Back to Journals » International Journal of Nanomedicine » Volume 21
Extracellular Vesicle-Associated Non-Coding RNAs in Preeclampsia: Mechanistic Insights, Biomarker Discovery, and Emerging Nanomedicine Concepts
Authors Zhang Z ![]() , Du J
, Du J ![]() , Chen T, Teng Y
, Chen T, Teng Y ![]() , Chen J
, Chen J ![]() , Li Y, Zhang Y, Long Y, Li C, Wu Z, Li N, Yang Z
, Li Y, Zhang Y, Long Y, Li C, Wu Z, Li N, Yang Z
Received 1 April 2026
Accepted for publication 17 June 2026
Published 29 June 2026 Volume 2026:21 612044
DOI https://doi.org/10.2147/IJN.S612044
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Kamakhya Misra
Zhongsong Zhang,1,2,* Jiahui Du,3,* Tianze Chen,4,* Yuanyin Teng,5 Junhao Chen,6 Yanpei Li,7 Yang Zhang,8,9 Yingfei Long,1 Chanyu Li,1 Zhu Wu,10 Na Li,1 Zhiling Yang1,*
1Department of Obstetrics and Gynecology, The First Affiliated Hospital of Chengdu Medical College, Chengdu, Sichuan, 610500, People’s Republic of China; 2School of Clinical Medicine, Chengdu Medical College, Chengdu, Sichuan, 610500, People’s Republic of China; 3School of Laboratory Medicine and Life Sciences, Wenzhou Medical University, Wenzhou, Zhejiang, 325035, People’s Republic of China; 4Center for Reproductive Medicine, Yangzhou Maternal and Child Health Care Hospital Affiliated to Yangzhou University, Yangzhou, Jiangsu, 225000, People’s Republic of China; 5Institute of Hematology, Zhejiang University, Hangzhou, Zhejiang, 310003, People’s Republic of China; 6Department of Urology, The Second Affiliated Hospital of Kunming Medical University, Kunming, Yunnan, 650101, People’s Republic of China; 7School of Pharmacy, Chengdu University of Traditional Chinese Medicine, Chengdu, Sichuan, 611137, People’s Republic of China; 8Department of Vascular Surgery, Fuwai Yunnan Cardiovascular Hospital, Chinese Academy of Medical Sciences, Kunming, Yunnan, 650102, People’s Republic of China; 9Department of Vascular Surgery, Affiliated Cardiovascular Hospital of Kunming Medical University, Kunming, Yunnan, 650102, People’s Republic of China; 10The Hunan Provincial Key Laboratory of TCM Agricultural Biogenomics, Changsha Medical University, Changsha, Hunan, 410219, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Na Li, Email [email protected] Zhiling Yang, Email [email protected]
Abstract: Preeclampsia (PE) is a complex multisystem disorder that affects 2– 8% of pregnancies worldwide and poses substantial risks to maternal and fetal health. Current diagnostic approaches rely largely on clinical signs and angiogenic biomarkers, and available treatments remain primarily supportive; they do not directly reverse the placental or systemic mechanisms that drive the disease. Extracellular vesicles (EVs), including small EVs often termed exosomes, carry non-coding RNAs (ncRNAs) that may contribute to placental–maternal communication in both normal and pathological pregnancy. In PE, altered EV-associated microRNAs, long non-coding RNAs, and circular RNAs have been detected in placental tissues, trophoblast-derived systems, maternal plasma or serum, urine, amniotic fluid, and other pregnancy-related samples. However, these matrices should not be assumed to indicate definitive tissue or cellular origins without appropriate source-attribution methods. This review summarizes current evidence on EV-associated ncRNAs in PE from three perspectives: mechanistic studies, biomarker discovery, and exploratory nanomedicine strategies. First, we discuss how dysregulated EV-associated ncRNAs may contribute to trophoblast dysfunction, immune-inflammatory imbalance, endothelial injury, and angiogenic dysregulation. Second, we evaluate EV-associated ncRNAs as candidate liquid-biopsy biomarkers, emphasizing that most reported signatures remain at the discovery or early validation stage. Their clinical implementation will require standardized EV isolation, RNA profiling, normalization procedures, and validation in independent longitudinal cohorts. Third, we discuss engineered EVs and EV-mimetic nanocarriers as experimental platforms for ncRNA delivery and distinguish these preclinical therapeutic concepts from clinically established PE management. Rather than suggesting immediate diagnostic or therapeutic readiness, this review highlights the opportunities and limitations of EV-associated ncRNAs as a framework for future PE research. Key challenges include EV heterogeneity, limited discrimination among vesicular subtypes, uncertain tissue origins of circulating EV cargo, poor reproducibility across cohorts, safety concerns during pregnancy, scalable manufacturing, and ethical considerations related to maternal–fetal interventions. Future studies integrating rigorously characterized EV populations, multi-omics profiling, functional validation, and longitudinal clinical sampling are essential to determine whether EV-associated ncRNAs can be translated into reliable PE biomarkers or safe nanomedicine-based interventions.
Keywords: preeclampsia, extracellular vesicles, small extracellular vesicles, non-coding RNAs, liquid biopsy, biomarker discovery, EV-mimetic nanocarriers
Introduction
Preeclampsia (PE) is a multisystem disorder defined by new onset hypertension accompanied by proteinuria or other end organ dysfunction after 20 weeks of gestation. It affects 2 to 8% of pregnancies worldwide and remains a leading cause of maternal and perinatal morbidity and mortality.1,2 PE accounts for 11–14% of global maternal deaths, greatly endangering the health of pregnant women.1,3 Current diagnostic strategies rely mainly on clinical signs and biomarkers, including the soluble fms-like tyrosine kinase-1 to placental growth factor (sFlt-1/PlGF) ratio.3–5 However, a distinction should be made between diagnosis after clinical presentation and prediction before disease onset, because most available approaches are more established for diagnosis or short-term risk assessment than for reliable early prediction.4,6 Therapeutic options remain largely supportive. They include antihypertensive agents, magnesium sulfate to prevent eclampsia, and delivery as the only definitive intervention. This strategy can prevent severe maternal complications but may also lead to medically indicated preterm birth and related neonatal morbidity. These approaches do not directly reverse key pathophysiological processes, such as abnormal placentation, placental hypoxia, systemic inflammation, and endothelial dysfunction.7,8 Recent studies have highlighted the potential involvement of extracellular vesicles (EVs), particularly small EV-enriched preparations, in PE-associated placental–maternal communication.9,10 Therefore, there remains a need for biomarkers and therapeutic concepts that can be mechanistically informative, temporally useful, and clinically feasible, while recognizing that translation into routine PE care requires rigorous validation.9,11
EVs are membrane-bound particles released by many cell types, and small EVs are often operationally referred to as EVs when supported by appropriate characterization. Growing evidence indicates that EVs are involved in PE-related intercellular communication, although many studies analyze mixed EV or small EV-enriched fractions rather than vesicles with proven endosomal origin.12,13 These vesicles can carry bioactive cargo, including non-coding RNAs (ncRNAs) such as microRNAs, long non-coding RNAs, and circular RNAs. In PE, altered EV-associated ncRNAs have been detected in placental tissues, trophoblast models, maternal plasma or serum, urine, amniotic fluid, and other pregnancy-related samples.12,14–16 Importantly, these biofluids represent sampling matrices rather than definitive tissue origins; assigning a circulating EV-ncRNA signal to the placenta, maternal endothelium, immune cells, kidney, or fetal compartment requires additional source-attribution approaches, such as cell-specific markers, placental perfusion models, or paired tissue–biofluid analyses.17–19 Mechanistically, EV-associated ncRNAs may influence trophoblast migration and invasion, spiral artery remodeling, angiogenic balance, immune tolerance, inflammation, and endothelial function.20,21 For example, hypoxia-associated miRNAs such as miR-210 have been linked to mitochondrial metabolism and angiogenic regulation, whereas lncRNAs and circRNAs may participate in competing endogenous RNA networks that modulate inflammatory and vascular pathways.12,22 Nevertheless, many of these findings derive from cell models, animal experiments, small clinical cohorts, or cross-sectional studies, and should therefore be interpreted as mechanistic and associative evidence rather than proof of clinical utility.23
EV-associated ncRNAs are also being investigated as candidate liquid-biopsy biomarkers for PE; however, biomarker discovery should be clearly distinguished from clinically validated prediction or diagnosis.4,24 Sampling of plasma, serum, urine, and amniotic fluid offers opportunities to monitor pregnancy-related molecular changes in a minimally invasive manner.25,26 However, these matrices differ in biological composition, EV abundance, levels of contaminating non-vesicular RNA, and relevance to specific PE phenotypes.27,28 For early prediction, longitudinal sampling before 20 weeks of gestation is required to determine whether EV-associated ncRNA signatures can identify women at risk before clinical manifestations develop.29 For diagnosis or risk stratification after disease onset, EV-associated ncRNAs may provide complementary information on PE subtype, disease severity, angiogenic imbalance, renal involvement, or inflammatory burden. However, they should not yet be presented as replacements for established clinical assessment or angiogenic biomarkers.30,31 Major barriers include inconsistent EV isolation methods and nomenclature, incomplete discrimination among EV subtypes, pre-analytical variability, lack of standardized RNA normalization, small sample sizes, population heterogeneity, and insufficient independent validation. These challenges are particularly important in PE because early-onset and late-onset disease, fetal growth restriction-associated PE, PE with severe features, and superimposed PE may involve overlapping but non-identical EV-associated ncRNA profiles.32,33 Therapeutic delivery represents a distinct and less mature area of investigation than biomarker development. Engineered EVs, EV-mimetic nanoparticles, and hybrid nanovesicles have been explored in preclinical studies as experimental carriers for ncRNA mimics, inhibitors, or other modulators.34,35 Preclinical models suggest that EV-based delivery of regulatory miRNAs may modulate trophoblast function, angiogenic signaling, inflammation, and endothelial injury. However, these studies remain exploratory and should not be interpreted as evidence that placenta-specific targeting, fetal safety, or superiority over standard PE management has been established.17,36 In pregnancy, therapeutic nanomedicine must meet additional requirements, including biodistribution analysis across maternal organs, the placenta, and the fetus; evaluation of placental transfer; reproductive and developmental toxicity testing; immunogenicity assessment; dose control; scalable manufacturing; and ethical safeguards for maternal–fetal interventions.36,37 Thus, EV-based nanomedicine should currently be regarded as a promising experimental concept rather than a clinically ready therapeutic strategy for PE.
In this review, we examine EV-associated ncRNAs as exploratory molecular mediators and candidate biomarkers in PE and separately discuss emerging nanomedicine concepts for ncRNA delivery. We first summarize mechanistic evidence linking dysregulated EV-associated ncRNAs to trophoblast dysfunction, immune-inflammatory imbalance, endothelial injury, angiogenic dysregulation, and complement- or NET-related pathways. We then evaluate EV-associated ncRNAs detected in different sampling matrices as candidate biomarkers across pregnancy stages, with particular attention to prediction, diagnosis, and risk stratification. Finally, we discuss experimental EV-based and EV-mimetic delivery strategies, along with the methodological, safety-related, and translational barriers that must be addressed before clinical application. By adopting this structure, we aim to provide a balanced framework that highlights the promise of EV-associated ncRNAs in PE while avoiding premature claims of diagnostic or therapeutic readiness.
Extracellular Vesicle-Associated ncRNAs in the Pathogenesis of PE
The placenta is not merely the initiating site of PE but also an active signaling hub that continuously communicates with the maternal circulation. In this context, extracellular vesicles provide a nanoscale language through which placental stress can be translated into systemic endothelial dysfunction, immune imbalance, inflammatory activation, and angiogenic disturbance. Their ncRNA cargo, including miRNAs, lncRNAs, and circRNAs, is not randomly packaged but reflects disease stage, cellular origin, and pathological context (Figure 1). More importantly, these EV-associated ncRNAs converge on several therapeutically relevant axes, such as trophoblast invasion, spiral artery remodeling, endothelial repair, immune tolerance, oxidative stress, and angiogenic balance. These disease driving nodes provide rational molecular targets for early intervention and support the feasibility of using extracellular vesicles as natural or engineered carriers to restore protective ncRNA signals, inhibit pathogenic pathways, and enhance placenta directed therapy before irreversible maternal and fetal injury occurs.
 |
Figure 1 Placental extracellular vesicle-associated ncRNAs in the pathogenesis of PE. This schematic illustrates the generation and biological roles of placental extracellular vesicles (PEVs) during pregnancy and their potential involvement in PE-related pathophysiology. During chorionic villous development, placental cell populations, including cytotrophoblasts, syncytiotrophoblasts, Hofbauer cells, mesenchymal cells, and other cells at the maternal–fetal interface, release increasing amounts of PEVs into the intervillous space and maternal circulation. These vesicles include small EVs/exosome-enriched vesicles, microvesicles, and apoptotic bodies, and they carry diverse bioactive cargo, including proteins, lipids, nucleic acids, and EV-associated ncRNAs. In normal pregnancy, PEVs contribute to placentation, decidualization, immune tolerance, maternal vascular communication, and maternofetal signaling. In PE, placental stress, hypoxia, oxidative stress, inflammation, and trophoblast dysfunction may alter the abundance, subtype distribution, and ncRNA cargo of PEVs. Dysregulated EV-associated miRNAs, lncRNAs, and circRNAs may be transferred to maternal endothelial cells, immune cells, renal or vascular cells, and placental cells, thereby participating in PE-related processes such as impaired trophoblast functional phenotypes, angiogenic imbalance, endothelial activation, inflammatory cytokine production, altered maternal–fetal immune tolerance, complement activation, NET-associated vascular injury, and renal endothelial dysfunction. These mechanisms should be interpreted according to evidence source, because findings from human biofluid EV fractions, placental tissues, primary cells, cell-line models, placental explants, and animal models do not represent the same level of evidence. Adapted from Ortega MA, Fraile-Martínez O, García-Montero C et al Unfolding the role of placental-derived Extracellular Vesicles in Pregnancy: From homeostasis to pathophysiology. Front Cell Dev Biol. Copyright © 2022 by authors.38 Abbreviations: PEVs, placental extracellular vesicles; EVs, extracellular vesicles; PE, preeclampsia; ncRNAs, non-coding RNAs; miRNAs, microRNAs; lncRNAs, long non-coding RNAs; circRNAs, circular RNAs; NETs, neutrophil extracellular traps. |
Placenta- and Trophoblast-Associated EV ncRNAs
miRNAs Associated with Placental/Trophoblast EV Preparations
miR-210
MicroRNAs (miRNAs) have been implicated in trophoblast functional phenotypes that are relevant to early placentation, including cell proliferation, migration, invasion, survival, and responses to hypoxia (Figure 2).21 Several studies have reported altered miR-210 levels in PE placental tissues and maternal circulation, and some EV or small EV-enriched preparations from trophoblast-related systems or maternal blood have also been reported to contain miR-210. Where sequencing-based studies specify the mature strand, the transcript should be reported as miR-210-3p or miR-210-5p; when the original study does not provide arm-specific information, we refer to it as total miR-210 or miR-210 with the arm not specified.39,40 Functionally, miR-210 has been linked to hypoxia-associated trophoblast dysfunction by regulating targets such as iron-sulfur cluster assembly enzyme (ISCU) and caudal-type homeobox 2 (CDX2), which are involved in mitochondrial metabolism, trophoblast migration, and invasion.41 In HTR-8/SVneo cells and first-trimester placental explants, experimental miR-210 overexpression has been associated with reduced extravillous trophoblast outgrowth and altered ERK/MAPK-related signaling; however, these findings should be considered model-based observations rather than direct proof that miR-210 is sufficient or necessary for human PE development.42 Similarly, animal studies using hypoxia exposure or miR-210 manipulation provide evidence that miR-210 can influence placental adaptation and fetal growth under defined experimental conditions.42,43 Nevertheless, PE-like phenotypes in rodents do not fully recapitulate human PE, and these data support functional involvement rather than establishing miR-210 as a sole causal driver of the human disease. The oxygen concentrations used in vitro also require careful interpretation: 2–3% O2 may approximate aspects of physiological low oxygen during early placentation depending on gestational timing, whereas 1% O2 usually represents a more severe experimental hypoxic stress. Therefore, differences in oxygen tension, gestational stage, cell type, and EV isolation strategy may all affect the observed miR-210 response. Overall, miR-210, particularly miR-210-3p when specified, should be presented as an EV-associated candidate marker and mechanistic mediator of hypoxia-related trophoblast and endothelial dysfunction, rather than as a proven causal determinant or clinically established therapeutic target for PE.44
 |
Figure 2 Overview of dysregulated miRNAs and associated biological pathways in PE. This schematic summarizes representative miRNAs reported to be dysregulated in PE and their potential association with maternal circulating signals and placental pathophysiology. (A) The left pink highlighted panel summarizes dysregulated circulating miRNAs in PE and their associated functional categories, including transcriptional and epigenetic regulation, oxidative stress and mitochondrial function, neuronal function and ion transport, and extracellular matrix remodeling. Representative downstream molecules or related targets include MALAT1, SLC3A1, TUG1, H19, NEAT1, VPO1, MOTS-c, KCNA1, GPC1, and MMP-9. (B) The right Orange highlighted panel summarizes dysregulated placental miRNAs in PE and their potential involvement in angiogenesis, cell survival, oxidative stress, apoptosis, and immune responses. Representative downstream molecules or related targets include VEGFA, Ephrin-B2, EPHB4, CYR61, PIK3R2, IGF-1, HMOX1, TRAF6/RGS2, FOXO3, USF2/PPP3R1, HDAC2, and p53/PUMA. MiRNA names shown in red indicate miRNAs reported to be upregulated in PE, whereas miRNA names shown in blue indicate miRNAs reported to be downregulated in PE. Black upward and downward arrows next to target molecules indicate the reported direction of change of the corresponding downstream molecule or pathway in the summarized studies. Black connecting arrows indicate reported or proposed regulatory relationships between miRNAs and their associated targets or pathways. Bold text indicates major functional categories or pathway modules. The right highlighted panel represents placenta-associated dysregulated miRNAs, whereas the left highlighted panel represents circulating dysregulated miRNAs. These associations should be interpreted as a pathway-level summary and do not necessarily indicate direct causal regulation unless experimentally validated in the cited studies. Adapted from Oancea, Mihaela et al MicroRNAs in Preeclampsia: An Overview of Biomarkers and Potential Therapeutic Targets. International journal of molecular sciences. Copyright © 2025 by authors.45 Abbreviations: PE, preeclampsia; miRNAs, microRNAs; MALAT1, metastasis-associated lung adenocarcinoma transcript 1; SLC3A1, solute carrier family 3 member 1; TUG1, taurine-upregulated gene 1; H19, H19 imprinted maternally expressed transcript; NEAT1, nuclear paraspeckle assembly transcript 1; VPO1, vascular peroxidase 1; MOTS-c, mitochondrial open reading frame of the 12S rRNA-c; KCNA1, potassium voltage-gated channel subfamily A member 1; GPC1, glypican 1; MMP-9, matrix metalloproteinase-9; VEGFA, vascular endothelial growth factor A; EPHB4, ephrin type-B receptor 4; CYR61, cysteine-rich angiogenic inducer 61; PIK3R2, phosphoinositide-3-kinase regulatory subunit 2; IGF-1, insulin-like growth factor 1; HMOX1, heme oxygenase 1; TRAF6, TNF receptor-associated factor 6; RGS2, regulator of G-protein signaling 2; FOXO3, forkhead box O3; USF2, upstream transcription factor 2; PPP3R1, protein phosphatase 3 regulatory subunit B alpha; HDAC2, histone deacetylase 2; PUMA, p53 upregulated modulator of apoptosis. |
miR-141
Similarly, miR-141 has been reported to be increased in PE-associated placental or trophoblast EV preparations EVs, exacerbates trophoblast dysfunction under hypoxia by targeting CXCL12β and disrupting its signaling through CXCR2/4 receptors, leading to increased apoptosis, reduced invasion-like behavior, and altered angiogenesis-related or extracellular matrix remodeling pathways in experimental models.46 Rescue experiments with arachidonic acid, which counteracts CXCL12β suppression, restore invasion and reduce apoptosis, highlighting a direct mechanistic link.47 Given its hypoxia-specific induction and detectability in maternal circulating EV-enriched fractions, miR-141 qualifies as a biomarker for early PE detection and a therapeutic candidate, as miR-141 inhibition may improve trophoblast migration or invasion-like phenotypes in experimental systems, but whether this translates into improved spiral artery remodeling in vivo remains to be demonstrated.48
miR-146a-5p
miR-146a-5p has been reported to regulate trophoblast proliferation, migration, invasion-like behavior, and EMT-like marker expression by targeting Wnt2 in experimental models.42 Dual-luciferase assays confirm miR-146a-5p binding to the Wnt2 3’UTR in HEK-293T cells, while transfection studies in HTR-8 cells show that miR-146a-5p mimics reduce proliferation (via MTT and colony formation), These assays support a role for miR-146a-5p in regulating trophoblast cell behavior in vitro, rather than directly demonstrating changes in spiral artery remodeling.49 In a broader anti-inflammatory context, PMSC-derived exosomal miR-146a-5p targets TRAF6 and inhibits NF-κB signaling. This process promotes macrophage M2 polarization, as indicated by increased CD206 and Arg1 and reduced TNF-α, and alleviates hypoxia-induced trophoblast apoptosis in vitro. In L-NAME-induced PE mouse models, exosome injection reduced hypertension and proteinuria and improved fetal outcomes.18 Although its expression pattern varies across studies, with upregulation in some PE placentas and downregulation in others, the exosomal stability of miR-146a-5p and its regulatory roles in invasion, inflammation, and hypoxia responses support its potential as a candidate biomarker for monitoring PE severity. It may also represent an experimental therapeutic target for engineered EVs or EV-mimetic nanocarriers designed to integrate diagnostic and therapeutic functions in a theranostic strategy.45 Collectively, studies of these miRNAs provide useful insights into the diagnosis and treatment of the core molecular pathology of PE through placental EV-associated ncRNAs. These findings may support future monitoring based on liquid biopsy and interventions mediated by nanomaterials.50
lncRNAs Associated with Placental/Trophoblast EV Preparations
Long non-coding RNAs associated with placental or trophoblast-derived EV preparations may regulate trophoblast proliferation, migration, invasion-like behavior, endothelial responses, and angiogenesis-related signaling through lncRNA–miRNA–mRNA competing endogenous RNA axes, thereby influencing cellular phenotypes that are relevant to placental development and may indirectly relate to vascular adaptation in vivo.18,20 For example, Fu et al reported that the EV-associated lncRNA ENST00000559730 was increased in maternal plasma EV-enriched fractions from patients with early-onset PE and could act as a sponge for hsa-miR-661, thereby derepressing nudix hydrolase 16 (NUDT16), a key modulator of mRNA decapping and stability, and enhancing PI3K–Akt pathway activity associated with endothelial dysfunction and impaired trophoblast migration.51 In hypoxic HTR-8/SVneo cells, ENST00000559730 manipulation was associated with altered Akt phosphorylation and eNOS/NO-related signaling, reduced invasion in Transwell assays, and impaired tube formation in Matrigel-based angiogenesis-like assays. These observations indicate effects on trophoblast and endothelial-like experimental phenotypes, but they do not directly demonstrate defective spiral artery remodeling.51 Similarly, for MEG3, functional experiments were performed mainly in JEG-3 cells, a choriocarcinoma-derived trophoblast-like cell line. Therefore, reduced viability, wound-healing migration, and EMT-like marker changes in this model support a possible role in trophoblast-like cell behavior, but extrapolation to primary extravillous trophoblasts or human PE pathogenesis requires caution.52 For MALAT1, studies using primary trophoblasts exposed to PE-derived EV preparations provide more physiologically relevant in vitro evidence than tumor-derived cell lines. However, these experiments still represent controlled ex vivo/in vitro systems and should be interpreted as supporting functional involvement in oxidative stress and cell-cycle regulation, not as definitive evidence of disease causality in human PE.53,54 These exosomal lncRNA driven networks suggest a layered regulatory cascade that may amplify the hypoxic microenvironment in PE. They also support the potential of lncRNAs as stable candidate biomarkers for early detection through liquid biopsy and as experimental targets for nanoengineered antagomirs aimed at restoring trophoblast homeostasis.55
circRNAs Associated with Placental/Trophoblast EV Preparations
Although research on circRNAs associated with placental or maternal circulating EV preparations in PE remains in its early stages, their covalently closed-loop structure endows them with exceptional stability against RNase degradation, rendering them highly promising components of maternal circulating EVs and ideal candidates for liquid biopsy.18 Recent high-throughput sequencing of umbilical cord blood EVs or small EV-enriched fractions EVs from PE patients has identified 332 upregulated and 515 differentially expressed circRNAs. Many of these circRNAs are predicted to participate in ceRNA networks related to trophoblast invasion-like behavior, extracellular matrix remodeling, inflammatory signaling, and angiogenesis-related pathways.56
A representative example is EV-associated circDNAJB6, which originates primarily from decidual macrophages but is enriched in the placental microenvironment; it has been reported to be significantly upregulated in maternal plasma EVs or plasma EV-enriched fractions from patients with PE and may contribute to disease progression by sequestering miR-670-5p, thereby derepressing TOB2 and activating the PPARγ/NF-κB inflammatory axis.18 These findings support a role for circDNAJB6 in regulating trophoblast proliferation, invasion-like behavior, apoptosis, and inflammatory signaling in experimental systems. Whether these changes contribute to defective spiral artery remodeling in vivo remains an important hypothesis requiring direct vascular evidence.18 For instance, hsa_circ_0008726 suppresses trophoblast migration, invasion-like behavior, and EMT-like marker changes in JEG-3 and HTR-8/SVneo models by sequestering miR-345-3p and increasing RYBP expression. These cellular phenotypes are relevant to extravillous trophoblast biology but should not be presented as direct evidence of impaired spiral artery remodeling.57
These findings indicate that circRNAs do not act in isolation but form interconnected non-coding RNA networks with EV-associated miRNAs and lncRNAs. These networks may amplify dysregulation of hypoxia responses, inflammation, and vascular remodeling pathways in PE. The high stability, placenta enriched expression profiles, and integration of exosomal circRNAs into multilayered ceRNA networks support their potential as candidate molecules for future multiomics diagnostic panels. They may also represent targets requiring further validation for engineered nanovesicle based theranostic strategies in PE.55
EV-Associated ncRNAs and Maternal–Fetal Immune Imbalance
During PE development, placental ischemia, hypoxia, and oxidative stress may impair trophoblast invasion-like behavior and disrupt maternal–fetal immune and endothelial communication. These alterations are associated with abnormal placentation and vascular maladaptation, but direct evidence of defective spiral artery remodeling requires vascular or placental-bed assessment rather than trophoblast functional assays alone (Figure 3).58 Recent studies have shown that placenta derived EVs are important carriers of molecular communication at the maternal fetal interface. These EVs are continuously released during pregnancy and enter the maternal circulation, where they convert changes in the placental microenvironment into signals that can be detected by the maternal immune system.59 Notably, EV associated non-coding RNAs (ncRNAs), especially miRNAs, lncRNAs, and circRNAs, may amplify inflammation and disrupt maternal fetal immune balance by regulating immune cell recruitment, inflammatory cytokine expression, and immune tolerance related pathways. Therefore, this section reviews the key roles of EV associated ncRNAs in immune and inflammatory abnormalities in PE. It also explains how these ncRNAs may mediate maternal fetal immune imbalance and explores their potential translational value for early prediction and intervention.
 |
Figure 3 Placenta-derived extracellular vesicles in PE-related immune imbalance, vascular dysfunction, and translational opportunities. This schematic summarizes the potential roles of placenta-derived extracellular vesicles (PEVs) in PE. In PE, placental stress, including hypoxia and inflammatory injury, may alter the abundance, size distribution, surface molecules, lipid composition, protein cargo, and ncRNA content of PEVs. The central blue box indicates PEVs in PE as the core element of the schematic. Blue arrows indicate factors or conditions associated with altered PEV release and cargo composition, including increased and enlarged PEVs in PE, distinct profiles between early-onset and late-onset PE, hypoxia-related changes, and altered lipids, surface markers, proteins, and miRNAs. Purple arrows indicate downstream PE-related biological processes associated with PEVs, including vascular dysfunction, impaired angiogenesis, endothelial activation or injury, abnormal placentation, altered trophoblast behavior, aberrant immune responses, increased chemotaxis, NET formation, cytokine dysregulation, and clinical PE manifestations. The red/yellow burst symbols in the placenta indicate placental stress or injury. The yellow highlighted box at the bottom represents potential translational opportunities, including exploratory therapeutic targeting and differential diagnosis, screening, or early prediction of PE. Yellow arrows indicate possible translational directions derived from PEV research. Bold text identifies major functional modules or conceptual categories in the schematic. No black upward or downward arrows are used in this figure. These pathways should be interpreted as a conceptual summary; well-supported PE findings, such as altered PEV profiles, immune activation, endothelial dysfunction, and NET-related inflammation, should be distinguished from proposed EV-ncRNA-mediated mechanisms that still require further experimental and clinical validation. Adapted from Ortega, Miguel A et al Unfolding the role of placental-derived Extracellular Vesicles in Pregnancy: From homeostasis to pathophysiology. Frontiers in cell and developmental biology. Copyright © 2022 by authors.38 Abbreviations: PE, preeclampsia; PEVs, placenta-derived extracellular vesicles; EVs, extracellular vesicles; ncRNA, non-coding RNA; miRNA, microRNA; NETs, neutrophil extracellular traps. |
EV-Associated miRNAs and Macrophage/NK/Treg Function
In PE, the imbalance of maternal fetal interface and systemic immune tolerance is one of the core pathological features of the disease.3 Current evidence suggests that EV-associated miRNAs may influence macrophage polarization, NK-cell activation, and T-cell differentiation in several experimental systems. However, these systems differ substantially in physiological relevance. Primary human decidual macrophages and peripheral blood immune cells provide more direct human immune-cell information, whereas RAW264.7, THP-1, and Jurkat cells are transformed or species-specific models that require validation in primary decidual or peripheral immune cells.3,60 Rodent pregnancy models can test systemic immune and vascular consequences, but they should not be considered direct equivalents of human maternal–fetal immune tolerance.61 For instance, miR-494, overexpressed in decidual mesenchymal stem cell-derived EVs/small EVs EVs from PE patients, inhibits M2 macrophage polarization by suppressing prostaglandin E2 secretion, thereby promoting a pro-inflammatory M1 phenotype that exacerbates placental dysfunction, as demonstrated in in vitro co-culture experiments with human decidual macrophages where miR-494 mimic transfection reduced M2 markers like CD206 and arginase-1 while elevating M1-associated TNF-α and IL-6 levels.18 Similarly, EV-associated miR-21 from preeclamptic placentas shifts macrophages toward an M1 state, increasing pro-inflammatory cytokines such as IL-1β and IL-6, which was observed in primary macrophage cultures exposed to isolated EVs, leading to enhanced nuclear factor-κB signaling and reduced anti-inflammatory activity.62 Regarding natural killer (NK) cells, EV-miRNAs in PE modulate NK activity by inhibiting cytotoxicity; specifically, placenta-derived EVs or trophoblast-associated EV preparations carrying miR-517a EVs activate immune responses and disrupt Th1/Th2 balance, resulting in heightened NK activation and maternal immune intolerance, as evidenced by flow cytometry assays showing increased NK degranulation (CD107a expression) in peripheral blood NK cells incubated with EV preparations isolated from PE samples EVs preparations from PE samples compared to controls.63 This mechanism was further validated in animal models, where injection of miR-517a-enriched EVs into pregnant mice induced PE-like symptoms, including hypertension and proteinuria, alongside elevated NK infiltration at the maternal-fetal interface.63 For regulatory T cells (Treg), EV-miRNAs contribute to immune dysregulation by altering Treg differentiation and function. In Jurkat T-cell models, EV-mediated transfer of miR-519d-3p has been associated with changes in FOXP3-related markers and Th17/Treg-associated cytokine profiles.64 Because Jurkat cells are leukemia-derived T cells and do not fully recapitulate primary regulatory T cells or decidual T-cell biology, these findings should be considered hypothesis-generating and require validation in primary maternal immune cells.64,65 Overall, available studies support the hypothesis that EV-associated miRNAs may contribute to immune imbalance in PE, but the degree to which these pathways operate in human decidual immune networks remains to be established through primary-cell studies, spatial placental analysis, and longitudinal clinical cohorts.
EV-Associated lncRNAs/circRNAs and Inflammatory Pathways
In PE, exosomal long non-coding RNAs (lncRNAs) and circular RNAs (circRNAs) may contribute to inflammatory cascades and disrupt maternal fetal immune homeostasis. They can act by sponging microRNAs, modulating transcription factors, and amplifying proinflammatory signaling. These functions support their mechanistic relevance in PE pathogenesis and suggest their potential as candidate biomarkers and experimental therapeutic targets in theranostic strategies.66 Specifically, MALAT1 is upregulated in maternal plasma EV enriched fractions from patients with PE. Exosomal MALAT1 may promote vascular endothelial inflammation by competitively binding miR-150-5p and increasing vascular endothelial growth factor A (VEGFA) expression. This process may activate NF-κB signaling and increase proinflammatory cytokines, including TNF-α and IL-6. In vitro endothelial cell models showed that MALAT1 enriched EVs increased NF-κB p65 nuclear translocation by 1.8 fold and cytokine secretion by twofold compared with controls.23 Similarly, lncRNA NEAT1 in trophoblast derived EVs may exacerbate macrophage mediated inflammation in PE by stabilizing TLR4 mRNA. This effect promotes M1 polarization and IL-1β release. In RAW264.7 macrophages co-cultured with EV preparations from PE samples, TLR4 protein levels increased by 2.5 fold, accompanied by NLRP3 inflammasome activation. These findings were further supported in lipopolysaccharide induced PE mouse models, which showed increased placental inflammation and hypertension.67 Among circRNAs, exosomal circDNAJB6 derived from decidual macrophages may promote PE progression through the miR-670-5p and TOB2 axis. circDNAJB6 can sponge miR-670-5p, increase TOB2 expression, suppress PPARγ activity, and activate NF-κB signaling, thereby amplifying inflammatory responses, including increased IL-6 and TNF-α. In HTR-8/SVneo trophoblast cells, EV mediated transfer of circDNAJB6 reduced PPARγ activity by 60% and increased NF-κB phosphorylation. In pregnant rats, tail vein injection of circDNAJB6 loaded EVs induced PE like symptoms, including proteinuria, fetal growth restriction, and elevated serum inflammatory markers.18 Similarly, in endothelial cell models such as HUVECs, EV-associated ncRNAs including MALAT1 or circ_0001438 have been linked to NF-κB activation, EPAS1 expression, and increased inflammatory cytokine production. However, HUVECs represent fetal large-vessel endothelial cells and do not fully reproduce maternal uterine, placental-bed, or systemic microvascular endothelium in PE.68 Furthermore, circ_0004906 in circulating EV fractions from PE patients has been linked to miR-193a-3p regulation and JAK/STAT pathway activation, which may enhance monocyte chemotaxis and pro-inflammatory cytokine production. Experimental validation in THP-1 monocyte models showed a twofold increase in p-STAT3 levels after exosome uptake. Animal studies further showed that exosomal circ_0004906 exacerbated placental leukocyte infiltration and systemic inflammation.69,70 These exosomal lncRNAs and circRNAs may contribute to a proinflammatory microenvironment in PE by regulating ceRNA networks and signaling hubs. These findings provide mechanistic insights into the link between immune imbalance and clinical manifestations and may support future nanoparticle based interventions.68
EV-Associated ncRNAs, Angiogenic Signaling, Endothelial Dysfunction, and Innate Immune Vascular Injury
EV-Associated miRNAs and AKT/HIF-1α/VEGF-Related Angiogenic Signaling
Angiogenic imbalance is a central feature of PE, but the relationship among AKT, HIF-1α, VEGF-A, PlGF, and sFlt-1 is context-dependent and should not be described as a simple linear pathway. HIF-1α generally functions as a hypoxia-responsive transcription factor that can induce angiogenic and anti-angiogenic mediators, whereas impaired AKT signaling, altered VEGF bioavailability, and increased sFlt-1 may together contribute to endothelial dysfunction in PE.71,72 Therefore, EV-associated ncRNAs should be discussed as regulators of angiogenesis-related signaling rather than as direct determinants of a single AKT/HIF-1α/VEGF cascade.66 Maternal plasma EV-associated miR-210, or miR-210 detected in small EV-enriched fractions, has been reported to be altered in severe PE cohorts. In HUVEC-based endothelial models, exposure to PE-derived EV preparations or experimental miR-210 enrichment has been associated with changes in AKT phosphorylation, HIF-1α signaling, VEGF-A secretion, and endothelial tube-formation capacity. These findings suggest that EV-associated miR-210 may participate in angiogenesis-related endothelial dysfunction under PE-associated stress conditions. However, they should not be interpreted as evidence that HIF-1α stabilization directly reduces VEGF expression or that miR-210 is sufficient to cause human PE vascular disease.73 In rodent pregnancy models, administration of miR-210-enriched EV preparations has been associated with PE-like readouts, including increased blood pressure and altered placental vascular markers. These findings provide model-based support for mechanistic plausibility, but rodent PE-like phenotypes do not fully reproduce human PE angiogenic pathology.74 Placenta-associated EV miR-15a-5p has been linked to CDK1 and PI3K/AKT-related signaling in trophoblast-like models. In HTR-8/SVneo cells, miR-15a-5p manipulation has been associated with altered AKT activity, VEGF-related transcript levels, and apoptosis-related phenotypes. Because HTR-8/SVneo cells are trophoblast-like cells rather than endothelial or vascular relaxation models, these findings should be interpreted as trophoblast signaling data and should not be used as direct evidence for endothelial VEGF bioavailability or maternal vascular dysfunction.75 In hypoxic HUVECs, miR-146a-5p-containing EV preparations have been associated with increased NO production and reduced ET-1-related responses. These endothelial-cell findings suggest a possible vascular mechanism, whereas L-NAME-induced mouse data provide model-based in vivo support. Neither model alone establishes therapeutic efficacy or vascular normalization in human PE.76 Additionally, exosomal miR-125a-5p in PE may inhibit trophoblast migration by downregulating VEGFA through AKT suppression. In wound healing assays, EVs from PE samples reduced trophoblast migration by 35%. Luciferase reporter assays targeting the VEGFA 3′ UTR further linked this effect to reduced AKT and HIF1α activation.77 Regulation of the AKT, HIF1α, and VEGF axis by EV associated miRNAs may help clarify the endothelial pathology of PE. It may also have translational implications for liquid biopsy based early detection and targeted nanomedicine delivery.78
EV-Associated ncRNAs and RAS/NO/Endothelin Signaling
In PE, exosomal ncRNAs may regulate vascular pathways involved in endothelial dysfunction and hypertension. These pathways include the renin angiotensin system (RAS), nitric oxide (NO) bioavailability, and endothelin signaling. These findings may clarify key pathogenic mechanisms and support the potential of EV associated ncRNAs as candidate biomarkers for liquid biopsy based early detection and as experimental targets for nanomedicine based interventions.79 For example, maternal plasma EV associated miR-210 from patients with PE was identified through miRNA profiling. It may suppress endothelial NO synthase (eNOS) expression by targeting the eNOS 3′ untranslated region, leading to reduced NO production and impaired vasodilation. In human umbilical vein endothelial cells (HUVECs), EV preparations from PE samples reduced NO levels by 50% in the Griess assay and decreased eNOS activity. In pregnant mice, administration of these EV preparations exacerbated hypertension by impairing NO dependent vascular relaxation.73 Similarly, exosomal miR-155 from preeclamptic placentas may enhance RAS activation by inhibiting suppressor of cytokine signaling 1 (SOCS1). This effect may increase angiotensin II (Ang II) sensitivity and vascular inflammation. In trophoblast cell lines, qPCR analysis showed miR-155 upregulation, which was associated with a twofold increase in Ang II type 1 receptor (AT1R) expression and reactive oxygen species (ROS) production. In rat models, exosome injection induced PE like symptoms, including proteinuria.80 In HTR-8/SVneo cells, H19-enriched EV preparations have been associated with AT1R and ET-1-related changes. Because this is a trophoblast-like model, the result should be interpreted as placental-cell signaling evidence rather than direct evidence of maternal vascular RAS activation.23 In a related mechanism, placental exosomal circ_0001438 may promote endothelial dysfunction by sequestering miR-942-5p and upregulating the NLRP3 inflammasome. This process may reduce NO bioavailability and enhance RAS and endothelin 1 (ET-1) signaling. Cell based studies showed a 60% reduction in NO metabolites in HUVECs after exosome exposure, accompanied by NLRP3 activation detected by Western blotting. In pregnant rats, circ_0001438 delivery increased blood pressure and vascular permeability.81 Additionally, placenta associated EV miR-15a-5p from PE placentas may target CDK1 and inhibit PI3K/AKT signaling. This effect may reduce eNOS phosphorylation and NO synthesis while enhancing Ang II related effects. In trophoblast models, miR-15a-5p mimics reduced AKT activity by 40% and decreased eNOS levels, as quantified by immunoblotting. These changes were associated with RAS hyperactivity in clinical samples.7.79 Amniotic fluid EV associated miR-146a-5p may counteract PE progression by repressing HIF-1α, restoring NO bioavailability, and mitigating ET-1 induced vasoconstriction. In hypoxic HUVEC cultures, delivery of this miRNA increased NO levels by 1.8 fold and attenuated ET-1 responses. In L-NAME induced PE mice, exosome treatment improved vascular function.76 Among lncRNAs, exosomal NEAT1 in PE may exacerbate vascular inflammation by stabilizing TLR4. This effect may reduce NO levels and enhance RAS and ET-1 signaling. Macrophage co-culture experiments showed a twofold increase in TLR4 expression and a decline in NO levels. Rodent models further showed increased endothelin receptor expression.82,83 Moreover, exosomal lncRNA MALAT1 from patients with PE may bind miR-150-5p and increase ET-1 transcription, thereby impairing endothelial NO production. In vitro endothelial assays showed that MALAT1 enriched EVs increased ET-1 levels by 70% and reduced NO production. PE cohorts also showed associated RAS dysregulation.84 Collectively, these exosomal ncRNA driven interactions include miRNA mediated eNOS repression and lncRNA or circRNA mediated enhancement of RAS and ET-1 signaling. Together, they provide a multifaceted framework for vascular pathology in PE and may support future nanoparticle based diagnostic and therapeutic strategies85
Complement/NET Abnormalities in PE and the Proposed EV-ncRNA–Complement/NET Axis
Complement activation and NET formation are well-described features of PE-associated vascular inflammation,86 but the type of evidence varies across studies.87,88 Clinical biomarker studies have reported increased circulating complement activation fragments such as C3a, C5a, or soluble terminal complement complex in subsets of PE patients. Placental staining studies have described increased deposition of terminal complement components, including C5b-9, in placental tissues. Genetic studies have linked variants in complement-regulatory genes with susceptibility to severe PE or HELLP-like phenotypes in selected populations.89 In parallel, NET-related markers have been detected in maternal plasma, decidua, or placental tissues, and functional assays have shown that serum from PE patients can promote NET formation in neutrophils from healthy donors. These findings support complement and NET involvement in PE-associated inflammation and endothelial injury, but they do not by themselves identify EV-associated ncRNAs as upstream regulator.90,91 Some experimental studies have implicated EV preparations from PE samples in endothelial activation, oxidative stress, VCAM-1 or vWF expression, and interactions with NET-related endothelial injury. These data suggest that EVs may participate in vascular inflammation in PE. However, many of these studies focused on total EVs, surface proteins, lipids, or mixed vesicle cargo rather than specifically isolating ncRNA-dependent mechanisms.92 Therefore, EV effects on complement deposition or NET-associated endothelial injury should be distinguished from EV-ncRNA-specific regulation.93–95
At the RNA level, EV-associated miRNAs and other ncRNAs have emerged as important regulators of immunovascular pathways related to complement and NET biology. Placental and circulating miR-155, miR-210, miR-181a-5p, the miR-17/20 cluster, and several lncRNAs are consistently dysregulated in PE and enriched within placenta-derived EVs. These ncRNAs target transcripts involved in endothelial nitric oxide synthase (eNOS) signalling, NF-κB activation, TLR pathways, and inflammasome components.45,96,97 Morales-Prieto et al reported that placenta-derived EVs are enriched in immunomodulatory miRNAs that regulate T cell and macrophage polarisation. In PE, increased ncRNAs such as miR-155 and miR-146a may shift the balance toward Th1 and Th17 responses rather than Th2 and Treg responses, thereby promoting a proinflammatory state that favours complement activation and neutrophil priming.96 Experimental studies in other inflammatory settings, summarized in reviews by O’Brien and Lv, show that EV-miRNAs can directly repress complement regulators, such as factor H, or components of the TLR and NF-κB axis. In this way, EV-miRNAs may regulate complement activation and NETosis thresholds in recipient neutrophils and endothelial cells. These findings suggest that similar regulatory circuits may operate in pregnancy-specific vascular beds.98,99 In PE, integrative omics analyses of maternal plasma have identified circulating miRNA signatures associated with early complement activation and innate immune activation. Several of these transcripts, including miR-155, miR-210, and miR-223, have been detected within placenta-derived EV fractions. These findings support the idea that EV-associated ncRNAs may transmit signals that prime maternal leukocytes for inflammasome and complement activation.100,101 Complement fragments such as C3a and C5a are potent NET inducers. Hernández González et al emphasized that NET formation in PE is driven by converging cytokine, DAMP, and complement signals. When this environment is combined with EV cargo enriched in proinflammatory ncRNAs that sustain ROS production and NF-κB activation, neutrophils may shift toward a NET-prone phenotype. This phenotype can deposit DNA and histone scaffolds that support further C3 and C5 convertase assembly, thereby reinforcing a pathogenic cycle between complement activation and NET formation on the endothelial surface.90,102 The proposed EV-ncRNA–complement/NET axis remains largely hypothesis-generating. Several PE-associated EV-ncRNAs, including miR-155, miR-146a, miR-210, and miR-223, are biologically plausible candidates because they have been linked to TLR–NF-κB signaling, ROS production, inflammasome activation, endothelial dysfunction, or immune-cell activation in PE or other inflammatory contexts.62,103 However, direct evidence that PE-derived EV-associated ncRNAs regulate complement components, complement regulators, NETosis thresholds, or complement–NET feedback loops in human pregnancy remains limited.68,99,104 Future studies should test this axis using purified and well-characterized EV populations, EV-RNA depletion or rescue experiments, primary neutrophils and endothelial cells, complement functional assays, NETosis assays, and paired clinical samples with complement/NET biomarkers.98,105
Extracellular Vesicle ncRNA as a Nanoscale Biomarker for PE: From Discovery to Validation
Sampling Matrices, Putative Cellular Origins, and EV Subtypes
When evaluating EV-associated ncRNAs as nanoscale biomarkers in PE, it is essential to distinguish the biological origin of vesicles from the biofluid or tissue matrix used for sampling. Placental tissue, placental perfusate, trophoblast culture systems, maternal plasma or serum, urine, and amniotic fluid provide different levels of biological proximity to the maternal–fetal interface.47,106 However, detection of an ncRNA in plasma, urine, or amniotic fluid EV fractions does not by itself prove placental, trophoblast, endothelial, renal, immune-cell, or fetal origin. Source attribution requires additional evidence, including tissue-paired profiling, placental perfusion models, cell-specific EV markers, immunocapture, or single-vesicle analysis. Placenta-derived EVs or trophoblast-derived EVs (Table 1). For example, Pillay et al systematically proposed that miRNA profiles carried by placenta-derived or trophoblast-derived EVs are closely associated with PE, providing a tissue reference for subsequent liquid biopsy studies.107,108 On this basis, Awoyemi et al used a dual lobe perfusion model to isolate medium and large STB EVs and identified significant dysregulation of hsa-miR-193b-5p, miR-324-5p, miR-652-3p, miR-3196, miR-9-5p, miR-421, and the classical hypoxia related miR-210-3p in PE placental m/l-STB EVs. The elevation of miR-9-5p was also observed in maternal serum EV fractions, suggesting that placental EV miRNA profiles may be reflected in peripheral blood. This finding supports their potential role in linking exploratory placental samples with subsequent liquid biopsy markers.109 Aharon et al further analyzed miRNAs in parallel in maternal plasma and placental EVs and showed that some placenta enriched exomiRNAs, such as C19MC and C14MC cluster members, exhibit consistent directional changes in the maternal circulation during pregnancy induced hypertension, PE, and other placental vascular complications. This provides systematic evidence for future studies that pair placental EV profiles with maternal circulating EV signals in PE.110 In addition to miRNAs, lncRNAs such as H19 and MALAT1, as well as circRNAs related to angiogenesis and immune regulation, have also been detected in placental and trophoblast derived EVs. These long and circular ncRNAs are often associated with trophoblast invasion, epithelial mesenchymal transition, and inflammatory response phenotypes. Although current research remains largely mechanistic, these findings suggest that placental EV lncRNAs and circRNAs may provide an important source of tissue specific long RNA marker libraries for subsequent blood or amniotic fluid biopsy studies.69
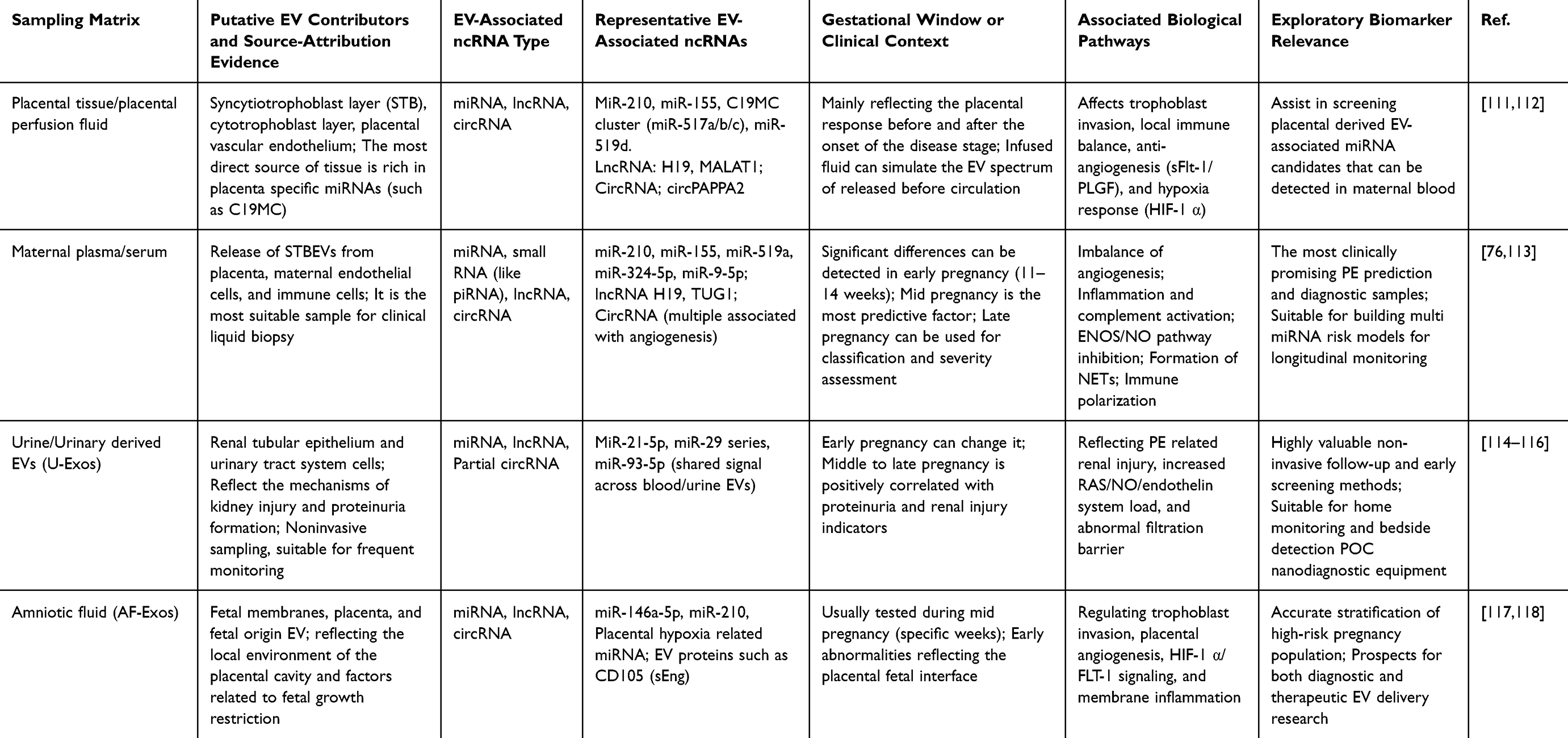 |
Table 1 Sampling Matrices, Putative EV Contributors, EV-Associated ncRNAs, and Biomarker Potential in PE |
Maternal peripheral blood remains the main clinical matrix for nanoscale liquid biopsy based on EV-associated ncRNAs. Early work by Pillay et al showed that NanoString based EV-associated miRNA profiling could distinguish early-onset PE from late-onset PE and reveal subtype-related differences in angiogenic, immune, and metabolic pathways. This work provided early methodological evidence that EV-associated miRNA profiles could be explored for PE biomarker discovery.107 Furthermore, Ghosh et al collected maternal plasma EV-enriched fractions during the first and second trimesters in a prospective cohort that included 14 women who later developed PE and 12 women with normal pregnancies. Using small RNA sequencing, they identified 148 differentially abundant EV-associated miRNAs in the first trimester, among which C19MC and C14MC cluster members showed marked gestational stage related changes. They constructed a combined EV-miRNA prediction model for the first and second trimesters using elastic-net regularized logistic regression. This model achieved an AUC of 0.956 in the preliminary cohort and predicted PE before clinical symptoms appeared. These findings provide preliminary proof-of-concept evidence for EV-miRNA based early prediction of PE during pregnancy.47 Gál et al further distinguished women with preterm PE and fetal growth restriction from controls based on first trimester plasma exosomal small RNA profiles, including miRNAs and piRNAs. They found that multiple exosomal miRNA combinations related to immune and vascular pathways had predictive value. This finding suggests that combined analysis of multiple small RNA ncRNA classes may outperform single miRNA markers.119 For established disease, Wang et al sequenced and qPCR validated maternal plasma EV-associated miRNAs from patients with early-onset PE and identified 10 candidate exomiRNAs, including miR-365b-3p and miR-765. The combined model achieved an AUC greater than 0.9 for distinguishing EOPE from healthy pregnancy. Multiple miRNAs were correlated with blood pressure levels and the placental sFlt-1/PLGF ratio, indicating potential value for diagnosis and disease course assessment.120 Chen et al focused on PE with severe features and found that a group of maternal plasma EV-associated miRNAs, including miR-210 and miR-155, were associated with endothelial injury, inflammation, disease severity, and laboratory indicators. These findings suggest that different clinical phenotypes may correspond to distinct EV-miRNA combinations. This also provides a basis for future stratification according to PE stage and subtype.73 Although studies of lncRNAs and circRNAs in maternal plasma or serum EV-enriched fractions remain limited, existing evidence suggests that certain exosomal lncRNAs, such as H19 and TUG1, and circRNAs differentially expressed in cord blood EVs are associated with placental hypoxia, angiogenic imbalance, and immune abnormalities. Incorporating these long and circular ncRNAs into liquid biopsy panels may improve model resolution across different PE subtypes and pregnancy stages.23,56,121
Compared with the first two EV sources, urine provides a more downstream perspective because the urinary system and kidneys are important sources of EVs. Urinary EVs retain the stability of classical EVs and offer advantages such as easy collection and suitability for repeated monitoring. Therefore, they are considered promising liquid biopsy matrices in various kidney diseases and may have clinical relevance for PE detection.122,123 Several reviews have emphasized that proteins and RNA in EVs can reflect the structural and functional status of glomeruli and tubules. In many acute and chronic kidney injury models, these EV cargos may provide information beyond traditional indicators such as serum creatinine and urinary protein.123–125 In PE, Illarionov et al collected urine samples during early and mid pregnancy and identified multiple urinary miRNAs, including miR-21-5p and the miR-29 family, that were associated with subsequent PE development using miRNA seq and qPCR. Many of these miRNAs have been linked to trophoblast function and vascular pathways in previous placental and peripheral blood studies. This suggests that some EV associated miRNA signals may be transmitted along the placental renal axis and detected in urine.114 More importantly, a recent multiplatform study analyzed EVs in both plasma and urine from women with PE. It found that several miRNAs, including miR-93-5p, were significantly upregulated in EVs from both biofluids, and their expression levels were positively correlated with blood pressure and proteinuria. This cross fluid consistency provides a useful example for comparing plasma and urine EV miRNA profiles across PE stages. It also suggests that urine based nanodiagnostic approaches may complement blood based testing in future clinical applications.126 Although research on urinary EV associated ncRNAs in PE remains limited, evidence from nephrology suggests that urinary EVs may help reflect PE related kidney injury and integrate information from the renal placental axis. These features support their potential use in wearable or bedside nanodiagnostic approaches, although further clinical validation is required.121,123
Because amniotic fluid is located at the placental fetal interface, amniotic fluid derived EVs may more directly reflect the local placental and fetal microenvironment. However, because sampling is invasive and clinically restricted, current studies mainly focus on high risk populations undergoing amniocentesis during mid pregnancy. From a biological perspective, however, amniotic fluid provides a valuable window for constructing placental and fetal cavity EV ncRNA profiles.127 For example, Gebara et al used imaging flow cytometry and single vesicle surface labeling analysis to show that EV concentrations were significantly increased in amniotic fluid from PE pregnancies. The proportion of VEGFR2 and LAP positive EVs with sEng enrichment was also increased, indicating antiangiogenic and inflammatory phenotypes. These findings provide a phenotypic basis for studying PE related amniotic fluid EVs.128 From the perspective of integrated diagnosis and therapy, Jin et al reported that amniotic fluid EV associated miR-146a-5p was significantly downregulated in PE pregnancy. AF EVs from normal pregnancy, or EVs enriched with miR-146a-5p, restored trophoblast cell migration, invasion, and tube formation in vitro. They also improved blood pressure, placental hypoxia markers, and the sFlt-1/PLGF imbalance in the L-NAME induced PE rat model.76 These findings suggest that AF EV associated miR-146a-5p may serve as a candidate biomarker for risk stratification in high risk populations during mid pregnancy. It may also represent a potential candidate molecule for EV based nanomedicine, consistent with the concept of integrated diagnostic and therapeutic strategies.
Current evidence suggests that EV miRNAs, especially C19MC and C14MC clusters, in the placenta, umbilical cord blood, and placental perfusion fluid are mainly used for exploratory research and correlation with histopathological features and molecular pathways.96 Maternal plasma and serum EVs carry major combinations of miRNAs, lncRNAs, and circRNAs throughout pregnancy, supporting applications from early pregnancy prediction to mid and late pregnancy diagnosis and subtype classification.59,126 Urinary EVs may reflect PE related renal injury and systemic vascular burden and may serve as a sampling matrix for frequent and low cost monitoring. Amniotic fluid EV ncRNAs provide a deeper phenotypic profile close to the placental fetal interface in high risk populations during mid pregnancy and may serve as natural carriers for local EV ncRNA based nanomedicine delivery.129,130 In future nanodiagnostic applications, integrating multisource EV ncRNAs may improve the sensitivity and specificity of PE prediction and subtype classification. This approach may also provide a biological and technical foundation for developing theranostic nanovesicles that combine homologous targeting, diagnostic readout, and drug delivery.
EV-Associated ncRNAs for PE Risk Prediction, Diagnosis, and Stratification
Before discussing EV-associated ncRNAs as potential biomarkers, the intended clinical question should be clearly defined. In PE, early prediction refers to estimating future disease risk before clinical onset, whereas diagnosis refers to identifying PE after established clinical criteria are met. After PE is suspected or diagnosed, biomarkers may further support risk assessment, severity evaluation, and subtype stratification.30,131 Therefore, EV-associated ncRNAs detected in maternal plasma or serum, urine, amniotic fluid, or placental and trophoblast associated EV preparations should be evaluated according to their intended clinical use rather than grouped into a single diagnostic category. Their performance should also be compared with established clinical, biophysical, and angiogenic tools, including maternal risk factors, mean arterial pressure, uterine artery Doppler indices, PlGF, the sFlt-1 and PlGF ratio, blood pressure, proteinuria, clinical symptoms, and routine laboratory indicators.4,132 Figure 4 summarizes this integrated workflow and illustrates how EV-associated ncRNAs may be positioned as complementary exploratory biomarkers for PE prediction, diagnosis, and stratification.
 |
Figure 4 Integration of PE biology and liquid-biopsy tools for EV-associated ncRNA-based risk prediction, diagnosis, and stratification. This schematic summarizes the clinical manifestations, placental biological alterations, and liquid-biopsy approaches relevant to PE assessment. (A) PE commonly presents after 20 weeks of gestation with clinical symptoms and signs such as hypertension, proteinuria, edema or swelling, excessive weight gain, headache, and visual disturbance. These manifestations support diagnosis of established disease together with maternal organ dysfunction, fetal assessment, and laboratory findings. (B) Placentas from PE pregnancies show multiple cellular and molecular abnormalities, including impaired trophoblast invasion and differentiation, increased pro-inflammatory and oxidative-stress responses, altered proliferation and senescence of placental mesenchymal/stromal cells, immune-cell imbalance involving Hofbauer cells, Tregs, macrophage polarization, neutrophil activation, and cytotoxic dNK-cell activity, as well as dysregulated angiogenic, inflammatory, and placental factors such as increased sFLT-1, sENG, PAPPA2, and DAMPs and decreased PAPP-A, hCG, PLGF, HLA-G, syncytin-1, VEGF, and placental steroid-related signals. (C) Liquid-biopsy techniques provide minimally invasive approaches to capture PE-associated maternal–fetal molecular information. Circulating fetal nucleated red blood cells, single circulating trophoblasts, cfDNA, cfRNA, and EVs or small EV/exosome-enriched fractions can be analyzed by FISH, cell imaging, PCR-based assays, DNA sequencing, RNA sequencing, and epigenetic profiling. Adapted from Ma Y, Chiang YW, Becker TM, Hyett J. Cell-Based and Cell-Free Non-Invasive Prenatal Analysis of Preeclampsia: An Updated Review of Liquid Biopsy. Biomedicines. Copyright © 2026 by authors.133 Abbreviations: PE, preeclampsia; EVs, extracellular vesicles; ncRNAs, non-coding RNAs; Tregs, regulatory T cells; dNK cells, decidual natural killer cells; sFLT-1, soluble fms-like tyrosine kinase-1; sENG, soluble endoglin; PAPPA2, pregnancy-associated plasma protein-A2; DAMPs, damage-associated molecular patterns; PAPP-A, pregnancy-associated plasma protein-A; hCG, human chorionic gonadotropin; PLGF, placental growth factor; HLA-G, human leukocyte antigen-G; VEGF, vascular endothelial growth factor; fNRBCs, fetal nucleated red blood cells; SCTs, single circulating trophoblasts; cfDNA, cell-free DNA; cfRNA, cell-free RNA; FISH, fluorescence in situ hybridization; PCR, polymerase chain reaction. |
Early Risk Prediction Before Clinical Onset
Early prediction and clinical diagnosis should be treated as distinct clinical questions. Diagnosis refers to identifying PE after clinical criteria have been met, usually after 20 weeks of gestation, whereas prediction refers to estimating the risk of future PE before symptoms or diagnostic criteria appear. In this review, early prediction refers to EV-associated ncRNA measurement before clinical onset, particularly before 20 weeks of gestation. These biomarkers should therefore be evaluated by longitudinal sampling and by their ability to improve risk models that already include maternal risk factors, mean arterial pressure, uterine artery Doppler indices, PlGF, and other angiogenic markers. These molecules have the advantages of high stability and strong tissue specificity, especially in liquid biopsy and non-invasive detection, providing candidate biomarkers for further validation rather than currently reliable clinical toolsfor the entire PE segment (Table 2).47,130 Early prediction of PE is crucial for improving maternal and infant health. The pathogenesis of PE is complex, involving placental dysplasia, immune response imbalance, and abnormal angiogenesis. The existing prediction methods often only diagnose after clinical symptoms appear, which misses the opportunity for early intervention. EVs provide a new pathway for early pregnancy warning by carrying miRNAs, lncRNAs, and circRNAs from various sources such as placenta, maternal blood, and urine.108,134 At present, many studies have shown that specific miRNAs can serve as biomarkers for predicting PE in early pregnancy (<20 weeks). miR-210 or miR-210-3p has been investigated as a candidate marker of hypoxia-associated placental stress in PE. However, its use for early prediction requires confirmation in longitudinal cohorts sampled before disease onset, standardized EV isolation, arm-specific miRNA reporting, and comparison with established screening tools.119,135 Through liquid biopsy technology, researchers have explored whether these markers can improve early risk assessment in preliminary cohorts by detecting miRNAs such as miR-210 in plasma. For example, Ghosh et al used maternal plasma EV-associated miRNAs during early pregnancy before 14 weeks of gestation to construct a model for predicting PE before clinical symptoms appear. The model achieved an AUC of up to 0.96.47 In addition to miRNAs, lncRNAs and circRNAs have increasingly been implicated in the early prediction of PE. For example, H19 is an lncRNA closely related to placental development and angiogenesis, and changes in H19 expression during early pregnancy may reflect abnormal placental function.119 In addition, circRNAs such as circPAPPA2 have been associated with placental hypoxia and may regulate angiogenic pathways by acting as miRNA sponges. These findings further support the potential of circRNAs for early PE prediction.73 For clinical translation, liquid biopsy offers a minimally invasive approach for dynamic monitoring and shows potential for early PE prediction. Small RNAs such as miRNAs, lncRNAs, and circRNAs can remain stable in body fluids, including plasma, urine, and amniotic fluid, when carried by EVs. This stability supports their potential use as accessible biomarkers.45 A recent study showed that combined analysis of EV-associated miRNAs and lncRNAs in maternal plasma may identify pregnancies at high risk of PE before clinical onset, with favorable sensitivity and specificity.119 In addition, urinary EV-associated miRNAs have shown PE-related changes during early pregnancy and may be suitable for future bedside or home monitoring.128 With technological advances and larger validation cohorts, EV-associated ncRNAs may become useful tools for early PE prediction, diagnosis, and precision intervention.
 |
Table 2 Summary of Different Exosomatic miRNA at PE Stages |
Assessment of Established PE and Risk Stratification After Clinical Presentation
After clinical presentation, the relevant question changes from prediction to diagnosis, severity assessment, and risk stratification.138 At this stage, EV-associated ncRNAs may be explored as molecular indicators of disease subtype, severity, angiogenic imbalance, renal involvement, endothelial dysfunction, or inflammatory burden. However, their diagnostic role remains unvalidated and should be considered complementary to established clinical criteria, symptoms, blood pressure, proteinuria, platelet count, liver enzymes, renal function, PlGF, the sFlt-1/PlGF ratio, and fetal/placental assessment.139 Currently, evaluating extracellular vesicle ncRNA levels in the plasma, serum, or other body fluids of pregnant women during the middle and late stages of pregnancy can provide a non-invasive, real-time liquid biopsy tool. This tool reflects the status of the placental or maternal-placental interface.140 This approach is expected to enable early warning and stratified management of PE, thereby improving maternal and fetal outcomes. In recent years, numerous studies have investigated this area, along with the underlying mechanisms and their clinical translational potential. For example, a recent study employed high-throughput sequencing (NGS) to analyze the differential miRNA expression profiles in peripheral blood exosomal EVs from women with early-onset PE compared with a control group and identified several significantly differentially expressed miRNAs. These differentially expressed miRNAs provide a molecular candidate pool for the future development of diagnostic or predictive models based on extracellular vesicle miRNAs.120 Furthermore, a recent systematic review indicates that placenta-derived or circulating extracellular vesicle miRNAs, such as miR-210 and miR-155, are significantly upregulated in patients with PE. These changes are closely associated with the inflammatory response, endothelial dysfunction, hypoxic response, and abnormal trophoblast behaviors, including migration, invasion, and apoptosis.106 The stability of these miRNAs, including their resistance to RNase, pH variations, and temperature changes, makes them highly suitable for liquid biopsy applications in plasma, serum, or exosomes.45 In addition to miRNAs, researchers have increasingly focused on exosome-associated lncRNAs. For instance, a recent review of pathological pregnancies, including PE, summarized that extracellular vesicle lncRNAs represent promising biomarkers due to their selective loading, high stability in circulation, and capacity to regulate gene expression via epigenetic and post-transcriptional mechanisms.141 Although research on lncRNA-based risk stratification during the clinical stages of PE (mid to late pregnancy) remains limited, the potential of lncRNAs as complements to miRNA biomarkers is widely recognized. Advances in ncRNA detection methodology are particularly promising. Recent reports have described the integration of microfluidics with nano-electrochemical biosensors for miRNA detection in early PE screening.142 Optimization of these technologies for exosome extraction and ncRNA quantification could facilitate the translation of laboratory findings into clinical-grade liquid biopsy tools. However, current research on mid-to-late pregnancy (the clinical diagnostic stage) still has notable limitations. Firstly, although reports on differentially expressed miRNAs are increasing, many studies have not strictly distinguished PE subtypes (such as early-onset versus late-onset), presence of fetal growth restriction (IUGR), or gestational age, leading to substantial heterogeneity in the results.119,143 Secondly, systematic large-scale studies on lncRNAs and circRNAs remain extremely scarce. For example, although reviews have highlighted the potential of circRNAs as PE biomarkers, their biogenesis, selective sorting into exosomes, and expression dynamics throughout mid-to-late pregnancy remain largely unknown.69 Therefore, although multiple basic and preliminary clinical studies support the feasibility of extracellular vesicle ncRNA for PE risk stratification and diagnosis in mid-to-late pregnancy, large-scale, multicenter, prospective studies are required to achieve true clinical translation. These studies should clarify the associations between ncRNAs and PE incidence, severity, and subtypes (such as early-onset and late-onset).80,144 At the same time, integrating ncRNA markers with traditional clinical, biochemical, and ultrasound indicators will help construct multiparameter risk models and evaluate their predictive value for maternal and fetal prognosis. For basic research, it is essential to explore the functional mechanisms of ncRNAs—whether they act merely as passive biomarkers or actively regulate the pathological processes of PE, potentially serving as therapeutic targets. This would open avenues for future nanomedicine interventions, such as EV-associated ncRNA delivery systems.45,145
The Advantages of EVs as Biomarkers
At present, when incorporating extracellular vesicle ncRNA (miRNA, lncRNA, and circRNA) into PE diagnosis and prediction systems, comparing EV-associated ncRNAs with traditional biomarkers can help determine whether they provide additional predictive or diagnostic value. However, therapeutic potential cannot be inferred from biomarker performance alone and must be evaluated separately through mechanistic studies, delivery experiments, pharmacokinetic and biodistribution analyses, and pregnancy-specific safety testing.126,146
Firstly, compared with traditional biomarkers, extracellular vesicle ncRNAs exhibit superior stability, specificity, and kinetic advantages. For example, small RNA sequencing studies of circulating EVs have shown that multiple miRNAs are detectable during the first trimester. Differential expression of these miRNAs may indicate PE risk before the appearance of clinical symptoms and traditional indicators.119 EVs possess a lipid bilayer membrane that effectively protects their contents from RNase degradation in plasma. The encapsulated ncRNAs are highly stable, can be stored for extended periods, and reflect maternal-fetal communication. This stability is superior to that of free miRNAs or protein markers.141
Secondly, from the perspective of subtype diagnosis and risk stratification, extracellular vesicle ncRNAs can help distinguish different PE subtypes (such as early-onset versus late-onset, and mild versus severe).47,147 For example, Scout Bowman Gibson et al developed a simplified method to evaluate circulating plasma small extracellular vesicles (sEVs) in maternal plasma. Their results showed that HIF-1α and its direct signaling partner miR-210 were detectable in systemic maternal sEVs, providing a foundation for understanding how sEV signaling contributes to early PE.135 These preliminary findings suggest that circulating EV-associated miRNAs may contribute to early PE risk assessment. However, their added predictive value must be validated against contemporary screening models incorporating maternal risk factors, mean arterial pressure, uterine artery Doppler, PlGF, and other angiogenic markers, rather than blood pressure or proteinuria alone. This highlights the high potential of extracellular vesicle miRNAs in early PE risk assessment. More importantly, such models can identify high-risk pregnant women before symptom onset and hold substantial clinical translational value.
In addition, unlike single proteins or angiogenic factors, ncRNAs (especially lncRNAs and circRNAs) can reflect more complex upstream regulatory networks. This capability is expected to reveal molecular mechanistic differences among PE subtypes.148 Previous reviews have indicated that extracellular vesicle-derived lncRNAs participate in maternal-fetal cell communication. They regulate trophoblast function in pathological pregnancies, including PE, by modulating gene transcription, competitively binding miRNAs, and influencing mRNA stability.141 Similarly, circRNAs have been identified as differentially expressed molecules in blood and placental samples from PE patients due to their stable circular structure and resistance to degradation. Certain circRNAs, such as hsa_circ_0036877, have been proposed as potential plasma biomarkers.149,150 Therefore, these ncRNAs can not only indicate PE risk but may also differentiate disease severity and pathogenic subtypes, for example by reflecting angiogenesis disorders versus immune- or inflammation-dominant PE through the circRNA-miRNA-mRNA ceRNA network.
Finally, incorporating extracellular vesicle ncRNAs is expected to significantly enhance clinical early prediction and personalized intervention. Based on early pregnancy samples, ncRNA panels can support risk stratification, intensified monitoring, and early preventive measures (such as low-dose aspirin). Meanwhile, elucidating the molecular signatures of different PE subtypes can lay the foundation for developing precise, subtype-specific treatment strategies in the future.151,152 Recent studies have emphasized the stability and therapeutic potential of extracellular vesicle carriers, suggesting that extracellular vesicle ncRNAs serve not only as biomarkers but also as potential therapeutic targets.
Multi-Biomarker Panels: Advancing from Single ncRNA to Integrated Multi-Omics Signatures
Multi-omics integration may help determine whether EV-associated ncRNAs add predictive or stratification value beyond established clinical, biophysical, and angiogenic variables. However, multi-omics biomarker performance should not be interpreted as evidence of therapeutic efficacy. Therapeutic relevance requires separate functional validation and delivery studies.45,153 Early studies mainly relied on qRT PCR or small scale microarrays to validate a limited number of extracellular vesicle miRNAs or lncRNAs. For example, several studies have focused on miR-210, miR-153-3p, and other molecules carried by extracellular vesicles derived from the placenta. These molecules affect trophoblast invasion and endothelial cell migration by regulating the AKT/HIF-1α/VEGF axis, TGF-β/Smad signaling, and oxidative stress related pathways, thereby contributing to placental ischemia, hypoxia, and vascular endothelial injury in PE.153 On this basis, researchers began to systematically characterize the miRNA profiles of extracellular vesicles. Using small RNA sequencing or high throughput microarrays, they performed unbiased transcriptomic profiling of extracellular vesicles derived from the peripheral blood or placenta of pregnant women with PE. These approaches can identify hundreds of differentially expressed miRNAs in a single experiment. Combined with GO/KEGG enrichment analysis and target gene prediction, these data showed that the miRNAs were mainly enriched in pathways related to immune regulation, angiogenesis, lipid metabolism, and endothelial function, providing a relatively comprehensive candidate pool for subsequent biomarker screening (Figure 5).109,139 For example, Shubhamoy Ghosh et al conducted a longitudinal cohort study in which extracellular vesicle miRNA (EV miRNA) in maternal plasma was sequenced at multiple time points during early pregnancy. They constructed a multi miRNA signature that predicted PE several weeks before clinical symptoms appeared and validated its high AUC in an independent cohort, demonstrating the technical feasibility of EV miRNA liquid biopsy for the early identification of high risk populations.47 At the mechanistic level, studies of single ncRNA function remain essential for linking omics discoveries to targeted interventions. For example, Wu et al found that placenta derived EVs or trophoblast derived EVs can transport miR-125b to umbilical vein endothelial cells. This process inhibits the expression of tight junction proteins such as VE cadherin, increases ROS generation, enhances endothelial barrier permeability, and reproduces the endothelial dysfunction phenotype of PE in vitro, providing direct evidence for the placental vascular axis.154 Li et al reported that maternal plasma exosomal miR-3198 regulates the proliferation, migration, and apoptosis of HTR-8/SVneo cells and primary trophoblast cells by targeting WNT3. miR-3198 is downregulated in patients with PE, and in vitro supplementation partially reverses trophoblast functional defects, providing a clear molecular target for engineered delivery and nanomedicine design based on exosome miRNA.145 Meanwhile, researchers have increasingly recognized that a single miRNA is insufficient to capture the high heterogeneity and diverse disease courses of PE. Therefore, research strategies have shifted toward multicenter and multi platform detection. High throughput platforms, including NanoString, TaqMan low-density chips, and digital PCR, are widely used to simultaneously quantify dozens to hundreds of EV miRNAs in limited samples. Algorithms such as LASSO regression, random forest, and extreme gradient boosting tree are then used to identify four or five highly informative miRNA combinations and construct comprehensive risk scores. These scores can distinguish early onset and late onset PE from other subtypes of gestational hypertension, supporting the transition from single indicator thresholds to multi biomarker pattern recognition.110,155,156 For example, Anat Aharon et al used NanoString to perform lineage specific analysis of maternal and placenta derived EV miRNAs. They found that several miRNA combinations not only distinguished placental and maternal pathological processes but were also significantly associated with adverse outcomes, including fetal growth restriction and premature birth. These findings suggest that EV miRNA panels may support the future classification and risk stratification of PE and related pregnancy vascular complications.110
 |
Figure 5 Extracellular miRNA (ex-miRNA) biomarker discovery and iterative selection for early prediction of PE. (A) Top 10 univariate ex-miRNA candidates. Heatmap showing expression patterns of the top 10 single ex-miRNA biomarkers with hierarchical clustering of ex-miRNAs and samples, demonstrating clear separation between PE cases and controls. (B) Verified bivariate ex-miRNA candidates (n = 110). Heatmap of log-transformed ratios for 110 validated bivariate biomarkers, clustered by miRNA composition and genomic location, with sample annotation indicating separation by diagnosis and time-to-diagnosis. (C) Putative cellular/tissue origins of candidate ex-miRNAs. Stacked bar plot summarizing the predicted contributing sources of candidate ex-miRNA biomarkers. (D–F) Three bivariate biomarkers selected by iterative machine learning, shown by cohort-wide heatmaps and PCA separation versus randomly selected bivariates. Adapted from Morey, Robert et al Discovery and verification of extracellular microRNA biomarkers for diagnostic and prognostic assessment of preeclampsia at triage. Science advances. Copyright © 2023 by authors.139 |
For nanoscale analysis of extracellular vesicle heterogeneity, emerging technologies such as single particle EV flow cytometry, nanoflow cytometry, and microfluidic single vesicle imaging have enabled researchers to classify and quantify placental EVs in maternal circulation or amniotic fluid. These analyses can be based on vesicle size, surface markers such as PLAP, and even single vesicle RNA content. Placental EVs with specific phenotypes, such as STB-EV, are significantly increased in patients with PE and are enriched in antiangiogenic signals, providing a technical foundation for applying single vesicle omics in PE.128,157,158 For example, Wei et al reported that SVAtlas, a single EV multi omics database, integrates proteomic and transcriptomic information from EVs across different diseases and fluid sources. This resource may support future cross cohort comparisons of PE exosomal ncRNA profiles with those from other pregnancy complications and cardiovascular disease cohorts, facilitating the identification of shared and disease specific nanofingerprints.158 At a systems level, integrated multi omics research is shifting the role of extracellular vesicle ncRNA from a single molecular marker to a multidimensional network node For example, Fang et al developed the CPMAAD system, which integrates PE related transcriptomic, methylation, multi chip, and clinical phenotype data from GEO. This system provides an integrated visualization interface for analyses spanning genes, pathways, and phenotypes. In this framework, ncRNAs from extracellular vesicles in maternal blood or placental tissue can be analyzed together with tissue transcriptomic and methylation profiles to reconstruct multilayer interaction networks linking ncRNA, mRNA, proteins, metabolites, and clinical phenotypes.155 Furthermore, integrated analyses of the placental transcriptome and metabolome showed that amino acid and lipid metabolic disorders are closely associated with inflammatory signaling pathways. These disorders are also significantly correlated with classical PE genes such as FLT1 and ENG and with several miRNA/lncRNA modules. In addition, multi omics studies integrating bulk RNA seq, single cell transcriptomics, immune infiltration features, and virtual drug screening have identified BIRC3, which is related to necrotic apoptosis, as a potential diagnostic and therapeutic target for PE. These findings provide structured information for future pathway design linking upstream signals, exosomal ncRNAs, and engineered nanocarriers.159,160 At the level of prediction models and clinical translation, the multi omics longitudinal cohort reported by Ivana Marić analyzed six types of omics data, including transcriptomic, proteomic, metabolomic, and immunophenotypic data. The study used nested cross validation and stacked ensemble learning to achieve high precision prediction of PE risk before 16 weeks of pregnancy. Its 9 component urinary metabolome model achieved an AUC of approximately 0.83 to 0.88 in the independent cohort, providing a paradigm for an early screening framework based on multi omics and machine learning.161 Subsequently, Zhao et al conducted a multicenter study that integrated early pregnancy clinical features, plasma proteomic and metabolomic profiles, and laboratory indicators. Using a random forest model, they established predictive models for early onset and late onset PE and showed that combining multi omics features with conventional clinical variables performed significantly better than single dimensional models. This approach can be translated into the development of integrated biomarkers combining extracellular vesicle ncRNA, proteomic and metabolomic profiles, and clinical variables. Through a three stage pathway involving training, validation, and independent testing, this strategy may gradually approach real clinical application scenario (Figure 6).162
 |
Figure 6 Development and validation of a random forest–based multi-omics model for early prediction of PE. (A) The top 10 metabolic and proteomic features were selected to generate 968 candidate biomarker combinations, and random forest models were trained and internally validated using three-fold cross-validation. (B) QQ plot summarizing the distribution of AUC values across all tested combinations, highlighting the optimal metabolite and protein panels. (C) Model performance comparison (AUC) in the training and test sets using clinical variables alone or combined with laboratory, metabolomic, and proteomic features. (D) ROC curves of the optimal model in training and test cohorts. (E) Spearman correlation heatmap showing relationships between predictors and model-derived risk scores in the overall dataset. This workflow supports a clinically translatable strategy for integrated biomarker development and can be further extended toward “extracellular vesicle ncRNA + proteome/metabolome + clinical variables” within a training–validation–independent testing framework. Adapted from Zhao Q, Li J, Diao Z et al Early prediction of preeclampsia from clinical, multi-omics and laboratory data using random forest model. BMC Pregnancy Childbirth. Copyright © 2025 by authors.162 |
In addition, multiple systematic reviews and meta-analyses have shown that, compared with single ncRNA markers, models based on multi ncRNA panels or combinations of ncRNAs with traditional serological indicators generally achieve higher sensitivity and specificity. When ncRNAs are incorporated into a high dimensional feature space with proteins, metabolites, and genetic variants, algorithms such as regularized regression, gradient boosting tree, and deep learning can significantly improve diagnostic OR values and AUC. These findings provide an important basis for evidence based evaluation of the future incorporation of extracellular vesicle ncRNA into routine PE screening.153,161–163 However, multi omics integration also presents several challenges, including large sample size requirements, substantial batch effects, insufficient data sharing, and limited standardization of analytical workflows. Reviews on the application of high dimensional technologies in PE research emphasize the need for future multicenter, large sample, prospective cohorts that follow extracellular vesicle research standards such as MISEV. Unified exosome isolation, quantification, and sequencing workflows should be embedded into existing prenatal screening and follow up pathways and combined with cloud databases and visualization tools to enable a smoother transition from laboratory discovery to standardized clinical testing.158,161,163,164 Looking ahead, the shift from single candidate extracellular vesicle ncRNAs to nanovesicle centered multi omics integration is moving PE diagnosis from single indicator static thresholds toward multidimensional dynamic risk curves. At the same time, this shift provides evidence based targets and patient stratification tools for engineering extracellular vesicles or biomimetic nanocarriers based on key ncRNA pathways. It also establishes methodological and translational frameworks for realizing extracellular vesicle ncRNA mediated integration of PE diagnosis and treatment.145,156,163
Extracellular Vesicle ncRNA as Therapeutic Target and Nano Drug Delivery Carrier: Towards Integrated Theranostics of PE
EV-Associated RNA as a Therapeutic Target for PE
The therapeutic relevance of EV-associated ncRNAs should be evaluated separately from their biomarker performance. A molecule that predicts PE risk or correlates with disease severity is not necessarily a valid therapeutic target. Therapeutic candidacy requires evidence that modulating the ncRNA alters disease-relevant pathways in appropriate pregnancy-relevant models, together with delivery feasibility, biodistribution, dose–response relationships, and maternal–fetal safety assessment. With advances in extracellular vesicle isolation and purification technologies, extracellular vesicle ncRNA can be viewed not only as a pathological signal but also as a nanoscale regulatory node that can be therapeutically targeted through the combined use of natural extracellular vesicles and synthetic nanomaterials.45,165 We consider this one of the most promising directions in current precision therapy research for PE. The following section discusses the evidence chain and nanomedicine design strategies for targeting extracellular vesicle RNA in PE therapy (Table 3).
 |
Table 3 Extracellular Vesicle RNA as a Therapeutic Target for PE and Its Nano Delivery Platform |
Multi omics and exosomal transcriptomic analyses have shown that miRNAs, lncRNAs, and circRNAs carried by placenta derived, decidual immune cell derived, and endothelial derived EVs form a cross tissue regulatory network. Among these molecules, miR-210, miR-155, miR-146a-5p, lncRNA H19, and several circRNAs are positioned at key intersections of trophoblast invasion, spiral artery remodeling, antiangiogenic and proangiogenic balance, and inflammatory responses, providing a theoretical basis for selecting target EV-associated ncRNAs.45,56,69 For example, the systematic review by Cretu et al showed that miR-210 is consistently upregulated in the PE placenta and maternal circulation. By targeting multiple genes related to mitochondrial metabolism and angiogenesis, miR-210 is associated with HIF-1α related hypoxia signaling and with reduced trophoblast invasion like and angiogenesis related phenotypes in experimental systems. These data support the functional involvement of miR-210 but do not establish direct causality in human PE progression or spiral artery remodeling. Experimental evidence suggests that miR-210 may enhance the HIF-1α dependent hypoxic feedback loop and inhibit trophoblast invasion and angiogenesis. Therefore, miR-210 is considered one of the most representative pathogenic extracellular vesicle miRNA targets, suggesting that anti-miR-210 nanomaterials may have a defined pathway of action.134 Similarly, extracellular vesicle circRNA DNAJB6 derived from decidual macrophages is upregulated in patients with PE. By acting as a miRNA sponge, circRNA DNAJB6 impairs trophoblast proliferation, migration, and invasion and promotes inflammation and endothelial dysfunction. Thus, circRNA DNAJB6 may represent a pathogenic extracellular vesicle circRNA upstream hub and provides a useful example for circRNA targeted intervention.19 In contrast to these pathogenic ncRNAs, some extracellular vesicle ncRNAs exhibit protective phenotypes. For example, MSC secreted EVs can deliver lncRNA H19 to trophoblast cells, where it competitively binds let-7b, releases FOXO1 from let-7b mediated inhibition, and activates AKT signaling. This process significantly enhances the invasion and migration of HTR-8/SVneo cells, inhibits apoptosis, and partially improves trophoblast migration, invasion like behavior, and apoptosis related phenotypes in vitro, thereby promoting a placental phenotype closer to that of normal pregnancy. This mechanism supports the H19/let-7b/FOXO1 axis as a therapeutic target that can be enhanced through exosome delivery (Figure 7).168 In addition, several reviews have summarized evidence that MSC Exos enhance metabolic activity, restore vascular balance, and regulate the immune microenvironment in PE animal models. These reviews indicate that MSC Exos may be safer than cell therapy because of their low immunogenicity and lack of proliferative capacity. MSC Exos can be regarded as natural nanomaterials loaded with nucleic acid drugs and may also serve as platforms for surface ligand modification and future placenta targeted delivery.152,169
 |
Figure 7 Experimental evidence for let-7b/FOXO1 regulation of trophoblast-like cell migration, invasion-like behavior, and apoptosis-related phenotypes in PE-related models. This figure summarizes representative experimental data related to the let-7b/FOXO1 axis in PE-associated trophoblast dysfunction. HTR-8/SVneo trophoblast-like cells were transfected with mimic-NC, let-7b mimic, inhibitor-NC, or let-7b inhibitor. (A) Heatmap showing differentially expressed miRNAs in the GEO dataset GSE96985. The top annotation bar indicates sample type, with turquoise representing normal samples and salmon/pink representing PE samples; the heatmap color gradient represents relative miRNA expression intensity. The red box in panel A highlights the differentially expressed let-7 family members, including hsa-let-7c-5p and hsa-let-7b-5p, which were selected from the miRNA expression heatmap as candidate miRNAs for subsequent analysis. (B) qRT-PCR analysis of let-7b expression in placental tissues from normal and PE pregnancies. (C) Bioinformatic prediction of the let-7b binding site in the FOXO1 3′UTR. The highlighted box in panel C indicates the predicted miRNA–target gene information, including miRNA name, target score, seed location, gene symbol, and 3′UTR length. The red text in panel C indicates the predicted miRNA–target gene pair, showing that hsa-let-7b-3p was predicted to target FOXO1, with 2308 representing the NCBI Gene ID of FOXO1 and NM_002015 representing the GenBank accession number of the FOXO1 transcript. (D) Dual-luciferase reporter assay assessing the interaction between let-7b and FOXO1 using wild-type and mutant FOXO1 3′UTR constructs. (E) FOXO1 mRNA expression measured by qRT-PCR after let-7b mimic or inhibitor treatment. (F and G) FOXO1 and phosphorylated FOXO1 protein expression assessed by Western blot and quantified by densitometry. (H and I) Trophoblast-like cell migration and invasion-like behavior evaluated using Transwell assays; representative images and quantitative cell counts are shown. (J) Apoptosis-related changes assessed by TUNEL staining; representative images and quantitative apoptosis rates are shown. Black bar labels indicate control or experimental transfection groups as shown in each panel. The asterisk (*) indicates p < 0.05 versus the normal group or mimic-NC group, as appropriate; the hash symbol (#) indicates p < 0.05 versus the inhibitor-NC group. Bold panel letters identify individual experimental panels. These data should be interpreted as model-based evidence from HTR-8/SVneo trophoblast-like cells and associated tissue or database analyses, rather than direct evidence of spiral artery remodeling or clinical therapeutic efficacy in human PE. Adapted from Chen, Yang et al MSC-Secreted Exosomal H19 Promotes Trophoblast Cell Invasion and Migration by Downregulating let-7b and Upregulating FOXO1. Molecular therapy. Nucleic acids. Copyright © 2020 by authors.168 Abbreviations: PE, preeclampsia; EV, extracellular vesicle; H19, H19 imprinted maternally expressed transcript; FOXO1, forkhead box O1; GEO, Gene Expression Omnibus; miRNA, microRNA; qRT-PCR, quantitative reverse-transcription polymerase chain reaction; 3′UTR, 3′ untranslated region; WT, wild type; Mut, mutant; NC, negative control; p-FOXO1, phosphorylated FOXO1; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; TUNEL, terminal deoxynucleotidyl transferase dUTP nick-end labeling; SD, standard deviation. |
At the specific RNA target level, miR-146a-5p, a typical anti inflammatory and pro repair miRNA, has been repeatedly validated through two independent exosome mediated pathways. On the one hand, miR-146a-5p enriched in amniotic fluid derived EVs (AF Exos) promotes trophoblast proliferation and migration in vitro and significantly alleviates hypertension, proteinuria, and placental ischemia in PE rat models. This mechanism is closely related to inhibition of HIF-1α/FT-1 expression and downregulation of antiangiogenic signals, suggesting that the HIF-1α/FT-1 axis is not only a diagnostic biomarker but also a miR-146a-5p mediated therapeutic target.76 In antiphospholipid antibody induced placental injury models, hUCMSC-EV associated miR-146a-5p reduces trophoblast apoptosis and inflammatory signaling and improves selected placental functional readouts. When vascular remodeling is assessed histologically, these findings can be interpreted as model specific evidence of improved placental vascular remodeling. Otherwise, they should be regarded as indirect evidence of improved placental function rather than direct proof of restored spiral artery remodeling. In this context, Lv et al used human umbilical cord MSC exosomes (hUCMSC Exos) to deliver miR-146a-5p in an antiphospholipid antibody induced placental injury model. This treatment inhibited TRAF6/NF-κB signaling, downregulated IL-1β and IL-18, regulated Cleaved-CASP3, BAX, and BCL2 expression, and reduced trophoblast apoptosis and inflammation. These findings highlight the central role of the miR-146a-5p/TRAF6 axis in pregnancy associated immune placental disease and support the mechanistic feasibility of miR-146a enriched exosomes or biomimetic nanoparticles for placental immunomodulation in PE and autoimmune associated pregnancy complications.166 At present, current basic studies and systematic reviews indicate at multiple levels that extracellular vesicle ncRNA is not only a liquid biopsy fingerprint reflecting placental and maternal endothelial status but also a key node in the pathological network. The miRNA/lncRNA/circRNA target spectrum built around extracellular vesicle ncRNA can guide RNA cargo selection for nanomedicine design.65,165,170
At the therapeutic level, current strategies for engineered EVs mainly include two categories. The first is to design functional EVs that use MSC Exos, placenta derived EVs, or trophoblast derived EVs as natural carriers. These EVs can be loaded with miRNA mimics or antisense oligonucleotides and modified on the surface with trophoblast targeting or placenta specific peptides to improve enrichment in the placenta or spiral artery wall. The second is to develop synthetic nanoparticles, such as PEG-PLA/DOTAP polymer nanoparticles and ionizable lipid nanoparticles, that achieve placental tropism through physicochemical regulation and endogenous plasma protein adsorption. These platforms can then be loaded with siRNA or miRNA drugs against targets identified from the extracellular vesicle ncRNA network.48,171 For example, Li et al developed polymer nanoparticles based on COOH-PEG5k-PLA8k and the cationic lipid DOTAP. These nanoparticles were coupled with a peptide that recognizes chondroitin sulfate A on the syncytiotrophoblast surface, forming T-NPsiSFLT1 which efficiently delivers siRNA-FLT1 to HTR-8/SVneo cells and mouse placenta. In various PE animal models, this system significantly reduces placental and serum sFlt-1 levels, improves endothelial function and blood pressure, and increases fetal weight and placental vascular density. Thus, elevated exosomal or serum sFlt-1 can be transformed from a classic diagnostic marker into a therapeutic target that can be precisely knocked down using nanomaterials.171 Recent studies have identified an LNP55 platform that achieves placenta biased distribution through adsorption of β2-glycoprotein I, based on high throughput in vivo screening of ionizable lipid nanoparticles. After loading VEGF mRNA, a single intravenous administration of this platform in an inflammation and hypoxia induced PE mouse model maintains blood pressure control throughout late pregnancy, reduces serum sFlt-1, restores placental vascular structure and the immune microenvironment, and significantly improves fetal growth outcomes. This finding provides proof of concept for extending similar LNP platforms to miRNA inhibitors or lncRNA/circRNA modulators.48 In addition, placenta tropic vectors such as LNP55 can achieve efficient placental transfection while encapsulating mRNA encoding VEGF, with controllable overall toxicity. Using similar materials, anti-miR-210, miR-146a-5p mimics, or lncRNA H19 overexpression vectors could potentially be encapsulated to form an RNA nanomedicine library that closely matches the multi omics target spectrum of EVs. This strategy could align diagnostic markers and therapeutic molecules along the same disease axis.165,172 MSC Exos themselves can be regarded as biogenic nanomaterials, and their membrane protein profiles confer immune evasion and tissue penetration capacity. Through surface engineering, such as coupling trophoblast targeting peptides or integrin recognition ligands, MSC Exos are expected to achieve higher placental localization and lower maternal liver and kidney enrichment without substantially increasing clearance rates.152,169 Under the concept of theranostics, multiple cohort studies and reviews propose using miR-210, miR-155, miR-146a-5p, H19, and circRNA fingerprints in maternal plasma EVs or plasma EV enriched fractions as indicators for disease stratification and pharmacodynamic monitoring. On the one hand, these fingerprints can be used to identify high risk pregnant women with markedly abnormal miRNA axes who are most likely to benefit from specific RNA nanotherapies. On the other hand, their levels can be dynamically tracked after administration to evaluate the extent of in vivo modulation of disease axes such as sFlt-1, HIF-1α, and NF-κB. This approach may help advance exosomal ncRNA toward an integrated node that is both measurable and therapeutically actionable.56,69,172
Multi omics integration, combining EV-associated miRNA/lncRNA/circRNA profiles with placental transcriptomic and proteomic analyses, can also evaluate potential off target networks before administration. It can predict angiogenic or immune related side effects caused by specific miRNA targeting and support co delivery strategies, such as loading anti-miR-210 together with proangiogenic miR-126 or H19 in the same nanocarrier, to reconstruct a more complete placental repair pathway at the network level.165,169,172 Although no PE nanomedicines targeting extracellular vesicle ncRNA have entered clinical trials, Chinese patents related to MSC Exos for PE treatment, safety data for MSC Exos in non pregnancy indications, and the superior efficacy of placenta tropic LNPs in rodent models together provide a feasible foundation for future Phase I studies of ncRNA targeted nanotherapy guided by extracellular vesicle markers in high risk populations. This transition also marks a new stage in extracellular vesicle ncRNA theragnostic, shifting from passive disease signaling to active therapeutic targets and drug carriers.45,173 Therapeutic studies should be interpreted with particular caution because most evidence comes from cell line assays, ex vivo placental or endothelial systems, and rodent PE like models. Improvements in trophoblast migration, endothelial tube formation, blood pressure, proteinuria, or fetal growth in these models support preclinical plausibility but do not establish clinical efficacy, fetal safety, or therapeutic benefit in human PE. Clinical translation will require validation in primary human placental and immune cells, ex vivo placental perfusion systems, clinically relevant animal models, and pregnancy specific toxicology studies.
EVs as Natural Nanocarriers for ncRNA Delivery
From the perspective of theranostics, extracellular vesicles can be regarded as active nanomaterials that carry and deliver ncRNA. This property may enable precise intervention in key PE pathways and real time monitoring of disease progression through liquid biopsy. This represents one of the most promising directions in extracellular vesicle nanomedicine (Figure 8).169,174,175 EVs are approximately 30 to 150 nm in diameter and are enclosed by a lipid bilayer enriched in cholesterol, sphingomyelin, integrins, transmembrane proteins, and other membrane proteins. Their natural nanovesicle structure protects loaded miRNAs, lncRNAs, and circRNAs from nuclease degradation and is associated with low immunogenicity, good biocompatibility, and active uptake and tissue homing mediated by ligand receptor and endocytic pathways. Compared with traditional synthetic nanoparticles, extracellular vesicles may more readily cross vascular and placenta related barriers.174,176 Pregnancy related studies have shown that PE associated EV-associated miRNA signatures are enriched in EVs derived from maternal plasma, placenta, and amniotic fluid. These molecules may reflect the functional status of trophoblasts, endothelial cells, and immune cells and can serve as non invasive biomarkers. In addition, they may serve as pharmacologically targetable therapeutic nodes and natural carriers for ncRNA based precision interventions.177,178 For example, Lin et al showed that miR-146a-5p in amniotic fluid derived EVs from normal pregnancy is significantly downregulated in patients with PE. Treatment of hypoxia simulated trophoblast cell lines HTR-8/SVneo and JEG-3 with normal amniotic fluid EVs or miR-146a-5p mimics significantly restores cell proliferation and migration and inhibits HIF-1α/FLT-1 overexpression. In a rat PE model, intravenous injection of normal amniotic fluid EVs reduces blood pressure and 24 hour urinary protein and improves placental perfusion and fetal weight. These findings suggest that miR-146a-5p enriched AF Exos may exert nanodrug like anti hypoxic and antiangiogenic effects by inhibiting the HIF-1α/FLT-1 axis.76 In addition, extracellular vesicles derived from human umbilical cord mesenchymal stem cells (hUCMSC) show natural tropism toward the placenta and injured vascular endothelium. Multiple in vitro and in vivo studies have evaluated hUCMSC derived EVs as nanoplatforms for ncRNA delivery.169 For example, Jiang et al found that hUCMSC Exos transport miR-140-5p to trophoblast cells, downregulate FSTL3, promote trophoblast proliferation, migration, and invasion under hypoxic conditions, and inhibit inflammatory factor expression. Systemic administration significantly improves PE like phenotypes in rats.167 Chen et al reported that hUCMSC Exos overexpressing miR-342-5p enhance trophoblast invasion and improve placental angiogenesis by inhibiting PDCD4, thereby reducing blood pressure and proteinuria in spontaneously hypertensive rats and delaying PE progression.179 Extracellular vesicles derived from MSCs can also transport miR-139-5p, activate the ERK/MMP-2 pathway by downregulating protein tyrosine phosphatase, and significantly enhance trophoblast invasion and migration.180 MSC derived EVs can also transport lncRNA H19 and activate the lncRNA H19/let-7b/FOXO1/AKT cascade, thereby enhancing trophoblast invasion, inhibiting apoptosis, and supporting uterine spiral artery remodeling.168,169 Multiple studies have shown that MSC Exos derived from bone marrow, adipose tissue, placenta, or decidua carry combinations of ncRNAs that collectively regulate four major networks, including trophoblast function, angiogenesis, inflammation, and oxidative stress. These findings provide a strong preclinical basis for constructing multitarget and network based exosomal ncRNA intervention strategies.169 Notably, in addition to placental and stem cell sources, EVs secreted by the gut commensal bacterium Akkermansia muciniphila can also alleviate PE like phenotypes in mice by regulating maternal immunity and improving placental angiogenesis. This finding suggests that extracellular vesicle nanoparticles from multiple tissues and microenvironments may form a complementary carrier library for systemic remodeling of the maternal fetal interface.178
 |
Figure 8 Extracellular vesicles as natural nanocarriers enabling theranostic ncRNA delivery. This schematic illustrates EVs as bioactive nanomaterials that can be engineered in vitro to enhance ncRNA loading, prolong circulation, and improve in vivo delivery efficiency. By incorporating membrane coatings (eg., PEGylation), immune-evasion signals (eg., CD47/SIRPα), targeting ligands, and hybrid designs with liposomes or inorganic nanoparticles, EVs can reduce mononuclear phagocyte system (MPS) clearance and achieve preferential accumulation at target tissues. In the target microenvironment, stimuli-responsive strategies (eg., pH-sensitive interactions) enable controlled ncRNA release and therapeutic modulation of disease-relevant pathways. In parallel, EV-based platforms support bioimaging and externally triggered guidance (eg., magnetic fields or near-infrared irradiation), highlighting their translational potential for integrated diagnosis and treatment of PE through liquid biopsy–guided monitoring. Adapted from Zou, Zaijun et al Current Knowledge and Future Perspectives of Exosomes as Nanocarriers in Diagnosis and Treatment of Diseases. Int J Nanomedicine. Copyright © 2023 by authors.174 |
As carriers, extracellular vesicles can be regarded as highly evolved biological nanocarriers. They have a relatively concentrated particle size distribution and stable zeta potential. Membrane enriched integrins, transmembrane proteins, and glycoproteins provide complex targeting codes that enable cellular entry through clathrin mediated endocytosis, micropinocytosis, and lipid raft dependent pathways and may facilitate the endosomal and lysosomal escape of nucleic acid cargoes.174–176 Multiple nanomedicine reviews have suggested that, compared with lipid nanoparticles (LNP), polymer micelles, and inorganic nanoparticles, extracellular vesicles may offer advantages in circulation half life, immunogenicity, organ accumulation, and nucleic acid protection. They may also provide higher loading efficiency for nucleic acid macromolecules, making them particularly suitable for fragile payloads such as miRNA, siRNA, circRNA, and ASO.174,175 In terms of loading technologies, endogenous loading can be achieved by transfecting donor cells, whereas exogenous loading can be performed using electroporation, ultrasound, freeze thaw cycles, or membrane extrusion. Multiple reviews of engineered EVs have systematically compared the effects of different loading processes on particle size, drug loading capacity, and membrane protein retention, providing a technical basis for encapsulating PE related ncRNAs, such as miR-146a-5p and miR-140-5p, into standardized exosome based therapeutics.174–176
In terms of material modification, nanotechnology is gradually upgrading natural vesicles into intelligent therapeutic carriers. On the one hand, covalent or non covalent grafting of polyethylene glycol (PEG), placenta homing peptides, or anchored anti-sFlt-1/Ang-II receptor antibody fragments onto the EV surface can prolong circulation time and enhance targeting to the placenta and injured endothelium.174 On the other hand, membrane fusion based hybrid EVs (MFHE) fuse nucleic acid rich EVs with liposomes or polymer nanoparticles with high drug loading capacity through freeze thaw cycles, PEG mediated fusion, or membrane extrusion. This strategy substantially improves drug loading capacity and surface functionalization while preserving EV membrane markers and biocompatibility, providing a paradigm for single particle co delivery of ncRNAs and small molecule or protein drugs.181 Furthermore, several teams have developed inorganic and organic extracellular vesicle hybrid platforms by embedding superparamagnetic iron oxide, gold nanorods, or quantum dots into the extracellular vesicle membrane or lumen. These platforms integrate MRI, photoacoustic, or fluorescence imaging with RNA therapy, enabling real time visualization of extracellular vesicle distribution and therapeutic effects in vivo and laying a materials science foundation for exosome based ncRNA therapeutics.182 At the same time, various nanomaterials have also been used for extracellular vesicle processing and enrichment. For example, aptamer modified magnetic graphene oxide nanoparticles can efficiently capture placental or MSC derived extracellular vesicles from blood. This approach provides an enrichment strategy for preparing high purity and recyclable therapeutic extracellular vesicles and lays the foundation for subsequent clinical grade formulation production and quality control.183
Research on EVs in pregnancy related diseases has gradually shifted from mechanistic exploration to animal experiments and the design of early intervention strategies. Systematic reviews indicate that EVs play a central role in regulating immune tolerance, trophoblast invasion, and angiogenesis at the maternal fetal interface. Various pregnancy complications are accompanied by changes in EV cargo and ncRNA profiles, providing theoretical support for EV based therapies and biomarker development.175,177 Mouse experiments by Chen et al showed that oral or intravenous administration of EVs from specific sources, such as Akkermansia EVs or MSC Exos, can improve the placental vascular network and alleviate PE like phenotypes. These findings suggest that EV administration may be feasible and may have an acceptable safety window during pregnancy.178
Mouse experiments by Chen et al showed that oral or intravenous administration of EVs from specific sources, such as Akkermansia EVs or MSC Exos, can improve the placental vascular network and alleviate PE like phenotypes. These findings suggest that EV administration may be feasible and may have an acceptable safety window during pregnancy.176,184 At the same time, guidelines from the International EV Society and other organizations emphasize the need to standardize extracellular vesicle preparation, including large scale production, purity and potency testing, immunogenicity evaluation, and long term follow up.184,185 These requirements also represent the manufacturing and regulatory thresholds that the PE field must address before promoting the clinical application of extracellular vesicle ncRNA nanomaterials. Extracellular vesicle ncRNA is a key molecular node in the pathological network of PE and may help establish a true theragnostic loop for early prediction, real time monitoring, and personalized intervention in the future.
EV-Mimetic and Hybrid Nanocarriers as Long-Term Experimental Delivery Platforms
Constructing exosome like lipid nanoparticles, including exosome membrane coated LNPs and multicomponent hybrid carriers, is considered a key step in shifting exosomal ncRNA from a passive biomarker to an active therapeutic tool for PE intervention. This strategy may help establish a continuous translational chain linking liquid biopsy, risk stratification, and precise drug delivery, thereby providing a more controllable materials science framework for improving pregnancy safety in clinical practice. (Table 4 and Figure 9).93,186,187
 |
Figure 9 Experimental nanocarrier strategies for placenta-oriented delivery of nucleic acids and small-molecule drugs. This schematic summarizes representative nanocarrier platforms that may be explored for pregnancy- and PE-related drug delivery research. Therapeutic payloads, including siRNA, mRNA, and small-molecule drugs, can be incorporated into lipid-based, polymer-based, protein-based, EV-based, or EV-mimetic nanocarriers. These platforms may be engineered to improve payload stability, circulation properties, tissue interaction, and placenta-oriented delivery in preclinical models. In the context of PE, such systems could provide experimental tools for testing whether functionally validated EV-associated ncRNA targets can be modulated in pregnancy-relevant settings. However, these approaches remain preclinical and should not be interpreted as established placenta-specific or fetal-safe therapies. Their translational development requires rigorous evaluation of maternal biodistribution, placental accumulation, transplacental transfer, fetal exposure, immunogenicity, off-target uptake, dose–response relationships, manufacturing reproducibility, and long-term maternal and offspring safety. Abbreviations: PE, preeclampsia; EV, extracellular vesicle; ncRNA, non-coding RNA; siRNA, small interfering RNA; mRNA, messenger RNA. Adapted from Huang Y, Zhang X, Xue L, He C. Nano-Enabled Therapeutics: Novel Strategies for Preeclampsia Treatment. Int J Nanomedicine. Copyright © 2026 by authors.188 |
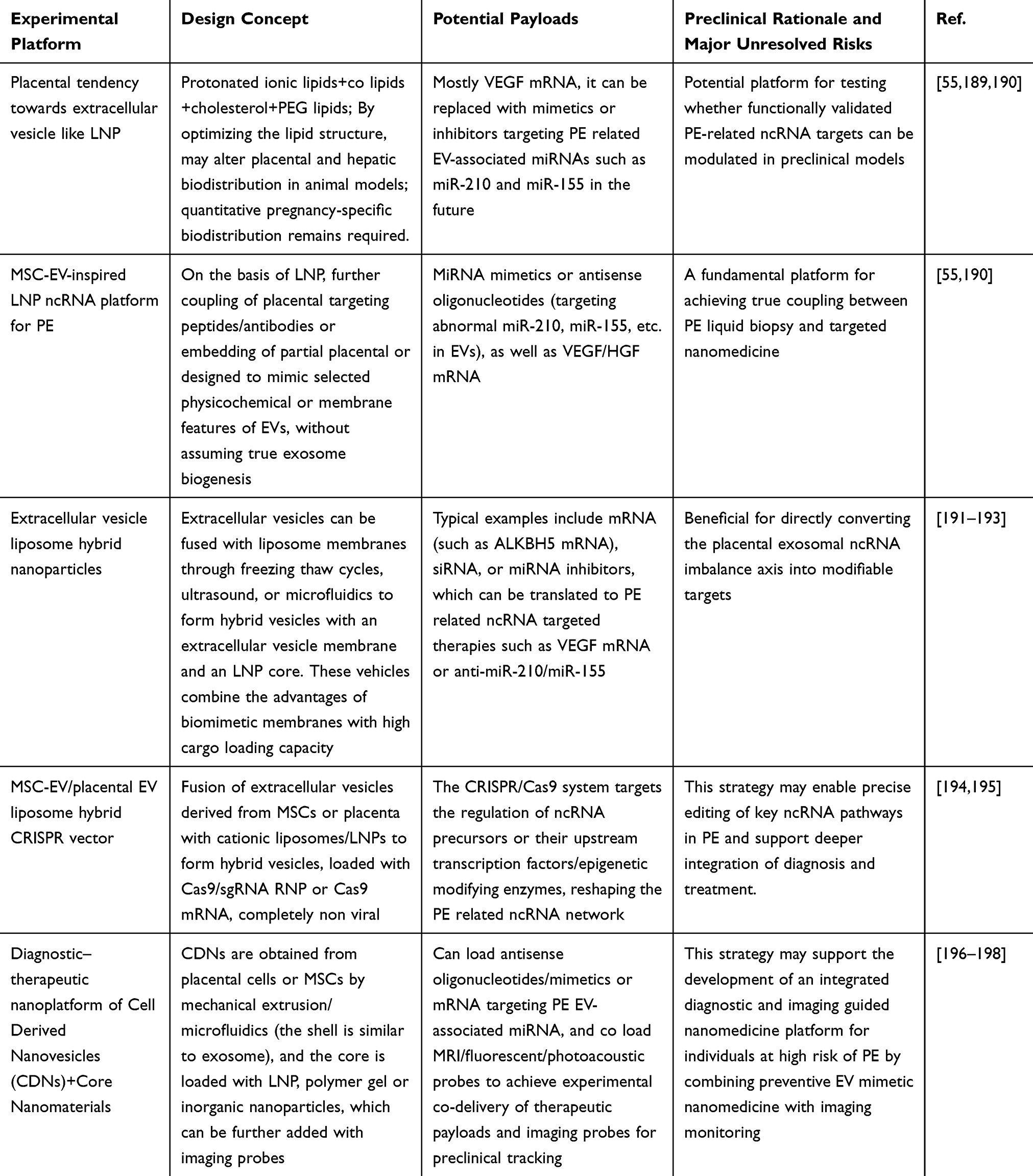 |
Table 4 Experimental EV-Inspired and Hybrid Nanocarrier Platforms for Future PE-Related ncRNA Delivery Research |
Previous studies have shown that human pregnancy associated extracellular vesicles (EVs) play a double edged role in the pathophysiology of PE. On the one hand, multiple miRNAs in placenta derived EVs, including miR-210, miR-155, and miR-9-5p, participate in disease development by regulating angiogenesis, endothelial function, and immune responses. On the other hand, these ncRNAs are enriched in vesicles that can be stably detected in peripheral blood and urine, providing highly specific molecular readouts for early non invasive diagnosis and dynamic monitoring.106 Existing large sample cohort studies have shown that the abundance of circulating EVs and miRNA/lncRNA profiles during pregnancy are significantly altered in patients with PE. Among these EVs, CD9 positive small EVs are significantly increased in both early onset and late onset PE and are strongly associated with placental dysfunction and adverse maternal fetal outcomes. These findings provide a quantifiable template for subsequent carrier amplification and signal reuse.109,186 Therefore, multiple reviews and translational studies have suggested that microRNA/lncRNA based interventions, such as antisense oligonucleotides, miRNA mimics, or CRISPR gene editing, represent a core direction for future molecular therapy in PE. These strategies target exosomal ncRNA axes closely related to placental ischemia, the antiangiogenic axis involving sFlt-1/PLGF imbalance, and systemic endothelial inflammation. Combining synthetic nanocarriers with EVs may help improve therapeutic efficacy while minimizing placental and maternal vascular toxicity.116,199 In the field of extracellular vesicle like LNPs, Kelsey L. Swingle et al recently identified a highly placenta oriented ionizable lipid nanoparticle, LNP-55, through high throughput material screening. This nanoparticle has a particle size of approximately 80 to 100 nm. LNP-55 adopts a classic LNP architecture composed of protonated ionizable lipids, helper lipids, cholesterol, and PEG lipids. Its structural parameters encode selective uptake by decidual and trophoblast cells while reducing distribution to traditional LNP target organs such as the liver48 In addition, the team encapsulated VEGF mRNA in LNP-55 for intravenous administration in a mouse PE model overexpressing sFlt-1. This treatment significantly improved placental blood flow, reduced systolic blood pressure, corrected abnormalities in placental small artery remodeling, and improved fetal growth restriction. Safety assessment suggested limited transplacental transfer and minimal histological changes in extraplacental tissues.48 This study and related work demonstrate that material structure encoded placental tropism is feasible at the animal level.189,200 At the same time, multiple reviews on gene and nucleic acid therapy for PE have noted that traditional adenoviral or AAV vectors are severely limited during pregnancy because of immunogenicity and integration risks. Degradable LNP systems can achieve controllable dosing and repeated administration. They can also regulate circulation half life and tissue affinity by adjusting lipid pKa, hydrophobic chain length, and PEG density, providing an ideal framework for accurately delivering inhibitors of pathogenic miRNAs, such as miR-210, miR-155, and miR-34a-3p, or miRNA mimics that promote angiogenesis and antioxidant activity to the placenta.55,189 Furthermore, Selvakumar et al summarized the evidence chain for microRNA based therapies in PE treatment and noted that most promising targets are located in pathways related to placental angiogenesis, trophoblast invasion, and immune tolerance. If high local exposure can be achieved at the maternal placental interface through extracellular vesicle like LNPs, this approach may reverse focal placental lesions without substantially affecting systemic blood pressure regulation.55 This type of LNP system can be further coupled on the surface with placenta specific peptides or antibodies, such as those targeting placental growth factor receptors or specific integrins. Alternatively, placental EV membrane proteins can be partially incorporated through membrane fusion. This approach combines bottom up material optimization with top down membrane biomimicry, thereby approximating the tissue tropism of natural placental EVs without sacrificing formulation stability.190,201
Compared with purely synthetic LNPs, rapidly developing hybrid carrier strategies integrate EVs with nanomaterials to form hybrid nanovesicles. This approach has shown improved performance in RNA therapy for tumors and inflammatory diseases and may be translated to pregnancy complications.202 A typical approach uses freeze thaw cycles, ultrasound, or microfluidics to induce the fusion of natural EVs with liposome membranes, thereby generating membrane fusion based hybrid EVs. This strategy retains cell specific protein barcodes on the EV membrane, such as adhesion molecules, integrins, and tetraspanins, while introducing the high loading capacity and surface modifiability of liposomes. It can therefore support the co delivery of nucleic acid drugs and small molecule drugs.202,203 Liu et al reviewed evidence showing that such hybrid vesicles can improve the encapsulation efficiency and cytoplasmic release of mRNA or siRNA during RNA intervention. Endosomal escape can also be enhanced by introducing pH sensitive or enzyme sensitive lipids or high entropy polymers, thereby overcoming the limited loading capacity and inefficient release of natural EVs.181 For example, Wu et al developed ALKBH5 mRNA loaded exosome liposome hybrid nanoparticles that restored m6A demethylase expression, reprogrammed tumor metabolism, and significantly inhibited tumor progression in colorectal cancer models. This study showed that an exosome membrane and LNP core structure can efficiently deliver mRNA while maintaining good in vivo safety and immune compatibility. This model has strong translational relevance for designing proangiogenic mRNA or miRNA inhibitor delivery systems targeting placental endothelial cells and trophoblast cells.204 At the same time, a Cas9 GFP delivery system constructed by hybridizing mesenchymal stem cell (MSC) EVs with cationic liposomes achieved efficient gene editing with low toxicity in HEK293T cells. This finding suggests that placental or immune tolerance related MSC-EVs could be combined with LNPs in the future to achieve in situ gene editing or epigenetic regulation of key PE related ncRNAs or upstream transcription factors.205 Another important type of exosome like platform is cell derived nanovesicles (CDNs), which are manufactured from cell membranes through top down mechanical extrusion or microfluidic methods. Their particle size and membrane protein profiles are highly similar to those of natural EVs, but they offer higher yield and more controllable composition. Therefore, CDNs are considered a scalable manufacturing alternative to natural EVs.190,196 Jang et al showed that CDNs derived from MSCs or immune cells can effectively load siRNA or miRNA and reproduce the immunoregulatory and targeted migration properties of natural EVs in various disease models.196 Poinsot et al systematically reviewed engineered and biomimetic exosome nanocapsules and noted that introducing programmable polymer nanoparticles or magnetic and photothermal inorganic nanomaterials into the CDN core can enable multimodal imaging and local controlled release. This strategy may integrate diagnosis, drug release, and efficacy monitoring into the same nanoplatform, which is a key prerequisite for true theranostics.201 In the PE context, an attractive strategy is to use EVs or CDNs from normal pregnant placentas or placental MSCs as biological shells and load their cores with ionizable LNPs or polymer nanogels. These cores could encapsulate antisense oligonucleotides, circRNA sponges, or therapeutic mRNAs, such as VEGF or HGF, that target pathogenic miRNAs such as miR-210 and miR-155, thereby restoring placental angiogenesis and endothelial homeostasis at the molecular level. At the same time, fluorescence or magnetic resonance contrast agents could be incorporated into the carrier to enable real time imaging of placental perfusion and nanodrug distribution, allowing the same nanovesicle to be both therapeutic and traceable.200,201
Embedding these materials science strategies into the clinical pathway of PE requires a deep understanding of placental EV biology and liquid biopsy systems. Large scale cohort studies have shown that EV abundance and miRNA profiles in the peripheral blood and urine of patients with PE are closely associated with pregnancy outcomes and long term cardiovascular risk. Therefore, high throughput detection methods such as CD9-ELISA may enable EV based stratification in early pregnancy.38,186,206 Studies showing long term blood pressure reduction, improved left ventricular function, and vascular remodeling in spontaneously hypertensive rats treated with normotensive pregnancy placental EVs support the concept that healthy placental EVs have endogenous cardiovascular protective effects. This provides conceptual support for designing normal placenta like EV material hybrid carriers for preventive intervention in high risk pregnant women.38,126,206 At the same time, multiple reviews on placenta targeted drug delivery and EV therapy for pregnancy related diseases have noted that applications in tumors and genetic diseases are mostly at the clinical trial stage, whereas the PE field remains at an early stage, with promising preclinical evidence but limited clinical validation. The main challenges include the lack of toxicological standards for nanomedicine during pregnancy, the high cost of long term follow up for placental and fetal exposure, and insufficient standardization of process scale up and quality control for EV preparations or EV enriched fractions.116,199 Therefore, future development of exosome like LNPs and hybrid carriers that combine synthetic nanocarriers with exosome components in PE should rely on existing experience from LNP mRNA therapy and EV based or EV mimetic nanomedicine in cancer. This approach can help gradually establish safety windows and dose exploration strategies for prenatal applications. In parallel, EV ncRNA indicators obtained from liquid biopsy should be embedded into clinical trial designs. The same set of EV-associated RNA readouts should be used to evaluate disease risk, drug exposure, and treatment response, thereby realizing the closed loop use of exosomal ncRNA as both therapeutic targets and diagnostic markers. This strategy may enable materials science driven nanotechnology to support the integrated precision diagnosis and treatment of PE.201,205
Challenges and Future Directions
Methodology and Translation Challenges
The major obstacle to translating EV-associated ncRNAs in PE is not the identification of a single candidate miRNA, lncRNA, circRNA, or nanocarrier, but the lack of standardized methodology across the entire workflow from sample collection, EV separation, vesicle characterization, RNA profiling, functional validation, nanomedicine manufacturing, and clinical trial design.58,116,207 Without methodological harmonization, even biologically plausible EV-ncRNA signals may fail to become reproducible biomarkers or regulated therapeutic products. The current comprehensive review on pregnancy related EVs has repeatedly emphasized that EVs undertake highly dynamic signal exchange functions at the maternal fetal interface, participating in normal implantation and placental vascular remodeling, as well as deeply intervening in the occurrence and development of pregnancy complications such as PE. Therefore, EV-associated ncRNAs should currently be viewed as promising but exploratory molecular tools whose clinical translation depends on technical reproducibility, biological source attribution, longitudinal validation, and pregnancy-specific safety assessment.58,116,207
The first challenge is EV heterogeneity (Figure 10). In PE, circulating EVs may originate from placental trophoblasts, maternal endothelial cells, platelets, leukocytes, renal cells, and other maternal or fetal compartments. These vesicles differ in size, density, membrane markers, biogenesis pathways, and ncRNA cargo. As a result, changes in a circulating miRNA cannot be simply attributed to the placenta or to one EV subtype without additional evidence.58,116,207 Studies comparing maternal blood EVs and placental tissue-derived EVs have shown that PE involves both placenta-enriched ncRNA alterations and broader maternal EV remodeling.110 Similarly, placental medium/large EV fractions and serum EV fractions may show partially overlapping but distinct miRNA profiles, suggesting that separation strategy strongly influences the observed signal.109 For this reason, future PE studies should distinguish tissue or cellular origin from biofluid sampling matrix: plasma, serum, urine, and amniotic fluid provide accessible sampling sources, but they do not by themselves define the biological origin of EV cargo. Source attribution will require approaches such as placental perfusion models, paired tissue–biofluid profiling, cell-specific surface markers, immunocapture strategies, single-vesicle analysis, and integration with placental histopathology or single-cell transcriptomic references.109
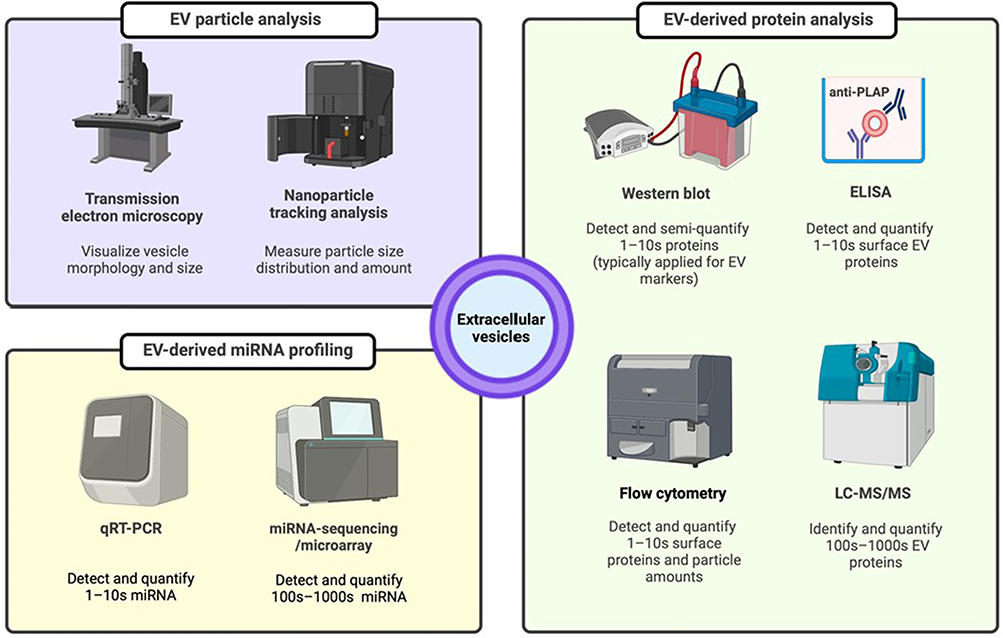 |
Figure 10 Methodological workflow for EV characterization and EV-associated miRNA profiling. This schematic summarizes the core analytical modules required for standardized EV-associated ncRNA research in PE. EV particle analysis, including transmission electron microscopy and nanoparticle tracking analysis, is used to assess vesicle morphology, size distribution, and particle concentration. EV-derived protein analysis, including Western blot, ELISA, flow cytometry, and LC–MS/MS, supports the detection or quantification of EV-associated surface and internal protein markers and helps evaluate vesicle enrichment, purity, and potential contamination. EV-derived miRNA profiling, including qRT-PCR, miRNA sequencing, and microarray analysis, enables detection and quantification of candidate EV-associated miRNAs for mechanistic studies and biomarker discovery. In PE research, combining particle-level characterization, protein-marker validation, and RNA profiling is essential to improve reproducibility, distinguish EV-associated signals from non-vesicular RNA contamination, and support longitudinal validation of candidate biomarkers before clinical translation. Adapted from Chaemsaithong P, Luewan S, Taweevisit Met al Placenta-Derived Extracellular Vesicles in Pregnancy Complications and Prospects on a Liquid Biopsy for Hemoglobin Bart’s Disease. Int J Mol Sci. Copyright © 2023 by authors.208 Abbreviations: EV, extracellular vesicle; PE, preeclampsia; ncRNA, non-coding RNA; miRNA, microRNA; TEM, transmission electron microscopy; NTA, nanoparticle tracking analysis; ELISA, enzyme-linked immunosorbent assay; LC–MS/MS, liquid chromatography–tandem mass spectrometry; qRT-PCR, quantitative reverse-transcription polymerase chain reaction. |
A second challenge is the inconsistent use of EV terminology (Figure 10).60,135 Many studies broadly use the term EVs, although isolated preparations may contain mixed populations, including small EVs, microvesicles, apoptotic bodies, lipoproteins, ribonucleoprotein complexes, and protein-bound extracellular RNAs.40,45 To improve clarity, EVs should be used as a general term when vesicle biogenesis has not been demonstrated. Small EVs should be used for preparations supported by size-based and marker-based characterization, whereas exosomes should be reserved for vesicles with evidence of endosomal origin.135 This terminological distinction is particularly important in PE because different vesicle populations may have different diagnostic and functional implications.209,210 Consistent reporting according to MISEV-based principles, including particle number, size distribution, EV-enriched and EV-negative markers, isolation method, purity assessment, and functional controls, is essential before EV-associated ncRNAs can be compared across studies.60,209,210 For biomarker translation, EV-associated ncRNA panels should be evaluated in prospective longitudinal cohorts using clinically relevant comparator models.33 These should include maternal risk factors, mean arterial pressure, uterine artery Doppler indices, PlGF, the sFlt-1/PlGF ratio, blood pressure, proteinuria, clinical symptoms, and routine laboratory markers.211 Added value should be assessed by discrimination, calibration, net reclassification, decision-curve analysis, and clinical usefulness, rather than by statistical significance alone.
The emergence of nanomedicine and EVs has provided new tools to solve some problems, but it has also brought new methodological and regulatory challenges.190,212–214 PE-related EV-ncRNA studies have used EDTA plasma, heparin plasma, serum, urine, placental perfusate, and amniotic fluid collected at different gestational ages and processed under different centrifugation, filtration, freezing, storage, and thawing conditions. These variables can alter EV yield, vesicle integrity, platelet activation, hemolysis, contaminating non-vesicular RNA, and apparent miRNA abundance. In addition, ultracentrifugation, density-gradient separation, size-exclusion chromatography, polymer precipitation, immunoaffinity capture, commercial kits, and microfluidic systems each introduce distinct biases in EV recovery, purity, scalability, and RNA yield.190,212,213 Thus, future biomarker studies should report pre-analytical variables in detail and should ideally compare or validate candidate EV-ncRNAs across standardized platforms rather than relying on a single isolation protocol. RNA-level standardization is equally important, including RNA extraction efficiency, spike-in controls, endogenous normalization strategies, sequencing depth, qPCR validation, batch-effect correction, and transparent reporting of low-abundance transcripts.190,212–214
Clinical validation represents another major barrier. Many currently reported EV-ncRNA signatures in PE are derived from small cohorts, cross-sectional comparisons, or samples obtained after disease onset.33,209 Such studies are valuable for discovery, but they cannot establish clinical prediction, diagnostic superiority, or subtype-specific utility without independent longitudinal validation. For early prediction, EV-ncRNA profiles should be measured before 20 weeks of gestation and tested for their incremental value beyond maternal risk factors, blood pressure, uterine artery Doppler, and angiogenic biomarkers such as the sFlt-1/PlGF ratio.132,215 For diagnosis and risk stratification after disease onset, EV-ncRNAs should be evaluated according to clinically meaningful phenotypes, including early-onset versus late-onset PE, PE with fetal growth restriction, severe-feature PE, superimposed PE, renal-dominant PE, and inflammation- or angiogenesis-dominant subtypes.30,32 Without PE subtype stratification, mixed cohorts may obscure biologically relevant EV-ncRNA patterns and lead to inconsistent biomarker performance.
The therapeutic translation of EV-based or EV-mimetic nanomedicine is even less mature than biomarker development and should be discussed separately. Engineered EVs, MSC-derived EVs, placental EV-inspired vesicles, lipid nanoparticle–EV hybrids, and synthetic exosome-mimetic carriers offer experimental opportunities for delivering miRNA mimics, antagomirs, siRNAs, mRNAs, or other regulatory molecules.36,216 However, these platforms also introduce substantial manufacturing and regulatory challenges. Natural EVs may show donor-to-donor variability, batch heterogeneity, variable cargo loading, incomplete potency definition, and difficulty in precise dose quantification. Synthetic or hybrid vesicles may improve manufacturability but may not fully reproduce the biodistribution, uptake pathways, or biological behavior of native pregnancy-related EVs at the maternal–fetal interface.217–219 Therefore, claims regarding placenta-specific targeting, fetal protection, or superiority over standard PE management should remain cautious until supported by rigorous biodistribution, pharmacokinetic, pharmacodynamic, and safety data.
Safety is a central issue for any EV-based nanomedicine intended for use during pregnancy.36 Unlike non-pregnant populations, PE therapy must consider two interconnected biological systems: the mother and the fetus. Key safety questions include maternal immune activation, complement activation, cytokine release, coagulation effects, off-target uptake by liver, spleen, kidney, lung, and vascular endothelium, placental accumulation, transplacental transfer, fetal exposure, embryo–fetal developmental toxicity, reproductive toxicity, genotoxicity of nucleic acid cargo, and the consequences of repeated dosing.220–222 In addition, surface engineering strategies that enhance targeting or circulation time, such as ligand conjugation, PEGylation, membrane fusion, or immune-evasion motifs, may also alter immunogenicity, clearance, placental transport, and long-term tissue retention.219,223 Thus, pregnancy-specific toxicology should include maternal organ toxicity, placental structure and function, fetal growth, fetal organ development, neonatal outcomes, and long-term offspring follow-up rather than relying only on standard adult nanomedicine safety assays.223,224
Regulatory and manufacturing requirements also remain unresolved.209,225 Clinical-grade EV or EV-mimetic products would require defined starting materials, controlled cell culture conditions, scalable purification, sterility and endotoxin testing, viral and adventitious-agent safety, potency assays, stability testing, release criteria, and batch-to-batch comparability under GXP-compliant conditions.226,227 For ncRNA-loaded EV products, additional quality attributes should include loading efficiency, cargo integrity, release kinetics, off-target gene modulation, and correlation between ncRNA dose and biological response.228,229 At present, there is no widely accepted regulatory pathway for preventive or therapeutic EV-ncRNA nanomedicine in pregnancy, and pregnant individuals have historically been excluded from early-phase drug trials. Future clinical development should therefore proceed through carefully staged studies, beginning with robust non-pregnant safety data, pregnancy-relevant animal models, ex vivo human placental perfusion systems, and ethically designed maternal–fetal follow-up protocols.230,231 Among potential nucleic-acid strategies, antisense oligonucleotides, miRNA mimics or inhibitors, siRNAs, and mRNA-based approaches may be considered experimental modalities for modulating PE-related pathways in preclinical systems.55,232 CRISPR-based approaches should be mentioned only as highly theoretical and currently unsuitable for near-term PE translation because of unresolved concerns regarding off-target editing, placental and fetal exposure, irreversibility, delivery specificity, reproductive safety, and ethical regulation during pregnancy.233,234
Finally, computational tools, multi-omics integration, and artificial intelligence may help organize complex EV-ncRNA datasets, but they should not be presented as immediate clinical solutions.235,236 These approaches may support EV subtype classification, source attribution, biomarker prioritization, PE subtype stratification, and prediction-model training. However, AI-based models are vulnerable to small sample size, batch effects, population bias, missing clinical metadata, and overfitting.237,238 For precision-medicine applications, EV-ncRNA algorithms should be trained and externally validated in diverse longitudinal pregnancy cohorts with standardized sample handling and transparent model reporting. In this context, AI, multi-omics, and placenta-targeted nanomedicine should be viewed as future-enabling strategies rather than clinically validated platforms.17,36,216,225 Overall, the next stage of the field should prioritize harmonized EV methodology, reproducible ncRNA quantification, longitudinal and subtype-specific validation, pregnancy-specific safety testing, and transparent regulatory standards before EV-associated ncRNAs can be translated into reliable PE biomarkers or safe therapeutic delivery systems.
Future Directions
To address current methodological and translational challenges, future research should advance technology, biology, and clinical translation in parallel. At the same time, actionable research pathways should be extracted from fragmented mechanistic studies and early translational exploration to link multi tissue derived extracellular vesicle ncRNA, nanomedicine, and pregnancy specific clinical scenarios. This approach may help establish a precision obstetric therapeutic framework that protects both maternal and fetal safety.58,117,208 The first key future direction is to construct a systematic EV-associated ncRNA atlas that spans the entire course of pregnancy, multiple tissues, and different PE subpopulations. Through standardized sample collection and multi omics integration, future studies can analyze dynamic EV-associated RNA communication networks among the placenta, maternal vasculature, immune system, kidney, and fetal compartment.109 Existing studies have demonstrated significant temporal and subtype specific differences in placenta derived EVs, miRNAs, and lncRNAs during normal pregnancy and PE. Combining multiple platforms to analyze plasma and urine EVs and distinguish small, medium, and large EV subgroups can help capture disease related signals more accurately, laying the foundation for a feasible liquid biopsy panel.47,126,208 However, most current cohorts have limited sample sizes and highly heterogeneous enrolled populations. Sampling time points are often concentrated after symptom onset, which limits their ability to support early pregnancy risk stratification and clinical trial enrollment stratification. Therefore, future prospective follow up cohorts based on high risk and multicenter populations are needed. These cohorts should collect extracellular vesicle ncRNA, protein, and clinical phenotype data in parallel on the same platform and adopt unified processing procedures and updated guidelines such as MISEV2023 to reduce technical batch effects.47,138,210
The second direction is to improve resolution by moving from overall EV level analysis to precise single vesicle and multidimensional spatial measurements. This should be deeply integrated with single cell genomics and spatial transcriptomics to define the heterogeneity and functional specialization of EVs from different sources in the placental microenvironment and distal vascular beds.128 For example, single particle flow cytometry and imaging flow cytometry based on markers such as placental alkaline phosphatase can quantitatively distinguish the abundance and phenotypic differences of placenta derived EVs across different pregnancy states. Analysis of single particle EV RNA in amniotic fluid and umbilical cord blood may reveal abnormal fetal signals in early pregnancy, suggesting the value of single vesicle resolution measurements for identifying high risk fetal and maternal phenotypes.128,157 In terms of computational methods, next generation tools such as miRTalk aim to infer cell cell communication networks from single cell transcriptomic and EV miRNA data. These tools can identify which placental or immune cell populations act as senders or receivers of specific extracellular vesicle ncRNA axes, providing topological target information for blocking harmful signals or amplifying protective signals.239 Integrating single vesicle and single cell data can also help distinguish communication patterns between early onset and late onset PE. It may also explain why the same EV miRNA shows opposite trends in placental tissue and circulating EVs, a common source of confusion in current studies. In the future, a three dimensional data framework integrating single cell, single vesicle, spatial, and liquid biopsy data should be systematically constructed while following MISEV2023 and pregnancy sample specific standards. This framework may reduce the non reproducibility of results across laboratories.128,210
The third direction is to develop computationally empowered diagnostic and prognostic models. These models should move beyond single miRNAs or small candidate panels and use machine learning and deep learning to integrate high dimensional extracellular vesicle ncRNA profiles, clinical indicators, and imaging features into interpretable multimodal models for early screening, phenotype stratification, and efficacy prediction.47,126,240–242 For example, an elastic net regularized logistic regression model based on early pregnancy plasma EV miRNA achieved predictive performance for late onset PE with an AUC close to 0.96 in small samples. This finding suggests that stable risk scores can be constructed in limited samples through appropriate feature selection and regularization. At the same time, systematic evaluations have assessed the performance of various machine learning and deep learning algorithms for surface enhanced Raman spectroscopy (SERS) fingerprinting and high throughput fluorescence signals from single particle EVs. These studies provide algorithmic baselines for future label free and high throughput EV detection devices.47,240,242 Furthermore, reviews on AI and EV precision medicine emphasize that clinical implementation requires large scale annotated EV databases, explicit consideration of preprocessing and batch effects during model construction, and interpretable AI methods to avoid black box predictions that weaken clinician trust.241–243 For PE, a syndrome with multiple causes and phenotypes, future research should integrate secreted miRNA/lncRNA profiles, sFlt-1/PlGF and other protein markers, and quantitative placental MRI indicators into unified models. It should also construct terminal phenotype oriented clinical trial stratification strategies based on EV ncRNA features and develop lightweight bedside models, such as those integrated into obstetric monitoring equipment or out of hospital blood pressure self testing systems, to achieve dynamic risk monitoring.50,241–243
The fourth direction, which is most directly related to nanomaterials, is to develop more placenta targeted, safe, and manufacturable nanomedicine platforms along a continuum from natural EVs and exosome like nanocarriers to LNPs and hybrid carriers. These platforms may transform ncRNAs from disease markers into intervention targets and even therapeutic agents.190,244 At the fundamental level, recent reviews on engineered EVs, EV mimetic nanovesicles, and exosome like nanovesicles suggest that cell engineering, membrane engineering, and topological control can optimize the in vivo distribution, tissue residence time, and loading efficiency of EVs. Moreover, engineered EVs from different sources, including immune cells, mesenchymal stem cells, and tumor cells, have shown favorable pharmacokinetic and pharmacodynamic characteristics in cardiovascular, metabolic, and tumor models. These findings provide a materials science template for adapting similar strategies to pregnancy complications.190,244 At the same time, important progress has been made in lipid nanoparticle (LNP) based treatment of pregnancy related disorders. A placenta tropic VEGF-A mRNA LNP, LNP-55, can restore placental perfusion, lower blood pressure, and improve fetal outcomes in a mouse model, with limited fetal exposure. This finding demonstrates that efficient intervention within the maternal placental safety window can be achieved through rational design of LNP composition and surface ligands.48 Based on these advances, future directions in PE include using placenta specific ligands, such as PLAP antibody fragments or syncytin binding peptides, or smart materials that respond to acidic or oxidative microenvironments within lesions to construct EV membrane coated LNPs or EV LNP hybrid vesicles. These platforms could deliver siRNA, ASO, or miRNA mimics or inhibitors to downregulate harmful factors such as sFlt-1 or restore protective miRNA axes. In addition, combining MSC derived EVs with synthetic nanomaterials may generate multifunctional nanovesicles with both immunoregulatory and anti inflammatory activity and precise drug delivery capacity.199,212 In this process, systematic evaluation of immunogenicity, placental transport, and long term safety of different material systems in pregnant animals will be a key bridge between materials chemistry and clinical obstetrics.169,199
Finally, from the perspective of clinical translation, exosome ncRNA related therapeutics must move beyond proof-of-concept validation and be embedded in rigorous product development and regulatory frameworks. This constitutes the fifth major direction for future research.245–247 On the one hand, registered clinical trials of cell derived EV therapy have rapidly accumulated in tumors, neurological diseases, and immune diseases. Relevant systematic reviews have identified common bottlenecks that hinder translation, including a lack of quantifiable product quality attributes, difficulty in GMP level scale production, and mismatches between clinical endpoint design and mechanism of action. These lessons are particularly important for pregnancy related EV based nanodrugs that are approaching clinical research.246,248 On the other hand, regulatory review of EV diagnostic and drug products emphasizes early communication with regulatory agencies. This includes clarifying during product design whether the product should be classified as an in vitro diagnostic, advanced therapy medicinal product, or combination product. A full process quality control system should also be established for source cells, purification processes, impurity profiles, and stability in accordance with MISEV2023 and EV production standard.199,209,245 In addition, multiple reviews on the clinical application of EVs have suggested that the ideal future path is the joint development of companion diagnostics and therapeutic carriers. This would involve synchronous evaluation of liquid biopsy EV ncRNA panels and corresponding EV or exosome nanodrugs in the same population, allowing biomarker guided adjustment of treatment timing and dose. Maternal long term cardiovascular outcomes should also be incorporated into follow up indicators.245–247 Therefore, from the perspective of future research planning, PE is a suitable pilot disease model for exosome ncRNA therapy because it has a clearly defined target organ, the placenta, a relatively short disease window, and substantial unmet therapeutic needs. If basic researchers, nanomaterial scientists, AI experts, and clinical obstetricians work together under unified standards and regulatory consensus, exosome ncRNA may progress from mechanistic research to clinical practice in this field.208,245,247
Conclusion
EVs associated noncoding RNAs (ncRNAs) provide an important window into placental maternal communication in PE and may connect placental stress with maternal endothelial, immune, inflammatory, and angiogenic dysfunction. Across placental tissue, maternal plasma or serum, urine, amniotic fluid, and other pregnancy related sampling matrices, EV associated miRNAs, lncRNAs, and circRNAs have potential as exploratory biomarkers for PE risk assessment, subtype stratification, and longitudinal monitoring. However, these signals should not be considered clinically actionable until EV isolation is standardized, vesicles are rigorously characterized, RNA quantification is reproducible, findings are validated in independent cohorts, and biofluid signals are clearly linked to tissue origin. The same biology also supports experimental EV based and EV mimetic nanocarrier strategies for ncRNA delivery. Nevertheless, therapeutic translation remains at an early preclinical stage and is constrained by EV heterogeneity, uncertain biodistribution, immunogenicity, maternal and fetal toxicity, placental transfer, manufacturing scalability, quality control, and regulatory uncertainty. Future progress will depend on harmonized EV methodology, longitudinal multicenter cohorts, PE subtype specific analyses, pregnancy relevant safety models, and cautious integration of multi omics and computational tools. Only after these barriers are addressed can EV associated ncRNAs be realistically evaluated as reliable biomarkers or safe nanomedicine-based interventions for PE.
Data Sharing Statement
No new data has been generated, all references are cited in the manuscript.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was funded by Chengdu Medical College High Citation Project Number: CYFY-GQ70.
Disclosure
The authors declare that there are no competing interests associated with the manuscript.
References
1. Bokuda K, Ichihara A. Preeclampsia up to date—what’s going on? Hypertension Research. 2023;46(8):1900–46. doi:10.1038/s41440-023-01323-w
2. Vera-Ponce VJ, Loayza-Castro JA, Ballena-Caicedo J, Valladolid-Sandoval LAM, Zuzunaga-Montoya FE, Gutierrez De Carrillo CI. Global prevalence of preeclampsia, eclampsia, and HELLP syndrome: a systematic review and meta-analysis. Front Reprod Health. 2025;7:1706009. doi:10.3389/frph.2025.1706009
3. Deer E, Herrock O, Campbell N, et al. The role of immune cells and mediators in preeclampsia. Nat Rev Nephrol. 2023;19(4):257–270. doi:10.1038/s41581-022-00670-0
4. Zeisler H, Llurba E, Chantraine F, et al. Predictive value of the sFlt-1:PlGF ratio in women with suspected preeclampsia. N Engl J Med. 2016;374(1):13–22. doi:10.1056/NEJMoa1414838
5. Yang YF, Li ZH, Liu TH, et al. Evaluating the role of DNA methyltransferases in trophoblast fusion and preeclampsia development: insights from methylation-regulated genes. FASEB J Off Publ Fed Am Soc Exp Biol. 2024;38(13):e23706. doi:10.1096/fj.202302705R
6. Zhu Y, Zhang Y, Hu W, et al. First-trimester pan-immune-inflammation value predicts preeclampsia in a dose-dependent linear pattern. BMC Pregnancy Childbirth. 2025;25:1250. doi:10.1186/s12884-025-08406-1
7. Roberts JM. Preeclampsia epidemiology(ies) and pathophysiology(ies). Best Pract Res Clin Obstet Gynaecol. 2024;94:102480. doi:10.1016/j.bpobgyn.2024.102480
8. Karadzov Orlic N, Joksić I. Preeclampsia pathogenesis and prediction - where are we now: the focus on the role of galectins and miRNAs. Hypertens Pregnancy. 2025;44(1):2470626. doi:10.1080/10641955.2025.2470626
9. Ghafourian M, Mahdavi R, Akbari Jonoush Z, et al. The implications of exosomes in pregnancy: emerging as new diagnostic markers and therapeutics targets. Cell Commun Signal CCS. 2022;20(1):51. doi:10.1186/s12964-022-00853-z
10. Gonzalez Fernandez J, Moncayo Arlandi J, Ochando A, Simon C, Vilella F. The role of extracellular vesicles in intercellular communication in human reproduction. Clinical Science. 2023;137(3):281–301. doi:10.1042/CS20220793
11. Kumar MA, Baba SK, Sadida HQ, et al. Extracellular vesicles as tools and targets in therapy for diseases. Signal Transduct Target Ther. 2024;9(1):27. doi:10.1038/s41392-024-01735-1
12. Salomon C, Rice GE. Role of exosomes in placental homeostasis and pregnancy disorders. Prog Mol Biol Transl Sci. 2017;145:163–179. doi:10.1016/bs.pmbts.2016.12.006
13. Zhang Y, Zhang J, Chen S, et al. Unveiling the network regulatory mechanism of ncRNAs on the ferroptosis pathway: implications for preeclampsia. Int J Womens Health. 2024;16:1633–1651. doi:10.2147/IJWH.S485653
14. Nishi K, Modi D. Placental exosomes in pregnancy and preeclampsia. Am J Reprod Immunol N Y N. 2024;91(5):e13857. doi:10.1111/aji.13857
15. Mitchell MD, Peiris HN, Kobayashi M, et al. Placental exosomes in normal and complicated pregnancy. Am J Obstet Gynecol. 2015;213(4 Suppl):S173–181. doi:10.1016/j.ajog.2015.07.001
16. Zhong XL, Huang Y, Du Y, et al. Unlocking the therapeutic potential of exosomes derived from nasal olfactory mucosal mesenchymal stem cells: restoring synaptic plasticity, neurogenesis, and neuroinflammation in schizophrenia. Schizophr Bull. 2023;50(3):600–614. doi:10.1093/schbul/sbad172
17. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367(6478):eaau6977. doi:10.1126/science.aau6977
18. Shan Y, Hou B, Wang J, Chen A, Liu S. Exploring the role of exosomal MicroRNAs as potential biomarkers in preeclampsia. Front Immunol. 2024;15:1385950. doi:10.3389/fimmu.2024.1385950
19. Zhang L, Tian X, Liu M, et al. Exosomal circDNAJB6 derived from decidual macrophages promotes preeclampsia progression via the miR-670-5p/TOB2 axis and by subsequently regulating the PPARγ/NF-κB pathway. J Transl Med. 2025;23(1):1154. doi:10.1186/s12967-025-07220-9
20. Chen A, Yu R, Jiang S, Xia Y, Chen Y. Recent advances of MicroRNAs, long non-coding RNAs, and circular RNAs in preeclampsia. Front Physiol. 2021;12:659638. doi:10.3389/fphys.2021.659638
21. Hayder H, Shan Y, Chen Y, O’Brien JA, Peng C. Role of microRNAs in trophoblast invasion and spiral artery remodeling: implications for preeclampsia. Front Cell Dev Biol. 2022;10:995462. doi:10.3389/fcell.2022.995462
22. Nair S, Salomon C. Extracellular vesicles and their immunomodulatory functions in pregnancy. Semin Immunopathol. 2018;40(5):425–437. doi:10.1007/s00281-018-0680-2
23. Gong RQ, Nuh AM, Cao HS, Ma M. Roles of exosomes-derived lncRNAs in preeclampsia. Eur J Obstet Gynecol Reprod Biol. 2021;263:132–138. doi:10.1016/j.ejogrb.2021.06.015
24. Biró O, Alasztics B, Molvarec A, Joó J, Nagy B, Rigó JJ. Various levels of circulating exosomal total-miRNA and miR-210 hypoxamiR in different forms of pregnancy hypertension. Pregnancy Hypertens. 2017;10:207–212. doi:10.1016/j.preghy.2017.09.002
25. Salomon C, Torres MJ, Kobayashi M, et al. A gestational profile of placental exosomes in maternal plasma and their effects on endothelial cell migration. PLoS One. 2014;9(6):e98667. doi:10.1371/journal.pone.0098667
26. Yáñez-Mó M, Siljander PRM, Andreu Z, et al. Biological properties of extracellular vesicles and their physiological functions. J Extracell Vesicles. 2015;4:27066. doi:10.3402/jev.v4.27066
27. Pillay P, Maharaj N, Moodley J, Mackraj I. Placental exosomes and pre-eclampsia: maternal circulating levels in normal pregnancies and, early and late onset pre-eclamptic pregnancies. Placenta. 2016;46:18–25. doi:10.1016/j.placenta.2016.08.078
28. Vickers KC, Palmisano BT, Shoucri BM, Shamburek RD, Remaley AT. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat Cell Biol. 2011;13(4):423–433. doi:10.1038/ncb2210
29. Hromadnikova I, Kotlabova K, Ivankova K, Krofta L. First trimester screening of circulating C19MC microRNAs and the evaluation of their potential to predict the onset of preeclampsia and IUGR. PLoS One. 2017;12(2):e0171756. doi:10.1371/journal.pone.0171756
30. Rana S, Lemoine E, Granger JP, Karumanchi SA. Preeclampsia: pathophysiology, challenges, and perspectives. Circ Res. 2019;124(7):1094–1112. doi:10.1161/CIRCRESAHA.118.313276
31. Brown MA, Magee LA, Kenny LC, et al. Hypertensive disorders of pregnancy: ISSHP classification, diagnosis, and management recommendations for international practice. Hypertens Dallas Tex 1979. 2018;72(1):24–43. doi:10.1161/HYPERTENSIONAHA.117.10803
32. Burton GJ, Redman CW, Roberts JM, Moffett A. Pre-eclampsia: pathophysiology and clinical implications. BMJ. 2019;
33. Mateescu B, Kowal EJK, van Balkom BWM, et al. Obstacles and opportunities in the functional analysis of extracellular vesicle RNA – an ISEV position paper. J Extracell Vesicles. 2017;6(1):1286095. doi:10.1080/20013078.2017.1286095
34. Anton L, Olarerin-George AO, Schwartz N, et al. miR-210 Inhibits trophoblast invasion and is a serum biomarker for preeclampsia. Am J Pathol. 2013;183(5):1437–1445. doi:10.1016/j.ajpath.2013.07.021
35. Guerby P, Tasta O, Swiader A, et al. Role of oxidative stress in the dysfunction of the placental endothelial nitric oxide synthase in preeclampsia. Redox Biol. 2021;40:101861. doi:10.1016/j.redox.2021.101861
36. Lener T, Gimona M, Aigner L, et al. Applying extracellular vesicles based therapeutics in clinical trials – an ISEV position paper. J Extracell Vesicles. 2015;4. doi:10.3402/jev.v4.30087
37. Menjoge AR, Rinderknecht A, Navath RS, et al. Transfer of PAMAM dendrimers across human placenta: prospects of its use as drug carrier during pregnancy. J Control Release off J Control Release Soc. 2011;150(3):326–338. doi:10.1016/j.jconrel.2010.11.023
38. Ortega MA, Fraile-Martínez O, García-Montero C, et al. Unfolding the role of placental-derived extracellular vesicles in pregnancy: from homeostasis to pathophysiology. Front Cell Dev Biol. 2022;10:1060850. doi:10.3389/fcell.2022.1060850
39. Khalilian S, Bijanvand A, Abedinlou H, Ghafouri-Fard S. A review on the role of miR-210 in human disorders. Pathol - Res Pract. 2023;241:154244. doi:10.1016/j.prp.2022.154244
40. Salimbayeva D, Kurmanova A, Nurmakova A, Smailov M, Kypshakbayeva Z. Placental biomarkers of preeclampsia: systematic review. Bratisl Med J. 2025;126(8):2001–2014. doi:10.1007/s44411-025-00198-2
41. Chan SY, Zhang YY, Hemann C, Mahoney CE, Zweier JL, Loscalzo J. MicroRNA-210 controls mitochondrial metabolism during hypoxia by repressing the iron-sulfur cluster assembly proteins ISCU1/2. Cell Metab. 2009;10(4):273–284. doi:10.1016/j.cmet.2009.08.015
42. Phipps EA, Thadhani R, Benzing T, Karumanchi SA. Pre-eclampsia: pathogenesis, novel diagnostics and therapies. Nat Rev Nephrol. 2019;15(5):275–289. doi:10.1038/s41581-019-0119-6
43. McCarthy FP, Kingdom JC, Kenny LC, Walsh SK. Animal models of preeclampsia; uses and limitations. Placenta. 2011;32(6):413–419. doi:10.1016/j.placenta.2011.03.010
44. Frazier S, McBride MW, Mulvana H, Graham D. From animal models to patients: the role of placental microRNAs, miR-210, miR-126, and miR-148a/152 in preeclampsia. Clin Sci Lond Engl. 2020;134(8):1001–1025. doi:10.1042/CS20200023
45. Oancea M, Mihu D, Braicu C, et al. MicroRNAs in preeclampsia: an overview of biomarkers and potential therapeutic targets. Int J Mol Sci. 2025;26(12):5607. doi:10.3390/ijms26125607
46. Wu D, Zhou B, Hong L, et al. Trophoblast cell-derived extracellular vesicles regulate the polarization of decidual macrophages by carrying miR-141-3p in the pathogenesis of preeclampsia. Sci Rep. 2024;14:24529. doi:10.1038/s41598-024-76563-y
47. Ghosh S, Thamotharan S, Fong J, Lei MYY, Janzen C, Devaskar SU. Circulating extracellular vesicular microRNA signatures in early gestation show an association with subsequent clinical features of pre-eclampsia. Sci Rep. 2024;14(1):16770. doi:10.1038/s41598-024-64057-w
48. Swingle KL, Hamilton AG, Safford HC, et al. Placenta-tropic VEGF mRNA lipid nanoparticles ameliorate murine pre-eclampsia. Nature. 2025;637(8045):412–421. doi:10.1038/s41586-024-08291-2
49. Peng P, Song H, Xie C, et al. miR-146a-5p-mediated suppression on trophoblast cell progression and epithelial-mesenchymal transition in preeclampsia. Biol Res. 2021;54(1):30. doi:10.1186/s40659-021-00351-5
50. Kim HI, Park J, Zhu Y, Wang X, Han Y, Zhang D. Recent advances in extracellular vesicles for therapeutic cargo delivery. Exp Mol Med. 2024;56(4):836–849. doi:10.1038/s12276-024-01201-6
51. Fu X, Wu M, Chen Y, et al. The expression profile of plasmatic exosomal lncRNAs in early-onset preeclampsia by sequencing. Am J Transl Res. 2022;14(6):3806–3823.
52. Liu H, Cai X, Liu J, Zhang F, He A, Li R. The MEG3 lncRNA promotes trophoblastic cell growth and invasiveness in preeclampsia by acting as a sponge for miR-21, which regulates BMPR2 levels. Eur J Histochem EJH. 2021;65(4):3323. doi:10.4081/ejh.2021.3323
53. Maliha E, Pinti A, Bassim P, Toumi H, El Hage R. Composite indices of femoral neck strength in young adult male handball players. J Clin Densitom off J Int Soc Clin Densitom. 2022;25(4):637–640. doi:10.1016/j.jocd.2021.11.012
54. Granitzer S, Widhalm R, Ellinger I, et al. LAT1-NRF2 axis controls sFlt-1/PlGF imbalance and oxidative stress in preeclampsia. Nat Commun. 2025;16:9112. doi:10.1038/s41467-025-64160-0
55. Selvakumar SC, Preethi KA, Ross K, Sekar D. The emerging role of microRNA-based therapeutics in the treatment of preeclampsia. Placenta. 2024;158:38–47. doi:10.1016/j.placenta.2024.09.018
56. Cao M, Wen J, Bu C, et al. Differential circular RNA expression profiles in umbilical cord blood exosomes from preeclampsia patients. BMC Pregnancy Childbirth. 2021;21(1):303. doi:10.1186/s12884-021-03777-7
57. Shu C, Xu P, Han J, Han S, He J. Upregulation of circRNA hsa_circ_0008726 in pre-eclampsia inhibits trophoblast migration, invasion, and EMT by regulating miR-345-3p/RYBP axis. Reprod Sci Thousand Oaks Calif. 2022;29(10):2829–2841. doi:10.1007/s43032-021-00804-y
58. Hula N, Escalera D, Goulopoulou S. Extracellular vesicles in preeclampsia: drivers of vascular dysfunction and inflammation. Am J Physiol Heart Circ Physiol. 2025;329(6):H1560–H1574. doi:10.1152/ajpheart.00584.2025
59. Pan H, Ouyang B, Zhang H, Zhao C. Non-coding RNAs: the architects of placental development and pregnancy success. Mol Genet Genomics MGG. 2025;300(1):39. doi:10.1007/s00438-025-02244-8
60. Winter HE, Murrieta-Coxca JM, Álvarez D, et al. Enhanced capture of preeclampsia-derived extracellular vesicles from maternal plasma by monocytes and T lymphocytes. J Reprod Immunol. 2025;167:104417. doi:10.1016/j.jri.2024.104417
61. Yu MY, Jia HJ, Zhang J, Ran GH, Liu Y, Yang XH. Exosomal miRNAs-mediated macrophage polarization and its potential clinical application. Int Immunopharmacol. 2023;117:109905. doi:10.1016/j.intimp.2023.109905
62. Liu X, Fei H, Yang C, et al. Trophoblast-Derived extracellular vesicles promote preeclampsia by regulating macrophage polarization. Hypertens Dallas Tex 1979. 2022;79(10):2274–2287. doi:10.1161/HYPERTENSIONAHA.122.19244
63. Xing Y, Kang L, Chen L, Li Y, Lu D. Research progress of exosomes in pathogenesis and treatment of preeclampsia. J Obstet Gynaecol Res. 2024;50(12):2183–2194. doi:10.1111/jog.16106
64. Cao SQ, Jiang TX, Guo YY, Lin R, Lin L. MiR-519d-3p from placenta-derived exosomes induce immune intolerance regulating immune cells, contributing to the pathogenesis of preeclampsia. Immunol Invest. 2025;54(4):522–543. doi:10.1080/08820139.2025.2450234
65. David M, Maharaj N. The immune-modulatory dynamics of exosomes in preeclampsia. Arch Gynecol Obstet. 2025;311(6):1477–1487. doi:10.1007/s00404-025-08013-7
66. Nair S, Salomon C. Extracellular vesicles as critical mediators of maternal-fetal communication during pregnancy and their potential role in maternal metabolism. Placenta. 2020;98:60–68. doi:10.1016/j.placenta.2020.06.011
67. Tartaglia E, Bordoni V, Oliva A, et al. T helper profile in pregnant women recovered from COVID-19. J Reprod Immunol. 2022;153:103661. doi:10.1016/j.jri.2022.103661
68. Mora-Palazuelos C, Villegas-Mercado CE, Avendaño-Félix M, et al. The role of ncRNAs in the immune dysregulation of preeclampsia. Int J Mol Sci. 2023;24(20):15215. doi:10.3390/ijms242015215
69. Shi Y, Shen F, Chen X, Sun M, Zhang P. Current understanding of circular RNAs in preeclampsia. Hypertens Res Off J Jpn Soc Hypertens. 2024;47(6):1607–1619. doi:10.1038/s41440-024-01675-x
70. Bao D, Zhuang C, Jiao Y, Yang L. The possible involvement of circRNA DMNT1/p53/JAK/STAT in gestational diabetes mellitus and preeclampsia. Cell Death Discov. 2022;8:121. doi:10.1038/s41420-022-00913-w
71. Kamrani A, Akbari M, Heris JA, Yousefi M. Regulation of PI3K/Akt in preeclampsia: an examination of its pathological role and therapeutic potential. Naunyn Schmiedebergs Arch Pharmacol. 2026;399(1):33–46. doi:10.1007/s00210-025-04472-6
72. Gianni-Barrera R, Di Maggio N, Melly L, et al. Therapeutic vascularization in regenerative medicine. Stem Cells Transl Med. 2020;9(4):433–444. doi:10.1002/sctm.19-0319
73. Chen Z, Zhang W, Wu M, Huang H, Zou L, Luo Q. Pathogenic mechanisms of preeclampsia with severe features implied by the plasma exosomal mirna profile. Bioengineered. 2021;12(2):9140–9149. doi:10.1080/21655979.2021.1993717
74. Motta-Mejia C, Kandzija N, Zhang W, et al. Placental vesicles carry active endothelial nitric oxide synthase and their activity is reduced in preeclampsia. Hypertens Dallas Tex 1979. 2017;70(2):372–381. doi:10.1161/HYPERTENSIONAHA.117.09321
75. Wang Y, Du X, Wang J. Transfer of miR-15a-5p by placental exosomes promotes pre-eclampsia progression by regulating PI3K/AKT signaling pathway via CDK1. Mol Immunol. 2020;128:277–286. doi:10.1016/j.molimm.2020.10.019
76. Lin J, Huang X, Zhang J, et al. Amniotic fluid-derived exosomal miR-146a-5p ameliorates preeclampsia phenotypes by inhibiting HIF-1α/FLT-1 expression. Placenta. 2025;162:35–44. doi:10.1016/j.placenta.2025.02.013
77. Xueya Z, Yamei L, Sha C, et al. Exosomal encapsulation of miR-125a-5p inhibited trophoblast cell migration and proliferation by regulating the expression of VEGFA in preeclampsia. Biochem Biophys Res Commun. 2020;525(3):646–653. doi:10.1016/j.bbrc.2020.02.137
78. JoshuaJ F, DanielR M, EvanN P, JamesS C, OliviaJ H, AnthonyV P. Mitochondrial isolation, cryopreservation and preliminary biochemical characterisation from placental cytotrophoblast and syncytiotrophoblast. Placenta. 2019;82:1–4. doi:10.1016/j.placenta.2019.05.004
79. Xu P, Ma Y, Wu H, Wang YL. Placenta-Derived MicroRNAs in the pathophysiology of human pregnancy. Front Cell Dev Biol. 2021;9:646326. doi:10.3389/fcell.2021.646326
80. Wang Z, Liu D, Dai Y, et al. Elevated placental microRNA-155 is a biomarker of a preeclamptic subtype. Hypertens Dallas Tex 1979. 2023;80(2):370–384. doi:10.1161/HYPERTENSIONAHA.122.19914
81. Li X, Yang R, Xu Y, Zhang Y. Circ_0001438 participates in the pathogenesis of preeclampsia via the circ_0001438/miR-942/NLRP3 regulatory network. Placenta. 2021;104:40–50. doi:10.1016/j.placenta.2020.11.005
82. Mishima S, Mitsui T, Tani K, et al. Endothelin-1 production via placental (pro)renin receptor in a mouse model of preeclampsia. Placenta. 2023;138:44–50. doi:10.1016/j.placenta.2023.05.002
83. ling JL, lin YD, Han Q, le ZH, Pan M, ying YJ. LncRNA-NEAT1 blocks the Wnt/β-catenin signaling pathway by targeting miR-217 to inhibit trophoblast cell migration and invasion. J Assist Reprod Genet. 2024;41(8):2107–2115. doi:10.1007/s10815-024-03124-7
84. Verdonk K, Saleh L, Lankhorst S, et al. Association studies suggest a key role for endothelin-1 in the pathogenesis of preeclampsia and the accompanying renin-angiotensin-aldosterone system suppression. Hypertens Dallas Tex 1979. 2015;65(6):1316–1323. doi:10.1161/HYPERTENSIONAHA.115.05267
85. Sutton EF, Gemmel M, Powers RW. Nitric oxide signaling in pregnancy and preeclampsia. Nitric Oxide Biol Chem. 2020;95:55–62. doi:10.1016/j.niox.2019.11.006
86. Blakey H, Sun R, Xie L, et al. Pre-eclampsia is associated with complement pathway activation in the maternal and fetal circulation, and placental tissue. Pregnancy Hypertens. 2023;32:43–49. doi:10.1016/j.preghy.2023.04.001
87. Pierik E, Prins JR, Van Goor H, et al. Dysregulation of complement activation and placental dysfunction: a potential target to treat preeclampsia? Front Immunol. 2019;10:3098. doi:10.3389/fimmu.2019.03098
88. Kumar V, Stewart JH. The complement system in human pregnancy and preeclampsia. Front Immunol. 2025;16:1617140. doi:10.3389/fimmu.2025.1617140
89. Gardikioti A, Venou TM, Gavriilaki E, et al. Molecular advances in preeclampsia and HELLP syndrome. Int J Mol Sci. 2022;23(7):3851. doi:10.3390/ijms23073851
90. Hernández González LL, Mayoral L P-C, Hernández-Huerta MT, et al. Targeting neutrophil extracellular trap formation: exploring promising pharmacological strategies for the treatment of preeclampsia. Pharm Basel Switz. 2024;17(5):605. doi:10.3390/ph17050605
91. Moodley M, Moodley J, Naicker T. The role of neutrophils and their extracellular traps in the synergy of pre-eclampsia and HIV infection. Curr Hypertens Rep. 2020;22(6):41. doi:10.1007/s11906-020-01047-z
92. Ramos A, Youssef L, Molina P, et al. Circulating extracellular vesicles and neutrophil extracellular traps contribute to endothelial dysfunction in preeclampsia. Front Immunol. 2024;15:1488127. doi:10.3389/fimmu.2024.1488127
93. Rosenfeld CS. Placenta extracellular vesicles: messengers connecting maternal and fetal systems. Biomolecules. 2024;14(8):995. doi:10.3390/biom14080995
94. Han C, Han L, Huang P, Chen Y, Wang Y, Xue F. Syncytiotrophoblast-Derived extracellular vesicles in pathophysiology of preeclampsia. Front Physiol. 2019;10:1236. doi:10.3389/fphys.2019.01236
95. Awoyemi T, Cerdeira AS, Zhang W, et al. Preeclampsia and syncytiotrophoblast membrane extracellular vesicles (STB-EVs). Clin Sci Lond Engl 1979. 2022;136(24):1793–1807. doi:10.1042/CS20220149
96. Morales-Prieto DM, Favaro RR, Markert UR. Placental miRNAs in feto-maternal communication mediated by extracellular vesicles. Placenta. 2020;102:27–33. doi:10.1016/j.placenta.2020.07.001
97. Lv Y, Lu C, Ji X, et al. Roles of microRNAs in preeclampsia. J Cell Physiol. 2019;234(2):1052–1061. doi:10.1002/jcp.27291
98. O’Brien K, Breyne K, Ughetto S, Laurent LC, Breakefield XO. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat Rev Mol Cell Biol. 2020;21(10):585–606. doi:10.1038/s41580-020-0251-y
99. Lv J, Xiong X. Extracellular vesicle microRNA: a promising biomarker and therapeutic target for respiratory diseases. Int J Mol Sci. 2024;25(17):9147. doi:10.3390/ijms25179147
100. Nair S, Ormazabal V, Lappas M, McIntyre HD, Salomon C. Extracellular vesicles and their potential role inducing changes in maternal insulin sensitivity during gestational diabetes mellitus. Am J Reprod Immunol N Y N. 2021;85(2):e13361. doi:10.1111/aji.13361
101. Mirzakhani H, Handy DE, Lu Z, et al. Integration of circulating microRNAs and transcriptome signatures identifies early-pregnancy biomarkers of preeclampsia. Clin Transl Med. 2023;13(11):e1446. doi:10.1002/ctm2.1446
102. Murugesan S, Hussey H, Saravanakumar L, et al. Extracellular vesicles from women with severe preeclampsia impair vascular endothelial function. Anesth Analg. 2022;134(4):713–723. doi:10.1213/ANE.0000000000005812
103. Hu XQ, Zhang L. MicroRNAs in uteroplacental vascular dysfunction. Cells. 2019;8(11):1344. doi:10.3390/cells8111344
104. Barnes MVC, Pantazi P, Holder B. Circulating extracellular vesicles in healthy and pathological pregnancies: a scoping review of methodology, rigour and results. J Extracell Vesicles. 2023;12(11):e12377. doi:10.1002/jev2.12377
105. Wang Z, Zhao G, Zeng M, Feng W, Liu J. Overview of extracellular vesicles in the pathogenesis of preeclampsia†. Biol Reprod. 2021;105(1):32–39. doi:10.1093/biolre/ioab060
106. Gerede A, Stavros S, Danavasi M, et al. MicroRNAs in preeclampsia: bridging diagnosis and treatment. J Clin Med. 2025;14(6). doi:10.3390/jcm14062003
107. Pillay P, Vatish M, Duarte R, Moodley J, Mackraj I. Exosomal microRNA profiling in early and late onset preeclamptic pregnant women reflects pathophysiology. Int J Nanomed. 2019;14:5637–5657. doi:10.2147/IJN.S208865
108. Pillay P, Moodley K, Moodley J, Mackraj I. Placenta-derived exosomes: potential biomarkers of preeclampsia. Int J Nanomed. 2017;12:8009–8023. doi:10.2147/IJN.S142732
109. Awoyemi T, Jiang S, Rahbar M, et al. MicroRNA analysis of medium/large placenta extracellular vesicles in normal and preeclampsia pregnancies. Front Cardiovasc Med. 2024;11:1371168. doi:10.3389/fcvm.2024.1371168
110. Aharon A, Rebibo-Sabbah A, Ahmad RS, et al. Associations of maternal and placental extracellular vesicle miRNA with preeclampsia. Front Cell Dev Biol. 2023;11:1080419. doi:10.3389/fcell.2023.1080419
111. Hromadnikova I, Dvorakova L, Kotlabova K, Krofta L. The prediction of gestational hypertension, preeclampsia and fetal growth restriction via the first trimester screening of plasma exosomal C19MC microRNAs. Int J Mol Sci. 2019;20(12):2972. doi:10.3390/ijms20122972
112. Gan L, Liu Z, Wei M, et al. MiR-210 and miR-155 as potential diagnostic markers for pre-eclampsia pregnancies. Medicine. 2017;96(28):e7515. doi:10.1097/MD.0000000000007515
113. Yin L, Zhang Y, Fu G, et al. Knowledge mapping of exosomes in preeclampsia: a bibliometric analysis (2008-2023). Front Endocrinol. 2025;16:1546554. doi:10.3389/fendo.2025.1546554
114. Illarionov RA, Maltseva AR, Pachuliia OV, et al. Urinary miRNA expression in pre-eclampsia during early and mid-pregnancy. Non Cod RNA. 2024;10(6):61. doi:10.3390/ncrna10060061
115. Phipps EA, Khankin EV. Extracellular vesicles in preeclampsia: evolving contributors to proteinuria. J Am Soc Nephrol JASN. 2017;28(11):3135–3137. doi:10.1681/ASN.2017070789
116. He X, Liu S, Wang Y, et al. Extracellular vesicles in pregnancy-related disorders: from mechanisms to clinical implications. Sci China Life Sci. 2025. doi:10.1007/s11427-024-2937-4
117. Grossini E, Surico D, Venkatesan S, Ola Pour MM, Aquino CI, Remorgida V. Extracellular vesicles and pregnancy-related hypertensive disorders: a descriptive review on the possible implications “from bench to bedside. Biology. 2025;14(3):240. doi:10.3390/biology14030240
118. Zhaoer Y, Mingming G, Wei Z, Dan Y, Yating Q, Ruizhe J. Extracellular vesicles for the treatment of preeclampsia. Tissue Cell. 2022;77:101860. doi:10.1016/j.tice.2022.101860
119. Gál L, Fóthi Á, Orosz G, Nagy S, Than NG, Orbán TI. Exosomal small RNA profiling in first-trimester maternal blood explores early molecular pathways of preterm preeclampsia. Front Immunol. 2024;15:1321191. doi:10.3389/fimmu.2024.1321191
120. Wang W, Zhang W, Chen L, et al. Identification of differentially expressed miRNAs in plasma exosomes from patients with early-onset pre-eclampsia using next generation sequencing. Heliyon. 2024;10(3):e24543. doi:10.1016/j.heliyon.2024.e24543
121. Tang C, Hu W. Biomarkers and diagnostic significance of non-coding RNAs in extracellular vesicles of pathologic pregnancy. J Assist Reprod Genet. 2024;41(10):2569–2584. doi:10.1007/s10815-024-03268-6
122. Wang Y, Zhang M. Urinary exosomes: a promising biomarker for disease diagnosis. Lab Med. 2023;54(2):115–125. doi:10.1093/labmed/lmac087
123. Zhou Z, Zhang D, Wang Y, et al. Urinary exosomes: a promising biomarker of drug-induced nephrotoxicity. Front Med. 2023;10:1251839. doi:10.3389/fmed.2023.1251839
124. Gaudio C, D’Arpino E, Stefani S, et al. Urinary exosomes in nephrology: a new frontier for diagnosis and prognosis of kidney diseases. Int J Mol Sci. 2025;26(17):8679. doi:10.3390/ijms26178679
125. Zhou J, Gong G, Tan H, et al. Urinary microRNA-30a-5p is a potential biomarker for ovarian serous adenocarcinoma. Oncol Rep. 2015;33(6):2915–2923. doi:10.3892/or.2015.3937
126. Kothandan VK, Ouyang Y, Sadovsky E, et al. A multi-platform assessment of extracellular vesicles from the plasma and urine of women with preeclampsia. Placenta. 2025;166:96–102. doi:10.1016/j.placenta.2024.12.014
127. Atukorala I, Hannan N, Hui L. Immersed in a reservoir of potential: amniotic fluid-derived extracellular vesicles. J Transl Med. 2024;22(1):348. doi:10.1186/s12967-024-05154-2
128. Gebara N, Scheel J, Skovronova R, et al. Single extracellular vesicle analysis in human amniotic fluid shows evidence of phenotype alterations in preeclampsia. J Extracell Vesicles. 2022;11(5):e12217. doi:10.1002/jev2.12217
129. Khan A, Raza F, He N. Nanoscale extracellular vesicle-enabled liquid biopsy: advances and challenges for lung cancer detection. Micromachines. 2024;15(10):1181. doi:10.3390/mi15101181
130. Wang D, Shen Y, Qian H, Jiang J, Xu W. Emerging advanced approaches for liquid biopsy: in situ nucleic acid assays of extracellular vesicles. Theranostics. 2024;14(19):7309–7332. doi:10.7150/thno.102437
131. Duley L, Meher S, Abalos E. Management of pre-eclampsia. BMJ. 2006;332(7539):463–468. doi:10.1136/bmj.332.7539.463
132. Poon LC, Shennan A, Hyett JA, et al. The International Federation of Gynecology and Obstetrics (FIGO) Initiative on Preeclampsia (PE): a pragmatic guide for first trimester screening and prevention. Int J Gynaecol Obstet Off Organ Int Fed Gynaecol Obstet. 2019;145(Suppl 1):1–33. doi:10.1002/ijgo.12802
133. Ma Y, Chiang YW, Becker TM, Hyett J. Cell-Based and cell-free non-invasive prenatal analysis of preeclampsia: an updated review of liquid biopsy. Biomedicines. 2026;14(4):851. doi:10.3390/biomedicines14040851
134. Cretu OE, Dirlau AA, Neacsu AV, Nenciu AE, Ceausu I. The role of microRNA-210 in the pathogenesis and diagnosis of preeclampsia-a systematic review. J Clin Med. 2025;14(21):7593. doi:10.3390/jcm14217593
135. Bowman-Gibson S, Chandiramani C, Stone ML, et al. Streamlined analysis of maternal plasma indicates small extracellular vesicles are significantly elevated in early-onset preeclampsia. Reprod Sci Thousand Oaks Calif. 2024;31(9):2771–2782. doi:10.1007/s43032-024-01591-y
136. Cen Y, Bao J, Zhou T, Yin F, Xu Z, Zhou M. Maternal plasma microRNA signatures for early detection of early-onset preeclampsia. Int J Biol Macromol. 2025;320(Pt 1):145654. doi:10.1016/j.ijbiomac.2025.145654
137. Luo YW, Liu CG, Kirby JA, Chu C, Zang D, Chen J. The emerging role of extracellular vesicle-derived lncRNAs and circRNAs in tumor and mesenchymal stem cells: the biological functions and potential for clinical application. Cancers. 2025;17(13):2186. doi:10.3390/cancers17132186
138. Cooke WR, Jones GD, Redman CW, Vatish M. Small RNAs in the pathogenesis of preeclampsia. Placenta. 2024;157:21–27. doi:10.1016/j.placenta.2024.06.009
139. Morey R, Poling L, Srinivasan S, et al. Discovery and verification of extracellular microRNA biomarkers for diagnostic and prognostic assessment of preeclampsia at triage. Sci Adv. 2023;9(51):eadg7545. doi:10.1126/sciadv.adg7545
140. Lewis E, Lee SJ, Park HR. Placenta-derived extracellular vesicles in fetal health: emerging insights into brain development and environmental interactions. Exp Mol Med. 2025;57(12):2729–2738. doi:10.1038/s12276-025-01601-2
141. Wang M, Zheng L, Ma S, Lin R, Li J, Yang S. Biogenesis and function of exosome lncRNAs and their role in female pathological pregnancy. Front Endocrinol. 2023;14. doi:10.3389/fendo.2023.1191721
142. A KP, Sekar D. Advancements in microRNA-based electrochemical biosensors for preeclampsia detection. Hypertens Res Off J Jpn Soc Hypertens. 2024;47(6):1752–1754. doi:10.1038/s41440-024-01672-0
143. Grezzana GB, Stein AT, Markoski MM. Towards early diagnosis of preeclampsia: emerging roles of circulating microRNAs. Eur J Obstet Gynecol Reprod Biol. 2026;316:114822. doi:10.1016/j.ejogrb.2025.114822
144. Su S, Yang F, Zhong L, Pang L. Circulating noncoding RNAs as early predictive biomarkers in preeclampsia: a diagnostic meta-analysis. Reprod Biol Endocrinol RBE. 2021;19:177. doi:10.1186/s12958-021-00852-8
145. Li Y, Yu Y, Li D, Li L. Exosomal encapsulation of miR-3198 promotes proliferation and migration of trophoblasts in preeclampsia. J Assist Reprod Genet. 2024;41(5):1403–1416. doi:10.1007/s10815-024-03104-x
146. Muskan M, Abeysinghe P, Cecchin R, Branscome H, Morris KV, Kashanchi F. Therapeutic potential of RNA-enriched extracellular vesicles: the next generation in RNA delivery via biogenic nanoparticles. Mol Ther. 2024;32(9):2939–2949. doi:10.1016/j.ymthe.2024.02.025
147. Tang P, Wang J, Wang L, Li S, Cao Y. High throughput sequencing analysis of exosomal miRNA expressions in early-onset preeclampsia patients. Clin Lab. 2023;69(2). doi:10.7754/Clin.Lab.2022.220430
148. Chen Y, Zhang Y, Xie S, Zhou X, Zhu L, Cao Y. Establishment of a placental lncRNA-mRNA expression network for early-onset preeclampsia. BMC Pregnancy Childbirth. 2024;24:329. doi:10.1186/s12884-024-06481-4
149. Jiang H, Meng T, Li Z. Role of circular RNAs in preeclampsia (Review). Exp Ther Med. 2024;28(4):372. doi:10.3892/etm.2024.12661
150. Jia N, Li J. Role of circular RNAs in preeclampsia. Dis Markers. 2019;2019:7237495. doi:10.1155/2019/7237495
151. Vafadar A, Heidari Z, Bolbolizadeh P, Razmjoue D, Ehtiati S, Savardashtaki A. Exosomes in preeclampsia. Clin Chim Acta Int J Clin Chem. 2026;578:120549. doi:10.1016/j.cca.2025.120549
152. Margiana R. Mesenchymal stem cell-derived exosomes in preeclampsia: a next-generation therapeutic tool. Cell Biochem Funct. 2024;42(1):e3908. doi:10.1002/cbf.3908
153. Liu J, Zhao Q, Zhu Y. Non-coding RNAs as diagnostic biomarkers for preeclampsia: a systematic review and meta-analysis. BMC Pregnancy Childbirth. 2025;25(1):949. doi:10.1186/s12884-025-08116-8
154. Wu S, Li Q, Liu X, et al. Placental exosomal miR-125b triggered endothelial barrier injury in preeclampsia. Placenta. 2023;137:31–37. doi:10.1016/j.placenta.2023.04.006
155. Fang Y, Fan M, Li H, et al. CPMOAD: comprehensive preeclampsia multi-omics analysis database. J Transl Med. 2025;23(1):1026. doi:10.1186/s12967-025-07094-x
156. Cao C, Saxena R, Gray KJ. Placental origins of preeclampsia: insights from multi-omic studies. Int J Mol Sci. 2024;25(17):9343. doi:10.3390/ijms25179343
157. Li Z, Tao M, Huang M, et al. Quantification of placental extracellular vesicles in different pregnancy status via single particle analysis method. Clin Chim Acta Int J Clin Chem. 2023;539:266–273. doi:10.1016/j.cca.2022.12.021
158. Wei Z, Zhou N, Jing M, et al. SVAtlas: a comprehensive single extracellular vesicle omics resource. Nucleic Acids Res. 2025:gkaf1189. doi:10.1093/nar/gkaf1189
159. Mumcu A, Sarıdoğan E, Düz SA, et al. Multi-omics analysis of placental metabolomics and transcriptomics datasets reveals comprehensive insights into the pathophysiology of preeclampsia. J Pharm Biomed Anal. 2025;256:116701. doi:10.1016/j.jpba.2025.116701
160. Lin Q, Huang P, Kang Y, Lu Y, Shi G. Integrated multi-omics analysis reveals necroptosis-related biomarker BIRC3 for early diagnosis and therapeutic targeting in preeclampsia. PLoS One. 2025;20(12):e0337546. doi:10.1371/journal.pone.0337546
161. Marić I, Contrepois K, Moufarrej MN, et al. Early prediction and longitudinal modeling of preeclampsia from multiomics. Patterns N Y N. 2022;3(12):100655. doi:10.1016/j.patter.2022.100655
162. Zhao Q, Li J, Diao Z, et al. Early prediction of preeclampsia from clinical, multi-omics and laboratory data using random forest model. BMC Pregnancy Childbirth. 2025;25(1):531. doi:10.1186/s12884-025-07582-4
163. Youssef L, Testa L, Crovetto F, Crispi F. 10 Role of high dimensional technology in preeclampsia (omics in preeclampsia). Best Pract Res Clin Obstet Gynaecol. 2024;92:102427. doi:10.1016/j.bpobgyn.2023.102427
164. Rosenberg EA, Seely EW. Update on preeclampsia and hypertensive disorders of pregnancy. Endocrinol Metab Clin North Am. 2024;53(3):377–389. doi:10.1016/j.ecl.2024.05.012
165. Munjas J, Sopić M, Stefanović A, et al. Non-Coding RNAs in preeclampsia-molecular mechanisms and diagnostic potential. Int J Mol Sci. 2021;22(19):10652. doi:10.3390/ijms221910652
166. Lv Q, Wang Y, Tian W, et al. Exosomal miR-146a-5p derived from human umbilical cord mesenchymal stem cells can alleviate antiphospholipid antibody-induced trophoblast injury and placental dysfunction by regulating the TRAF6/NF-κB axis. J Nanobiotechnology. 2023;21(1):419. doi:10.1186/s12951-023-02179-5
167. Jiang Y, Luo T, Xia Q, Tian J, Yang J. microRNA-140-5p from human umbilical cord mesenchymal stem cells-released exosomes suppresses preeclampsia development. Funct Integr Genomics. 2022;22(5):813–824. doi:10.1007/s10142-022-00848-6
168. Chen Y, Ding H, Wei M, et al. MSC-Secreted exosomal H19 promotes trophoblast cell invasion and migration by downregulating let-7b and upregulating FOXO1. Mol Ther Nucleic Acids. 2020;19:1237–1249. doi:10.1016/j.omtn.2019.11.031
169. Shi H, Yang Z, Cui J, Tao H, Ma R, Zhao Y. Mesenchymal stem cell-derived exosomes: a promising alternative in the therapy of preeclampsia. Stem Cell Res Ther. 2024;15(1):30. doi:10.1186/s13287-024-03652-0
170. Göhner C, Plösch T, Faas MM. Immune-modulatory effects of syncytiotrophoblast extracellular vesicles in pregnancy and preeclampsia. Placenta. 2017;60(Suppl 1):S41–S51. doi:10.1016/j.placenta.2017.06.004
171. Li L, Yang H, Chen P, et al. Trophoblast-targeted nanomedicine modulates placental sFLT1 for preeclampsia treatment. Front Bioeng Biotechnol. 2020:8. doi:10.3389/fbioe.2020.00064
172. Hu L, Ma J, Cao M, et al. Exosomal mRNA and lncRNA profiles in cord blood of preeclampsia patients. J Matern Fetal Neonatal Med Off J Eur Assoc Perinat Med Fed Asia Ocean Perinat Soc Int Soc Perinat Obstet. 2022;35(25):8199–8209. doi:10.1080/14767058.2021.1966413
173. Zhao W, Li K, Li L, et al. Mesenchymal stem cell-derived exosomes as drug delivery vehicles in disease therapy. Int J Mol Sci. 2024;25(14):7715. doi:10.3390/ijms25147715
174. Zou Z, Li H, Xu G, Hu Y, Zhang W, Tian K. Current knowledge and future perspectives of exosomes as nanocarriers in diagnosis and treatment of diseases. Int J Nanomed. 2023;18:4751–4778. doi:10.2147/IJN.S417422
175. Sen S, Xavier J, Kumar N, Ahmad MZ, Ranjan OP. Exosomes as natural nanocarrier-based drug delivery system: recent insights and future perspectives. 3 Biotech. 2023;13(3):101. doi:10.1007/s13205-023-03521-2
176. Li T, Li X, Han G, et al. The therapeutic potential and clinical significance of exosomes as carriers of drug delivery system. Pharmaceutics. 2022;15(1):21. doi:10.3390/pharmaceutics15010021
177. Buca D, Bologna G, D’Amico A, et al. Extracellular vesicles in feto-maternal crosstalk and pregnancy disorders. Int J Mol Sci. 2020;21(6):2120. doi:10.3390/ijms21062120
178. Chen Y, Ou Z, Pang M, et al. Extracellular vesicles derived from Akkermansia muciniphila promote placentation and mitigate preeclampsia in a mouse model. J Extracell Vesicles. 2023;12(5):e12328. doi:10.1002/jev2.12328
179. Chen Y, Jin J, Chen X, Xu J, An L, Ruan H. Exosomal microRNA-342-5p from human umbilical cord mesenchymal stem cells inhibits preeclampsia in rats. Funct Integr Genomics. 2023;23(1):27. doi:10.1007/s10142-022-00931-y
180. Liu H, Wang F, Zhang Y, Xing Y, Wang Q. Exosomal microRNA-139-5p from mesenchymal stem cells accelerates trophoblast cell invasion and migration by motivation of the ERK/MMP-2 pathway via downregulation of protein tyrosine phosphatase. J Obstet Gynaecol Res. 2020;46(12):2561–2572. doi:10.1111/jog.14495
181. Liu A, Yang G, Liu Y, Liu T. Research progress in membrane fusion-based hybrid exosomes for drug delivery systems. Front Bioeng Biotechnol. 2022;10:939441. doi:10.3389/fbioe.2022.939441
182. Chan MH, Chang ZX, Huang CYF, Lee LJ, Liu RS, Hsiao M. Integrated therapy platform of exosomal system: hybrid inorganic/organic nanoparticles with exosomes for cancer treatment. Nanoscale Horiz. 2022;7(4):352–367. doi:10.1039/d1nh00637a
183. Huang W, Yu Y, Yang C, et al. Aptamer decorated magnetic graphene oxide nanoparticles for effective capture of exosomes. Chem Eng J. 2022;431:133849. doi:10.1016/j.cej.2021.133849
184. Reiner AT, Witwer KW, Van Balkom BWM, et al. Concise review: developing best-practice models for the therapeutic use of extracellular vesicles. Stem Cells Transl Med. 2017;6(8):1730–1739. doi:10.1002/sctm.17-0055
185. Quintero D, Perucca Orfei C, Kaplan LD, de Girolamo L, Best TM, Kouroupis D. The roles and therapeutic potentialof mesenchymal stem/stromal cells and their extracellular vesicles in tendinopathies. Front Bioeng Biotechnol. 2023;11:1040762. doi:10.3389/fbioe.2023.1040762
186. Popova AK, Vashukova ES, Illarionov RA, et al. Extracellular vesicles as biomarkers of pregnancy complications. Int J Mol Sci. 2024;25(22):11944. doi:10.3390/ijms252211944
187. Narumi R, Suzuki H, Ogoyama M, et al. Determination of the number of circulating small extracellular vesicles in pregnancy using the novel marker CD9. Int J Mol Sci. 2025;26(22):10906. doi:10.3390/ijms262210906
188. Huang Y, Zhang X, Xue L, He C. Nano-Enabled therapeutics: novel strategies for preeclampsia treatment. Int J Nanomed. 2026;21:578148. doi:10.2147/IJN.S578148
189. Wang L, Mu Q, Zhang W, et al. Placental targeted drug delivery: a review of recent progress. Nanoscale. 2025;17(14):8316–8335. doi:10.1039/d4nr05338a
190. Li L, Wang F, Zhu D, Hu S, Cheng K, Li Z. Engineering exosomes and exosome-like nanovesicles for improving tissue targeting and retention. Fundam Res. 2025;5(2):851–867. doi:10.1016/j.fmre.2024.03.025
191. Wang X, Munson MJ, Friis K, et al. Hybrid extracellular vesicles for efficient loading and functional delivery of mRNA. J Extracell Vesicles. 2025;14(12):e70201. doi:10.1002/jev2.70201
192. Abdel-Bar HM, Tandiono S, Liam-Or R, et al. Optimizing exosome lipid hybrid nanoparticles for enhanced siRNA delivery and improved therapeutic anticancer efficacy in vivo. ACS Nano. 2025;19(50):42658–42674. doi:10.1021/acsnano.5c16991
193. Evers MJW, Van de Wakker SI, De Groot EM, et al. Functional siRNA delivery by extracellular vesicle–liposome hybrid nanoparticles. Adv Healthc Mater. 2022;11(5):2101202. doi:10.1002/adhm.202101202
194. Lin Y, Wu J, Gu W, et al. Exosome–liposome hybrid nanoparticles deliver CRISPR/Cas9 system in MSCs. Adv Sci. 2018;5(4):1700611. doi:10.1002/advs.201700611
195. Berggreen AH, Petersen JL, Lin L, Benabdellah K, Luo Y. CRISPR delivery with extracellular vesicles: promises and challenges. J Extracell Biol. 2023;2(9):e111. doi:10.1002/jex2.111
196. Jang HJ, Shim KS, Lee J, et al. Engineering of cell derived-nanovesicle as an alternative to exosome therapy. Tissue Eng Regen Med. 2024;21(1):1–19. doi:10.1007/s13770-023-00610-4
197. Zhu L, Zhong Y, Wu S, et al. Cell membrane camouflaged biomimetic nanoparticles: focusing on tumor theranostics. Mater Today Bio. 2022;14:100228. doi:10.1016/j.mtbio.2022.100228
198. Aafreen S, Feng J, Wang W, Liu G. Theranostic extracellular vesicles: a concise review of current imaging technologies and labeling strategies. Extracell Vesicles Circ Nucleic Acids. 2023;4(1):107–132. doi:10.20517/evcna.2023.01
199. Zheng S, Georgiou HM, Kokkinos MI, Mirabito Colafella KM, Brennecke SP, Kalionis B. Therapeutic potential of extracellular vesicles for treating human pregnancy disorders. Extracell Vesicles Circ Nucleic Acids. 2025;6(2):287–309. doi:10.20517/evcna.2025.07
200. Amiri A, Bagherifar R, Ansari Dezfouli E, Kiaie SH, Jafari R, Ramezani R. Exosomes as bio-inspired nanocarriers for RNA delivery: preparation and applications. J Transl Med. 2022;20(1):125. doi:10.1186/s12967-022-03325-7
201. Poinsot V, Pizzinat N, Ong-Meang V. Engineered and mimicked extracellular nanovesicles for therapeutic delivery. Nanomater Basel Switz. 2024;14(7):639. doi:10.3390/nano14070639
202. Chen H, Yao H, Chi J, et al. Engineered exosomes as drug and RNA co-delivery system: new hope for enhanced therapeutics? Front Bioeng Biotechnol. 2023;11:1254356. doi:10.3389/fbioe.2023.1254356
203. Zheng L, Hu B, Zhao D, et al. Recent progresses of exosome–liposome fusions in drug delivery. Chin Chem Lett. 2024;35(2):108647. doi:10.1016/j.cclet.2023.108647
204. Wu S, Yun J, Tang W, et al. Therapeutic m6A eraser ALKBH5 mRNA-Loaded exosome-liposome hybrid nanoparticles inhibit progression of colorectal cancer in preclinical tumor models. ACS Nano. 2023;17(12):11838–11854. doi:10.1021/acsnano.3c03050
205. Gharehchelou B, Mehrarya M, Sefidbakht Y, et al. Mesenchymal stem cell-derived exosome and liposome hybrids as transfection nanocarriers of Cas9-GFP plasmid to HEK293T cells. PLoS One. 2025;20(1):e0315168. doi:10.1371/journal.pone.0315168
206. Feng Y, Lau S, Chen Q, et al. Normotensive placental extracellular vesicles provide long-term protection against hypertension and cardiovascular disease. Am J Obstet Gynecol. 2024;231(3):350.e1–350.e24. doi:10.1016/j.ajog.2023.12.030
207. Afzal A, Khan M, Gul Z, et al. Extracellular vesicles: the next frontier in pregnancy research. Reprod Sci. 2024;31(5):1204–1214. doi:10.1007/s43032-023-01434-2
208. Chaemsaithong P, Luewan S, Taweevisit M, et al. Placenta-Derived extracellular vesicles in pregnancy complications and prospects on a liquid biopsy for hemoglobin Bart’s disease. Int J Mol Sci. 2023;24(6):5658. doi:10.3390/ijms24065658
209. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7(1):1535750. doi:10.1080/20013078.2018.1535750
210. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
211. Ali M, Ahmed M, Memon M, et al. Preeclampsia: a comprehensive review. Clin Chim Acta Int J Clin Chem. 2024;563:119922. doi:10.1016/j.cca.2024.119922
212. Fan X, Zhang Y, Liu W, et al. A comprehensive review of engineered exosomes from the preparation strategy to therapeutic applications. Biomater Sci. 2024;12(14):3500–3521. doi:10.1039/d4bm00558a
213. Zhang P, Chen K, Liu W, et al. Engineered exosomes: a promising design platform for overcoming cancer therapy resistance. Front Cell Dev Biol. 2025:13. doi:10.3389/fcell.2025.1608480
214. Park SJ, Jung HI. Critical challenges and future direction in extracellular vesicle research and commercialization. BioChip J. 2025;19(3):411–423. doi:10.1007/s13206-025-00232-z
215. O’Gorman N, Wright D, Syngelaki A, et al. Competing risks model in screening for preeclampsia by maternal factors and biomarkers at 11-13 weeks gestation. Am J Obstet Gynecol. 2016;214(1):103.e1–103.e12. doi:10.1016/j.ajog.2015.08.034
216. Dave KM, Pinky PP, Manickam S. Molecular engineering of extracellular vesicles for drug delivery: strategies, challenges, and perspectives. J Control Release off J Control Release Soc. 2025;386:114068. doi:10.1016/j.jconrel.2025.114068
217. Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJA. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–345. doi:10.1038/nbt.1807
218. Jang SC, Kim OY, Yoon CM, et al. Bioinspired exosome-mimetic nanovesicles for targeted delivery of chemotherapeutics to malignant tumors. ACS Nano. 2013;7(9):7698–7710. doi:10.1021/nn402232g
219. Sato YT, Umezaki K, Sawada S, et al. Engineering hybrid exosomes by membrane fusion with liposomes. Sci Rep. 2016;6:21933. doi:10.1038/srep21933
220. Dobrovolskaia MA, Shurin M, Shvedova AA. Current understanding of interactions between nanoparticles and the immune system. Toxicol Appl Pharmacol. 2016;299:78–89. doi:10.1016/j.taap.2015.12.022
221. Liu J, Liu Z, Pang Y, Zhou H. The interaction between nanoparticles and immune system: application in the treatment of inflammatory diseases. J Nanobiotechnology. 2022;20(1):127. doi:10.1186/s12951-022-01343-7
222. Szebeni J. Complement activation-related pseudoallergy: a stress reaction in blood triggered by nanomedicines and biologicals. Mol Immunol. 2014;61(2):163–173. doi:10.1016/j.molimm.2014.06.038
223. Yang Q, Lai SK. Anti-PEG immunity: emergence, characteristics, and unaddressed questions. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2015;7(5):655–677. doi:10.1002/wnan.1339
224. Mistry HD, Klossner R, Kallol S, et al. Effects of aldosterone on the human placenta: insights from placental perfusion studies. Placenta. 2022;123:32–40. doi:10.1016/j.placenta.2022.03.129
225. Gimona M, Brizzi MF, Choo ABH, et al. Critical considerations for the development of potency tests for therapeutic applications of mesenchymal stromal cell-derived small extracellular vesicles. Cytotherapy. 2021;23(5):373–380. doi:10.1016/j.jcyt.2021.01.001
226. Mendt M, Kamerkar S, Sugimoto H, et al. Generation and testing of clinical-grade exosomes for pancreatic cancer. JCI Insight. 2018;3(8):e99263. doi:10.1172/jci.insight.99263
227. Bisaccia P, Magarotto F, D’Agostino S, et al. Extracellular vesicles from mesenchymal umbilical cord cells exert protection against oxidative stress and fibrosis in a rat model of bronchopulmonary dysplasia. Stem Cells Transl Med. 2024;13(1):43–59. doi:10.1093/stcltm/szad070
228. Esfandyari S, Elkafas H, Chugh RM, Park HS, Navarro A, Al-Hendy A. Exosomes as biomarkers for female reproductive diseases diagnosis and therapy. Int J Mol Sci. 2021;22(4):2165. doi:10.3390/ijms22042165
229. Nazri HM, Greaves E, Quenby S, Dragovic R, Tapmeier TT, Becker CM. The role of small extracellular vesicle-miRNAs in endometriosis. Hum Reprod Oxf Eng. 2023;38(12):2296–2311. doi:10.1093/humrep/dead216
230. Ma Y, Hou B, Zong J, Liu S. Potential molecular mechanisms and clinical implications of piRNAs in preeclampsia: a review. Reprod Biol Endocrinol RBE. 2024;22(1):73. doi:10.1186/s12958-024-01247-1
231. Zhou S, Li J, Yang W, et al. Noninvasive preeclampsia prediction using plasma cell-free RNA signatures. Am J Obstet Gynecol. 2023;229(5):553.e1–553.e16. doi:10.1016/j.ajog.2023.05.015
232. Ogoyama M, Takahashi H, Suzuki H, Ohkuchi A, Fujiwara H, Takizawa T. Non-Coding RNAs and prediction of preeclampsia in the first trimester of pregnancy. Cells. 2022;11(15):2428. doi:10.3390/cells11152428
233. Anzalone AV, Koblan LW, Liu DR. Genome editing with CRISPR-Cas nucleases, base editors, transposases and prime editors. Nat Biotechnol. 2020;38(7):824–844. doi:10.1038/s41587-020-0561-9
234. Mishra S, Rehan S, Barekzai AM, Sharma A, Raghav A. Emerging frontiers in genome editing: from CRISPR to next-generation technologies. Methods. 2026;S1046–2023(26)00131–3. doi:10.1016/j.ymeth.2026.05.011
235. Kelly CJ, Karthikesalingam A, Suleyman M, Corrado G, King D. Key challenges for delivering clinical impact with artificial intelligence. BMC Med. 2019;17(1):195. doi:10.1186/s12916-019-1426-2
236. Riley RD, Ensor J, Snell KIE, et al. Calculating the sample size required for developing a clinical prediction model. BMJ. 2020;368:m441. doi:10.1136/bmj.m441
237. Subramanian I, Verma S, Kumar S, Jere A, Anamika K. Multi-omics data integration, interpretation, and its application. Bioinforma Biol Insights. 2020;14:1177932219899051. doi:10.1177/1177932219899051
238. Singh A, Shannon CP, Gautier B, et al. DIABLO: an integrative approach for identifying key molecular drivers from multi-omics assays. Bioinformatics. 2019;35(17):3055–3062. doi:10.1093/bioinformatics/bty1054
239. Shao X, Yu L, Li C, et al. Extracellular vesicle-derived miRNA-mediated cell-cell communication inference for single-cell transcriptomic data with miRTalk. Genome Biol. 2025;26(1):95. doi:10.1186/s13059-025-03566-x
240. Athaya T, Li X, Hu H. A deep learning method to integrate extracelluar miRNA with mRNA for cancer studies. Bioinforma Oxf Engl. 2024;40(11):btae653. doi:10.1093/bioinformatics/btae653
241. Greenberg ZF, Graim KS, He M. Towards artificial intelligence-enabled extracellular vesicle precision drug delivery. Adv Drug Deliv Rev. 2023;199:114974. doi:10.1016/j.addr.2023.114974
242. Del R MC, Lu Y, Jalali M, et al. Evaluation of machine learning and deep learning models for the classification of a single extracellular vesicles spectral library. Sens Diagn. 2025;4(10):869–883. doi:10.1039/D5SD00091B
243. Thakur A, Rai D. Global requirements for manufacturing and validation of clinical grade extracellular vesicles. J Liq Biopsy. 2024;6:100278. doi:10.1016/j.jlb.2024.100278
244. Si C, Gao J, Ma X. Engineered exosomes in emerging cell-free therapy. Front Oncol. 2024;14:1382398. doi:10.3389/fonc.2024.1382398
245. Stawarska A, Bamburowicz-Klimkowska M, Runden-Pran E, et al. Extracellular vesicles as next-generation diagnostics and advanced therapy medicinal products. Int J Mol Sci. 2024;25(12):6533. doi:10.3390/ijms25126533
246. Li B, Huang Y, Wu B, et al. Screening and Clinical Verification of Extracellular Vesicle Biomarkers. In: Wang Q, Zheng L, editors. Extracellular Vesicles: From Bench to Bedside. Springer Nature; 2024:265–274. doi:10.1007/978-981-99-8365-0_15
247. Zou Y, Zhou Y, Li G, Dong Y, Hu S. Clinical applications of extracellular vesicles: recent advances and emerging trends. Front Bioeng Biotechnol. 2025;13. doi:10.3389/fbioe.2025.1671963
248. Van Dorpe S, Tummers P, Denys H, Hendrix A. Towards the clinical implementation of extracellular vesicle-based biomarker assays for cancer. Clin Chem. 2024;70(1):165–178. doi:10.1093/clinchem/hvad189
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Overcoming Obstacles in Liquid Biopsy Developments for Prostate Cancer
Jang A, Rauterkus GP, Vaishampayan UN, Barata PC
OncoTargets and Therapy 2022, 15:897-912
Published Date: 26 August 2022
An Investigation of Exosome Concentration and Exosomal microRNA (miR-155 and miR-222) Expression in Pregnant Women with Gestational Hypertension and Preeclampsia
Ntsethe A, Mackraj I
International Journal of Women's Health 2022, 14:1681-1689
Published Date: 7 December 2022
Tailored Extracellular Vesicles from Dental Stem Cells: Advances in Specific Modifications for Enhanced Therapeutic Applications
Liu X, Li Z, Fu J, Wang R, He J, Yao J, Ye Q, He Y
International Journal of Nanomedicine 2025, 20:8327-8341
Published Date: 26 June 2025
Regenerative Potential of Exosomes from 3D Spheroid-Cultured Stem Cells Compared with 2D Culture in Dentin–Pulp Regeneration: A Systematic Review
Jos Erry HW, Margono A, Bagio DA, Suniarti DF, Kusuma I, Budi HS, Mauludin R, Dalimunthe RS, Tambun DV, Amir LR
Clinical, Cosmetic and Investigational Dentistry 2026, 18:614885
Published Date: 2 June 2026