Back to Journals » Drug Design, Development and Therapy » Volume 20
Fibroblast Growth Factor 21: Mechanisms, Therapeutic Potential, and Clinical Translation in Metabolic Dysfunction
Authors Rama K, Jahagirdar V, Sanyal AJ ![]()
Received 26 December 2025
Accepted for publication 13 April 2026
Published 28 April 2026 Volume 2026:20 560034
DOI https://doi.org/10.2147/DDDT.S560034
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Mariana Carmen Chifiriuc
Kaanthi Rama,* Vinay Jahagirdar,* Arun J Sanyal
Stravitz-Sanyal Institute for Liver Disease and Metabolic Health, Division of Gastroenterology, Hepatology and Nutrition, Virginia Commonwealth University School of Medicine, Richmond, VA, 23298, USA
*These authors contributed equally to this work
Correspondence: Arun J Sanyal, Stravitz-Sanyal Institute for Liver Disease and Metabolic Health, MCV Box 980342, Richmond, VA, 23298-0342, USA, Tel +1-804 828 6314, Email [email protected]
Abstract: Fibroblast growth factor 21 (FGF21) is an endocrine hepatokine that coordinates systemic energy metabolism by linking hepatic nutrient sensing with adipose and central signalling pathways. Experimental and clinical studies identify FGF21 as a critical modulator of lipid oxidation, insulin sensitivity, and inflammatory balance, processes central to the pathogenesis of metabolic dysfunction-associated steatohepatitis (MASH). Endogenous FGF21 rises in response to fasting, lipotoxicity, and mitochondrial stress, yet this compensatory increase is insufficient in chronic metabolic disease, reflecting target-tissue resistance. Pharmacologic augmentation with engineered FGF21 analogues demonstrates robust reductions in hepatic fat, serum transaminases, and fibrosis biomarkers, along with improvements in triglyceride and adiponectin levels as well as liver histology. In a Phase 2b trial, efruxifermin reversed cirrhosis in 39% of participants. These agents act across the MASH cascade, mitigating lipotoxic injury, inflammation, and stellate-cell activation while favorably modifying cardiometabolic risk. FGF21 therefore represents a unifying therapeutic axis that integrates hepatic and systemic metabolic correction. Ongoing Phase 3 studies will determine whether these biochemical and histologic improvements translate into long-term clinical benefit, positioning FGF21 analogues as cornerstone therapies for metabolic disease. This review aims to synthesize current evidence on the molecular mechanisms and therapeutic potential of FGF21 in metabolic dysfunction-associated steatohepatitis. It highlights emerging clinical data on FGF21 analogues and their role in targeting key pathways of disease progression, with implications for future therapeutic strategies.
Keywords: fibroblast growth factor 21, MASH, MASLD, insulin resistance, hepatic fibrosis, efruxifermin, cardiometabolic disease
Introduction
Metabolic dysfunction-associated steatotic liver disease (MASLD) has emerged as the most prevalent chronic liver condition worldwide, affecting an estimated 38% of the global population.1 Metabolic dysfunction-associated steatohepatitis (MASH), the progressive form of MASLD, is increasingly common and closely parallels the rising burden of obesity and type 2 diabetes. Clinically, MASH is defined by hepatic steatosis, inflammation, and hepatocellular injury, with a subset progressing to advanced fibrosis, cirrhosis, hepatocellular carcinoma, and liver-related mortality, while also conferring substantial cardiovascular risk. Current treatment remains centered on lifestyle intervention, while approved and emerging pharmacologic options such as resmetirom and semaglutide offer important advances but are limited by restricted indications, adverse effects, and incomplete coverage of the metabolic, inflammatory, and fibrotic pathways that drive disease progression. In this context, fibroblast growth factor 21 (FGF21) analogues are attractive because they target multiple components of MASH biology simultaneously, with potential to improve steatosis, inflammation, fibrosis, and broader cardiometabolic risk.
FGF21 is a hepatokine that orchestrates systemic adaptation to nutritional and metabolic stress. First described in 2000, FGF21 has emerged as a central mediator linking hepatic energy metabolism with adipose tissue, skeletal muscle, the pancreas, and the central nervous system.2 Unlike paracrine members of the fibroblast growth factor family, which act locally in development and repair, FGF21 belongs to the endocrine subgroup characterized by poor heparin binding, enabling its secretion into the circulation.3 Its discovery reframed hepatokines as endocrine effectors that maintain whole-body metabolic balance. In rodent models, hepatic FGF21 expression increases during fasting, ketogenic diets, cold exposure, and mitochondrial stress.4 FGF21 acts through a receptor complex comprising fibroblast growth factor receptor 1c (FGFR1c) and the co-receptor β-Klotho (KLB). The presence of KLB dictates tissue sensitivity, explaining why FGF21 primarily targets adipose tissue and select brain regions.5 Pharmacologic analogues of FGF21 have since become leading candidates for the treatment of metabolic dysfunction-associated steatohepatitis (MASH) and related cardiometabolic diseases.
This review aims to provide a comprehensive and mechanistic overview of FGF21 biology and its therapeutic relevance in MASH. It integrates preclinical and clinical evidence to contextualize the role of FGF21 analogues across the metabolic, inflammatory, and fibrotic spectrum of disease. By synthesizing emerging trial data, this review highlights FGF21 as a promising multi-target therapeutic strategy in metabolic liver disease.
The Fibroblast Growth Factor Family and FGF21’s Position
The FGF superfamily includes twenty-two proteins with diverse roles in development, repair, and metabolism.6 Based on their signalling mode, FGFs are divided into three subfamilies: Paracrine FGFs, such as FGF1, 10, 16, 18, 20, and 22, act locally through high-affinity binding to heparan sulphate to regulate growth and angiogenesis. Intracrine FGFs (FGF11, 14) remain within cells and modulate ion channels rather than cell-surface receptors. Endocrine FGFs, FGF19 (rodent FGF15), FGF21, and FGF23 circulate systemically and require Klotho co-receptors for receptor activation.7 FGF21 signals through a heterodimeric receptor complex comprising FGFR isoforms (primarily FGFR1c) and the obligate co-receptor β-Klotho, which confers tissue specificity by restricting signalling to KLB-expressing metabolic organs. This receptor requirement is central to its endocrine mechanism, linking ligand binding to downstream ERK and AKT pathway activation.
FGF21 is structurally distinct because of its weak heparin affinity, a feature that enables endocrine action.8 Its C-terminal domain binds β-Klotho, while the N-terminal domain engages FGFR1c, FGFR2c, and FGFR3c to trigger ERK and AKT signalling. This dual-binding requirement confines FGF21 activity to KLB-expressing tissues, maintaining specificity despite broad FGF-receptor (FGFR) distribution. Importantly, it is distinguished from FGF19 by its inability to bind FGFR4.9
Compared with other members of the FGF family, FGF21 is structurally distinct due to its low affinity for heparan sulphate, which enables endocrine secretion rather than local paracrine signalling. Unlike paracrine FGFs that act at the tissue level, FGF21 circulates systemically and requires the β-Klotho co-receptor in complex with FGFR1c to mediate its effects, conferring tissue specificity. Functionally, FGF21 is uniquely adapted for metabolic regulation, coordinating lipid, glucose, and energy homeostasis, in contrast to other FGFs that primarily regulate development, repair, or mineral metabolism.
FGF21 Signalling and Physiologic Role
Tissue Sources
The liver is the dominant source of circulating FGF21 under normal physiological conditions. Other tissues, including adipose tissue, skeletal muscle, pancreas, and heart, contribute under stress or injury.10 Cold exposure stimulates FGF21 production in brown and white adipose tissue to enhance thermogenesis, while skeletal muscle releases FGF21 as a stress-induced myokine in mitochondrial dysfunction or intense exercise.11,12
FGF21 expression shows marked tissue specificity, with the liver serving as the primary source of circulating FGF21 under basal physiological conditions. In contrast, adipose tissue, skeletal muscle, pancreas, and heart contribute to FGF21 production in a context-dependent manner, particularly during metabolic stress, cold exposure, mitochondrial dysfunction, or injury. Notably, hepatic FGF21 is predominantly endocrine in function, whereas extrahepatic expression often reflects localized autocrine or paracrine signalling, highlighting organ-specific differences in both magnitude and functional role.
Regulation of Expression
FGF21 expression is governed by multiple nutrient-sensing and stress-response pathways that enable the liver to adapt to fluctuating metabolic states. During fasting, hepatic peroxisome proliferator-activated receptor alpha (PPARα) acts as a major transcriptional driver, linking FGF21 expression to enhanced fatty acid oxidation and ketogenesis.13 Under conditions of protein restriction, activation of the GCN2-eIF2α-ATF4 pathway upregulates FGF21, integrating amino acid availability with metabolic reprogramming.14 In contrast, carbohydrate overload induces hepatic FGF21 via carbohydrate response element-binding protein (ChREBP), establishing a feedback mechanism that suppresses simple-sugar preference and moderates glycemic flux.15 Beyond nutrient sensing, mitochondrial and endoplasmic reticulum stress markedly increase FGF21 transcription through ATF4 and CHOP activation, positioning it as a sentinel for cellular stress across diverse physiological contexts.16
FGF21 transcription is highly tissue-specific and regulated by distinct transcription factors depending on metabolic context. As described above, in the liver (hepatocytes), the primary source of circulating FGF21, expression is driven by PPARα and cAMP response element-binding protein H (CREBH) during fasting, carbohydrate-responsive element-binding protein (ChREBP) in response to glucose, and activating transcription factor 4 (ATF4) with C/EBP homologous protein (CHOP) under cellular stress.16,17 In adipose tissue, specificity is governed by specificity protein 1 (Sp1) for basal transcription, while c-Jun N-terminal kinase (JNK) signalling enhances FGF21 production in obesity, and Rev-erb alpha modulates signalling through repression of beta-Klotho (β-Klotho), the FGF21 coreceptor.18 In skeletal muscle, FGF21 expression is minimal at baseline but induced during mitochondrial stress through myogenic differentiation 1 (MyoD) and activating transcription factor 2 (ATF2), with additional regulation via the phosphoinositide 3-kinase (PI3K)/Akt/mechanistic target of rapamycin (mTOR) pathway.12 In contrast, cardiac tissue has minimal intrinsic FGF21 production and largely depends on endocrine FGF21 derived from adipose tissue, particularly during injury. Collectively, these mechanisms highlight how FGF21 integrates nutrient, stress, and tissue-specific signals through context-dependent transcriptional control.19
These combined inputs make FGF21 a dynamic mediator of metabolic resilience, integrating dietary, hormonal, and stress cues into a unified endocrine signal.
Target Tissues and Receptors
FGF21 exerts its biological effects through interaction with FGFR isoforms 1c or 3c, which must heterodimerize with the co-receptor β-Klotho to initiate downstream signalling.5,20 Because β-Klotho expression is tissue-restricted, the physiologic actions of FGF21 are confined mainly to metabolic organs that express both receptor components. Adipose tissue represents the principal peripheral target, where FGF21 enhances insulin sensitivity, promotes lipolysis, and augments energy expenditure.21 In the central nervous system, β-Klotho is highly expressed in the hypothalamus, allowing FGF21 to influence appetite, circadian rhythm, and even alcohol consumption.22 Additional sites of action include the pancreas, where FGF21 modulates insulin secretion, and the heart, where it mitigates oxidative and metabolic stress during injury or overload.23,24 The spatial precision conferred by β-Klotho thus ensures that endocrine FGF21 signalling remains physiologically specific despite widespread FGFR distribution.
While FGFR1c is the predominant receptor mediating the metabolic effects of FGF21 in adipose tissue, FGFR2c also contributes to FGF21 signalling in specific contexts, particularly in the liver and epithelial tissues. In the presence of the co-receptor β-Klotho, FGFR2c can facilitate FGF21-induced metabolic and proliferative signalling, although its role is less well defined compared to FGFR1c. FGFR3c may provide complementary or tissue-specific signalling, highlighting that FGF21 activity is mediated through a spectrum of fibroblast growth factor receptors depending on tissue context.
Physiologic Functions
FGF21 coordinates a comprehensive adaptive response that spans energy, glucose, lipid, and behavioral regulation. In the fasting state, FGF21 promotes hepatic fatty acid oxidation and ketogenesis while supporting gluconeogenic pathways that maintain systemic energy supply.25 By stimulating glucose transporter 1 (GLUT1) expression in adipocytes, it enhances glucose uptake and improves insulin responsiveness, effectively lowering circulating glucose and insulin levels.26 FGF21 also reduces plasma triglyceride concentrations by suppressing hepatic lipogenesis and stimulating lipoprotein lipase-mediated clearance.27 It promotes adiponectin secretion, an adipokine that enhances insulin sensitivity and exerts anti-inflammatory effects.28 Beyond peripheral metabolism, FGF21 acts on the brain to modulate sweet-taste preference and alcohol intake, linking hepatic metabolic signals to behavioral regulation.29 Collectively, these actions establish FGF21 as a key endocrine integrator that aligns hepatic nutrient sensing with whole-body metabolic and neurobehavioral adaptation. Figure 1 shows the triggers for FGF21 release and its target sites.
 |
Figure 1 Triggers, signalling pathways, and tissue-specific effects of fibroblast growth factor 21 (FGF21). FGF21 is induced in response to diverse metabolic and cellular stressors, including fasting, protein restriction, carbohydrate excess, mitochondrial and endoplasmic reticulum stress, cold exposure, intense exercise, hyperglycemia, and cardiac pressure overload, through nutrient- and stress-sensing pathways such as PPARα, GCN2-eIF2α-ATF4, ChREBP, PERK, and ATF4/CHOP. Once secreted, primarily from the liver, FGF21 acts via FGFR1c/β-Klotho signalling on multiple target tissues. In the liver, it enhances fatty acid oxidation, suppresses SREBP-1c-mediated lipogenesis, and promotes ketogenesis. In adipose tissue, it increases insulin-independent glucose uptake, lipolysis, and adiponectin secretion. Systemically, these effects improve insulin sensitivity. In the pancreas, FGF21 reduces glucotoxic stress and modulates insulin secretion, while in the central nervous system it suppresses sweet-taste preference, alters circadian feeding behaviour, and increases sympathetic outflow to brown adipose tissue thermogenesis. In the heart, FGF21 improves fatty acid oxidation efficiency and limits oxidative stress and hypertrophic signalling. Together, these coordinated actions position FGF21 as a central endocrine regulator of metabolic homeostasis across organs. (Upward arrow: increases, downward arrow: decreases). |
FGF21 in Obesity, Insulin Resistance and Type 2 Diabetes
FGF21 plays a central role in the metabolic adaptations associated with obesity, insulin resistance, and type 2 diabetes mellitus. It is produced primarily by the liver in response to metabolic stress and acts through FGFR1c and β-Klotho in adipose tissue and the central nervous system to regulate nutrient handling and energy balance. The rise in circulating FGF21 seen in obesity and diabetes reflects a compensatory signal that becomes inadequate as receptor expression and downstream responsiveness decline.30
Obesity
FGF21 influences body weight through effects on adipose tissue, the central nervous system, and whole-body energy expenditure. In white adipose tissue, it enhances insulin-independent glucose uptake, increases fatty acid oxidation, and promotes the release of adiponectin, which improves systemic insulin sensitivity.31 In brown and beige adipose depots, FGF21 stimulates thermogenesis and increases energy expenditure through activation of UCP1 and mitochondrial biogenesis.32 Central actions reduce simple-sugar intake and modulate reward pathways, lowering preference for sweet foods.33 These mechanisms collectively oppose the metabolic consequences of caloric excess.
In diet-induced obese rodents and non-human primates, FGF21 administration produces consistent reductions in body weight, fat mass, and circulating triglycerides.27 Treated animals exhibit higher energy expenditure, increased thermogenesis, and improved adipose tissue insulin signalling. Acute weight reduction often depends on adipose tissue, but chronic effects on energy balance occur even when adipose-mediated pathways are impaired, suggesting both adipose-dependent and independent mechanisms.34 Across models, improvements in hepatic steatosis accompany weight loss, indicating a systemic correction of lipid metabolism.
Human studies show that FGF21 analogues such as LY2405319, pegbelfermin, efruxifermin, and pegozafermin are safe and well tolerated. However, weight loss in obesity trials has been modest and inconsistent. In early trials, treated patients experienced small reductions in body weight despite large improvements in triglycerides, HDL cholesterol, and adiponectin.35–37 Efruxifermin and BFKB8488A showed mild reductions in body weight, but changes rarely matched preclinical observations.38,39 This contrast highlights limitations in translating FGF21-mediated weight loss from animals to humans.
Insulin Resistance
FGF21 enhances insulin sensitivity by improving glucose uptake in adipose tissue through induction of GLUT1, reducing hepatic glucose output, and suppressing de novo lipogenesis.26 It shifts cellular metabolism toward fatty acid oxidation and lowers circulating non-esterified fatty acids, which reduces hepatic substrate burden. The FGF21 adiponectin axis further amplifies systemic insulin sensitization. These molecular actions position FGF21 as a key counter-regulatory hormone in states of metabolic overload.
In rodent models of insulin resistance, FGF21 lowers blood glucose, enhances insulin sensitivity, and improves glucose tolerance independent of insulin secretion.40 These improvements occur rapidly and are tightly linked to β-Klotho expression in adipose tissue. In FGF21-deficient mice, insulin resistance worsens, hepatic steatosis progresses, and inflammatory signalling rises, confirming FGF21’s endocrine role in metabolic homeostasis.41
In insulin-resistant humans, FGF21 analogues improve adiponectin levels, triglycerides, HDL cholesterol, and hepatic fat content.42 Yet glycemic endpoints have shown limited improvement. Pegbelfermin and BFKB8488A improved insulin sensitivity indices but did not significantly reduce fasting glucose or HbA1c in most patients with insulin resistance.37,39 These data underscore a discrepancy between molecular potential and clinical metabolic outcomes.
Type 2 Diabetes Mellitus
FGF21 reduces hepatic gluconeogenesis, increases peripheral glucose uptake, and enhances fatty acid oxidation. In models of diabetes, it decreases pancreatic stress, improves β-cell survival, and reduces glucotoxicity.43 These actions support the physiological hypothesis that FGF21 should have antidiabetic potential.
FGF21 markedly lowers blood glucose in diabetic rodents and non-human primates. It improves glucose tolerance and reduces HbA1c in animals with marked insulin resistance. In these models, both acute and chronic glucose-lowering effects are reproducible and robust.40,44 These findings drove initial enthusiasm for the translation of FGF21 analogues into antidiabetic therapies.
Despite strong preclinical efficacy, clinical trials in type 2 diabetes have not met primary glycemic endpoints. LY2405319 improved triglycerides and HDL cholesterol but produced only small reductions in fasting glucose and HbA1c.45 Pegbelfermin, efruxifermin, and BFKB8488A produced similar metabolic improvements but modest glycemic benefit.37,39,46 The failure to replicate preclinical glucose-lowering effects likely reflects several factors:
- FGF21 resistance: Obesity and diabetes are associated with high endogenous FGF21 levels and reduced β-Klotho expression, limiting target-tissue sensitivity.47
- Pharmacokinetics: Even optimized analogues may not fully mimic physiological pulsatility or tissue distribution patterns.
- Interspecies differences: Human adipose and CNS responses differ significantly from rodents, especially in thermogenesis.
- Complexity of metabolic disease: Redundant hormonal pathways in humans may blunt the antidiabetic effect of isolated FGF21 signalling.
Together, these limitations explain why FGF21 analogues show strong effects on lipids, adiponectin, and hepatic fat but only modest glycemic responses in people with obesity and type 2 diabetes.
Effects on Systemic Metabo-Inflammation
FGF21 plays an important role in regulating systemic metabolic inflammation, or metaflammation, which is a hallmark of obesity, insulin resistance, and type 2 diabetes. Chronic nutrient excess drives activation of inflammatory pathways in adipose tissue, liver, and immune cells. FGF21 counters these processes by improving metabolic homeostasis and by exerting direct anti-inflammatory actions on multiple cell types.
FGF21 reduces inflammatory signalling in adipocytes, preadipocytes, and macrophages. It lowers expression of key pro-inflammatory cytokines, including TNF-α, IL-1β, and IL-6, while increasing anti-inflammatory cytokines such as IL-10. These effects arise through suppression of the NF-κB pathway and activation of Nrf2-dependent antioxidant responses.48 Experimental models of obesity and type 2 diabetes show that FGF21 administration reduces tissue inflammation, decreases circulating inflammatory markers, and improves glucose metabolism as inflammatory tone declines.49
Distinct signalling pathways mediate these anti-inflammatory effects in different tissues. In adipose tissue, FGF21 signals through FGFR substrate 2 and ERK1/2 to reduce cytokine expression and improve adipocyte function. In macrophages, FGF21 modulates oxidative stress responses through activation of Nrf2.48 Together, these pathways attenuate local tissue inflammation and reduce systemic metabolic stress.
In metabolic syndrome and fatty liver disease, circulating FGF21 levels correlate with markers of inflammation such as IL-6, TNF-α, and hs-CRP.50,51 Impaired FGF21 responses are associated with exaggerated postprandial inflammation, suggesting that endogenous FGF21 normally acts as a brake on inflammatory activation during nutrient excess.52 In diet-induced obesity, loss of FGF21 signalling worsens adipose tissue inflammation and hepatic injury, whereas endogenous FGF21 protects against progression of metaflammation.49 In contrast, pharmacologic FGF21 analogues in clinical trials have shown variable anti-inflammatory effects, and are discussed in later sections.
Overall, FGF21 functions as an endocrine regulator of systemic metaflammation. By decreasing inflammatory cytokine production, enhancing antioxidant defence, and alleviating metabolic stress, FGF21 interrupts the cycle that links nutrient overload with chronic inflammation. These actions complement its metabolic effects and contribute to its therapeutic potential in obesity, insulin resistance, type 2 diabetes, and metabolic liver disease.
Rationale for the Use of FGF21 in MASH
Over the course of the last two decades, the core elements of the pathogenic underpinning of MASH have been established.53 MASH progression is driven by a tightly interconnected cascade linking lipid accumulation, oxidative stress, inflammation, and fibrosis. Hepatic lipid overload, driven by increased de novo lipogenesis (via sterol regulatory element-binding protein 1c {SREBP-1c}) and impaired β-oxidation, leads to lipotoxicity with accumulation of toxic lipid species such as free fatty acids and ceramides. This promotes mitochondrial dysfunction and generation of reactive oxygen species (ROS), activating oxidative stress pathways including c-Jun N-terminal kinase (JNK) and nuclear factor erythroid 2-related factor 2 (Nrf2). Oxidative stress in turn triggers hepatocyte injury and release of damage-associated molecular patterns (DAMPs), which activate Kupffer cells and recruit inflammatory cells through nuclear factor-kappa B (NF-κB) signalling and inflammasome pathways (eg., NLRP3).
This inflammatory milieu amplifies hepatocellular injury and perpetuates cytokine release, including tumor necrosis factor alpha (TNF-α), interleukin-1 beta (IL-1β), and interleukin-6 (IL-6), creating a feed-forward loop between oxidative stress and inflammation. Chronic inflammation then activates hepatic stellate cells through transforming growth factor beta (TGF-β) and platelet-derived growth factor (PDGF) signalling, driving extracellular matrix deposition and fibrogenesis. Importantly, these processes are not linear but bidirectional, as fibrosis further exacerbates hypoxia and oxidative stress, reinforcing disease progression.
FGF21 integrates into this pathway as a stress-responsive hepatokine that counteracts multiple nodes of injury. It enhances fatty acid oxidation, reduces lipogenesis, and improves mitochondrial function, thereby limiting lipotoxicity and oxidative stress, while simultaneously suppressing NF-κB-mediated inflammation and promoting anti-inflammatory signalling. These combined metabolic and anti-inflammatory effects position FGF21 analogues as promising therapeutic agents to interrupt disease progression and promote fibrosis regression in MASH.
Figure 2 shows the effects of FGF21 in MASH.
 |
Figure 2 Hepatocyte stress-induced FGF21 signalling and downstream metabolic, inflammatory, and antifibrotic effects. Hepatocellular lipotoxicity, oxidative stress, and endoplasmic reticulum stress induce FGF21 expression and secretion as an adaptive response. Circulating FGF21 signals through the FGFR1/β-Klotho receptor complex to coordinate protective actions across metabolic and immune pathways. In the liver, FGF21 reduces hepatic fat content and serum transaminases. It suppresses inflammatory signalling by lowering TNF-α, IL-6, and IL-1β, promoting IL-10 production, activating Nrf2, and favouring anti-inflammatory macrophage polarization. Metabolic effects include inhibition of SREBP-1c-mediated de novo lipogenesis, activation of PPARα-driven fatty acid oxidation, increased adiponectin, improved insulin sensitivity, higher HDL cholesterol, and reduced triglycerides. FGF21 also exerts antifibrotic effects by limiting hepatic stellate cell activation, decreasing expression of α-SMA and COL1A1, and improving non-invasive fibrosis markers such as PRO-C3 and ELF score. Collectively, these integrated actions position FGF21 as a central stress-response hepatokine that counteracts the key drivers of MASH progression. (Upward arrow: increases, downward arrow: decreases). |
As we will explore below, the emerging consensus is that FGF21 analogues consistently improve hepatic steatosis, lipid profiles, and insulin sensitivity, with several agents demonstrating clinically meaningful signals for MASH resolution and fibrosis improvement. However, variability in antifibrotic efficacy across trials and agents highlights ongoing uncertainty, particularly in advanced fibrosis and cirrhosis, where results have been more modest or inconsistent. Clinically, reductions in liver fat and improvements in non-invasive biomarkers are reproducible, but translation to hard histologic and long-term outcomes remains an area of active investigation.
Strengths of FGF21 analogues include their multimodal mechanism targeting lipid metabolism, inflammation, and fibrosis simultaneously, along with generally favourable safety profiles. Limitations include potential FGF21 resistance, heterogeneity in receptor engagement and pharmacokinetics across agents, and incomplete durability data. Overall, while FGF21-based therapies represent a promising class, further phase 3 data are needed to define their long-term clinical benefit, optimal patient selection, and positioning relative to other metabolic and antifibrotic therapies.
Effects on Hepatic Metabolism
FGF21 regulates key aspects of hepatic lipid and glucose metabolism. It increases fatty acid oxidation, suppresses de novo lipogenesis, and reduces hepatic triglyceride accumulation.25 These actions are mediated through PPARα activation, enhanced mitochondrial β-oxidation, and inhibition of SREBP-1 driven lipogenesis.54 In models of MASLD, FGF21 reduces hepatic steatosis, improves insulin sensitivity, and restores metabolic flexibility.55
FGF21 analogues such as pegbelfermin, efruxifermin, and pegozafermin produce substantial reductions in liver fat in clinical trials.36,38,56 Decreases in ALT, AST, triglycerides, and PRO-C3 accompany these improvements, indicating both metabolic and structural benefit. Through these coordinated actions, FGF21 addresses the metabolic overload that initiates and perpetuates MASLD.
Effects on Oxidative Stress, Mitochondrial Health, and ER Stress
Lipotoxicity and mitochondrial dysfunction are central drivers of MASLD progression.57 FGF21 functions as a stress-responsive hepatokine that counters these processes. It activates Nrf2-dependent antioxidant pathways, which reduce reactive oxygen species and enhance cellular resilience.58 FGF21 improves mitochondrial capacity, supports mitochondrial biogenesis, and restores proteostasis in hepatocytes exposed to lipid excess.
FGF21 also reduces ER stress by improving protein folding capacity and maintaining cellular homeostasis during nutrient overload.59 In preclinical studies, it protects hepatocytes from lipotoxicity-induced apoptosis and prevents progression from simple steatosis to steatohepatitis.60 These actions reduce cellular injury signals that fuel inflammation and fibrosis in MASLD.
Effects on Innate Immune Activation
Innate immune activation amplifies hepatocellular injury in MASLD.61 FGF21 suppresses macrophage-mediated inflammation through inhibition of NF-κB and activation of Nrf2.48 It reduces production of pro-inflammatory cytokines such as TNF-α, IL-1β, IL-6, and IFN-γ while increasing anti-inflammatory IL-10.62 FGF21 acts directly on macrophages and indirectly via adipose tissue, where it reduces inflammatory cytokine release from adipocytes and preadipocytes.49
FGF21 also modulates neutrophil and monocyte function, enhancing phagocytosis and oxidative burst capacity.63 Despite this, the overall effect remains anti-inflammatory in the setting of metabolic disease. In MASLD, circulating FGF21 levels correlate with inflammatory markers, and impaired FGF21 signalling is associated with exaggerated postprandial inflammation. Administration of FGF21 analogues improves hepatic inflammatory markers in clinical studies, whereas systemic anti-inflammatory effects were primarily demonstrated in preclinical models.36,48,64
Effects on Fibrosis
Fibrosis is the major determinant of prognosis in MASLD.65 FGF21 has consistent antifibrotic actions across multiple models. It reduces stellate-cell activation, decreases collagen deposition, and lowers expression of fibrogenic genes such as α-SMA and collagen I. These effects arise from direct antagonism of myofibroblast differentiation, inhibition of TGF-β1/Smad signalling cascades, induction of apoptosis in activated myofibroblasts, and regulation of cytoglobin expression in hepatic stellate cells.66,67
FGF21 analogues have shown meaningful antifibrotic activity in humans. In clinical trials of pegozafermin and efruxifermin, fibrosis biomarkers such as PRO-C3, ELF score, and liver stiffness decline during treatment.64,68 In the HARMONY and BALANCE trials, some patients demonstrated histologic improvement in fibrosis stage, indicating a direct disease-modifying effect. The combination of metabolic correction, anti-inflammatory activity, and stellate-cell suppression positions FGF21 as a strong antifibrotic candidate in MASLD.
Emerging Data on Regeneration
FGF21 has shown regenerative effects in peripheral nerve injury by reducing oxidative damage and regulating autophagy, suggesting broader roles in tissue repair.69 FGF21 accelerates cutaneous wound healing by promoting keratinocyte migration and differentiation via SIRT1-dependent autophagy.70 In diabetic wound models, FGF21 treatment restored endothelial cell proliferation, migration, and tube-forming ability under hyperglycemic conditions while reducing inflammatory responses and mitochondrial damage.71
Although limited, emerging data suggest that FGF21 may support hepatic regeneration. In animal models, FGF21 improves mitochondrial health and reduces oxidative and ER stress, conditions that favour hepatocyte survival and recovery.72 In zebrafish models of targeted liver cell ablation, FGF21 increased the regeneration area of damaged liver and improved survival rates by activating AMPK-mTOR-mediated autophagy while reducing apoptosis and oxidative stress.73 These findings complement the clinical data on fibrosis reversal in MASH discussed previously, suggesting FGF21 supports both hepatocyte survival during acute injury and structural remodelling in chronic disease. These findings remain preliminary but raise the possibility that FGF21 may contribute to hepatic regeneration in addition to its metabolic and antifibrotic actions.
Types of FGF21 Available for Use as Drugs and Differences Across FGF21 Analogues
Native FGF21 has a short plasma half-life of less than two hours, rapid renal clearance, and susceptibility to proteolytic degradation. These properties limit its direct clinical use. To overcome these limitations, several engineered analogues have been developed to prolong half-life, improve receptor affinity, and optimize tissue distribution. Modifications include PEGylation, Fc-fusion, and amino-acid substitutions that resist cleavage by dipeptidyl peptidase IV.74 Each class of analogue differs in pharmacokinetics, receptor selectivity, and clinical performance.
Table 1 summarises the various FGF21 agents available for use as drugs.
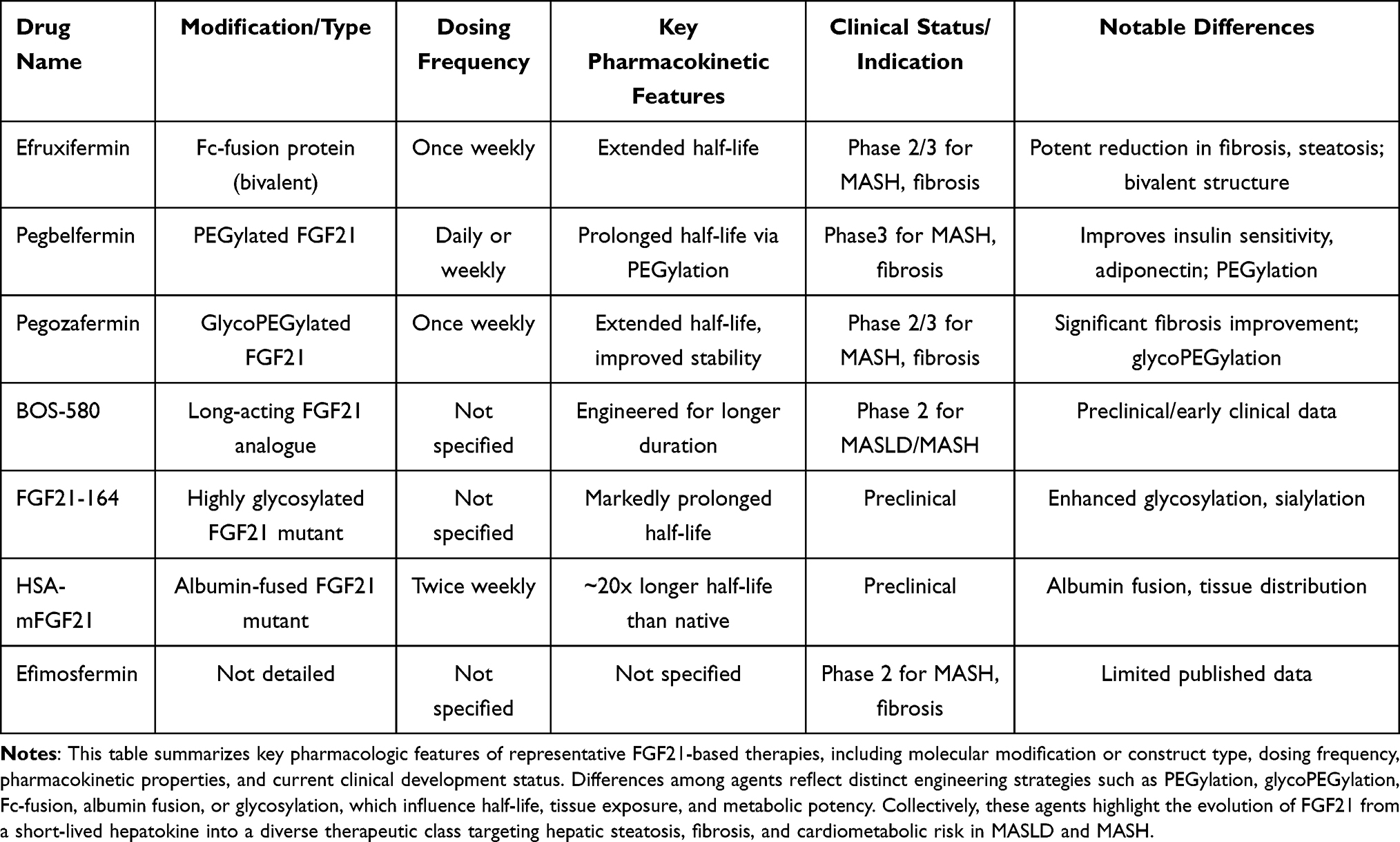 |
Table 1 Comparative Characteristics of FGF21 Analogues in Development for MASLD/MASH |
Early Studies and Clinical Development of FGF21 Analogues
Pegbelfermin (BMS-986036)
Pegbelfermin is a PEGylated recombinant FGF21 analogue designed for once-daily or once-weekly dosing. It was the first FGF21 analogue to show clear metabolic and hepatic efficacy in humans, establishing proof of concept for this therapeutic class.75
In a randomized phase 2a, 16-week, double-blind, placebo-controlled trial in patients with biopsy-confirmed NASH (F1-F3), pegbelfermin achieved significant reductions in absolute hepatic fat fraction compared with placebo.36 More than half of patients in pegbelfermin groups achieved ≥30% relative reduction in hepatic fat fraction, a threshold associated with histological response. Significant decreases in PRO-C3 and increased adiponectin levels were seen in pegbelfermin groups compared to placebo. Improvements were also observed in lipid profiles and markers of liver injury.
In the phase 2b FALCON-1 and FALCON-2 trials in biopsy-proven NASH, once-weekly pegbelfermin for 24–48 weeks produced approximately 30–40% relative reductions in hepatic fat fraction, improvements in ALT and AST, and decreases in fibrosis markers such as PRO-C3 and ELF.76,77 Adverse events were mainly mild gastrointestinal symptoms and transient injection-site reactions, indicating a favourable safety profile. However, histologic NASH resolution and fibrosis improvement endpoints were achieved in only a modest proportion of patients.
Pegbelfermin has not advanced into phase 3 development for MASH. The limited histologic response in FALCON, coupled with the emergence of more potent and durable FGF21 analogues, led to strategic discontinuation of further late-phase trials in favor of next-generation agents.
Efruxifermin (AKR-001)
Efruxifermin is an Fc-fusion FGF21 analogue with a terminal half-life of about 3–4 days, enabling once-weekly dosing. It provides balanced FGFR1c, FGFR2c, and FGFR3c activity while remaining β-Klotho dependent, and currently represents the most advanced FGF21 agent in development.78
Early 2a studies in patients with NASH and metabolic risk factors showed that efruxifermin rapidly reduced liver fat, improved triglycerides and HDL cholesterol, and increased adiponectin.38 The BALANCED phase 2a trial demonstrated that efruxifermin significantly reduced hepatic fat fraction in patients with biopsy proven NASH and F1-F3 fibrosis, by up to 13% vs 0.3% in placebo. Beyond hepatic steatosis, efruxifermin improved multiple metabolic and hepatic parameters, including increasing adiponectin levels and improved lipid profiles compared to placebo. These trials demonstrated good tolerability, with mostly mild gastrointestinal events, and established dose ranges suitable for longer-term histologic studies.
In the phase 2b HARMONY trial, weekly efruxifermin for 24 weeks in patients with NASH and fibrosis stage F2-F3 resulted in NASH resolution without fibrosis worsening in up to 41% of participants, and at least one-stage fibrosis improvement in 39%.46,64 Liver fat reduced by about 60–70% on MRI-PDFF, and fibrosis markers such as PRO-C3 and TIMP-1 improved. Liver fat normalised (≤5%) in 30–50% of patients on efruxifermin, compared to 2% with placebo. The efficacy appeared preserved in patients taking GLP-1 receptor agonists and was unrelated to baseline statin use.
The SYMMETRY phase 2b trial in compensated NASH cirrhosis confirmed beneficial effects on liver stiffness, collagen turnover, and metabolic parameters, with a consistent safety profile.79 The primary outcome was reduction of at least one stage of fibrosis without worsening MASH at week 36, and was not achieved. However, at week 96, efruxifermin demonstrated a clinically meaningful benefit, with 29% of patients achieving fibrosis reduction without worsening MASH compared to 11% with placebo. Multiple secondary endpoints, including reductions in hepatic fat fraction, liver stiffness by MRE and biomarkers of liver injury and fibrosis were noted in the treatment group. These findings positioned efruxifermin as a leading candidate among FGF21 analogues.
Efruxifermin is now being evaluated in the SYNCHRONY phase 3 program, including a large randomized, placebo-controlled trial in non-cirrhotic MASH with fibrosis stage F2-F3 (NCT06215716). This study will assess NASH resolution and fibrosis regression over long-term treatment and will provide definitive data on efficacy, safety, and durability. Additional phase 3 trials in compensated cirrhosis due to MASH are also underway (NCT06528314).
Pegozafermin (BIO89-100)
Pegozafermin is a glyco-PEGylated FGF21 analogue with site-specific modifications that extend its half-life to about 90 hours. It combines potent FGFR1c/β-Klotho activation with infrequent dosing and a strong triglyceride-lowering profile.80
Multiple-ascending-dose and short-term studies in patients with NASH or dyslipidemia showed that pegozafermin substantially reduced triglycerides, increased HDL cholesterol, and lowered hepatic fat content, with good tolerability and low immunogenicity.56,80 These results provided early evidence of dual hepatic and cardiovascular benefit.
The ENLIVEN phase 2b trial demonstrated that pegozafermin significantly improved fibrosis (27% of patients receiving 44 mg every 2 weeks vs 7% with placebo) and NASH resolution (37% vs 2% in placebo) in patients with biopsy-confirmed NASH and F2-F3 fibrosis.68 Substantial reductions were noted in liver fat content measured by MRI-PDFF, with improvements in ELF scores, liver stiffness and liver volume. In the ENTRIGUE trial, pegozafermin reduced triglycerides by 50–60% and improved HDL cholesterol in patients with severe hypertriglyceridemia, supporting its development both for MASH and for atherogenic dyslipidemia.81
Phase 3 development of pegozafermin includes the ENLIGHTEN-Fibrosis trial (NCT06318169), a randomized, placebo-controlled study in adults with MASH and fibrosis stage F2-F3. This trial evaluates whether pegozafermin can achieve fibrosis improvement without worsening MASH or MASH resolution without worsening fibrosis after one year of treatment. A companion phase 3 study, ENLIGHTEN-Cirrhosis, is assessing pegozafermin in compensated MASH cirrhosis (NCT06419374) to determine its efficacy and safety in more advanced disease.
BFKB8488A (Roche/Genentech)
BFKB8488A is a humanized monoclonal antibody that selectively agonizes the FGFR1/β-Klotho complex, avoiding FGFR2 and FGFR3 activation. It is unique among FGF21-based therapies as a receptor-targeted biologic with long dosing intervals and high specificity.
In a Phase I trial in obese and diabetic adults, single and multiple doses of BFKB8488A produced dose-dependent reductions in hepatic fat, plasma triglycerides, and body weight, along with marked increases in adiponectin.39 Participants also reported reduced preference for sweet foods, consistent with FGF21’s central role in dietary reward and sugar intake. The safety profile was favourable, with mostly mild gastrointestinal symptoms. Detailed histologic data in MASH remain limited compared to efruxifermin and pegozafermin.
As of November 2025, BFKB8488A has not entered phase 3 trials for MASH. Ongoing and planned phase 2 studies will determine whether its selective FGFR1/β-Klotho activation and long-acting antibody format justify progression to large-scale histologic and outcome-based trials.
Potential Risks and Safety Profile of FGF21 Analogues
Across clinical trials, FGF21 analogues have demonstrated an excellent overall safety profile. The most frequent adverse events include mild gastrointestinal symptoms such as nausea, diarrhea, or decreased appetite, which are typically transient and dose-related.82 Injection-site reactions are infrequent and self-limited. Laboratory findings show modest, reversible decreases in serum bone formation markers, possibly reflecting FGF21 effects on bone turnover observed in animal studies.83,84 However, no increase in fracture incidence or bone mineral density loss has been reported in human trials. Minor reductions in blood pressure and heart rate have been noted, attributed to improved autonomic tone and metabolic efficiency rather than direct cardiovascular toxicity.85 No significant hepatotoxicity, renal impairment, or immune-mediated hypersensitivity has been documented to date. Given the endocrine breadth of FGF21, long-term surveillance for potential effects on thyroid, reproductive, and bone metabolism remains warranted, but cumulative trial data support an excellent benefit-to-risk balance.
Future Role of FGF21 in MASH and Beyond
FGF21 analogues represent a paradigm shift toward multi-target metabolic therapy for MASH. Unlike agents that act on single pathways, such as FXR or PPARs, FGF21 addresses the full metabolic-inflammatory-fibrotic spectrum. It improves hepatic steatosis, insulin sensitivity, lipid metabolism, and systemic inflammation while mitigating fibrosis progression. The next generation of trials will determine whether histologic and biochemical benefits translate into clinical outcomes such as reduced progression to cirrhosis, decompensation, and cardiovascular events. Combination strategies with GLP-1 receptor agonists or thyroid hormone receptor β agonists (eg., resmetirom) may further enhance efficacy through complementary mechanisms. Beyond MASH, FGF21 analogues are being explored for the treatment of metabolic and fibrotic disorders, including chronic kidney disease, heart failure with preserved ejection fraction, and obesity-related cardiomyopathy.19 The hormone’s pleiotropic effects on mitochondrial efficiency, oxidative stress, and inflammation make it an attractive candidate for systemic metabolic modulation.
Role of FGF21 Beyond MASH to Other Liver Diseases
FGF21 is primarily studied in the context of MASH, but preclinical and clinical data suggest broader relevance. In alcohol-associated liver disease, elevated FGF21 levels are associated with a protective response, and FGF21 administration can reverse disease progression and reduce fibrosis, indicating therapeutic potential.86 Experimental models of toxin-induced hepatic fibrosis (eg., dimethylnitrosamine-induced) show that FGF21 attenuates fibrogenesis by inhibiting hepatic stellate cell activation and modulating TGF-β/Smad2/3 and NF-κB signalling pathways, suggesting direct antifibrotic actions independent of metabolic dysfunction.87 Additionally, FGF21 deficiency has been linked to increased risk and severity of HCC, and its presence may mitigate oncogenic processes in the liver, although clinical translation in HCC remains investigational.88 FGF21’s effects on fibrosis appear to be mediated by both direct actions on hepatic stellate cells and indirect effects via modulation of inflammation, lipid metabolism, and cellular stress responses.
Beyond the Liver: FGF21 in Extrahepatic Fibrotic and Metabolic Diseases
Fibroblast growth factor 21 (FGF21) plays a significant antifibrotic and metabolic regulatory role in extrahepatic tissues, including the lung, kidney, heart, adipose tissue, and pancreas, beyond its established hepatic effects.
FGF21 exerts antifibrotic effects in the lung: In idiopathic pulmonary fibrosis (IPF), FGF21 levels are elevated, and animal models show that FGF21 deficiency worsens lung fibrosis, while administration of FGF21 analogs mitigates fibrogenesis, primarily by inhibiting alveolar epithelial cell apoptosis rather than directly altering fibroblast phenotype.89 Similarly, in the kidney, FGF21 is upregulated in response to fibrotic injury and directly inhibits renal fibrosis by suppressing the Wnt/β-catenin signalling pathway, as demonstrated in mouse models of unilateral ureteral obstruction.90 In the cardiovascular system, FGF21 is associated with reduced risk and progression of atherosclerosis, coronary artery disease, and heart failure, likely through its anti-inflammatory, antioxidative, and metabolic actions.91 Elevated FGF21 levels predict the incidence and progression of cardiovascular and metabolic diseases, and experimental models suggest FGF21 protects against cardiac injury and remodelling.92 Metabolically, FGF21 regulates glucose and lipid homeostasis in adipose tissue, enhances insulin sensitivity, promotes browning of white adipose tissue, and increases energy expenditure.34 In the pancreas, FGF21 has anti-inflammatory effects and may preserve β-cell function.10 Elevated FGF21 is observed in obesity, type 2 diabetes, and metabolic syndrome, and is associated with disease severity and complications, including nephropathy and atheromatosis.93
Summary and Future Directions
FGF21 has emerged as one of the most promising metabolic hormones in the treatment of MASH. Over the past two decades, the understanding of FGF21 has progressed from its initial description as afasting-induced hepatokine to its current recognition as asystemic regulator of energy homeostasis, lipid turnover, and inflammatory balance. Through its actions on the liver, adipose tissue, skeletal muscle, and brain, FGF21 coordinates an integrated response to metabolic stress that directly targets the pathogenic drivers of MASH.
FGF21 improves insulin resistance by enhancing fatty acid oxidation, reducing de novo lipogenesis, and correcting hepatic and adipose metabolism. These metabolic effects are reinforced by anti-inflammatory and antifibrotic actions, partly through the FGF21 adiponectin axis. In preclinical models, FGF21 supplementation reverses steatohepatitis, lowers oxidative and ER stress, and down-regulates fibrogenic gene expression, supporting pharmacologic replacement in MASH, where endogenous FGF21 is elevated but functionally resistant.
Clinical trials with pegbelfermin, efruxifermin, and pegozafermin confirm reductions in hepatic fat, triglycerides, and non-invasive fibrosis markers, along with increases in HDL cholesterol and adiponectin, with good tolerability. These agents also improve systemic insulin sensitivity and lipid balance, suggesting dual hepatic and cardiovascular benefit in a population with high cardiometabolic risk. Key remaining questions include whether these improvements translate into fewer cirrhosis-related events and cardiovascular outcomes, and whether long-term maintenance therapy will be required to sustain benefit.
Combination therapy is another promising frontier. FGF21 analogues may complement agents that target parallel metabolic pathways, such as GLP-1 receptor agonists, FXR agonists, or THR-β modulators, such as resmetirom. These combinations could enhance weight loss, lipid metabolism, and fibrosis regression beyond the effect of any single drug. Moreover, FGF21’s favorable safety and cardiovascular profile make it an ideal backbone for such regimens. Precision medicine approaches that incorporate genetic, metabolic, and transcriptomic data may also help identify subgroups, such as those with pronounced dyslipidemia or adiponectin deficiency, who are most likely to benefit.
Beyond MASH, FGF21’s physiological role extends to other chronic diseases characterized by metabolic stress and fibrosis. Studies in animal models show beneficial effects in cardiac hypertrophy, renal fibrosis, and obesity-related cardiomyopathy. These data highlight FGF21’s potential as a systemic antifibrotic and metabolic modulator rather than a liver-specific intervention. Its impact on mitochondrial function, oxidative balance, and cellular survival pathways suggests applicability to a wide range of chronic fibrotic and metabolic conditions.
In conclusion, FGF21 represents a unifying concept in metabolic medicine, a single pathway capable of influencing lipid flux, insulin action, inflammation, and fibrosis across organ systems. Among the many agents in development for MASH, it is distinguished by its broad action and its ability to improve the metabolic phenotype underlying both hepatic and cardiovascular disease. As phase 3 data mature, FGF21-based therapies may redefine the management of MASH, shifting the focus from isolated hepatic endpoints toward holistic metabolic restoration. The ongoing evolution of long-acting analogues and rational combination strategies offers an opportunity not only to alter the course of MASH but to establish FGF21 as a cornerstone therapy for systemic metabolic fibrosis.
Disclosure
This work was supported from intramural funds from Stravitz-Sanyal Institute for Liver disease and Metabolic Health, VCU, Richmond, VA. The sponsor had no role in defining the project, its implementation, data analysis or contents of this manuscript. Dr Arun Sanyal reports stock options from Inversago, Durect, Rivus, Northsea, consultant from Intercept, Gilead, Merck, Boehringer Ingelheim, NovoNordisk, consultant from Eli Lilly, Amgen, Alnylam, Regeneron, Altimmune, consultant from 89 Bio, Madrigal, Boston Pharma, Glaxo Smith Kline, consultant from Genentech, Pliant, Avant Sante, Surrozen, Poxel, Hanmi, consultant from LG Chem, Akero Therapeutics, Salix, Zydus, Path AI, consultant from Histoindex, Genfit, Inventiva, Livergenix, Institution has received grants from Intercept, Merck, Echosens, Novo Nordisk, Institution has received grants from Eli Lilly, Amgen, Boehringer Ingelheim, Institution has received grants from Gilead, Hanmi and Zydus, Royalties from Elsevier and Wolter Kluwers, outside the submitted work. The authors report no other conflicts of interest in this work.
References
1. Wong VW, Ekstedt M, Wong GL-H, et al. Changing epidemiology, global trends and implications for outcomes of NAFLD. J Hepatol. 2023;79(3):842–17. doi:10.1016/j.jhep.2023.04.036
2. Nishimura T, Nakatake Y, Konishi M, et al. Identification of a novel FGF, FGF-21, preferentially expressed in the liver. Biochim Biophys Acta. 2000;1492(1):203–206. doi:10.1016/S0167-4781(00)00067-1
3. Goetz R, Beenken A, Ibrahimi OA, et al. Molecular insights into the klotho-dependent, endocrine mode of action of fibroblast growth factor 19 subfamily members. Mol Cell Biol. 2007;27(9):3417–3428. doi:10.1128/MCB.02249-06
4. Badman MK, Koester A, Flier JS, et al. Fibroblast growth factor 21-deficient mice demonstrate impaired adaptation to ketosis. Endocrinology. 2009;150(11):4931–4940. doi:10.1210/en.2009-0532
5. Ding X, Boney-Montoya J, Owen B, et al. βKlotho is required for fibroblast growth factor 21 effects on growth and metabolism. Cell Metab. 2012;16(3):387–393. doi:10.1016/j.cmet.2012.08.002
6. Zhang X, Ibrahimi OA, Olsen SK, et al. Receptor specificity of the fibroblast growth factor family. The complete mammalian FGF family. J Biol Chem. 2006;281(23):15694–15700. doi:10.1074/jbc.M601252200
7. Itoh N, Ohta H, Konishi M. Endocrine FGFs: evolution, physiology, pathophysiology, and pharmacotherapy. Front Endocrinol. 2015;6:154. doi:10.3389/fendo.2015.00154
8. Beenken A, Mohammadi M. The structural biology of the FGF19 subfamily. Adv Exp Med Biol. 2012;728:1–24.
9. Yang C, Jin C, Li X, et al. Differential specificity of endocrine FGF19 and FGF21 to FGFR1 and FGFR4 in complex with KLB. PLoS One. 2012;7(3):e33870. doi:10.1371/journal.pone.0033870
10. Fisher FM, Maratos-Flier E. Understanding the physiology of FGF21. Annu Rev Physiol. 2016;78:223–241. doi:10.1146/annurev-physiol-021115-105339
11. Lee P, Brychta RJ, Linderman J, et al. Mild cold exposure modulates fibroblast growth factor 21 (FGF21) diurnal rhythm in humans: relationship between FGF21 levels, lipolysis, and cold-induced thermogenesis. J Clin Endocrinol Metab. 2013;98(1):E98–102. doi:10.1210/jc.2012-3107
12. Ribas F, Villarroya J, Hondares E, et al. FGF21 expression and release in muscle cells: involvement of MyoD and regulation by mitochondria-driven signalling. Biochem J. 2014;463(2):191–199. doi:10.1042/BJ20140403
13. Moyers JS, Shiyanova TL, Mehrbod F, et al. Molecular determinants of FGF-21 activity-synergy and cross-talk with PPARgamma signaling. J Cell Physiol. 2007;210(1):1–6. doi:10.1002/jcp.20847
14. Laeger T, Albarado D, Burke S, et al. Metabolic responses to dietary protein restriction require an increase in FGF21 that is delayed by the absence of GCN2. Cell Rep. 2016;16(3):707–716. doi:10.1016/j.celrep.2016.06.044
15. Lundsgaard AM, Fritzen AM, Sjøberg KA, et al. Circulating FGF21 in humans is potently induced by short term overfeeding of carbohydrates. Mol Metab. 2017;6(1):22–29. doi:10.1016/j.molmet.2016.11.001
16. Wan XS, Lu X-H, Xiao Y-C, et al. ATF4- and CHOP-dependent induction of FGF21 through endoplasmic reticulum stress. Biomed Res Int. 2014;2014:807874. doi:10.1155/2014/807874
17. Kim H, Mendez R, Zheng Z, et al. Liver-enriched transcription factor CREBH interacts with peroxisome proliferator-activated receptor α to regulate metabolic hormone FGF21. Endocrinology. 2014;155(3):769–782. doi:10.1210/en.2013-1490
18. Jager J, Wang F, Fang B, et al. The nuclear receptor rev-erbα regulates adipose tissue-specific FGF21 signaling. J Biol Chem. 2016;291(20):10867–10875. doi:10.1074/jbc.M116.719120
19. Packer M. The adipokine hypothesis of heart failure with a preserved ejection fraction: a novel framework to explain pathogenesis and guide treatment. J Am Coll Cardiol. 2025;86(16):1269–1373. doi:10.1016/j.jacc.2025.06.055
20. Kilkenny DM, Rocheleau JV. The FGF21 receptor signaling complex: klothoβ, FGFR1c, and other regulatory interactions. Vitam Horm. 2016;101:17–58.
21. Holland WL, Adams A, Brozinick J, et al. An FGF21-adiponectin-ceramide axis controls energy expenditure and insulin action in mice. Cell Metab. 2013;17(5):790–797. doi:10.1016/j.cmet.2013.03.019
22. Bookout AL, de Groot MHM, Owen BM, et al. FGF21 regulates metabolism and circadian behavior by acting on the nervous system. Nat Med. 2013;19(9):1147–1152. doi:10.1038/nm.3249
23. Coate KC, Hernandez G, Thorne CA, et al. FGF21 is an exocrine pancreas secretagogue. Cell Metab. 2017;25(2):472–480. doi:10.1016/j.cmet.2016.12.004
24. Planavila A, Redondo-Angulo I, Villarroya F. FGF21 and cardiac physiopathology. Front Endocrinol. 2015;6:133. doi:10.3389/fendo.2015.00133
25. Potthoff MJ, Inagaki T, Satapati S, et al. FGF21 induces PGC-1alpha and regulates carbohydrate and fatty acid metabolism during the adaptive starvation response. Proc Natl Acad Sci U S A. 2009;106(26):10853–10858. doi:10.1073/pnas.0904187106
26. Ge X, Chen C, Hui X, et al. Fibroblast growth factor 21 induces glucose transporter-1 expression through activation of the serum response factor/Ets-like protein-1 in adipocytes. J Biol Chem. 2011;286(40):34533–34541. doi:10.1074/jbc.M111.248591
27. Schlein C, Talukdar S, Heine M, et al. FGF21 lowers plasma triglycerides by accelerating lipoprotein catabolism in white and brown adipose tissues. Cell Metab. 2016;23(3):441–453. doi:10.1016/j.cmet.2016.01.006
28. Lin Z, Tian H, Lam KL, et al. Adiponectin mediates the metabolic effects of FGF21 on glucose homeostasis and insulin sensitivity in mice. Cell Metab. 2013;17(5):779–789. doi:10.1016/j.cmet.2013.04.005
29. Talukdar S, Owen B, Song P, et al. FGF21 regulates sweet and alcohol preference. Cell Metab. 2016;23(2):344–349. doi:10.1016/j.cmet.2015.12.008
30. Zhang X, Yeung DCY, Karpisek M, et al. Serum FGF21 levels are increased in obesity and are independently associated with the metabolic syndrome in humans. Diabetes. 2008;57(5):1246–1253. doi:10.2337/db07-1476
31. Hotta Y, Nakamura H, Konishi M, et al. Fibroblast growth factor 21 regulates lipolysis in white adipose tissue but is not required for ketogenesis and triglyceride clearance in liver. Endocrinology. 2009;150(10):4625–4633. doi:10.1210/en.2009-0119
32. Chen MZ, Chang JC, Zavala-Solorio J, et al. FGF21 mimetic antibody stimulates UCP1-independent brown fat thermogenesis via FGFR1/βKlotho complex in non-adipocytes. Mol Metab. 2017;6(11):1454–1467. doi:10.1016/j.molmet.2017.09.003
33. Von Holstein-Rathlou S, BonDurant L, Peltekian L, et al. FGF21 mediates endocrine control of simple sugar intake and sweet taste preference by the liver. Cell Metab. 2016;23(2):335–343. doi:10.1016/j.cmet.2015.12.003
34. BonDurant LD, Ameka M, Naber MC, et al. FGF21 regulates metabolism through adipose-dependent and -independent mechanisms. Cell Metab. 2017;25(4):935–944.e4. doi:10.1016/j.cmet.2017.03.005
35. Kim AM, Somayaji VR, Dong JQ, et al. Once-weekly administration of a long-acting fibroblast growth factor 21 analogue modulates lipids, bone turnover markers, blood pressure and body weight differently in obese people with hypertriglyceridaemia and in non-human primates. Diabetes Obes Metab. 2017;19(12):1762–1772. doi:10.1111/dom.13023
36. Sanyal A, Charles ED, Neuschwander-Tetri BA, et al. Pegbelfermin (BMS-986036), a PEGylated fibroblast growth factor 21 analogue, in patients with non-alcoholic steatohepatitis: a randomised, double-blind, placebo-controlled, phase 2a trial. Lancet. 2019;392(10165):2705–2717. doi:10.1016/S0140-6736(18)31785-9
37. Charles ED, Neuschwander-Tetri BA, Pablo Frias J, et al. Pegbelfermin (BMS-986036), PEGylated FGF21, in patients with obesity and type 2 diabetes: results from a randomized phase 2 study. Obesity. 2019;27(1):41–49. doi:10.1002/oby.22344
38. Harrison SA, Ruane PJ, Freilich BL, et al. Efruxifermin in non-alcoholic steatohepatitis: a randomized, double-blind, placebo-controlled, phase 2a trial. Nat Med. 2021;27(7):1262–1271. doi:10.1038/s41591-021-01425-3
39. Wong C, Dash A, Fredrickson J, et al. Fibroblast growth factor receptor 1/Klothoβ agonist BFKB8488A improves lipids and liver health markers in patients with diabetes or NAFLD: a phase 1b randomized trial. Hepatology. 2023;78(3):847–862. doi:10.1002/hep.32742
40. Xu J, Stanislaus S, Chinookoswong N, et al. Acute glucose-lowering and insulin-sensitizing action of FGF21 in insulin-resistant mouse models--association with liver and adipose tissue effects. Am J Physiol Endocrinol Metab. 2009;297(5):E1105–14. doi:10.1152/ajpendo.00348.2009
41. So WY, Cheng Q, Xu A, et al. Loss of fibroblast growth factor 21 action induces insulin resistance, pancreatic islet hyperplasia and dysfunction in mice. Cell Death Dis. 2015;6(3):e1707. doi:10.1038/cddis.2015.80
42. Nie Z, Xu J, Liu Y, et al. Effects and safety of FGF21 analogs on glycemic parameters, lipid profiles, and adiponectin in overweight and obese adults: a meta-analysis of randomized controlled trials. Int J Endocrinol. 2025;2025(1):9943228. doi:10.1155/ije/9943228
43. Kharitonenkov A, Shiyanova TL, Koester A, et al. FGF-21 as a novel metabolic regulator. J Clin Invest. 2005;115(6):1627–1635. doi:10.1172/JCI23606
44. Berglund ED, Li CY, Bina HA, et al. Fibroblast growth factor 21 controls glycemia via regulation of hepatic glucose flux and insulin sensitivity. Endocrinology. 2009;150(9):4084–4093. doi:10.1210/en.2009-0221
45. Gaich G, Chien J, Fu H, et al. The effects of LY2405319, an FGF21 analog, in obese human subjects with type 2 diabetes. Cell Metab. 2013;18(3):333–340. doi:10.1016/j.cmet.2013.08.005
46. Harrison SA, Frias JP, Neff G, et al. Safety and efficacy of once-weekly efruxifermin versus placebo in non-alcoholic steatohepatitis (HARMONY): a multicentre, randomised, double-blind, placebo-controlled, phase 2b trial. Lancet Gastroenterol Hepatol. 2023;8(12):1080–1093. doi:10.1016/S2468-1253(23)00272-8
47. Kruse R, Vienberg SG, Vind BF, et al. Effects of insulin and exercise training on FGF21, its receptors and target genes in obesity and type 2 diabetes. Diabetologia. 2017;60(10):2042–2051. doi:10.1007/s00125-017-4373-5
48. Yu Y, He J, Li S, et al. Fibroblast growth factor 21 (FGF21) inhibits macrophage-mediated inflammation by activating Nrf2 and suppressing the NF-κB signaling pathway. Int Immunopharmacol. 2016;38:144–152. doi:10.1016/j.intimp.2016.05.026
49. Wang N, Zhao -T-T, Li S-M, et al. Fibroblast growth factor 21 exerts its anti-inflammatory effects on multiple cell types of adipose tissue in obesity. Obesity. 2019;27(3):399–408. doi:10.1002/oby.22376
50. Tucker B, McClelland RL, Allison MA, et al. Relationship of fibroblast growth factor 21 levels with inflammation, lipoproteins and non-alcoholic fatty liver disease. Atherosclerosis. 2020;299:38–44. doi:10.1016/j.atherosclerosis.2020.03.009
51. Li X, Zheng K, Liu L, et al. Relationship of postprandial fibroblast growth factor 21 with lipids, inflammation and metabolic dysfunction-associated fatty liver disease during oral fat tolerance test. Front Endocrinol. 2024;15:1343853. doi:10.3389/fendo.2024.1343853
52. Sharma S, Dixon T, Jung S, et al. Dietary methionine restriction reduces inflammation independent of FGF21 action. Obesity. 2019;27(8):1305–1313. doi:10.1002/oby.22534
53. Friedman SL, Neuschwander-Tetri BA, Rinella M, et al. Mechanisms of NAFLD development and therapeutic strategies. Nat Med. 2018;24(7):908–922. doi:10.1038/s41591-018-0104-9
54. Yano K, Yamaguchi K, Seko Y, et al. Hepatocyte-specific fibroblast growth factor 21 overexpression ameliorates high-fat diet-induced obesity and liver steatosis in mice. Lab Invest. 2022;102(3):281–289. doi:10.1038/s41374-021-00680-9
55. Xu J, Lloyd DJ, Hale C, et al. Fibroblast growth factor 21 reverses hepatic steatosis, increases energy expenditure, and improves insulin sensitivity in diet-induced obese mice. Diabetes. 2009;58(1):250–259. doi:10.2337/db08-0392
56. Loomba R, Lawitz EJ, Frias JP, et al. Safety, pharmacokinetics, and pharmacodynamics of pegozafermin in patients with non-alcoholic steatohepatitis: a randomised, double-blind, placebo-controlled, phase 1b/2a multiple-ascending-dose study. Lancet Gastroenterol Hepatol. 2023;8(2):120–132. doi:10.1016/S2468-1253(22)00347-8
57. Choi W, Woo GH, Kwon T-H, et al. Obesity-Driven metabolic disorders: the interplay of inflammation and mitochondrial dysfunction. Int J Mol Sci. 2025;26(19):9715.
58. Gómez-Sámano M, Grajales-Gómez M, Zuarth-Vázquez JM, et al. Fibroblast growth factor 21 and its novel association with oxidative stress. Redox Biol. 2017;11:335–341. doi:10.1016/j.redox.2016.12.024
59. Jiang S, Yan C, Fang Q-C, et al. Fibroblast growth factor 21 is regulated by the IRE1α-XBP1 branch of the unfolded protein response and counteracts endoplasmic reticulum stress-induced hepatic steatosis. J Biol Chem. 2014;289(43):29751–29765. doi:10.1074/jbc.M114.565960
60. Fisher FM, Chui PC, Nasser IA, et al. Fibroblast growth factor 21 limits lipotoxicity by promoting hepatic fatty acid activation in mice on methionine and choline-deficient diets. Gastroenterology. 2014;147(5):1073–83.e6. doi:10.1053/j.gastro.2014.07.044
61. Cai J, Zhang XJ, Li H. The role of innate immune cells in nonalcoholic steatohepatitis. Hepatology. 2019;70(3):1026–1037. doi:10.1002/hep.30506
62. Li JY, Wang N, Khoso MH, et al. FGF-21 elevated IL-10 production to correct LPS-Induced inflammation. Inflammation. 2018;41(3):751–759. doi:10.1007/s10753-018-0729-3
63. Wang WF, Ma L, Liu M-Y, et al. A novel function for fibroblast growth factor 21: stimulation of NADPH oxidase-dependent ROS generation. Endocrine. 2015;49(2):385–395. doi:10.1007/s12020-014-0502-9
64. Noureddin M, Frias JP, Neff GW, et al. Safety and efficacy of once-weekly efruxifermin versus placebo in metabolic dysfunction-associated steatohepatitis (HARMONY): 96-week results from a multicentre, randomised, double-blind, placebo-controlled, phase 2b trial. Lancet. 2025;406(10504):719–730. doi:10.1016/S0140-6736(25)01073-6
65. Younossi ZM, de Avila L, Petta S, et al. Predictors of fibrosis, clinical events and mortality in MASLD: data from the Global-MASLD study. Hepatology. 2025. doi:10.1097/HEP.0000000000001617
66. Dolivo DM, Larson SA, Dominko T. Fibroblast growth factor 2 as an antifibrotic: antagonism of myofibroblast differentiation and suppression of pro-fibrotic gene expression. Cytokine Growth Factor Rev. 2017;38:49–58. doi:10.1016/j.cytogfr.2017.09.003
67. Sato-Matsubara M, Matsubara T, Daikoku A, et al. Fibroblast growth factor 2 (FGF2) regulates cytoglobin expression and activation of human hepatic stellate cells via JNK signaling. J Biol Chem. 2017;292(46):18961–18972. doi:10.1074/jbc.M117.793794
68. Loomba R, Sanyal AJ, Kowdley KV, et al. Randomized, controlled trial of the FGF21 analogue pegozafermin in NASH. N Engl J Med. 2023;389(11):998–1008. doi:10.1056/NEJMoa2304286
69. Lu Y, Li R, Zhu J, et al. Fibroblast growth factor 21 facilitates peripheral nerve regeneration through suppressing oxidative damage and autophagic cell death. J Cell Mol Med. 2019;23(1):497–511. doi:10.1111/jcmm.13952
70. Chen X, Tong G, Fan J, et al. FGF21 promotes migration and differentiation of epidermal cells during wound healing via SIRT1-dependent autophagy. Br J Pharmacol. 2022;179(5):1102–1121. doi:10.1111/bph.15701
71. Li Z, Qiu X, Guan G, et al. The role of FGF-21 in promoting diabetic wound healing by modulating high glucose-induced inflammation. Heliyon. 2024;10(9):e30022. doi:10.1016/j.heliyon.2024.e30022
72. Ye D, Wang Y, Li H, et al. Fibroblast growth factor 21 protects against Acetaminophen-induced hepatotoxicity by potentiating peroxisome proliferator-activated receptor coactivator protein-1α-mediated antioxidant capacity in mice. Hepatology. 2014;60(3):977–989. doi:10.1002/hep.27060
73. Qiang W, Shen T, Noman M, et al. Fibroblast growth factor 21 augments autophagy and reduces apoptosis in damaged liver to improve tissue regeneration in zebrafish. Front Cell Dev Biol. 2021;9:756743. doi:10.3389/fcell.2021.756743
74. Hecht R, Li Y-S, Sun J, et al. Rationale-Based engineering of a potent long-acting FGF21 analog for the treatment of type 2 diabetes. PLoS One. 2012;7(11):e49345. doi:10.1371/journal.pone.0049345
75. Krupinski J M.N, Kozhich A, Chiney M, Morin P, Christian R. Effects of BMS-986036 (pegylated fibroblast growth factor 21) on hepatic steatosis and fibrosis in a mouse model of nonalcoholic steatohepatitis. Hepatology. 2016;64:749A.
76. Loomba R, Sanyal AJ, Nakajima A, et al. Pegbelfermin in patients with nonalcoholic steatohepatitis and stage 3 fibrosis (FALCON 1): a randomized phase 2b study. Clin Gastroenterol Hepatol. 2024;22(1):102–112.e9. doi:10.1016/j.cgh.2023.04.011
77. Abdelmalek MF, Sanyal AJ, Nakajima A, et al. Pegbelfermin in patients with nonalcoholic steatohepatitis and compensated cirrhosis (FALCON 2): a randomized phase 2b study. Clin Gastroenterol Hepatol. 2024;22(1):113–123.e9. doi:10.1016/j.cgh.2023.04.012
78. Stanislaus S, Hecht R, Yie J, et al. A novel Fc-FGF21 with improved resistance to proteolysis, increased affinity toward β-Klotho, and enhanced efficacy in mice and cynomolgus monkeys. Endocrinology. 2017;158(5):1314–1327. doi:10.1210/en.2016-1917
79. Noureddin M, Rinella ME, Chalasani NP, et al. Efruxifermin in compensated liver cirrhosis caused by MASH. N Engl J Med. 2025;392(24):2413–2424. doi:10.1056/NEJMoa2502242
80. Rosenstock M, Tseng L, Pierce A, et al. The novel GlycoPEGylated FGF21 analog pegozafermin activates human FGF receptors and improves metabolic and liver outcomes in diabetic monkeys and healthy human volunteers. J Pharmacol Exp Ther. 2023;387(2):204–213. doi:10.1124/jpet.123.001618
81. Bhatt DL, Bays HE, Miller M, et al. The FGF21 analog pegozafermin in severe hypertriglyceridemia: a randomized phase 2 trial. Nat Med. 2023;29(7):1782–1792. doi:10.1038/s41591-023-02427-z
82. Jeong C, Han N, Jeon N, et al. Efficacy and safety of fibroblast growth factor-21 analogs for the treatment of metabolic dysfunction-associated steatohepatitis: a systematic review and meta-analysis. Clin Pharmacol Ther. 2024;116(1):72–81. doi:10.1002/cpt.3278
83. Wei W, Dutchak PA, Wang X, et al. Fibroblast growth factor 21 promotes bone loss by potentiating the effects of peroxisome proliferator-activated receptor γ. Proc Natl Acad Sci U S A. 2012;109(8):3143–3148. doi:10.1073/pnas.1200797109
84. Talukdar S, Zhou Y, Li D, et al. A Long-Acting FGF21 molecule, PF-05231023, decreases body weight and improves lipid profile in non-human primates and type 2 diabetic subjects. Cell Metab. 2016;23(3):427–440. doi:10.1016/j.cmet.2016.02.001
85. Huang Z, Xu A, Cheung BMY. The potential role of fibroblast growth factor 21 in lipid metabolism and hypertension. Curr Hypertens Rep. 2017;19(4):28. doi:10.1007/s11906-017-0730-5
86. Desai BN, Singhal G, Watanabe M, et al. Fibroblast growth factor 21 (FGF21) is robustly induced by ethanol and has a protective role in ethanol associated liver injury. Mol Metab. 2017;6(11):1395–1406. doi:10.1016/j.molmet.2017.08.004
87. Xu P, Zhang Y, Liu Y, et al. Fibroblast growth factor 21 attenuates hepatic fibrogenesis through TGF-β/smad2/3 and NF-κB signaling pathways. Toxicol Appl Pharmacol. 2016;290:43–53. doi:10.1016/j.taap.2015.11.012
88. Singhal G, Kumar G, Chan S, et al. Deficiency of fibroblast growth factor 21 (FGF21) promotes hepatocellular carcinoma (HCC) in mice on a long term obesogenic diet. Mol Metab. 2018;13:56–66. doi:10.1016/j.molmet.2018.03.002
89. Ghanem M, Archer G, Justet A, et al. FGF21 signaling exerts antifibrotic properties during pulmonary fibrosis. Am J Respir Crit Care Med. 2025;211(3):486–498. doi:10.1164/rccm.202311-2021OC
90. Zhong W, Jiang Y, Wang H, et al. Fibroblast growth factor 21 alleviates unilateral ureteral obstruction-induced renal fibrosis by inhibiting Wnt/β-catenin signaling pathway. Biochim Biophys Acta Mol Cell Res. 2024;1871(2):119620. doi:10.1016/j.bbamcr.2023.119620
91. Tan H, Yue T, Chen Z, et al. Targeting FGF21 in cardiovascular and metabolic diseases: from mechanism to medicine. Int J Biol Sci. 2023;19(1):66–88. doi:10.7150/ijbs.73936
92. Lakhani I, Gong M, Wong WT, et al. Fibroblast growth factor 21 in cardio-metabolic disorders: a systematic review and meta-analysis. Metabolism. 2018;83:11–17. doi:10.1016/j.metabol.2018.01.017
93. Iglesias P, Selgas R, Romero S, et al. Biological role, clinical significance, and therapeutic possibilities of the recently discovered metabolic hormone fibroblastic growth factor 21. Eur J Endocrinol. 2012;167(3):301–309. doi:10.1530/EJE-12-0357
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Association of Serum Leptin and Adiponectin Concentrations with Type 2 Diabetes Biomarkers and Complications Among Saudi Women
Mohammed Saeed W, Nasser Binjawhar D
Diabetes, Metabolic Syndrome and Obesity 2023, 16:2129-2140
Published Date: 13 July 2023
Lipotoxicity: A New Perspective in Type 2 Diabetes Mellitus
Chen B, Li T, Wu Y, Song L, Wang Y, Bian Y, Qiu Y, Yang Z
Diabetes, Metabolic Syndrome and Obesity 2025, 18:1223-1237
Published Date: 24 April 2025
Investigating the Pathogenesis and Treatment of Type 2 Diabetes from the Perspective of Adipose Tissue
Kang Z, Jin Z, Wu L, Sun A, Dong L, Li J, Li Q, Tong X
Diabetes, Metabolic Syndrome and Obesity 2025, 18:2343-2360
Published Date: 14 July 2025
Resmetirom: An Update on Therapy for Metabolic Dysfunction-Associated Steatohepatitis (MASH)
Van Kleef LA, Michel M, Alkhouri N, Brouwer WP, Schattenberg JM
Drug Design, Development and Therapy 2026, 20:559331
Published Date: 17 March 2026
Metabolic Crosstalk in Diabetic Kidney Disease: Synergistic Effects of Glucotoxicity and Lipotoxicity
Cao J, Dun Z, Tian D, Qiao Z, Wang Y
Diabetes, Metabolic Syndrome and Obesity 2026, 19:580866
Published Date: 23 March 2026