Back to Journals » OncoTargets and Therapy » Volume 14
Gαi1 Promoted Proliferation, Migration and Invasion via Activating the Akt-mTOR/Erk-MAPK Signaling Pathway in Renal Cell Carcinoma
Authors Chen Z ![]() , Zhang Y
, Zhang Y ![]() , Wu X, Zhang J, Xu W, Shen C, Zheng B
, Wu X, Zhang J, Xu W, Shen C, Zheng B
Received 22 December 2020
Accepted for publication 8 April 2021
Published 4 May 2021 Volume 2021:14 Pages 2941—2952
DOI https://doi.org/10.2147/OTT.S298102
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr XuYu Yang
Zhan Chen,1,2,* Yong Zhang,1,2,* Xiang Wu,1 Ji Zhang,1 Wei Xu,1,2 Cheng Shen,1,2 Bing Zheng1
1Department of Urology, The Second Affiliated Hospital of Nantong University, Nantong, People’s Republic of China; 2Medical Research Center, The Second Affiliated Hospital of Nantong University, Nantong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Bing Zheng
Department of Urology, The Second Affiliated Hospital of Nantong University, Nantong, 226001, People’s Republic of China
Tel +86-13813605988
Email [email protected]
Background: Renal cell carcinoma (RCC) accounts for about 2– 3% of all adult malignancies. G protein alpha inhibitory subunit 1 (Gαi1) plays a key role in mediating PI3K-Akt signaling upon activation of receptor tyrosine kinases (RTKs). However, little is known about its expression, regulation and biological function in RCC.
Methods: Gαi1 expression in RCC tissues and cells was detected by quantitative real-time PCR (qRT-PCR), Western blot and immunohistochemistry (IHC). The effect of Gαi1 silence on cell proliferation and apoptosis of 786-O and ACHN cells was detected by CCK-8 assay and flow cytometry. Wound-healing assay and Transwell assays were used to detect the cell invasion in RCC cells. The expression of CDK4, cyclin D1, MMP-2, MMP-9, Bax, Bcl-2, p/t-Akt, p/t-S6 and p/t-Erk was detected by Western blot and qRT-PCR. Furthermore, a nude mouse subcutaneous xenograft model was used to further evaluate the potential effects of Gail in vivo.
Results: In the present study, our data showed that Gαi1 expression was dramatically increased in RCC tissues compared with normal renal tissues. In addition, knocking down the expression of Gαi1 subsequently inhibited proliferation, migration and invasion of RCC cells in vivo and vitro. Furthermore, the expression of CDK4, cyclin D1, MMP-2 and MMP-9 was significantly reduced upon Gαi1 inhibition. Gαi1 positively regulates the activation of the mTOR and Erk pathways.
Conclusion: In conclusion, this study reveals Gαi1 promoted proliferation via activating the Akt-mTOR and Erk-MAPK signaling pathways in RCC, and Gαi1 may be a therapeutic and prognostic target for RCC.
Keywords: renal cell carcinoma, Gαi1, cell proliferation, cell migration, therapeutic targets
Introduction
Renal cell carcinoma (RCC) was the most common type of malignant tumor in kidney that accounts for more than 90% of renal cancer,1,2 among which 75–85% of them are clear cell renal cell carcinoma.3 RCC ranks among the 10 most common cancer types globally that causes about 140,000 deaths every year.4 Despite the continuous progress in medical treatment, the incidence of the disease has increased year by year.5 Patients with localized renal tumors generally have a favorable prognosis, with 5-year OS rates consistently between 60% and 70% after surgery.6 However, due to RCC is not sensitive to chemotherapy, the treatment options are very limited.7 The standard treatment for RCC is surgical resection.7 The prognosis of these patients with advanced disease is extremely poor, and the 5-year overall survival (OS) rate for patients with metastatic disease is less than 10%.8,9 Although immunotherapy, including interleukin-2 and interferon-α, is widely used in the treatment of metastatic renal cell carcinoma (mRCC), its efficacy is still controversial.10–12 Targeted drugs, including vascular endothelial growth factor receptor (VEGFR) inhibitors and rapamycin (mTOR) pathway inhibitors, provide new treatment regiments for mRCC patients. But the effect of these treatments is very limited and some patients suffer serious side effects such as hand–foot skin reactions, hypertension, diarrhea, etc.13,14 Thus, it is essential to identify the molecular mechanism underlying RCC to provide guidance for clinical decision-making and developing novel therapeutic strategies.
The G protein α inhibitory subunit (including Gαi1, Gαi2, and Gαi3) were initially identified by their ability to bind to GPCRs (G protein-coupled receptors) and inhibit adenylate cyclase (AC) activity.15 The traditional view is that G protein α inhibitory subunit (Gαi) routinely transduces G protein-coupled receptor (GPCR) signals, inhibits adenylate cyclase (AC) activity, and reduces the level of intracellular cAMP.16,17 Recent studies have shown that Gαi1/3 are physically bound to ligands that activated RTK (such as EGFR and KGFR), which encodes and activates the adaptor protein Gab1 to mediate downstream Akt-mTOR activation.18–20 Upon stimulation by EGF, Gαi1 can bind epidermal growth factor receptor (EGFR) as an adaptor protein, and promote proliferation and migration through the PI3K-Akt-mTOR pathway in vitro.21,22 In addition, Gαi1 mediates the activation of Akt-mTOR pathway induced by keratinocyte growth factor (KGF) and basic fibroblast growth factor (bFGF), and thus participates in the regulation of proliferation, differentiation, survival and migration of skin keratinocytes.19,20,22 In human gliomas, overexpression of Gαi1 leads to Akt activation and proliferation of glioma cells.23 However, the role of Gαi1 in the progression of RCC remains unclear.
This study aimed to investigate the expression and regulatory mechanisms of Gαi1 in RCC. Our study indicates that Gαi1 is highly expressed in RCC tissues. Our results further showed that suppression of Gαi1 expression significantly inhibited proliferation of RCC cells in vitro and the tumor growth of RCC cells in nude mice. In addition, the decline of CDK4, cyclin D1, MMP-2 and MMP-9 was significant after inhibiting Gαi1 in RCC cells. Furthermore, we found that the Akt-mTOR and Erk-MAPK pathways are downstream pathways of Gai1. Together, our data suggest that Gαi1 may be a potential therapeutic target for RCC patients.
Materials and Methods
Clinical Samples
From 2017 to 2019, altogether 43 paired RCC and adjacent normal tissue samples were collected from patients who were undergoing renal cancer surgery in The Second Affiliated Hospital of Nantong University. The patients did not receive any treatment before surgery. All patients participating in the study were informed and written informed consent was obtained. The study was approved by The Second Affiliated Hospital of Nantong University Ethics Committee, according to the Declaration of Helsinki.
Cell Culture
Human renal cell carcinoma cell lines 786-O, A498, ACHN, Caki1 and Caki2 were purchased from the Shanghai Institutes of Biological Sciences (Shanghai, China). 786-O, Caki1 and Caki2 Cells were cultured in 1640 medium (Gibco) supplemented with 10% fetal bovine serum (Gibco) and A498, ACHN cells were maintained in MEM (Gibco) medium with 10% FBS in a humidified incubator (Thermo Fisher Scientific) air atmosphere containing 5% CO2.
Lentiviral Infection
Gαi1 expression was knocked down using lentiviral vectors. The LV-shGαi1 sequences were as follows: 5′‐AGGATCAAACACATATGAA‐3′ and LV-shNC sequences were as follows: 5′‐TTCTCCCGAACGTGTCACG‐3′. Gαi1 knockdown (named as LV-shGαi1) lentivirus and negative control GV248 vector (termed as LV-shNC) infected the 786-O and ACHN cells, following the reagent manufacturer’s instructions (Genechem, Shanghai, China). Green fluorescence cells were observed with the fluorescence microscope and retroviral production and puromycin stable cell selection has been previously described cell infection efficiency was evaluated. The knockdown efficiency was determined by qRT-PCR and Western blot.
Western Blotting Analysis
Proteins were extracted from RCC patient tissues or cells in ice-cold RIPA buffer containing protease inhibitors and protein concentration was determined by BCA Protein Assay Kit. Western Blot assays were performed by well-established protocols as previously described.27 Image J was used for density analysis to quantify the intensity of the Western blotting band. The primary antibodies used in Western blotting were as follows: Gαi1 (1:1000, Proteintech), β-actin (1:5000, Proteintech), Akt (1:1000, CST), p-Akt (1:1000, CST), Erk (1:1000, CST), p-Erk (1:1000, CST), S6 (1:1000, CST) p-S6 (1:1000, CST), cyclin D1 (1:2000, Proteintech), CDK4 (1:1000, Proteintech), MMP-2 (1:1000, Proteintech), MMP-9 (1:1000, Proteintech), BAX (1:4000, Proteintech), Bcl2 (1:1000, Proteintech). The secondary antibody used in Western blotting was HRP goat anti-rabbit IgG (1:5000, Proteintech) and HRP goat anti-mouse IgG (1:5000, Proteintech).
RNA Isolation and Quantitative Real-Time PCR
Total RNA was isolated from RCC patient tissues or cells lysates using Trizol kit (Qiagen, Valencia, CA, USA). cDNAs were synthesized using a Thermo-script RT kit (Life Technologies, Rockville, MD, USA). Quantitative real-time PCRs were performed in in CFX96™ Real-Time System (Bio-Rad, Hercules, CA, USA), using SYBR PCR reagent (Takara, Shiga, Japan). GADPH is standardized as a gene expression level. The sequences of primers are as follows:
Gαi1-F: GCTCAACCAAATTACATCCCGAC
Gαi1-R: ATCTCTGACCTCCCACATCAAAC
CDK4-F: TGCTGGATGTCATTCACACAGA
CDK4-R: TTGATGAGGGGAAGAGGAATGC
Cyclin D1-F: TTCGTGGCCTCTAAGATGAAGG
Cyclin D1-R: GTTCCACTTGAGCTTGTTCACC
MMP-2-F: AGACCTGGATCGCCAAAGAAGC
MMP-2-R: TTCGTGTACCACTGTACAGGGTC
MMP-9-F: TCGAGAACTTGCAGGCCGCTGT
MMP-9-R: ATGGACCGTGTGCTTTCCTATG
BAX-F: CAGCTGACATGTTTTCTGACGG
BAX-R: AGCCCATGATGGTTCTGATCAG
BCL2-F: ATTGTGGCCTTCTTTGAGTTCG
BCL2-R: TTCAGGTACTCAGTCATCCACAG
GAPDH-F: ACTTGGTATCGTGGAAGGACTC
GAPDH-R: GTAGAGGCAGGGATGATGTTCTG
Immunohistochemistry
Renal cancer and normal tissue were used for immunohistochemistry. After baking for 60 min in a constant temperature oven at 60°C, tissue chips were dewaxed in xylene and hydrated in graded alcohols. Slides were blocked for 5 min, then primary antibody Gαi1 (1:100, Proteintech), Ki67 (1:8000, Proteintech) was added for incubating at 4°C overnight. After washing, secondary antibody HRP Goat Anti-Rabbit IgG (1:200, Proteintech) was added and incubated for 2 h at room temperature. Finally, the tissue specimens were stained with diaminobenzidine and exanimated with microscopic. Specimens were classified into negative, positive, ++ positive, or +++ positive, based on the sum of the staining intensity (varied from weak to strong) and staining extent scores.
CCK-8 Assay
Cell proliferation was measured by CCK-8 assay (CCK-8, Dojindo, Japan). Lentivirus infected 786-O and ACHN cells in exponential growth phase were seeded onto five 96-well plates (3×103 cells/well) in triplicate and cultured for 1, 2, 3, and 4 days. Four hours before absorbance measuring, 10 μL of CCK-8 solution was added. The absorbance was measured at 450 nm with a microplate reader after incubated at 37°C for 2 h.
Colony Formation Assay
Lentivirus infected cells 786-O and ACHN were seeded into 6-well plates (800 cells/well) and cultured in the RPMI-1640 or MEM medium with 20% FBS for 7 days. The colonies were fixed with 4% paraformaldehyde for 30 min. The colonies were then washed with PBS and stained with 0.1% crystal violet. Finally, The cell colonies were photographed.
Wound Healing Assay
786-O and ACHN cells (5×105 cells/well) were seeded on a 6-well plate. The cell layer scratches in each well were incubated with serum-free medium. Take pictures with the microscope every 6 hours, and repeat 3 times to calculate the cell migration rate of each group.
Transwell Assay
Transwell kit (8.0μm pore size polycarbonate filter) with a Matrigel overlay (BD, NJ) was used to evaluate the invasion ability of cells. 786-O cells and ACHN cells (1×105) with 200μL of FBS-free medium was added into the upper chamber, and 600μL of medium with 20% FBS was add into the lower chamber. After incubating for 24 hours at 37°C with 5% CO2, non-invasive cells were removed on the upper surface by a cotton swab. The invaded cells were fixed with 4% paraformaldehyde, stained with 0.1% crystal violet and counted.
Cell Apoptosis Were Detected by Flow Cytometry
To further quantify apoptotic cells, flow cytometry (FCM) analysis was performed according to the manufacturer’s instructions. 786-O and ACHN cells were seeded in a 6-well plate and cultured for 24 hours. After centrifugation (1000×g), the cells returned to normal and were suspended in binding buffer, and stained with 5μL PE Annexin V and 5μL 7-AAD. At least 5×103 cells/sample were collected and analyzed using an EPICS XL flow cytometer (BD LSR Fortessa, USA).
In vivo Xenograft Experiments
Female BALB/c nude mice aged 4–6 weeks were obtained from the Animal Research Center of Nantong University. Approximately 5.0×106 different 786-O cells (786-O-shNC, and 786-O-shGαi1) were subcutaneously injected into the left and right sides of the nude mice armpit. The mice body weight and tumor volume were measured every other week. The tumor volume calculation formula: tumor volume (mm3) = length×width2×0.52. All mice were euthanized at t30 daysafter seeding 786-O cells and tumors removed, weighed, fixed and embedded in IHC. The protocol for animal experiment was approved by the Animal Ethics Committee of Nantong University and carried out in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Statistical Analysis
Statistical analysis between the two groups was analyzed by Student’s t-test and comparisons involving multiple groups were analyzed by two-way ANOVA using GraphPad 5.02, followed by post hoc tests. All data are expressed as the mean ± SD, and p < 0.05 was considered represent statistically significant. All experiments were repeated at least three times.
Results
Gαi1 Expression is Upregulated in RCC Tissues and RCC Cell Lines
To investigate the role of Gαi1 in RCC, the mRNA and protein expression levels were detected by qRT-PCR, Western blot and immunohistochemistry (IHC) in a series of RCC cell lines and 43 pairs of RCC tissue samples (non-metastatic tumors and their adjacent normal tissue specimens). qRT-PCR result suggested that Gαi1 mRNA expression was significantly upregulated in fresh RCC tissues (Figure 1A). Western blotting and IHC analysis showed a significant increase of Gαi1 protein expression in RCC (Figure 1B and E). The expression of Gαi1 in five RCC cell lines examined by qRT-PCR and Western blotting revealed that Gαi1 is highly expressed in RCC cell lines (Figure 1C and D).
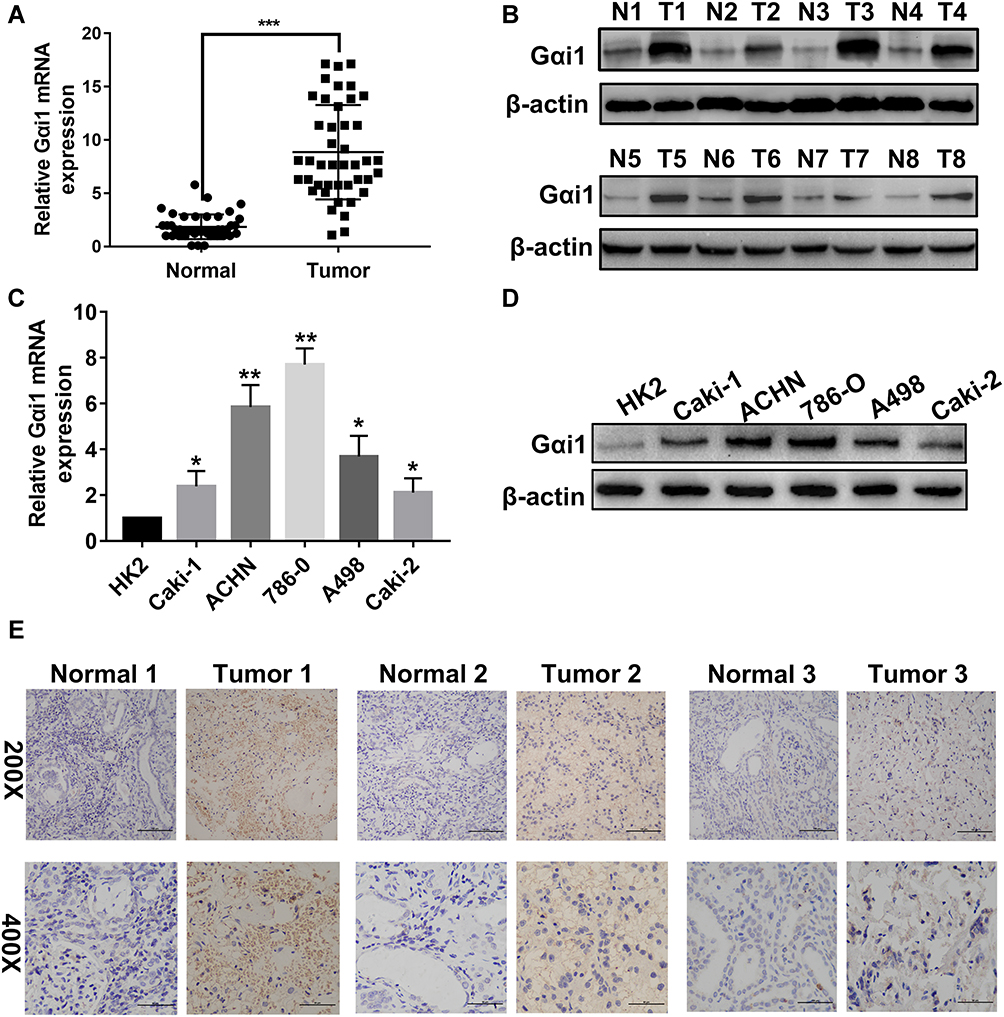 |
Figure 1 Gαi1 expression is upregulated in RCC tissues and RCC cell lines. The expression of Gαi1 mRNA (A) in RCC tumor tissues with different tumor stages and normal tissues was detected by by qRT-PCR. The relative quantification was calculated by the 2−ΔΔCt method and normalized based on GAPDH. Expression level of Gαi1 protein (B) in RCC tissues and adjacent normal tissues. Gαi1 mRNA (C) and protein expression (D) in RCC cell lines and normal epithelium cell of renal tubule HK2. IHC analysis (E) of Gαi1 in RCC and adjacent normal tissues. Results were presented as mean ± SD. *P<0.05, **P<0.01, ***P<0.001. Abbreviations: N, adjacent tissues; T, RCC tissues. |
Gαi1 Knockdown Inhibited RCC Cells Proliferation and Migration in vitro
In order to further investigate the underlying mechanism of Gαi1 in promoting RCC, we knockdown Gαi1 expression in RCC cell line 786-O cells and ACHN cells by lentiviruses-mediated expression of control shRNA (LV-shNC) and Gαi1-specific shRNA (LV-shGαi1) in vitro. The mRNA and protein expression of Gαi1 in 786-O and ACHN cells after virus infection was detected by Western blotting and qRT-PCR, which demonstrates efficient downregulation following infection with LV-shGαi1 (Figure 2A–D).
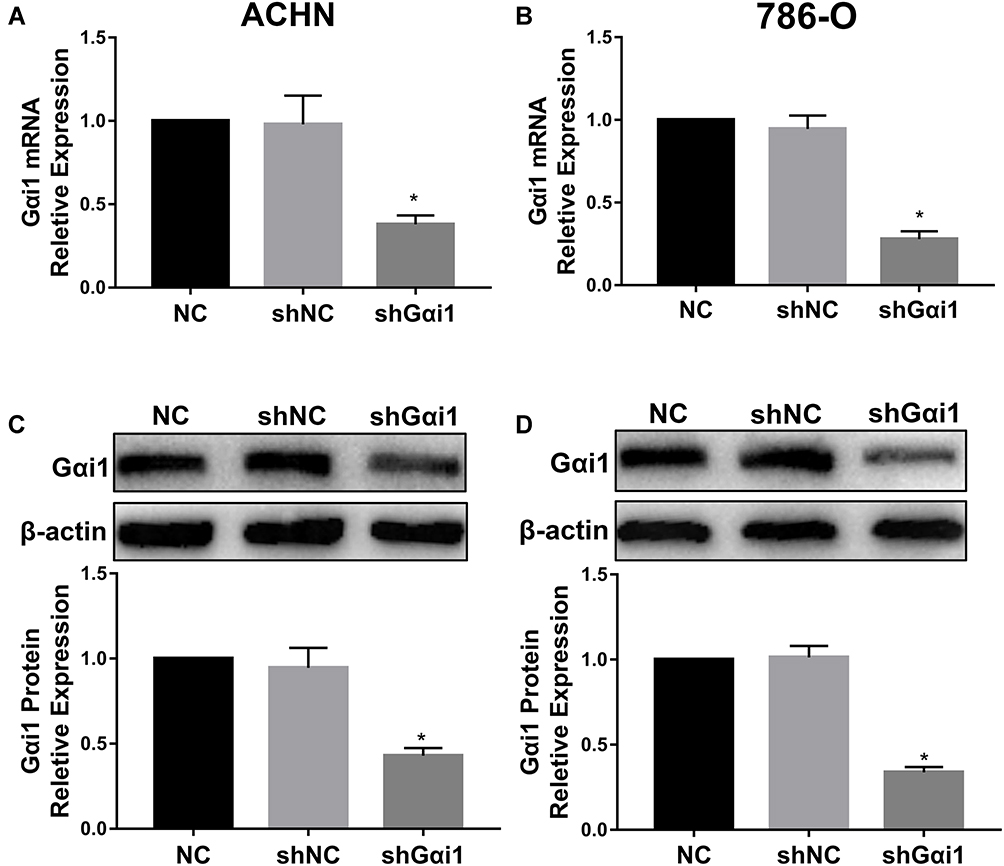 |
Figure 2 Gαi1 expression is knocked down by Gαi1 shRNA. Gαi1 knockdown cell models were constructed through the transfection of shNC or shGαi1. The knockdown efficiencies of Gαi1 mRNA (A and B) in 786-O and ACHN cells were detected by qRT-PCR. Gαi1 protein expression (C and D) level decreased significantly in group of shGαi1 whether in Western blot analysis. Results were presented as mean ± SD. *P<0.05. |
To understand the role of Gαi1 in the proliferation of RCC cells, we first performed a CCK-8 assay and a colony formation assay. CCK-8 assay demonstrated that the reduced proliferation of 786-O and ACHN cells after Gαi1 knocking down when compared to control cells (Figure 3A). Also, results from the colony formation assay indicated that silencing Gαi1 obviously inhibited the cell proliferation, which was demonstrated by the number of 786-O and ACHN cell colonies is significantly reduced (Figure 3B and E). Then, the cell migration of 786-O and ACHN cells was further evaluated (Figure 3C and F). The wound healing assay and transwell assay indicated that Gαi1 knockdown significantly impaired the invasive and migratory capabilities of 786-O and ACHN cells (Figure 3D and G). Furthermore, significant decline of CDK4, cyclin D1, MMP-2 and MMP-9 was observed after inhibiting the function of Gαi1 in 786-O and ACHN cells (Figure 4A–E).
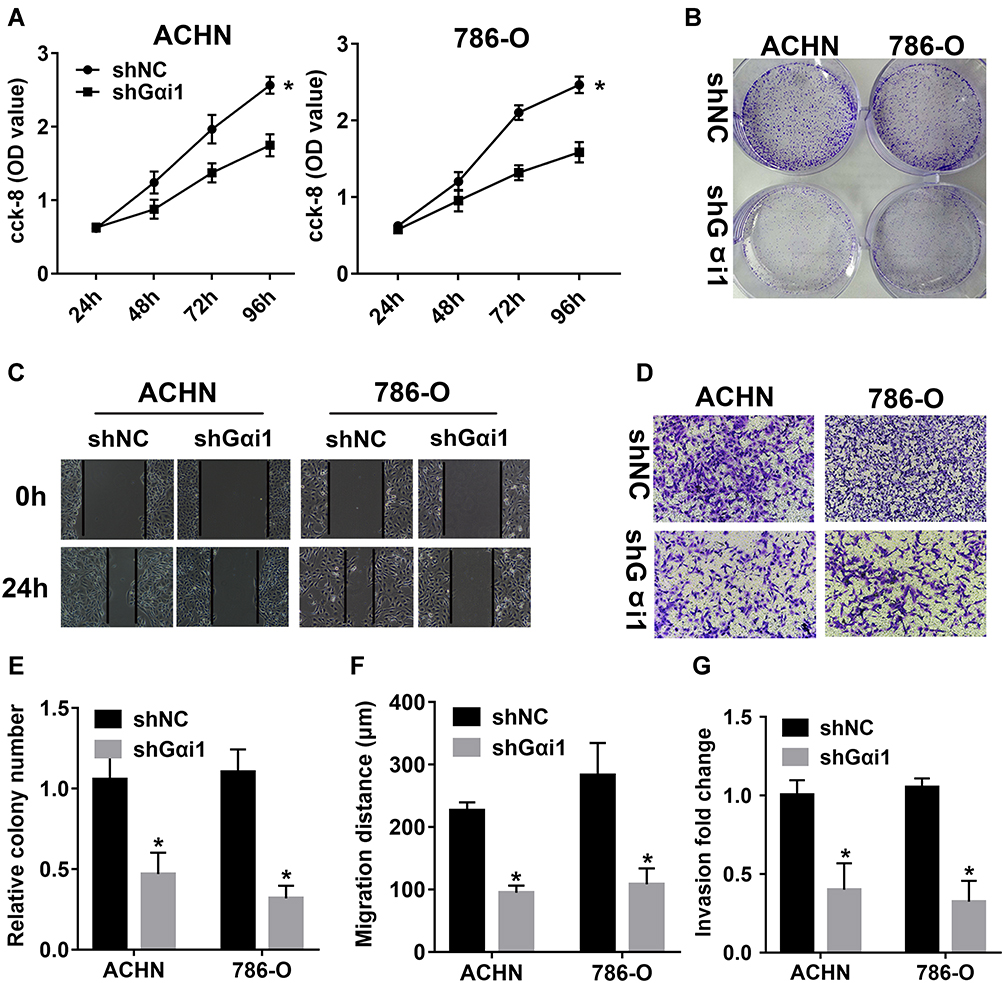 |
Figure 3 Gαi1 knockdown inhibited RCC cells proliferation and migration in vitro. (A) The effects of Gαi1 knockdown on cell proliferation of 786-O and ACHN cells were detected by CCK-8 assay. (B and E) Colony formation assay showed that RCC cell colony formation ability was significantly inhibited in Gαi1 knockdown group. (C and F) Wound-healing was utilized to assess the effects of Gαi1 knockdown on cell migration of RCC cells. (D and G) Transwell assays were used to detect the cell invasion in RCC cells between shNC and shGαi1 group. Results were presented as mean ± SD. *P<0.05. |
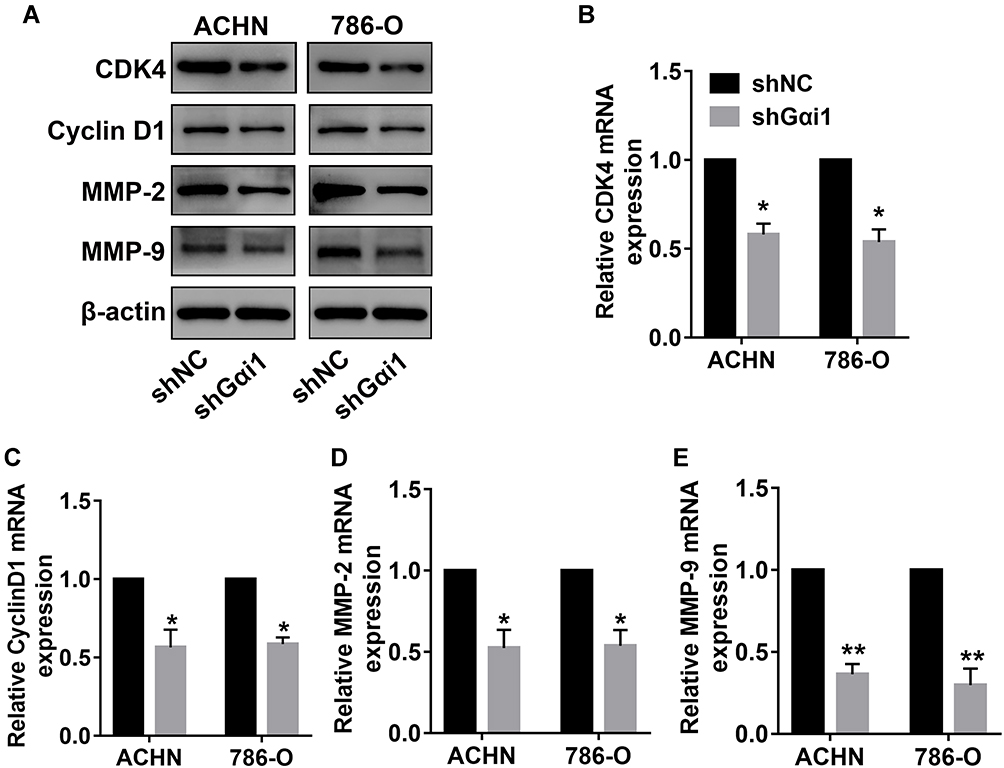 |
Figure 4 Gαi1 knockdown inhibited the expression of CDK4, cyclin D1, MMP-2 and MMP-9 in RCC cells. (A) Western blot analysis of CDK4, cyclin D1, MMP-2 and MMP-9 protein expression in 786-O and ACHN cells with Gαi1 knockdown. (B–E) The mRNA expression of CDK4, yclin D1, MMP-2 and MMP-9 was detected by qRT-PCR in 786-O cells of shNC and shGαi1 groups. Results were presented as mean ± SD. *P<0.05, **P<0.01. |
Gαi1 Knockdown Promoted Cells Apoptosis in vitro
Apoptotic dysfunction caused by dysregulation of apoptosis-related proteins plays an important role in the development of cancer.24 To determine whether Gαi1 influenced RCC cell apoptosis, flow cytometry assay was performed to detect apoptotic cells. We observed that Gαi1 knockdown obviously promoted apoptosis of 786-O and ACHN cells, respectively (Figure 5A and C). Besides, the mRNA expression and protein expression of Bax and Bcl-2 were detected by qRT-PCR and Western blot. The result of qRT-PCR and Western blot analysis showed that the expression of Bax was up-regulated and Bcl-2 was down-regulated in the Gαi1 knockdowning RCC cells (Figure 5B–E). These results indicated that knockdown of Gαi1 induced apoptosis of RCC cells.
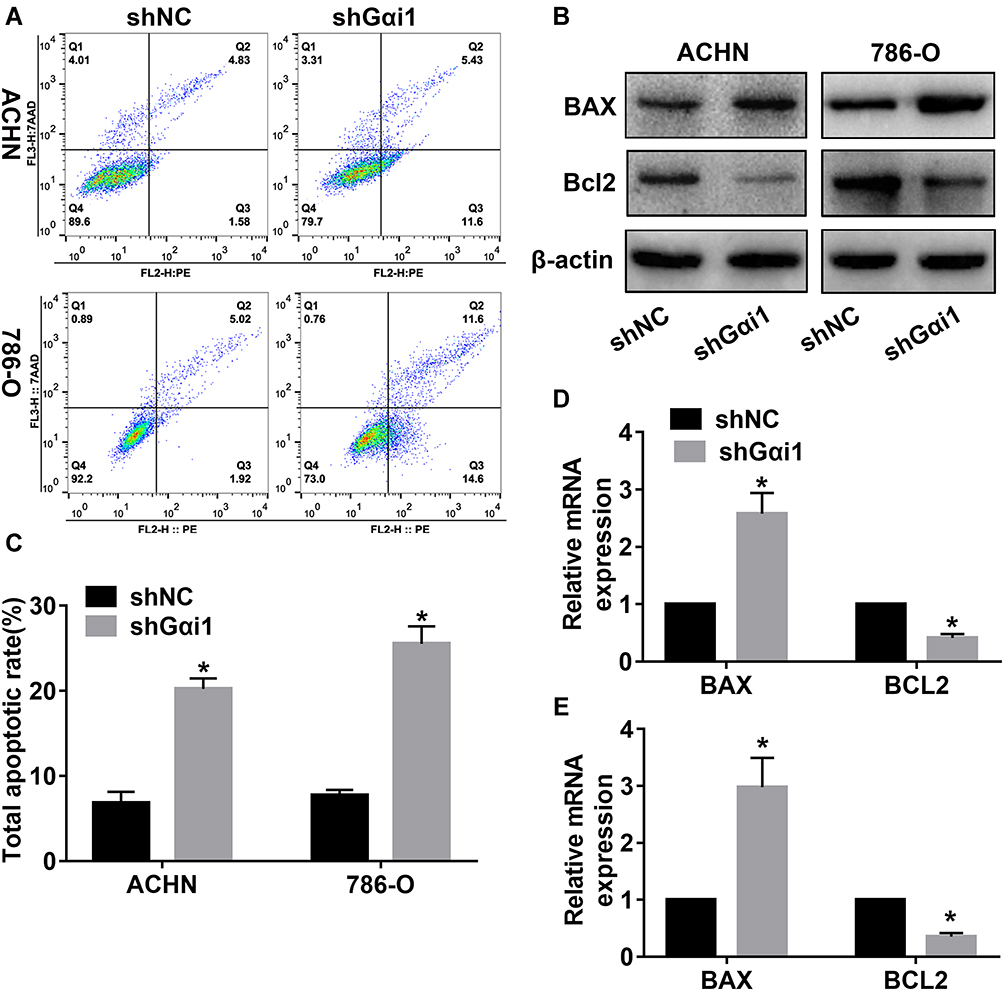 |
Figure 5 Gαi1 knockdown promoted cells apoptosis in vitro. (A and C) Flow cytometry was performed to assess the effects of Gαi1 knockdown on cell apoptosis of 786-O and ACHN cells. Western blot analysis of BAX and Bcl2 protein expression (B) in 786-O and ACHN cells with Gαi1 knockdown. Relative mRNA expression of BAX and Bcl2 was detected by qRT-PCR and normalized based on the shNC group in ACHN (D) and 786-O (E) cells. Results were presented as mean ± SD. *P<0.05. |
Gαi1 Knockdown Inhibited mTOR and Erk Signaling Pathways
Many lines of evidence show that Gαi1 forms a complex with RTKs (FGFR, EGFR and KGFR) and mediates the activation of downstream Gab1-PI3K-Akt signaling pathway. To investigate the efficacy of Gαi1 in the signaling pathways, we used Western blot analysis to examine the activation of these pathways. We found that knockdown of Gαi1 inhibited the ratio of p-Akt/t-Akt, p-S6/t-S6 and p-Erk/t-Erk in both 786-O and ACHN cells (Figure 6A and B). These results indicate that knocking down Gαi1 inhibits the Akt-mTOR and Erk-MAPK signaling pathways.
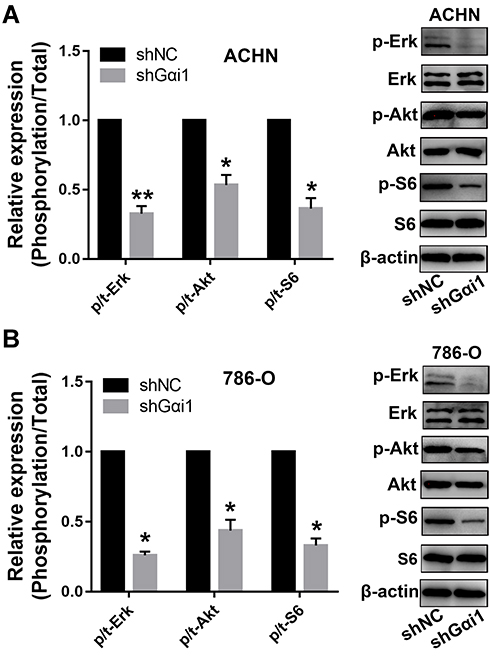 |
Figure 6 Gαi1 knockdown inhibited Akt-mTOR and Erk-MAPK signaling pathways. Western blot was used to analysis of p/t-AKT, p/t-S6 and p/t-ERK protein expression in 786-O and ACHN cells. Knockdown of Gαi1 inhibited Akt-mTOR and Erk-MAPK signaling pathways in 786-O (A) and ACHN (B) cells. Results were presented as mean ± SD. *P<0.05, **P<0.01. |
Gαi1 Knockdown Inhibited Tumor Growth in Nude Mice
According to previous results that knockdown of Gαi1 leaded to impaired proliferation of cancer cells, we further analyzed whether Gαi1 shRNA has an effect on tumor growth in vivo. To establish mice xenograft models, 786-O cells with or without Gαi1 knockdown were injected subcutaneously into nude mice respectively. Our results showed that tumor growth was significantly impaired in the group of Gαi1 knockdown compared with the control group (Figure 7A and B). And knockdown of Gαi1 resulted in a significant reduction in tumor weight as assessed at the end of experiment (Figure 7C and D). We found that Ki-67 staining was stronger in the control group compared to Gαi1-knocking down group (Figure 7E). In summary, the above results indicated that Gαi1 knockdown could inhibit RCC tumor growth in vivo.
 |
Figure 7 Gαi1 knockdown inhibited tumor growth in nude mice. Subcutaneous tumor model of 786-O cells with Gαi1 knockdown. (A) Gαi1 knockdown led to decreased tumor volume. (B) The photos of tumors derived from shNC and shGαi1 cells in nude mice. (C and D) The changes in volume weight were measured at the indicated weeks after mice were transplanted. (E) IHC analysis that Gαi1 knockdown reduced Ki67 protein expression. Results were presented as mean ± SD. *P<0.05. |
Discussion
Renal cancer have no specific presenting symptoms or signs. It has been reported that approximately 30% of RCC patients have a metastatic lesion at the time of initial diagnosis.25 The clinical prognosis of patients with metastatic RCC (mRCC) is extremely poor, with a median survival of only 13 months.26 The Akt/mTOR signaling pathway plays a very important role in the occurrence and development of renal cell carcinoma; therefore, targeting mTOR and block the Akt/mTOR signaling pathway is an potential effective way to treat mRCC.27,28 mTOR inhibitors are mainly targeted at mTORC1, which suppress tumor cells growth upon treatment of a variety of cell growth factors including vascular endothelial growth factor (VEGF) and platelet-derived growth factor (PDGF).29 The related drugs such as temsirolimus and everolimus were approved by the US FDA for the treatment of progressive metastatic RCC.30 However, most RCC patients have a shorter period of effective response to mTORC1 inhibitors, and eventually most patients acquire resistance.30,31 Therefore, there is an urgent need to find new molecular mechanisms to treat RCC and improve the prognosis of patients with metastatic RCC.
The results of this study indicate that Gαi1 may be a new anti-cancer target protein to slow down the growth of RCC cells. First, Gαi1 mRNA and protein are up-regulated in human RCC tissue (compared to surrounding kidney tissue) and RCC cells (compared to human normal renal tubular epithelial cells). Second, Gαi1 can promote RCC cell proliferation, migration and invasion, and inhibit RCC cell apoptosis. Third, the tumor growth of Gαi1 knocking down RCC cells was slower than that of control tumors. Fourth, in RCC cells, Gαi1 can activate the Akt-mTOR and Erk-MAPK signaling pathways.
Gαi protein is essential for EGF to activate the PI3K/Akt/mTORC1 pathway,19 while EGF and EGFR are commonly expressed in most human cancers.32 The activation of Akt and mTOR is important for tumor proliferation, survival, and metastasis. The function of Gαi1 in the progress of the cancer is crucial,33 so Gαi protein is likely to become a potential new target for anti-cancer therapy. The results of immunohistochemistry in this study showed that Gαi1 was highly expressed in RCC tissues, and the up-regulation of Gαi1 in RCC tissues was related to tumor staging, which suggests that Gαi1 plays an important role in the occurrence and development of RCC. Similar studies have shown that Gαi1 is highly expressed in human glioma tissue, and the up-regulation of Gαi1 in glioma tissue is related to tumor grade and Akt activation. This study showed that Gαi1 can promote the proliferation, migration and invasion of 786-O and ACHN cells in vitro. The establishment of subcutaneous xenogeneic tumor formation experiments in nude mice showed that Gαi1 can promote the proliferation of RCC cells.
The Akt/mTOR signaling pathway plays a vital role in the occurrence and development of renal cancer,34 and studies have shown that Gαi protein is essential for EGF to activate the PI3K/Akt/mTORC1 pathway.20 Therefore, this study also explored whether Gαi protein can regulate the proliferation, migration and invasion of RCC cells by activating the mTOR and Erk signaling pathway. Western Blot experiments showed that knockdown of Gαi1 inhibited the ratio of p/t-Akt, p/t-S6 and p/t-Erk in 786-O and ACHN cells.
In conclusion, we found that Gαi is highly expressed in human RCC tissues. And our further research shows that Gαi protein can regulate the proliferation, migration and invasion of RCC cells by activating the Akt-mTOR and Erk-MAPK signaling pathways. Therefore, we conducted a preliminary discussion on the role of Gαi protein in the development of RCC, which provides a new and potential therapeutic target for the treatment of RCC.
Funding
This work was supported by the Natural Science Foundation of Jiangsu province (BE2017682) and Nantong City (GJZ17087), and by the Scientific research project of “333 talent program” (BRA2016199).
Disclosure
The authors declare no conflict of interests.
References
1. Chow WH, Dong LM, Devesa SS. Epidemiology and risk factors for kidney cancer. Nat Rev Urol. 2010;7(5):245–257.
2. Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69(1):7–34. doi:10.3322/caac.21551
3. Moch H, Cubilla AL, Humphrey PA, et al. The 2016 WHO classification of tumours of the urinary system and male genital organs—part a: renal, penile, and testicular tumours. Eur Urol. 2016;70(1):93–105. doi:10.1016/j.eururo.2016.02.029
4. Rhoades SK, Bilen MA. A review of papillary renal cell carcinoma and MET inhibitors. Kidney Cancer. 2019;3(3):151–161. doi:10.3233/KCA-190058
5. Tahbaz R, Schmid M, Merseburger AS. Prevention of kidney cancer incidence and recurrence: lifestyle, medication and nutrition. Curr Opin Urol. 2018;28(1):62–79. doi:10.1097/MOU.0000000000000454
6. van den Heuvel C, van Ewijk A, Zeelen C, et al. Molecular profiling of druggable targets in clear cell renal cell carcinoma through targeted RNA sequencing. Front Oncol. 2019;9:117. doi:10.3389/fonc.2019.00117
7. Qu L, Ding J, Chen C, et al. Exosome-transmitted lncARSR promotes sunitinib resistance in renal cancer by acting as a competing endogenous RNA. Cancer Cell. 2016;29(5):653–668. doi:10.1016/j.ccell.2016.03.004
8. Liu X, Yang Z, Luo X, et al. Calcium-activated nucleotidase 1 silencing inhibits proliferation, migration, and invasion in human clear cell renal cell carcinoma. J Cell Physiol. 2019;234(12):22635–22647. doi:10.1002/jcp.28829
9. Pal SK, Ali SM, Yakirevich E, et al. Characterization of clinical cases of advanced papillary renal cell carcinoma via comprehensive genomic profiling. Eur Urol. 2018;73(1):71–78. doi:10.1016/j.eururo.2017.05.033
10. Atzpodien J, Buer J, Sel S, et al. [Chemoimmunotherapy in the systemic treatment of advanced renal carcinoma]. Urologe A. 1999;38(5):474–478. German. doi:10.1007/s001200050316
11. Buti S, Rovere RK, Donini M, et al. Changes in lymphocyte count induced by repeated cycles with low-dose interleukin-2 and interferon-α in 146 patients with renal cell carcinoma. Tumori J. 2012;98(1):45–52. doi:10.1177/030089161209800106
12. Maroto JP, Del Muro XG, Mellado B, et al. Phase II trial of sequential subcutaneous interleukin-2 plus interferon alpha followed by sorafenib in renal cell carcinoma (RCC). Clin Transl Oncol. 2013;15(9):698–704. doi:10.1007/s12094-012-0991-z
13. Pal SK, Quinn DI. Differentiating mTOR inhibitors in renal cell carcinoma. Cancer Treat Rev. 2013;39(7):709–719. doi:10.1016/j.ctrv.2012.12.015
14. Zarrabi K, Fang C, Wu S. New treatment options for metastatic renal cell carcinoma with prior anti-angiogenesis therapy. J Hematol Oncol. 2017;10(1):38. doi:10.1186/s13045-016-0374-y
15. Downes GB, Gautam N. The G protein subunit gene families. Genomics. 1999;62(3):544–552. doi:10.1006/geno.1999.5992
16. Alessi DR, James SR, Downes CP, et al. Characterization of a 3-phosphoinositide-dependent protein kinase which phosphorylates and activates protein kinase balpha. Curr Biol. 1997;7(4):261–269. doi:10.1016/S0960-9822(06)00122-9
17. Marrari Y, Crouthamel M, Irannejad R, et al. Assembly and trafficking of heterotrimeric G proteins. Biochemistry. 2007;46(26):7665–7677. doi:10.1021/bi700338m
18. Holgado-Madruga M, Emlet DR, Moscatello DK, et al. A Grb2-associated docking protein in EGF- and insulin-receptor signalling. Nature. 1996;379(6565):560–564. doi:10.1038/379560a0
19. Cao C, Huang X, Han Y, et al. Galpha(i1) and Galpha(i3) are required for epidermal growth factor-mediated activation of the Akt-mTORC1 pathway. Sci Signal. 2009;2(68):ra17. doi:10.1126/scisignal.2000118
20. Zhang YM, Zhang Z-Q, Liu -Y-Y, et al. Requirement of Galphai1/3-Gab1 signaling complex for keratinocyte growth factor-induced PI3K-AKT-mTORC1 activation. J Invest Dermatol. 2015;135(1):181–191. doi:10.1038/jid.2014.326
21. Marshall J, Zhou X-Z, Chen G, et al. Antidepression action of BDNF requires and is mimicked by Gαi1/3 expression in the hippocampus. Proc Natl Acad Sci U S A. 2018;115(15):E3549–E3558. doi:10.1073/pnas.1722493115
22. Sun J, Huang W, Yang S-F, et al. Galphai1 and Galphai3 mediate VEGF-induced VEGFR2 endocytosis, signaling and angiogenesis. Theranostics. 2018;8(17):4695–4709. doi:10.7150/thno.26203
23. Liu -Y-Y, Chen M-B, Cheng L, et al. microRNA-200a downregulation in human glioma leads to Galphai1 over-expression, Akt activation, and cell proliferation. Oncogene. 2018;37(21):2890–2902. doi:10.1038/s41388-018-0184-5
24. Mohammad RM, Muqbil I, Lowe L, et al. Broad targeting of resistance to apoptosis in cancer. Semin Cancer Biol. 2015;35(Suppl):S78–S103.
25. Cohen HT, McGovern FJ. Renal-cell carcinoma. N Engl J Med. 2005;353(23):2477–2490. doi:10.1056/NEJMra043172
26. Lalani A-KA, McGregor BA, Albiges L, et al. Systemic treatment of metastatic clear cell renal cell carcinoma in 2018: current paradigms, use of immunotherapy, and future directions. Eur Urol. 2019;75(1):100–110. doi:10.1016/j.eururo.2018.10.010
27. Zheng B, Mao J-H, Qian L, et al. Pre-clinical evaluation of AZD-2014, a novel mTORC1/2 dual inhibitor, against renal cell carcinoma. Cancer Lett. 2015;357(2):468–475. doi:10.1016/j.canlet.2014.11.012
28. Luo J, Luo X, Liu X, et al. DUSP9 suppresses proliferation and migration of clear cell renal cell carcinoma via the mTOR pathway. Onco Targets Ther. 2020;13:1321–1330. doi:10.2147/OTT.S239407
29. Sun SY. mTOR kinase inhibitors as potential cancer therapeutic drugs. Cancer Lett. 2013;340(1):1–8. doi:10.1016/j.canlet.2013.06.017
30. Brodziak A, Sobczuk P, Bartnik E, et al. Drug resistance in papillary RCC: from putative mechanisms to clinical practicalities. Nat Rev Urol. 2019;16(11):655–673. doi:10.1038/s41585-019-0233-z
31. Husseinzadeh HD, Garcia JA. Therapeutic rationale for mTOR inhibition in advanced renal cell carcinoma. Curr Clin Pharmacol. 2011;6(3):214–221. doi:10.2174/157488411797189433
32. Sigismund S, Avanzato D, Lanzetti L. Emerging functions of the EGFR in cancer. Mol Oncol. 2018;12(1):3–20. doi:10.1002/1878-0261.12155
33. Stitzlein L, Rao P, Dudley R. Emerging oral VEGF inhibitors for the treatment of renal cell carcinoma. Expert Opin Investig Drugs. 2019;28(2):121–130. doi:10.1080/13543784.2019.1559296
34. Mossmann D, Park S, Hall MN. mTOR signalling and cellular metabolism are mutual determinants in cancer. Nat Rev Cancer. 2018;18(12):744–757. doi:10.1038/s41568-018-0074-8
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.