewal properties of primitive leukemic cells and assessed to what extent a first- (IM) or a second-generation (DAS) TKI influenced these biological phenomena. We find that high BCR::ABL1 transcripts define both leukemic progenitors and primitive leukemic cells with different biological behaviors in terms of proliferation, clonogenicity and self-renewal. We also find that these cell populations exhibit different sensitivities to IM and DAS.
Our results raise several interesting considerations. Previous data indicated that CD34+ cells express higher BCR::ABL1 mRNA levels than differentiated PMN cells.27,28 We confirm these results and also report that there is a direct correlation between the amount of BCR::ABL1/GUSIS expressed in PB PMNs and those detected in BM CD34+ cells. Thus, the BCR::ABL1 levels found in PB essentially reflect the oncogenic transcripts expressed by BM CML progenitors. Our findings also expand the previous results published by Junia Melo’s group showing that BCR::ABL1 mRNA and protein expression are lower in CD34+ cells isolated from patients in the chronic phase of the disease as compared to those in blast crisis.36 Indeed, we found a direct correlation between BCR::ABL1 transcripts at diagnosis and the expression levels of the chimeric oncoprotein in CP-CML patients at diagnosis. Hence, considering that elevated BCR::ABL1 expression is a pivotal characteristic of the transition between the chronic phase and the acute phase of CML, our data suggest that the fate of the disease may be already determined at the time of diagnosis.
Another interesting finding is the direct correlation found in CD34+ cells between BCR::ABL1 protein expression and its phosphorylation levels as well as the activation of its common downstream targets. These results are in agreement with those previously reported by Hardik et al that, after lentiviral transduction of healthy CD34+ cells with different levels of BCR::ABL1, found that higher oncogenic expression was associated with increased phosphorylation of BCR::ABL1, ERK and STAT-5.14 However, in contrast with our findings, the authors failed to detect systematic differences in AKT phosphorylation. This discordance may be attributable to the fact that, unlike Hardik et al, we employed primary human specimens selected – at diagnosis – for higher or lower endogenous BCR::ABL1 transcripts. Finally, in agreement with previous evidence, we found a paradoxical increase in ERK phosphorylation after primary leukemic CD34+ cells were treated with IM.24 The involvement of a SRC-dependent mechanism behind this phenomenon was confirmed by its suppression after exposure to the dual ABL/SRC inhibitor DAS. Moreover, the greater increase in ERK phosphorylation after IM exposure detected in CD34+BAH compared to CD34BAL, suggests a stronger SRC signaling in cells expressing higher BCR::ABL1.
An unexpected observation of our study was the lower CFU number generated by CD34+BAH in short-term culture assays. Interestingly, this event was mostly dependent on a reduction in the number of leukemic BFU-Es. Our results are not consistent with those reported by Hardik et al that found an increased number of both erythroid and myeloid progenitors in CD34+ cells transduced with high copies of BCR::ABL1.14 Again, these contrasting results may be attributed to ectopic BCR::ABL1 expression but may also be due to different cytokine concentrations used in the culture medium employed for these experiments. It should also be noted that, as the reduction in erythroid progenitors detected after IM and DAS treatment was significantly superior to that observed for the other myeloid precursors, the former cells would appear to heavily depend on BCR::ABL1 catalytic activity for their expansion and survival.
Our data reporting the ability of high dose of IM or DAS to reduce more effectively the CFU number of CD34+BAH than CD34+BAL, are consistent with those published by Hardik et al14 (Hardik Modi, Blood 2007), suggesting an improvement of dependence on BCR::ABL1 catalytic activity for their survival and proliferation. Furthermore, these data accord to results by Kumari et al,13 indeed, they reported a persistence of CD34+ cells expressing low BCR::ABL1 after IM exposure indicating a rapid initial depletion of the leukemic clone expressing higher BCR::ABL1 levels.
To investigate the reduction in CFU numbers observed in CD34+BAH progenitors we performed LTC-IC assays. In agreement with previously published data generated in transgenic mouse models37 or primary leukemic cells,32 we confirm that high BCR::ABL1 expression increases the differentiation of long-term hematopoietic leukemic stem cells and decreases their self-renewal properties. Indeed, CD34+BAH primitive leukemia cells showed both a significantly lower LTC-IC frequency than CD34+BAL cells and a lower clonogenic potential. This observation raises an important clinical implication concerning why the CML clone expressing higher BCR::ABL1 levels does not exhaust more rapidly than clone with lower oncogene levels. It was demonstrated that blast cells acquire self-renewal ability resulting as reservoir promoting diseases progression.38 BCR::ABL1 leads an increased CML transformation and genetic aberrations by alteration of DNA repair process39 which can drive to self-renewal maintenance by neutralization of inhibiting mechanism within a leukemic stem cell.37 We hypothesize that cells expressing higher BCR::ABL1 gene expression levels more distinctly acquire these molecular alterations than those showing lower oncogene levels. Hence, although our data demonstrate that high BCR::ABL1 levels enhanced cell differentiation reducing thus their self-renewal properties, if possible that high oncogene expression does not abolish completely the repopulating ability of leukemic stem cells promoting their persistence and disease progression.
Moreover, we found that primitive leukemic cells were modestly responsive to IM or DAS suggesting that, unlike committed progenitors, cells displaying a more primitive phenotype are not dependent on BCR::ABL1 protein levels for their survival. Furthermore, the observation that high dose of IM and DAS comparable reduced the LTC-IC frequency of CD34+BAH and CD34+BAL highlight an important correlation with results obtained from our SCREEN study,15 suggesting that patients expressing high BCR::ABL1 gene-expression levels can benefit of high dose of IM or DAS.
Furthermore, the observation that high BCR::ABL1 levels are associated with an increasing number of cycling cells indicates that high oncogenic transcripts favor the differentiation of CD34+ cells, thus reducing both their clonogenic potential and self-renewal ability. Interestingly, the same results were observed in quiescent (CD34+CD38-) and more quiescent (CD34+CD38-CD45RA-CD71-) leukemic stem cells, strongly supporting the notion that high oncogene levels promote the expansion of the leukemic clone according to its expression level.
Conclusion
In conclusion, with this work we demonstrate that high BCR::ABL1 expression leads to increased proliferation and anti-apoptotic signaling in CD34+ committed leukemic progenitors, promoting also stem cell division. It remains to be established if the higher proliferation rates observed in this cell populations lead to the acquisition of additional genomic abnormalities that could explain the inferior outcome of CML patients exhibiting high BCR::ABL1 at diagnosis. An additional, but not mutually exclusive, explanation may be that higher BCR::ABL1 levels modify the expression profile of a plethora of genes ultimately responsible for disease progression and the development of TKIs resistance. Attempting to approach mechanisms leading to persistence of leukemic stem cells, we tracked a different profile of CD34+ cells expressing higher BCR::ABL1 compared to those showing low oncogene levels. Our intriguing observations implicate that according to BCR::ABL1 levels at diagnosis, CML patients should be subjected to different clinical management. CML patients showing higher BCR::ABL1 levels at diagnosis would be exposed at an higher risk to develop disease progression caused by higher proliferation rate which can induce additional molecular alterations which increase the persistence of peripheral blast cells.33–35 However, remain to establish why the reduced cell renewal property, showed by CD34+BAH, does not result in an auto-limitation of the disease. Lastly, Further experiments assessing DNA integrity and RNA expression analyses will have to be performed in both CD34+BAH and CD34+BAL cells to verify these hypotheses.
Abbreviations
AKT, V-Akt murine thymoma viral oncogene-like protein 1; BFU-E, burst-forming uniterythroid; BM, bone marrow; BOS, Bosutinib; BSA, bovine serum albumin; CFSE, carboxyfluorescein-succinimidyl ester; CFU, colony forming unit; CFU-E, colony forming unit-erythroid; CML, chronic myeloid leukemia; CP, chronic phase; CRKL, CRK like proto-oncogene, adaptor Protein; DAS, Dasatinib; DMSO, dimethyl sulfoxide; EDTA, ethylenediaminetetraacetic acid; ERK, extracellular signal-regulated kinase; FBS, fetal bovine serum; FITC, fluorescein-5-isothiocyanate; GUSB, β-glucuronidase; HD, healthy donor; IM, imatinib mesylate; LDA, limiting dilution analysis; LTC-IC, long-term culture-initiating cell; M-MLV, moloney murine leukemia virus; MNC, mononuclear cell; MSC, mesenchymal stem cell; PB, peripheral blood; PE, phycoerythrin; PMN, polymorphonuclear; PON, ponatinib; SCREEN, Sicily and Calabria CML REgional ENterprise; SDS-PAGE, sodium dodecyl sulfate poly-acrylamide gel electrophoresis; SRC, V-Src Avian Sarcoma (Schmidt-Ruppin A-2) Viral Oncogene Homolog; TFR, treatment-free remission; TKI, tyrosine kinase inhibitor.
Acknowledgments
This work was supported by Grant AIRC number 12958 and PRIN 2017 CUP 64I19000710001.
Disclosure
Fabio Stagno reports personal fees from Pfizer, Novartis, and Incyte, outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Massimino M, Stella S, Tirro E, et al. ABL1-directed inhibitors for CML: efficacy, resistance and future perspectives. Anticancer Res. 2020;40(5):2457–2465. doi:10.21873/anticanres.14215
2. Ren R. Mechanisms of BCR-ABL in the pathogenesis of chronic myelogenous leukaemia. Nat Rev Cancer. 2005;5(3):172–183. doi:10.1038/nrc1567
3. O’Hare T, Walters DK, Stoffregen EP, et al. In vitro activity of bcr-abl inhibitors AMN107 and BMS-354825 against clinically relevant imatinib-resistant abl kinase domain mutants. Cancer Res. 2005;65(11):4500–4505. doi:10.1158/0008-5472.CAN-05-0259
4. Rosti G, Castagnetti F, Gugliotta G, Baccarani M. Tyrosine kinase inhibitors in chronic myeloid leukaemia: which, when, for whom? Nat Rev Clin Oncol. 2017;14(3):141–154. doi:10.1038/nrclinonc.2016.139
5. Shah NP, Tran C, Lee FY, chen P, Norris D, Sawyers CL. Overriding imatinib resistance with a novel ABL kinase inhibitor. Science. 2004;305(5682):399–401. doi:10.1126/science.1099480
6. Etienne G, Guilhot J, Rea D, et al. Long-term follow-up of the French Stop Imatinib (STIM1) Study in patients with chronic myeloid leukemia. J Clin Oncol. 2017;35(3):298–305. doi:10.1200/JCO.2016.68.2914
7. Hochhaus A, Baccarani M, Silver RT, et al. European Leukemia Net 2020 recommendations for treating chronic myeloid leukemia. Leukemia. 2020;34(4):966–984. doi:10.1038/s41375-020-0776-2
8. Shah NP. NCCN guidelines updates: discontinuing TKI therapy in the treatment of chronic myeloid leukemia. J Natl Compr Canc Netw. 2019;17(5.5):611–613. doi:10.6004/jnccn.2019.5013
9. Barnes DJ, Palaiologou D, Panousopoulou E, et al. Bcr-Abl expression levels determine the rate of development of resistance to imatinib mesylate in chronic myeloid leukemia. Cancer Res. 2005;65(19):8912–8919. doi:10.1158/0008-5472.CAN-05-0076
10. Gorre ME, Mohammed M, Ellwood K, et al. Clinical resistance to STI-571 cancer therapy caused by BCR-ABL gene mutation or amplification. Science. 2001;293(5531):876–880. doi:10.1126/science.1062538
11. Jiang X, Forrest D, Nicolini F, et al. Properties of CD34+ CML stem/progenitor cells that correlate with different clinical responses to imatinib mesylate. Blood. 2010;116(12):2112–2121. doi:10.1182/blood-2009-05-222471
12. Keeshan K, Mills KI, Cotter TG, McKenna SL. Elevated Bcr-Abl expression levels are sufficient for a haematopoietic cell line to acquire a drug-resistant phenotype. Leukemia. 2001;15(12):1823–1833. doi:10.1038/sj.leu.2402309
13. Kumari A, Brendel C, Hochhaus A, Neubauer A, Burchert A. Low BCR-ABL expression levels in hematopoietic precursor cells enable persistence of chronic myeloid leukemia under imatinib. Blood. 2012;119(2):530–539. doi:10.1182/blood-2010-08-303495
14. Modi H, McDonald T, Chu S, Yee J-K, Forman SJ, Bhatia R. Role of BCR/ABL gene-expression levels in determining the phenotype and imatinib sensitivity of transformed human hematopoietic cells. Blood. 2007;109(12):5411–5421. doi:10.1182/blood-2006-06-032490
15. Vigneri P, Stagno F, Stella S, et al. High BCR–ABL/GUSIS levels at diagnosis of chronic Phase CML are associated with unfavorable responses to standard-dose imatinib. Clin Cancer Res. 2017;23(23):7189–7198. doi:10.1158/1078-0432.CCR-17-0962
16. Pittenger MF, Mackay AM, Beck SC, et al. Multilineage potential of adult human mesenchymal stem cells. Science. 1999;284(5411):143–147. doi:10.1126/science.284.5411.143
17. Massimino M, Tirro E, Stella S, et al. Impact of the breakpoint region on the leukemogenic potential and the TKI responsiveness of atypical BCR-ABL1 transcripts. Front Pharmacol. 2021;12:669469. doi:10.3389/fphar.2021.669469
18. Muller MC, Erben P, Saglio G, et al. Harmonization of BCR-ABL mRNA quantification using a uniform multifunctional control plasmid in 37 international laboratories. Leukemia. 2008;22(1):96–102. doi:10.1038/sj.leu.2404983
19. Bradeen HA, Eide CA, O’Hare T, et al. Comparison of imatinib mesylate, dasatinib (BMS-354825), and nilotinib (AMN107) in an N-ethyl-N-nitrosourea (ENU)-based mutagenesis screen: high efficacy of drug combinations. Blood. 2006;108(7):2332–2338. doi:10.1182/blood-2006-02-004580
20. Buffa P, Romano C, Pandini A, et al. BCR-ABL residues interacting with ponatinib are critical to preserve the tumorigenic potential of the oncoprotein. FASEB J. 2014;28(3):1221–1236. doi:10.1096/fj.13-236992
21. Yang M, Xi Q, Jia W, Wang X. Structure-based analysis and biological characterization of imatinib derivatives reveal insights towards the inhibition of wild-type BCR-ABL and its mutants. Bioorg Med Chem Lett. 2019;29(24):126758. doi:10.1016/j.bmcl.2019.126758
22. Saussele S, Haverkamp W, Lang F, et al. Ponatinib in the treatment of chronic myeloid leukemia and Philadelphia chromosome-positive acute leukemia: recommendations of a German Expert Consensus Panel with focus on cardiovascular management. Acta Haematol. 2020;143(3):217–231. doi:10.1159/000501927
23. Ponchio L, Duma L, Oliviero B, Gibelli N, Pedrazzoli P, Robustelli Della Cuna G. Mitomycin C as an alternative to irradiation to inhibit the feeder layer growth in long-term culture assays. Cytotherapy. 2000;2(4):281–286. doi:10.1080/146532400539215
24. Konig H, Holtz M, Modi H, et al. Enhanced BCR-ABL kinase inhibition does not result in increased inhibition of downstream signaling pathways or increased growth suppression in CML progenitors. Leukemia. 2008;22(4):748–755. doi:10.1038/sj.leu.2405086
25. Liu M, Miller CL, Eaves CJ. Human long-term culture initiating cell assay. Methods Mol Biol. 2013;946:241–256.
26. Massimino M, Vigneri P, Stella S, et al. Combined inhibition of Bcl2 and Bcr-Abl1 exercises anti-leukemia activity but does not eradicate the primitive leukemic cells. J Clin Med. 2021;10(23):5606. doi:10.3390/jcm10235606
27. Copland M, Hamilton A, Elrick LJ, et al. Dasatinib (BMS-354825) targets an earlier progenitor population than imatinib in primary CML but does not eliminate the quiescent fraction. Blood. 2006;107(11):4532–4539. doi:10.1182/blood-2005-07-2947
28. Primo D, Flores J, Quijano S, et al. Impact of BCR/ABL gene expression on the proliferative rate of different subpopulations of haematopoietic cells in chronic myeloid leukaemia. Br J Haematol. 2006;135(1):43–51. doi:10.1111/j.1365-2141.2006.06265.x
29. Smith KM, Yacobi R, Van Etten RA. Autoinhibition of Bcr-Abl through its SH3 domain. Mol Cell. 2003;12(1):27–37. doi:10.1016/S1097-2765(03)00274-0
30. Corbin AS, Agarwal A, Loriaux M, Cortes J, Deininger MW, Druker BJ. Human chronic myeloid leukemia stem cells are insensitive to imatinib despite inhibition of BCR-ABL activity. J Clin Invest. 2011;121(1):396–409. doi:10.1172/JCI35721
31. Cilloni D, Saglio G. Molecular pathways: BCR-ABL. Clin Cancer Res. 2012;18(4):930–937. doi:10.1158/1078-0432.CCR-10-1613
32. Udomsakdi C, Eaves CJ, Swolin B, Reid DS, Barnett MJ, Eaves AC. Rapid decline of chronic myeloid leukemic cells in long-term culture due to a defect at the leukemic stem cell level. Proc Natl Acad Sci U S A. 1992;89(13):6192–6196. doi:10.1073/pnas.89.13.6192
33. Eaves AC, Barnett MJ, Ponchio L, Cashman JD, Petzer AL, Eaves CJ. Differences between normal and CML stem cells: potential targets for clinical exploitation. Stem Cells. 1998;16(Suppl 1):77–83; discussion 9. doi:10.1002/stem.5530160809
34. Holyoake T, Jiang X, Eaves C, Eaves A. Isolation of a highly quiescent subpopulation of primitive leukemic cells in chronic myeloid leukemia. Blood. 1999;94(6):2056–2064. doi:10.1182/blood.V94.6.2056
35. Holyoake TL, Jiang X, Jorgensen HG, et al. Primitive quiescent leukemic cells from patients with chronic myeloid leukemia spontaneously initiate factor-independent growth in vitro in association with up-regulation of expression of interleukin-3. Blood. 2001;97(3):720–728. doi:10.1182/blood.V97.3.720
36. Barnes DJ, Schultheis B, Adedeji S, Melo JV. Dose-dependent effects of Bcr-Abl in cell line models of different stages of chronic myeloid leukemia. Oncogene. 2005;24(42):6432–6440. doi:10.1038/sj.onc.1208796
37. Schemionek M, Elling C, Steidl U, et al. BCR-ABL enhances differentiation of long-term repopulating hematopoietic stem cells. Blood. 2010;115(16):3185–3195. doi:10.1182/blood-2009-04-215376
38. Jamieson CH, Ailles LE, Dylla SJ, et al. Granulocyte-macrophage progenitors as candidate leukemic stem cells in blast-crisis CML. N Engl J Med. 2004;351(7):657–667. doi:10.1056/NEJMoa040258
39. Burke BA, Carroll M. BCR-ABL: a multi-faceted promoter of DNA mutation in chronic myelogeneous leukemia. Leukemia. 2010;24(6):1105–1112. doi:10.1038/leu.2010.67
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.