")
Back to Journals » ImmunoTargets and Therapy » Volume 12
Exploration of Immune Targets for Type 1 Diabetes and Latent Autoimmune Disease Immunotherapy
Authors Siddiqui K , Nawaz SS
Received 4 June 2023
Accepted for publication 9 September 2023
Published 29 September 2023 Volume 2023:12 Pages 91—103
DOI https://doi.org/10.2147/ITT.S417917
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Shurin
Khalid Siddiqui,* Shaik Sarfaraz Nawaz*
Strategic Center for Diabetes Research, College of Medicine, King Saud University, Riyadh, Saudi Arabia
*These authors contributed equally to this work
Correspondence: Khalid Siddiqui, Strategic Center for Diabetes Research, College of Medicine, King Saud University, P.O. Box 245, Riyadh, 11411, Saudi Arabia, Tel +966 114724179 Ext.3106 ; +966 530202763, Email [email protected]
Abstract: Type 1 diabetes (T1D) is an autoimmune disease that destroys pancreatic beta cells, which produce insulin in the islets of Langerhans. The risk of developing T1D is influenced by environmental factors, genetics, and autoantibodies. Latent autoimmune diabetes in adults (LADA) is a type of T1D that is genetically and phenotypically distinct from classic T1D. This review summarizes the accumulated information on the risk factors for T1D and LADA, and immunotherapy trials that offer insights into potential future combined therapeutic interventions for both T1D and LADA to slow the rate of islet cell loss and preserve beta cell function. Future research should also focus on improving intervention doses, conducting more thorough examinations of intervention responders, and/or combining minimally effective single-target immunotherapies to slow the rate of islet cell loss and preserve beta cell function.
Keywords: type 1 diabetes, LADA, immunotherapy, islet cells
Overview of Diabetes
Diabetes is a metabolic disease defined by hyperglycemia caused by deficiencies in insulin production, action, or both.1 Diabetes long-term complications include retinopathy, which can lead to vision loss; nephropathy, which can lead to renal failure; peripheral neuropathy, which can lead to foot ulcers, amputations, and Charcot’s joints; and autonomic neuropathy, which can lead to gastrointestinal, genitourinary, and cardiovascular symptoms, as well as sexual dysfunction. Atherosclerotic cardiovascular, peripheral arterial and cerebral vascular disease are more common in diabetic patients. In persons with diabetes, hypertension and anomalies in lipoprotein metabolism are frequently detected.1,2
Diabetes is classified into four types or categories by the American Diabetes Association: type 1 diabetes, type 2 diabetes, gestational diabetes mellitus, and diabetes caused or associated with specific conditions (such as neonatal diabetes and maturity-onset diabetes of the young), exocrine pancreas diseases (such as cystic fibrosis and pancreatitis), and drug- or chemical-induced diabetes (such as with glucocorticoid use, in the treatment of HIV/AIDS, or after organ transplantation).2
Immune system modulation is the goal of immunotherapy, a method that has proved effective in treating autoimmune diseases. However, immunotherapy treatment for T1D and LADA has had a low success rate. In this review, we summarize the accumulated information on the risk factors for type 1 diabetes and latent autoimmune diabetes in adults and immunotherapy trials that offer insights into potential future combined therapeutic interventions for both T1D and LADA to slow the rate of islet cell loss and preserve beta cell function. We highlight the most important immunotherapeutic techniques that have been evaluated thus far, with an emphasis on distinguishing traits.
Type 1 Diabetes
Type 1 diabetes (T1D) affects more than 1.2 million children and adolescents worldwide.3 T1D is an autoimmune disease characterized by the destruction of immune-mediated pancreatic beta cells in the islets of Langerhans.4 Although T1D can occur at any age, it is most common in children and young adults. For those with T1D, daily insulin injections are necessary to maintain a healthy blood glucose level and are required for people with T1D to survive.5,6 Insulin therapy does not cure T1D or completely avoid complications, the use of insulin has revolutionized T1D management.7–9 Apart from the acute complications of hypoglycemia (abnormally low blood glucose) and diabetic ketoacidosis (DKA), poor metabolic control results in poor growth and the onset of vascular complications at an early age.6
Risk Factors for T1D
The risk of developing preclinical or clinical T1D is influenced by environmental exposures and genetic predisposition.9 It is suggested that gluten diet, cow milk, vitamin D intake, gut microbiota, viral infections (enteroviruses, Coxsackieviruses), drugs (streptozotocin, pentamidine, and antibiotics), and epigenetic modifications all these factors play a role in T1D development.10
Risk Gene for T1D
According to genome-wide association studies (GWASs), T1D is a polygenic disease with approximately 50% genetic risk attributable to the HLA-DR-DQ haplotypes of the major histocompatibility complex (MHC) region of human leukocyte antigen (HLA) class II (HLA), which are responsible for the presentation by B lymphocyte cells, dendritic cells, and macrophages to cluster of differentiation CD4+ T lymphocyte cells.11
Most non-HLA-associated genes identified by GWASs are involved in immune regulation and function (insulin, protein tyrosine phosphatase non-receptor type 22 (PTPN22), interleukin 2 receptor subunit alpha (IL2RA), SH2B adaptor protein 3 (S12H2B3), protein tyrosine phosphatase non-receptor type 2 (PTPN2), cytotoxic T-lymphocyte associated protein-4 (CLTA4), interleukin 18 receptor accessory protein (IL18RAP), c-c motif chemokine receptor 5 (CCR5), interferon induced with helicase C domain 1 (IFIH1), a cluster of differentiation 226 (CD226), protein kinase c theta (PRKCQ), interleukin-2 (IL-2), BTB domain and CNC homology 2 (BACH2), ubiquitin associated and SH3 domain containing A (UBASH3A), a regulator of G protein signaling 1 (RGS1), T-cell activation Rho GTPase activation protein (TAGAP); insulin production and metabolism (Erb-B2 receptor tyrosine kinase 3 (ERBB3); protection from beta cell apoptosis (tumor necrosis factor alpha-induced protein 3 (TNFAIP3).12
The UK Biobank Affymetrix axiom array data was recently used to validate a genetic risk score (GRS) that used 67 single nucleotide polymorphisms (SNPs) (14 HLA DQA1-DQB1 SNPs, 21 other HLA SNPs, and 32 non-HLA SNPs) for newborn screening and future T1D incidence. T1D GRS considerably enhanced the discrimination of T1D patients from T2D and control participants.13 These data have made it possible to create GRS that could be used to identify those who are at high risk.14 Although identifying GRS can determine their lifelong risk for developing a disease, it cannot forecast those who have the preclinical disease and are experiencing the destruction of their beta cells.
Immune Pathogenesis
Islet autoimmunity in T1D manifests as persistent hyperglycemia and low c-peptide due to permanent loss of more than 70% of beta cell mass months to decades before the clinical disease.15 Blood glucose, HbA1c, and c-peptide were the only way to diagnose T1D before the discovery of the underlying immunological pathogenesis and biomarkers to identify those who have the disease.
T1D is characterized by the presence of specific autoantibodies (AAbs) for beta cell antigens as well as insulitis.10 Several other additional autoantigens have also shown their presence and are recognized in human T1D as shown in Table 1.
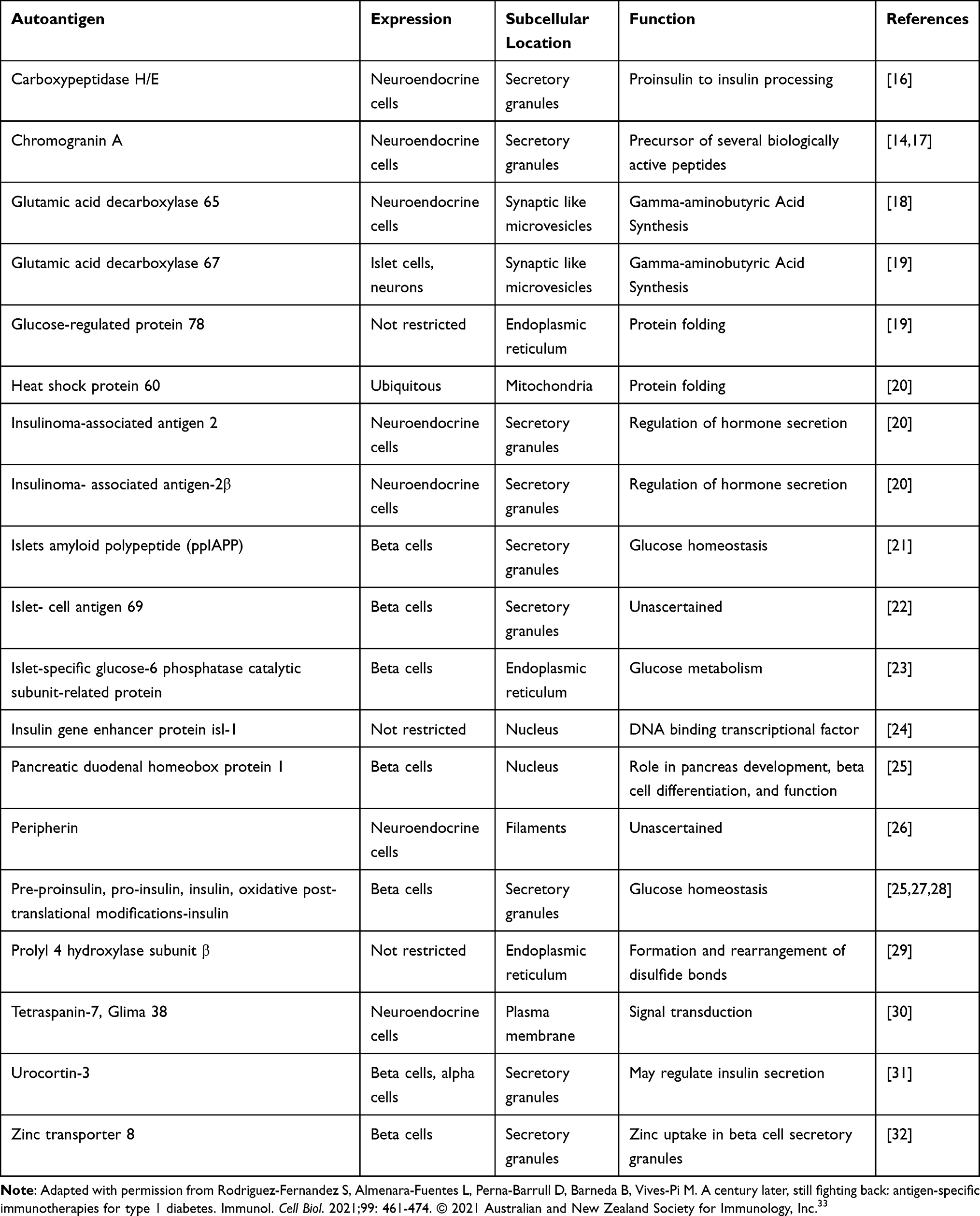 |
Table 1 Islet Autoantibodies to Pancreatic Insulin-Producing Cells Secreted in T1D |
Prospective, long-term investigations of those at risk of developing T1D have revealed that the disease proceeds progressively through discrete recognizable phases before the onset of symptoms at varying but predictable rates. Stage 1 is characterized as pre-symptomatic beta cell autoimmunity as shown by the presence of two or more islet autoantibodies with normoglycemia, stage 2 as pre-symptomatic beta cell autoimmune with dysglycemia, and stage 3 as symptomatic disease onset. The adoption of this staging classification will give T1D a standardized taxonomy, aid in the creation of treatments and the planning of clinical trials to prevent symptomatic disease, advance precision medicine, and offer a structure for an optimized benefit/risk ratio that will influence regulatory approval, reimbursement, and the adoption of interventions in the early stages of T1D to prevent symptomatic disease.34 Islet-targeting autoantibodies that target glutamic acid decarboxylase 65, insulinoma-associated protein 2, insulin, and zinc transporter 8 are all proteins linked to secretory granules in beta cells and are all biomarkers of T1D-associated autoimmunity that are discovered months to years before symptoms appear.35 These markers can be used to identify and study the risk of developing T1D.
At 5, 10, and 15 years of follow-up in the Colorado Diabetes Autoimmunity Study in the Young (DAISY), the Finnish T1D prediction and prevention (DIPP), and the German BABYDIAB and BABYDIET studies, the progression rates of T1D following seroconversion were 43.5%, 69.7%, and 84.2%, respectively.36 Understanding the immune mechanisms of T1D has made it possible to identify individuals who are more likely to experience clinical disease and to create a novel treatment that aims to delay the onset and reverse the effects of the disease.14
In recent years, the T1D Biomarker Working Group and its accompanying Core for Assay Validation (www.t1dbiomarkers.org) have been dedicated to transferring promising candidate biomarkers from the discovery arena to confirmation and validation testing through a collaborative and coordinated approach. T cell-related biomarkers would significantly speed up disease progression monitoring and the evaluation of T1D intervention therapy.37 Due to the extremely low frequency of diabetogenic T cells in peripheral blood, the low avidity contacts between autoreactive T-cell receptors (TCRs) and the HLA peptide complex, and the significant disease heterogeneity, this research has proven to be challenging.14 However, recent advances in single-cell technologies are making it possible to characterise diabetogenic T cells in high-dimensional phenotypic, transcriptional, and epigenetic detail.38 This might eventually lead to the discovery of accurate and sensitive immunological biomarkers.
Research Resources such as the Network for Pancreatic Organ Donors with Diabetes and Juvenile Diabetes Research Foundation (nPOD; http://www.jdrfnpod.org/) provide an organized framework for obtaining tissues from subjects with T1D or at risk of T1D, allowing for direct study of islet-infiltrating T cells, islet autoantigen reacting T-cells. Human peripheral autoreactive T cells reacted to epitopes that showed posttranslational changes such as disulfide bonds in insulin, islet-derived T cells with hybrid insulin peptide, fusions of proinsulin c-peptide with islet amyloid polypeptide (IAPP).39 Islet-infiltrating T lymphocytes that specifically target proinsulin peptides. The B:9–23 specific T cells of the islets responded to proinsulin, highlighting the relevance of proinsulin-specific T cells in the islet microenvironment.40 CD4 T cells identify epitopes generated by the covalent cross-linking of proinsulin peptides to other peptides found in cell secretory granules. These hybrid insulin peptides (HIPs) are antigenic to CD4 T cells. T cells that target hybrid peptides explain how immunological tolerance is disrupted in T1D.41 Understanding the TCR repertoire of pathogenic T cells in T1D may allow for their isolation and use as surrogate biomarkers of drug efficacy in individuals receiving immune therapies.14
Latent Autoimmune Diabetes in Adults
The pathogenesis and clinical manifestations of LADA are extremely diverse. Beyond the distinction between classic T1D and LADA, the extensive heterogeneity of autoimmune diabetes is evident. It differs from classic T1D/ T2D in terms of genetic background, autoimmune response, rate of decrease in pancreatic islet function, and clinical metabolic characteristics. It is characterized by genetic, phenotypic, and humoral variability, encompassing varying degrees of insulin resistance and autoimmunity.42,43 Although these patients did not require insulin at the time of diagnosis, they are believed to have a slowly progressing form of autoimmune diabetes with serum immunological markers of T1D.
The primary current diagnostic criteria for LADA include adult-onset diabetes (>30 years at diagnosis), the presence of diabetes-associated autoantibodies, and the lack of insulin demand for at least 6 months following diagnosis.43–45
The genetic distribution and phenotypic traits of the various LADA groups were related to the GADA level. From the lowest GADA quartile to the highest, a significant trend toward reduced insulin secretion and metabolic trait values was observed. Therefore, LADA patients with high GADA concentrations resembled but were not identical to, patients with T1D, while those with low GADA concentrations resembled patients with T2D. Although the risk genotypes for HLA-DQB1 and protein tyrosine phosphatase non-receptor 22 (PTPN22) were elevated in LADA, they were substantially less prevalent than in T1D.46
LADA accounts for 2–12% of all diabetes patients, with wide variations based on ethnicity, the autoantibody used for screening (most frequently an autoantibody against glutamic acid decarboxylase [GADA]), and the method of diagnosis.44,47 The human and an established rat model (LEW.1AR1/Ztm-IDDM) of LADA pancreas demonstrate variations in immune cell infiltration, as well as a shift in the ratio of macrophages to CD8 T cells in the islet infiltrate, owing to an increase in Interleukin 1 beta (IL-1β) and a reduction in tumor necrosis factor-α. In LADA, Interleukin 10 (IL-10), proliferating cell nuclear antigen (PCNA), and insulin expression rose, but caspase 3 gene expression decreased, the underlying pathophysiology in human and rat LADA pancreases was identical. LADA is a milder type of autoimmune diabetes in individuals of a late age.47
Risk Factor for Latent Autoimmune Diabetes in Adults
In the first genome-wide association study of LADA, GWAS signals indicate that LADA is a late-onset form of T1D, albeit with genetically attenuated potency of key T1D-associated HLA haplotypes and also with a genetic component similar to T2D. Functional studies look at how the glycolytic regulator PFKFB3 is placed at the nexus of autoimmune and metabolic diabetes.48 Independent effects of MHC class I reported in T1D were not observed in LADA, indicating that the association of MHC class I may be a genetic discriminator between LADA and childhood-onset T1D.49 Recent research has confirmed the genetic relationship between LADA and both T1D and T2D, as well as the subtype-specific signatures in the HLA and a novel LADA-specific locus called PFKFB3. GRS, which includes T1D-risk variations, is a useful tool for differentiating diabetes subtypes and identifying patients who are insulin-dependent in fast-developing states. Although genetic evidence supports the existence of LADA, more research is needed to fully determine its role in the diabetes spectrum.50
The Nord-Trondelag Health Study (HUNT) genotyped 60 SNPs known to be associated with T1D or T2D, including 14 tag SNPs for HLA haplotyping in 120 patients with T1D, 126 LADA, and 1090 T2D. Genetic heterogeneity of LADA is linked to varying degrees of autoimmune activity and is distinguishable from both T1D and T2D. Most strongly associated HLA haplotypes for T1D were significantly associated with patients with LADA, but primarily with LADA with high anti-GAD levels. There were no connections between LADA and non-HLA T1D loci. Tetraspanin-8 (TSPAN8) and the fat mass and obesity-associated (FTO) were two T2D-related genes that were generally associated with LADA, but primarily in patients with low anti-GAD LADA levels.51 The researchers investigated the relationship between altering beta-cell function in patients with latent autoimmune diabetes in adulthood and GAD autoantibody (GADA) titers. Initial GADA titers in LADA patients identified subjects with varying degrees of persistent autoimmunity and disease progression. Low GADA titer LADA patients shared T2D patients’ metabolic characteristics and decreased beta-cell function.52 In two large population-based studies (the Swedish case-control study and the Norwegian HUNT Study), obesity and overweight are linked to an increased risk of LADA, especially when combined with a family history of diabetes.53 BMI was higher in LADA than in T2D but lower in T1D. GAD antibody titer in LADA correlated negatively with BMI and c-peptide secretion. Beta cell function from the intravenous glucose tolerance test (IVGTT) in LADA was 228% higher than in T2D but 35% lower than in T1D. LADA had insulin sensitivity comparable to T2D but 41% higher than T1D.54 The systematic and meta-analysis study found polymorphisms of protein tyrosine phosphatase N22 (PTPN22), Insulin, transcriptional factor 7-like 2 (TCFL2), and variants of cytotoxic T-lymphocyte antigen-4 (CTLA-4) as risk factors for LADA.55,56
Furthermore, consumption of processed red meat was associated with an increased risk of LADA particularly in people with a family history of T1D or high-risk HLA genotypes (HLA-DQB1 and HLA-DRB1).57 The family history of T1D, as well as a family history of T2D, was linked to an elevated risk of LADA.58 Being overweight, physical inactivity, smoking, low birth weight, sweetened beverage consumption, and alcohol use are all associated with an increased risk of LADA.59–61 LADA may be avoided by making the same lifestyle changes as T2D, such as losing weight, increasing physical activity, and quitting smoking.
During the 7-year follow-up period, 119 (56.1%) of the 212 LADA patients required insulin. In the first year after diagnosis, high GADA titers, BMI≤ 25, ZnT8 and IA-2 positive, and sulfonylurea medication significantly enhance the development of insulin demand in LADA patients. The distinctive gut microbiota and related metabolites of LADA patients are linked to autoantibodies, glucose metabolism, islet function, and inflammatory factors, all of which may contribute to the disease’s etiology.62
Immunotherapy Approaches to Suppress Beta-Cell Autoimmunity
In recent years, various combination therapies that target the many pathways involved in beta cell destruction have been proposed. However, new promising combination therapies, such as those that combine immunomodulators with drugs that stimulate beta cell regeneration to restore normoglycemia, are being developed.63 In the pancreatic draining lymph node (pLN), a stem-like autoimmune progenitor gives rise to pLN autoimmune mediators. Using single-cell RNA sequencing and clonal analysis, researchers discovered that autoimmune CD8 T cells represent distinct T cell differentiation states and identified factors that drive the transition from a stem-like autoimmune progenitor to the autoimmune mediator in the pLN. Strategies that target the stem-like autoimmune progenitor pool have the potential to be novel and powerful immunotherapeutic interventions for T1D.64 Targeting the stem-like autoimmune progenitor pool may lead to the development of innovative and potent immunotherapeutic treatments for T1D. Moreover, researchers hypothesized that microneedle intradermal administration of human proinsulin peptide C19-A3 coupled to ultrasmall gold nanoparticles (GNPs) may increase antigen-specific immunotherapy by promoting tolerogenic dendritic cell immunomodulation.65 Liraglutide, a drug used to promote beta cell regeneration, and immunotherapy based on Phosphatidylserine (PS) in the membrane and encapsulating insulin A and B chains (PSAB-liposomes) shown to reduce hyperglycemia in an autoimmune T1D model with spontaneous onset.66 Recent data suggest that MBD2 (methyl-CpG-binding domain 2) could be a viable target for developing epigenetic-based T1D therapeutics in clinical settings. Mbd2 deficiency exacerbated the development of T1D in the NOD mice model. Th1 stimulation caused the Stat1 promoter to undergo DNA methylation and induction of MBD2 expression, which then bound to methylated CpG DNA within the Stat1 promoter, allowing MBD2 to maintain Th1 program homeostasis and prevent autoimmunity. By controlling the STAT1-IFN axis, MBD2 functions as a repressor to maintain the homeostasis of the Th1 program in T1D.67 Moreover, the deletion of Renalase (RNLS), a potential gene for T1D identified by a GWAS, made beta cells resistant to autoimmune destruction in a mouse model of T1D. In diabetic mice, oral pargyline protected transplanted beta cells, by delaying diabetes development. RNLS is a therapeutic target to prevent the loss of beta cells in T1D and a regulator of beta cell susceptibility.68 However, its precise role in LADA is unknown.
Circulating C-X-C motif chemokine receptor type 5-negative, programmed cell death protein 1-positive (CXCR5−PD-1hi) peripheral T helper (Tph) cells are linked to clinical T1D development. Tph cells are useful biomarkers of disease progression as well as a target for immunotherapy in T1D.69 A combination of six HLA-DRB1*0401-selective beta-cell peptides was administered intravenously to individuals with this genotype who had just developed T1D at dosages of 10, 100, and 500 g per month for 24 weeks. Treatment resulted in dose-dependent increases regulatory T cells (Treg) expression of the canonical transcription factor FOXP3 and altered expression of the Treg gene, as well as substantial modifications in islet-specific immune responses. Multiple-peptide immunotherapy shows promise as an approach to repair immune regulatory abnormalities key to the pathobiology of autoimmune diabetes in this first-in-human investigation.70,71
Immunotherapy Approaches from Intervention for Prevention of T1D
Despite over a century of insulin replacement treatment, there is still no cure for T1D etiology. Finding the “correct” therapy for the “right” patient at the “right” moment is still an unmet goal in T1D. For those with T1D, disease-modifying treatments continue to be an aspirational goal. Several therapies, including anti-inflammatory medications, and T or B cell-specific immunosuppressants, have been investigated thus far. In addition, a small number of clinical trial data have revealed minor benefits of immunotherapy in T1D summarized in Table 2.
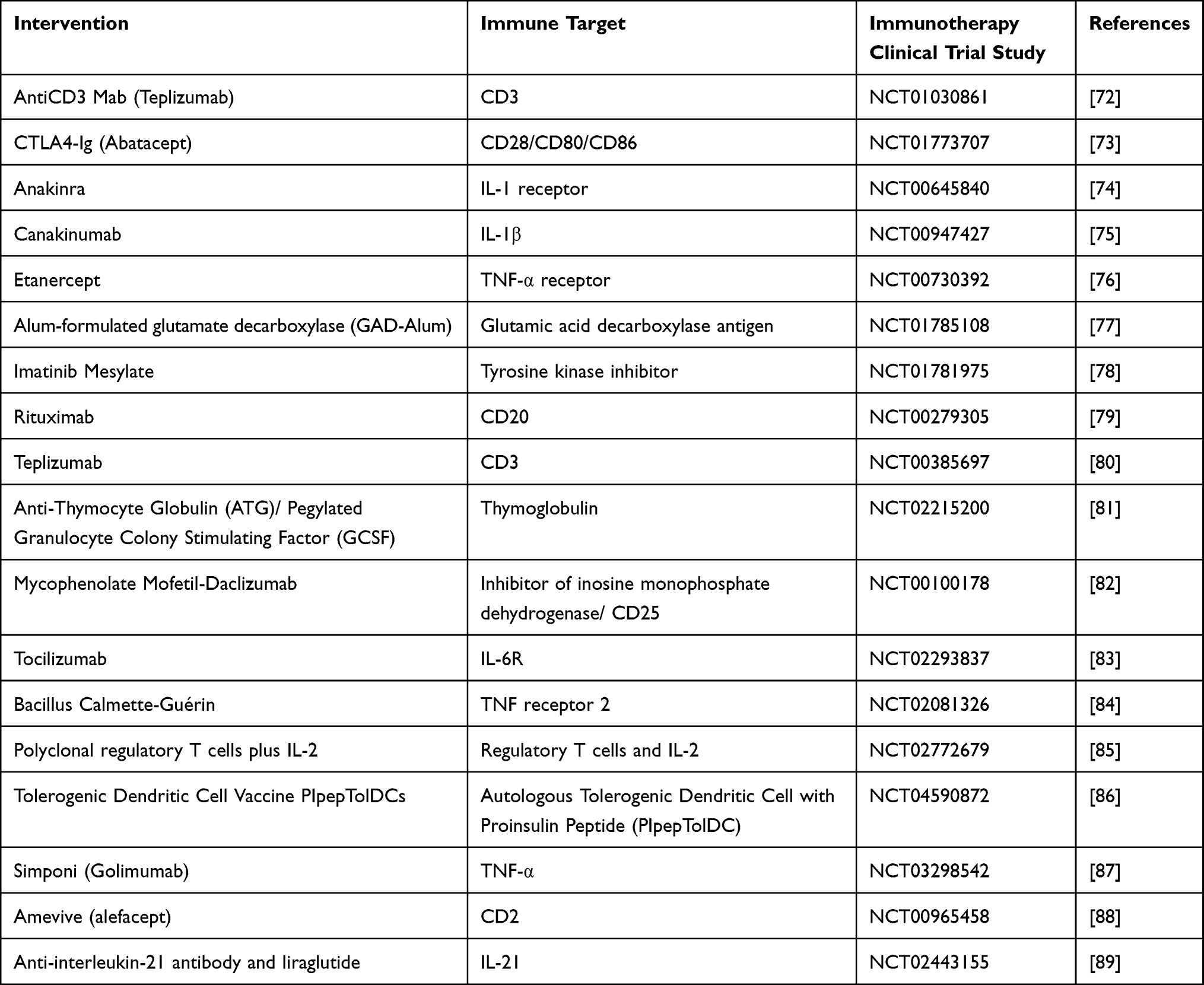 |
Table 2 Immune Interventions in T1D |
In the Phase 2 study, a single 14-day session of teplizumab (Fc receptor-nonbinding anti-CD3 monoclonal antibody) significantly reduced the development of clinical T1D in high-risk, non-diabetic relatives of diabetic patients with at least two autoantibodies and abnormal oral glucose tolerance test. The median delay in diabetes diagnosis was two years. The individuals most likely to respond were those lacking one T1D-associated MHC allele, HLA-DR3, but possessing HLA-DR4, as well as the absence of anti-ZnT8 antibodies (ClinicalTrials.gov Identifier: NCT01030861).72 Teplizumab is currently licensed in the United States for patients who are at high risk of developing T1D.90
Consequently, in multicenter, double-blind, randomised controlled trial T1D patients aged 6–45 years who have just received abatacept (10–1000 mg/kg per dose) intravenously on days 1, 14, and 28 and once a month for a total of 27 infusions for two years. Over two years, co-stimulation modulation with abatacept halted the decline in beta cell function. The positive impact shows that T-cell activation still occurs around the time of T1D clinical diagnosis (ClinicalTrials.gov Identifier: NCT00505375).73
Children who had been diagnosed with T1D received daily anakinra for 28 days and were observed for 6 months. Before and after anakinra therapy, blood was taken for microarray analysis. Anakinra-treated patients showed identical HbA1c and mixed-meal tolerance testing (MMTT) responses, but reduced insulin needs 1 and 4 months after diagnosis, and lower insulin-dose-adjusted HbA1c 1 month after diagnosis, compared to controls (ClinicalTrials.gov Identifier: NCT00645840).91 In clinical investigations, T1D patients aged 6–45 years were included in a randomized, placebo-controlled experiment. In newly diagnosed T1D, canakinumab (a human monoclonal anti-interleukin-1 antibody) 2 mg/kg (maximum 300 mg) subcutaneous injection monthly for 12 months was proved safe but ineffective as a single immunomodulatory medication (ClinicalTrials.gov, identifier: NCT00947427).75 In organ-specific autoimmune diseases, interleukin-1 inhibition may be more efficient when used in conjunction with therapies that target adaptive immunity. At a dose of 0.4 mg/kg (maximum dose of 25 mg/dose), twice a week, etanercept (TNF receptor inhibitor) is administered subcutaneously to 18 newly diagnosed T1D patients (11 men and 7 women, ages 7.8–18.2 years). Etanercept therapy reduced HbA1c and increased endogenous insulin production in young individuals with newly diagnosed T1D, indicating that beta cell activity was preserved (ClinicalTrials.gov identifier: NCT00730392).76
In a double-blind, placebo-controlled clinical trial, non-diabetic children between the ages of 4 and 17.9 who had autoantibodies to glutamate decarboxylase (GADA), insulin, or zinc-transporter 8 were randomized, stratified by 2 or more islet autoantibodies, to 2 injections of 20 g GAD-Alum or placebo, spaced 30 days apart. In young children with prediabetes, GAD-Alum as a subcutaneous prime and boost injection was safe, but it had little impact on the development of T1D (ClinicalTrials.gov identifier: NCT01122446).92,93 In recent-onset T1D, a double-blind placebo-controlled intervention with glutamic acid decarboxylase (GAD)-alum, vitamin D, and Ibuprofen was conducted (T1D). 64 T1D patients were randomized (age 10–17.99 years, fasting c-peptide 0.12 nmol/l, GADA-positive). A linear relationship was found between baseline c-peptide, HbA1c, and insulin/per kilogram/24 h and change in c-peptide AUC after 15 months. c-peptide was not preserved by ibuprofen, vitamin D, or GAD-alum. Baseline clinical and immunological variables, as well as vitamin D levels, all affected treatment success (Clinical Trial Registration Identifier: NCT01785108).94
In a multicenter phase 2 trial, randomly assigned, double-blind, placebo-controlled patients with newly diagnosed T1D aged between 18–45 years, positive for islet autoantibody, and with stimulated c-peptide of greater than 0.2 nmol L−1 on a mixed meal tolerance test (MMTT) were included. At a dose of 400 mg imatinib mesylate (4X100 mg of film-coated tablets per day) is administered. A 26-week treatment of imatinib retained beta cell function at 12 months (ClinicalTrials.gov, Identifier: NCT01781975). Determining the appropriate dosage and duration of treatment, safety, and efficacy of imatinib in children, the use of complementary medications in combination, and imatinib’s ability to delay or arrest the progression of diabetes in a population at risk. A double-blind experiment comprised 87 newly diagnosed T1D patients aged 8 to 40 rituximab treatment decreased CD19+ B cells, lowered glycated hemoglobin levels, and needed less insulin. The use of rituximab preserved beta-cell activity (ClinicalTrials.gov identifier: NCT00279305).95 The discovery that B lymphocytes contribute to the pathophysiology of T1D may offer up a new avenue in the treatment of this ailment. In this two-year trial, participants are given one of three teplizumab infusion regimens (14-day full dosage, 14-day reduced dose, or 6-day full dose). Teplizumab immunotherapy inhibits deterioration in beta-cell function and improves glycemic control with reduced insulin dosages (ClinicalTrials.gov, Identifier NCT00385697).15
The T1D TrialNet Study Group showed that low-dose anti-thymocyte globulin (ATG) (2.5 mg/kg) retained beta-cell function and lowered HbA1c for 1 year in new-onset T1D in a three-arm, randomized, double-masked, placebo-controlled phase 2b trial. 29 individuals received ATG plus pegylated granulocyte colony-stimulating factor (GCSF), while the other 29 received ATG only. Compared to placebo, low-dose ATG (2.5 mg/kg) results in improvements in immune cell subsets, a decrease in HbA1c, and long-term preservation of beta-cell function. While the advantages of low-dose ATG were diminished by the addition of GCSF (ClinicalTrials.gov Identifier: NCT02215200).81
Mycophenolate mofetil (MMF) alone or in combination with daclizumab (DZB) was evaluated in a multi-center, randomized, placebo-controlled, double-masked study to stop the loss of beta cells that produce insulin in newly diagnosed T1D (ClinicalTrials.gov Identifier: NCT00100178).81,82 Tocilizumab (IL-6R monoclonal antibody) was studied in a multicenter, randomized, placebo-controlled, double-blind experiment in people with newly diagnosed T1D. Immunophenotyping revealed decreases in IL-6R downstream signaling in T cells. However, there were no alterations in CD4 memory subsets, T helper 17 (Th17) cells, Tregs, or CD4+ T effector cell resistance to Treg suppression or prevent the rate of loss of residual beta cell function. During therapy, a dendritic cells (DC) subgroup declined, but when therapy ended, it returned to baseline (ClinicalTrials.gov Identifier: NCT02293837).83 Polyclonal Tregs and low-dose IL-2 were combined in a Phase I study (TILT trial), to improve Treg survival and expansion. Patients with T1D received a single infusion of autologous polyclonal Tregs, then one or two 5-day doses of human low-dose IL-2 (ld-IL-2). Therapy increased the amount of endogenous and infused Tregs, but it also increased the number of activated NK, mucosal-related invariant T, and clonal CD8+ T cells (ClinicalTrials.gov Identifier: NCT02772679).96 These findings have significant ramifications for the use of ld-IL-2 and Tregs in the treatment of autoimmune diseases in patients who already have active immunity. The safety and efficacy of a multi-dose Bacillus Calmette-Guérin (BCG) vaccination for the prevention of infectious illness in T1D in randomised, double-blind, placebo-controlled phase 2/3 trial was studied (ClinicalTrials.gov Identifier: NCT02081326).84 To modify autoimmune illnesses in an antigen-specific manner and to interfere in the pathophysiology of T1D, tolerogenic dendritic cells (tolDCs) are thought to be an appealing strategy. In T1D, C19-A3, a naturally produced proinsulin peptide, demonstrates activated immunological responses, and tolDCs that present this peptide can induce proinsulin-specific regulatory T cells (ClinicalTrials.gov Identifier: NCT04590872).97 Administration of autologous tolDCs pulsed with proinsulin peptide intradermally slow the gradual loss of beta-cell function with a shorter duration of T1D and retained c-peptide production.
Therapy Approaches for Prevention of LADA
The physician faces significant challenges in identifying and treating LADA. A consensus statement from a global expert panel discussed some key points for future action, such as a) screening for LADA, b) personalized medicine, c) the need for more randomized controlled comparative trials with hypoglycemic agents, d) further investigation of immune therapy, e) large-scale long-term studies in different patient populations, f) quality of life/lifestyle issues, g) studies including patients of different ethnic origin) the nature/quality of autoantibody assays.44
There are few clinical studies with LADA. Randomized, double-blind, placebo-controlled, dose-escalation clinical trial in a total of 47 LADA patients who received 4, 20, 100, or 500 Ag Diamyd subcutaneously at weeks 1 and 4. The ratio of CD4+ CD25+ to CD4+ CD25- cells rose. The stimulatory effects of 20 Ag Diamyd on both fasting and stimulated c-peptide were seen.98 In patients with LADA, strict glycemic management is the foundation for preventing or delaying beta-preserved cell loss and decreasing the onset of diabetes complications. Insulin, insulin sensitizers, sodium-glucose co-transporter 2 inhibitors, dipeptidyl peptidase-4 inhibitors, and receptor agonists are all used to treat LADA.70 In preliminary research, rosiglitazone, and insulin may help LADA patients maintain islet beta cell activity.99
A double-blind, randomized, controlled trial demonstrated that linagliptin medication reduces the rate of fall in C-peptide levels by raising endogenous glucagon-like Peptide 1 (GLP-1) levels to protect beta cells in LADA patients during a two-year follow-up.100 Recent data suggest that adding 2000 IU of vitamin D3 per day to the dipeptidyl peptidase-4 inhibitor saxagliptin may preserve beta cell function in LADA patients.101
Conclusion
Immunotherapies have shown promise in targeting the immune system, but they have not yet been able to achieve long-term glycemic control or preserve insulin secretion in people with type 1 diabetes (T1D) or latent autoimmune diabetes in adults (LADA). This is because the autoimmune destruction of beta cells in these conditions is often severe, and immunotherapies alone cannot reverse the damage. However, they may be able to slow the rate of beta cell loss, and the duration of glycemic control may be extended through the refinement of various immunotherapies. Single-target immunotherapies have not yet been able to fully restore T1D or LADA. However, progressive research in a clinical context should focus on improving intervention doses, doing more thorough examinations of intervention responders, and/or combining minimally effective single-target immunotherapies. This may lead to the development of more effective and durable immunotherapies for T1D and LADA.
Acknowledgments
The authors are thankful to the College of Medicine Research Centre (CMRC) and Deanship of Scientific Research, King Saud University, Riyadh, Saudi Arabia for supporting the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. American Diabetes A. Diagnosis and classification of diabetes mellitus. Diabet Care. 2013;36(Suppl 1):S67–S74.
2. ElSayed NA, Aleppo G, Aroda VR, et al. 2. Classification and diagnosis of diabetes: standards of care in diabetes-2023. Diabet Care. 2023;46(Suppl 1):S19–S40. doi:10.2337/dc23-S002
3. Sun H, Saeedi P, Karuranga S, et al. IDF Diabetes Atlas: global, regional, and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res Clin Pract. 2022;183:109119. doi:10.1016/j.diabres.2021.109119
4. Roep BO, Thomaidou S, van Tienhoven R, Zaldumbide A. Type 1 diabetes mellitus as a disease of the beta-cell (do not blame the immune system?). Nat Rev Endocrinol. 2021;17(3):150–161. doi:10.1038/s41574-020-00443-4
5. International Diabetes Federation. IDF diabetes atlas. Brussels, Belgium; 2021. Available from: https://www.diabetesatlas.org.
6. Dayan CM, Besser REJ, Oram RA, et al. Preventing type 1 diabetes in childhood. Science. 2021;373(6554):506–510. doi:10.1126/science.abi4742
7. Bluestone JA, Buckner JH, Herold KC. Immunotherapy: building a bridge to a cure for type 1 diabetes. Science. 2021;373(6554):510–516. doi:10.1126/science.abh1654
8. Tatovic D, Dayan CM. Replacing insulin with immunotherapy: time for a paradigm change in Type 1 diabetes. Diabet Med. 2021;38(12):e14696. doi:10.1111/dme.14696
9. Atkinson MA, Eisenbarth GS, Michels AW. Type 1 diabetes. Lancet. 2014;383(9911):69–82. doi:10.1016/S0140-6736(13)60591-7
10. Esposito S, Toni G, Tascini G, Santi E, Berioli MG, Principi N. Environmental Factors Associated With Type 1 Diabetes. Front Endocrinol. 2019;10:592. doi:10.3389/fendo.2019.00592
11. Erlich H, Valdes AM, Noble J, et al. HLA DR-DQ haplotypes and genotypes and type 1 diabetes risk: analysis of the type 1 diabetes genetics consortium families. Diabetes. 2008;57(4):1084–1092. doi:10.2337/db07-1331
12. Concannon P, Rich SS, Nepom GT. Genetics of type 1A diabetes. N Engl J Med. 2009;360(16):1646–1654. doi:10.1056/NEJMra0808284
13. Sharp SA, Rich SS, Wood AR, et al. Development and standardization of an improved type 1 diabetes genetic risk score for use in newborn screening and incident diagnosis. Diabet Care. 2019;42(2):200–207. doi:10.2337/dc18-1785
14. Warshauer JT, Bluestone JA, Anderson MS. New Frontiers in the Treatment of Type 1 Diabetes. Cell Metab. 2020;31(1):46–61. doi:10.1016/j.cmet.2019.11.017
15. Sherry NA, Kushner JA, Glandt M, Kitamura T, Brillantes AM, Herold KC. Effects of autoimmunity and immune therapy on beta-cell turnover in type 1 diabetes. Diabetes. 2006;55(12):3238–3245. doi:10.2337/db05-1034
16. Yang L, Zhou ZG, Tan SZ, et al. Carboxypeptidase-H autoantibodies differentiate a more latent subset of autoimmune diabetes from phenotypic type 2 diabetes among Chinese adults. Ann N Y Acad Sci. 2008;1150:263–266. doi:10.1196/annals.1447.037
17. Gottlieb PA, Delong T, Baker RL, et al. Chromogranin A is a T cell antigen in human type 1 diabetes. J Autoimmun. 2014;50:38–41. doi:10.1016/j.jaut.2013.10.003
18. Schlosser M, Banga JP, Madec AM, et al. Dynamic changes of GAD65 autoantibody epitope specificities in individuals at risk of developing type 1 diabetes. Diabetologia. 2005;48(5):922–930. doi:10.1007/s00125-005-1719-1
19. Rondas D, Crevecoeur I, D’Hertog W, et al. Citrullinated glucose-regulated protein 78 is an autoantigen in type 1 diabetes. Diabetes. 2015;64(2):573–586. doi:10.2337/db14-0621
20. Vermeulen I, Weets I, Asanghanwa M, et al. Contribution of antibodies against IA-2beta and zinc transporter 8 to classification of diabetes diagnosed under 40 years of age. Diabet Care. 2011;34(8):1760–1765. doi:10.2337/dc10-2268
21. Denroche HC, Verchere CB. IAPP and type 1 diabetes: implications for immunity, metabolism and islet transplants. J Mol Endocrinol. 2018;60(2):R57–R75. doi:10.1530/JME-17-0138
22. Pietropaolo M, Castano L, Babu S, et al. Islet cell autoantigen 69 kD (ICA69). Molecular cloning and characterization of a novel diabetes-associated autoantigen. J Clin Invest. 1993;92(1):359–371. doi:10.1172/JCI116574
23. Yang J, Danke NA, Berger D, et al. Islet-specific glucose-6-phosphatase catalytic subunit-related protein-reactive CD4+ T cells in human subjects. J Immunol. 2006;176(5):2781–2789.
24. Purcell AW, Sechi S, DiLorenzo TP. The evolving landscape of autoantigen discovery and characterization in type 1 diabetes. Diabetes. 2019;68(5):879–886. doi:10.2337/dbi18-0066
25. Moriyama H, Abiru N, Paronen J, et al. Evidence for a primary islet autoantigen (preproinsulin 1) for insulitis and diabetes in the nonobese diabetic mouse. Proc Natl Acad Sci U S A. 2003;100(18):10376–10381. doi:10.1073/pnas.1834450100
26. Doran TM, Morimoto J, Simanski S, et al. Discovery of phosphorylated peripherin as a major humoral autoantigen in type 1 diabetes mellitus. Cell Chem Biol. 2016;23(5):618–628. doi:10.1016/j.chembiol.2016.04.006
27. Nakayama M. Insulin as a key autoantigen in the development of type 1 diabetes. Diabetes Metab Res Rev. 2011;27(8):773–777. doi:10.1002/dmrr.1250
28. Strollo R, Vinci C, Man YKS, et al. Autoantibody and T cell responses to oxidative post-translationally modified insulin neoantigenic peptides in type 1 diabetes. Diabetologia. 2023;66(1):132–146. doi:10.1007/s00125-022-05812-4
29. Yang ML, Kibbey RG, Mamula MJ. Biomarkers of autoimmunity and beta cell metabolism in type 1 diabetes. Front Immunol. 2022;13:1028130. doi:10.3389/fimmu.2022.1028130
30. McLaughlin KA, Richardson CC, Ravishankar A, et al. Identification of tetraspanin-7 as a target of autoantibodies in type 1 diabetes. Diabetes. 2016;65(6):1690–1698. doi:10.2337/db15-1058
31. Gonzalez-Duque S, Azoury ME, Colli ML, et al. Conventional and neo-antigenic peptides presented by beta cells are targeted by circulating naive CD8+ T cells in type 1 diabetic and healthy donors. Cell Metab. 2018;28(6):946–960 e946. doi:10.1016/j.cmet.2018.07.007
32. Gomes KF, Semzezem C, Batista R, et al. Importance of zinc transporter 8 autoantibody in the diagnosis of type 1 diabetes in Latin Americans. Scientific Reports. 2017;7(1):207.
33. Rodriguez-Fernandez S, Almenara-Fuentes L, Perna-Barrull D, Barneda B, Vives-Pi M A century later, still fighting back: antigen-specific immunotherapies for type 1 diabetes. Immunol Cell Biol. 2021;99:461–474.
34. Insel RA, Dunne JL, Atkinson MA, et al. Staging presymptomatic type 1 diabetes: a scientific statement of JDRF, the Endocrine Society, and the American Diabetes Association. Diabet Care. 2015;38(10):1964–1974. doi:10.2337/dc15-1419
35. Katsarou A, Gudbjornsdottir S, Rawshani A, et al. Type 1 diabetes mellitus. Nat Rev Dis Primers. 2017;3(1):17016. doi:10.1038/nrdp.2017.16
36. Ziegler AG, Rewers M, Simell O, et al. Seroconversion to multiple islet autoantibodies and risk of progression to diabetes in children. JAMA. 2013;309(23):2473–2479. doi:10.1001/jama.2013.6285
37. Ahmed S, Cerosaletti K, James E, et al. Standardizing T-cell biomarkers in type 1 diabetes: challenges and recent advances. Diabetes. 2019;68(7):1366–1379. doi:10.2337/db19-0119
38. Kasmani MY, Ciecko AE, Brown AK, et al. Autoreactive CD8 T cells in NOD mice exhibit phenotypic heterogeneity but restricted TCR gene usage. Life Sci Alliance. 2022;5(10):e202201503. doi:10.26508/lsa.202201503
39. Babon JA, DeNicola ME, Blodgett DM, et al. Analysis of self-antigen specificity of islet-infiltrating T cells from human donors with type 1 diabetes. Nat Med. 2016;22(12):1482–1487. doi:10.1038/nm.4203
40. Michels AW, Landry LG, McDaniel KA, et al. Islet-derived CD4 T cells targeting proinsulin in human autoimmune diabetes. Diabetes. 2017;66(3):722–734. doi:10.2337/db16-1025
41. Delong T, Wiles TA, Baker RL, et al. Pathogenic CD4 T cells in type 1 diabetes recognize epitopes formed by peptide fusion. Science. 2016;351(6274):711–714. doi:10.1126/science.aad2791
42. Buzzetti R, Zampetti S, Maddaloni E. Adult-onset autoimmune diabetes: current knowledge and implications for management. Nat Rev Endocrinol. 2017;13(11):674–686. doi:10.1038/nrendo.2017.99
43. Buzzetti R, Maddaloni E, Gaglia J, Leslie RD, Wong FS, Boehm BO. Adult-onset autoimmune diabetes. Nat Rev Dis Primers. 2022;8(1):63. doi:10.1038/s41572-022-00390-6
44. Buzzetti R, Tuomi T, Mauricio D, et al. Management of latent autoimmune diabetes in adults: a consensus statement from an international expert panel. Diabetes. 2020;69(10):2037–2047. doi:10.2337/dbi20-0017
45. American Diabetes Association Professional Practice C. 3. Prevention or delay of type 2 diabetes and associated comorbidities: standards of medical care in diabetes-2022. Diabet Care. 2022;45(Suppl 1):S39–S45. doi:10.2337/dc22-S003
46. Andersen MK, Lundgren V, Turunen JA, et al. Latent autoimmune diabetes in adults differs genetically from classical type 1 diabetes diagnosed after the age of 35 years. Diabet Care. 2010;33(9):2062–2064. doi:10.2337/dc09-2188
47. Jorns A, Wedekind D, Jahne J, Lenzen S. Pancreas Pathology of Latent Autoimmune Diabetes in Adults (LADA) in patients and in a LADA rat model compared with type 1 diabetes. Diabetes. 2020;69(4):624–633. doi:10.2337/db19-0865
48. Cousminer DL, Ahlqvist E, Mishra R, et al. First genome-wide association study of latent autoimmune diabetes in adults reveals novel insights linking immune and metabolic diabetes. Diabet Care. 2018;41(11):2396–2403. doi:10.2337/dc18-1032
49. Mishra R, Akerlund M, Cousminer DL, et al. Genetic discrimination between LADA and childhood-onset type 1 diabetes within the MHC. Diabet Care. 2020;43(2):418–425. doi:10.2337/dc19-0986
50. Andersen MK. New insights into the genetics of latent autoimmune diabetes in adults. Curr Diab Rep. 2020;20(9):43. doi:10.1007/s11892-020-01330-y
51. Pettersen E, Skorpen F, Kvaloy K, Midthjell K, Grill V. Genetic heterogeneity in latent autoimmune diabetes is linked to various degrees of autoimmune activity: results from the Nord-Trondelag Health Study. Diabetes. 2010;59(1):302–310. doi:10.2337/db09-0923
52. Liu L, Li X, Xiang Y, et al. Latent autoimmune diabetes in adults with low-titer GAD antibodies: similar disease progression with type 2 diabetes: a nationwide, multicenter prospective study (LADA China Study 3). Diabet Care. 2015;38(1):16–21. doi:10.2337/dc14-1770
53. Hjort R, Ahlqvist E, Carlsson PO, et al. Overweight, obesity and the risk of LADA: results from a Swedish case-control study and the Norwegian HUNT Study. Diabetologia. 2018;61(6):1333–1343. doi:10.1007/s00125-018-4596-0
54. Zaharia OP, Bobrov P, Strassburger K, et al. Metabolic characteristics of recently diagnosed adult-onset autoimmune diabetes mellitus. J Clin Endocrinol Metab. 2018;103(2):429–437. doi:10.1210/jc.2017-01706
55. Dong F, Yang G, Pan HW, et al. The association of PTPN22 rs2476601 polymorphism and CTLA-4 rs231775 polymorphism with LADA risks: a systematic review and meta-analysis. Acta Diabetol. 2014;51(5):691–703. doi:10.1007/s00592-014-0613-z
56. Ramu D, Perumal V, Paul SFD. Association of common type 1 and type 2 diabetes gene variants with latent autoimmune diabetes in adults: a meta-analysis. J Diabetes. 2019;11(6):484–496. doi:10.1111/1753-0407.12879
57. Lofvenborg JE, Ahlqvist E, Alfredsson L, et al. Consumption of red meat, genetic susceptibility, and risk of LADA and type 2 diabetes. Eur J Nutr. 2021;60(2):769–779. doi:10.1007/s00394-020-02285-2
58. Hjort L, Novakovic B, Grunnet LG, et al. Diabetes in pregnancy and epigenetic mechanisms-how the first 9 months from conception might affect the child’s epigenome and later risk of disease. Lancet Diabetes Endocrinol. 2019;7(10):796–806. doi:10.1016/S2213-8587(19)30078-6
59. Carlsson S. Environmental (Lifestyle) Risk Factors for LADA. Curr Diabetes Rev. 2019;15(3):178–187. doi:10.2174/1573399814666180716150253
60. Lofvenborg JE, Andersson T, Carlsson PO, et al. Sweetened beverage intake and risk of latent autoimmune diabetes in adults (LADA) and type 2 diabetes. Eur J Endocrinol. 2016;175(6):605–614. doi:10.1530/EJE-16-0376
61. Rasouli B, Andersson T, Carlsson PO, et al. Alcohol and the risk for latent autoimmune diabetes in adults: results based on Swedish ESTRID study. Eur J Endocrinol. 2014;171(5):535–543. doi:10.1530/EJE-14-0403
62. Fang Y, Zhang C, Shi H, et al. Characteristics of the Gut microbiota and metabolism in patients with latent autoimmune diabetes in adults: a case-control study. Diabet Care. 2021;44(12):2738–2746. doi:10.2337/dc20-2975
63. Pozzilli P, Maddaloni E, Buzzetti R. Combination immunotherapies for type 1 diabetes mellitus. Nat Rev Endocrinol. 2015;11(5):289–297. doi:10.1038/nrendo.2015.8
64. Gearty SV, Dundar F, Zumbo P, et al. An autoimmune stem-like CD8 T cell population drives type 1 diabetes. Nature. 2022;602(7895):156–161. doi:10.1038/s41586-021-04248-x
65. Tatovic D, McAteer MA, Barry J, et al. Safety of the use of gold nanoparticles conjugated with proinsulin peptide and administered by hollow microneedles as an immunotherapy in type 1 diabetes. Immunother Adv. 2022;2(1):ltac002. doi:10.1093/immadv/ltac002
66. Villalba A, Rodriguez-Fernandez S, Perna-Barrull D, et al. Antigen-specific immunotherapy combined with a regenerative drug in the treatment of experimental type 1 diabetes. Sci Rep. 2020;10(1):18927. doi:10.1038/s41598-020-76041-1
67. Yue T, Sun F, Wang F, et al. MBD2 acts as a repressor to maintain the homeostasis of the Th1 program in type 1 diabetes by regulating the STAT1-IFN-gamma axis. Cell death and differentiation. 2022;29(1):218–229.
68. Cai EP, Ishikawa Y, Zhang W, et al. Genome-scale in vivo CRISPR screen identifies RNLS as a target for beta cell protection in type 1 diabetes. Nat Metab. 2020;2(9):934–945. doi:10.1038/s42255-020-0254-1
69. Ekman I, Ihantola EL, Viisanen T, et al. Circulating CXCR5(-)PD-1(hi) peripheral T helper cells are associated with progression to type 1 diabetes. Diabetologia. 2019;62(9):1681–1688. doi:10.1007/s00125-019-4936-8
70. Yin W, Luo S, Xiao Z, Zhang Z, Liu B, Zhou Z. Latent autoimmune diabetes in adults: a focus on beta-cell protection and therapy. Front Endocrinol. 2022;13:959011. doi:10.3389/fendo.2022.959011
71. Liu YF, Powrie J, Arif S, et al. Immune and metabolic effects of antigen-specific immunotherapy using multiple beta-cell peptides in type 1 diabetes. Diabetes. 2022;71(4):722–732. doi:10.2337/db21-0728
72. Herold KC, Bundy BN, Long SA, et al. An anti-CD3 antibody, teplizumab, in relatives at risk for type 1 diabetes. N Engl J Med. 2019;381(7):603–613. doi:10.1056/NEJMoa1902226
73. Orban T, Bundy B, Becker DJ, et al. Co-stimulation modulation with Abatacept in patients with recent-onset type 1 diabetes: a randomised, double-blind, placebo-controlled trial. Lancet. 2011;378(9789):412–419. doi:10.1016/S0140-6736(11)60886-6
74. Sumpter KM, Adhikari S, Grishman EK, White PC. Preliminary studies related to anti-interleukin-1beta therapy in children with newly diagnosed type 1 diabetes. Pediatr Diabetes. 2011;12(7):656–667. doi:10.1111/j.1399-5448.2011.00761.x
75. Moran A, Bundy B, Becker DJ, et al. Interleukin-1 antagonism in type 1 diabetes of recent onset: two multicentre, randomised, double-blind, placebo-controlled trials. Lancet. 2013;381(9881):1905–1915. doi:10.1016/S0140-6736(13)60023-9
76. Mastrandrea L, Yu J, Behrens T, et al. Etanercept treatment in children with new-onset type 1 diabetes: pilot randomized, placebo-controlled, double-blind study. Diabet Care. 2009;32(7):1244–1249. doi:10.2337/dc09-0054
77. Tavira B, Barcenilla H, Wahlberg J, Achenbach P, Ludvigsson J, Casas R. Intralymphatic glutamic acid decarboxylase-alum administration induced Th2-like-specific immunomodulation in responder patients: a pilot clinical trial in type 1 diabetes. J Diabetes Res. 2018;2018:9391845. doi:10.1155/2018/9391845
78. Gitelman SE, Bundy BN, Ferrannini E, et al. Imatinib therapy for patients with recent-onset type 1 diabetes: a multicentre, randomised, double-blind, placebo-controlled, phase 2 trial. Lancet Diabetes Endocrinol. 2021;9(8):502–514. doi:10.1016/S2213-8587(21)00139-X
79. Pescovitz MD, Greenbaum CJ, Bundy B, et al. B-lymphocyte depletion with rituximab and beta-cell function: two-year results. Diabet Care. 2014;37(2):453–459. doi:10.2337/dc13-0626
80. Sherry N, Hagopian W, Ludvigsson J, et al. Teplizumab for treatment of type 1 diabetes (Protégé study): 1-year results from a randomised, placebo-controlled trial. Lancet. 2011;378(9790):487–497. doi:10.1016/S0140-6736(11)60931-8
81. Haller MJ, Long SA, Blanchfield JL, et al. Low-dose anti-thymocyte globulin preserves C-Peptide, Reduces HbA(1c), and increases regulatory to conventional T-cell ratios in new-onset type 1 diabetes: two-year clinical trial data. Diabetes. 2019;68(6):1267–1276. doi:10.2337/db19-0057
82. Gottlieb PA, Quinlan S, Krause-Steinrauf H, et al. Failure to preserve beta-cell function with mycophenolate mofetil and daclizumab combined therapy in patients with new- onset type 1 diabetes. Diabet Care. 2010;33(4):826–832. doi:10.2337/dc09-1349
83. Greenbaum CJ, Serti E, Lambert K, et al. IL-6 receptor blockade does not slow beta cell loss in new-onset type 1 diabetes. JCI Insight. 2021;6(21):e150074.
84. Faustman DL, Lee A, Hostetter ER, et al. Multiple BCG vaccinations for the prevention of COVID-19 and other infectious diseases in type 1 diabetes. Cell Rep Med. 2022;3(9):100728. doi:10.1016/j.xcrm.2022.100728
85. Hartemann A, Bensimon G, Payan CA, et al. Low-dose interleukin 2 in patients with type 1 diabetes: a Phase 1/2 randomised, double-blind, placebo-controlled trial. Lancet Diabetes Endocrinol. 2013;1(4):295–305. doi:10.1016/S2213-8587(13)70113-X
86. Rios-Rios WJ, Sosa-Luis SA, Torres-Aguilar H. Current advances in using tolerogenic dendritic cells as a therapeutic alternative in the treatment of type 1 diabetes. World J Diabetes. 2021;12(5):603–615. doi:10.4239/wjd.v12.i5.603
87. Quattrin T, Haller MJ, Steck AK, et al. Golimumab and beta-cell function in youth with new-onset type 1 diabetes. N Engl J Med. 2020;383(21):2007–2017. doi:10.1056/NEJMoa2006136
88. Rigby MR, Harris KM, Pinckney A, et al. Alefacept provides sustained clinical and immunological effects in new-onset type 1 diabetes patients. J Clin Invest. 2015;125(8):3285–3296. doi:10.1172/JCI81722
89. von Herrath M, Bain SC, Bode B, et al. Anti-interleukin-21 antibody and liraglutide for the preservation of beta-cell function in adults with recent-onset type 1 diabetes: a randomised, double-blind, placebo-controlled, phase 2 trial. Lancet Diabetes Endocrinol. 2021;9(4):212–224. doi:10.1016/S2213-8587(21)00019-X
90. Beran D, Abidha C, Adler A, et al. Teplizumab approval for type 1 diabetes in the USA. Lancet Diabetes Endocrinol. 2023;11(2):78–80. doi:10.1016/S2213-8587(22)00384-9
91. White PC, Adhikari S, Grishman EK, Sumpter KM. A phase I study of anti-inflammatory therapy with rilonacept in adolescents and adults with type 1 diabetes mellitus. Pediatr Diabetes. 2018;19(4):788–793. doi:10.1111/pedi.12634
92. Elding Larsson H, Lundgren M, Jonsdottir B, Cuthbertson D, Krischer J, Di A. Safety and efficacy of autoantigen-specific therapy with 2 doses of alum-formulated glutamate decarboxylase in children with multiple islet autoantibodies and risk for type 1 diabetes: a randomized clinical trial. Pediatr Diabetes. 2018;19(3):410–419. doi:10.1111/pedi.12611
93. Mathieu C, Lahesmaa R, Bonifacio E, Achenbach P, Tree T. Immunological biomarkers for the development and progression of type 1 diabetes. Diabetologia. 2018;61(11):2252–2258. doi:10.1007/s00125-018-4726-8
94. Ludvigsson J, Routray I, Elluru S, et al. Combined vitamin D, ibuprofen and glutamic acid decarboxylase-alum treatment in recent onset Type I diabetes: lessons from the DIABGAD randomized pilot trial. Future Sci OA. 2020;6(7):FSO604. doi:10.2144/fsoa-2020-0078
95. Pescovitz MD, Greenbaum CJ, Krause-Steinrauf H, et al. Rituximab, B-lymphocyte depletion, and preservation of beta-cell function. N Engl J Med. 2009;361(22):2143–2152. doi:10.1056/NEJMoa0904452
96. Dong S, Hiam-Galvez KJ, Mowery CT, et al. The effect of low-dose IL-2 and Treg adoptive cell therapy in patients with type 1 diabetes. JCI Insight. 2021;6(18). doi:10.1172/jci.insight.147474
97. Nikolic T, Zwaginga JJ, Uitbeijerse BS, et al. Safety and feasibility of intradermal injection with tolerogenic dendritic cells pulsed with proinsulin peptide-for type 1 diabetes. Lancet Diabetes Endocrinol. 2020;8(6):470–472. doi:10.1016/S2213-8587(20)30104-2
98. Agardh CD, Cilio CM, Lethagen A, et al. Clinical evidence for the safety of GAD65 immunomodulation in adult-onset autoimmune diabetes. J Diabetes Complications. 2005;19(4):238–246. doi:10.1016/j.jdiacomp.2004.12.003
99. Zhou Z, Li X, Huang G, et al. Rosiglitazone combined with insulin preserves islet β cell function in adult‐onset latent autoimmune diabetes (LADA). Diabetes Metab Res Rev. 2005;21(2):203–208. doi:10.1002/dmrr.503
100. Yang Z, Zhou Z, Li X, Huang G, Lin J. Rosiglitazone preserves islet β-cell function of adult-onset latent autoimmune diabetes in 3 years follow-up study. Diabetes Res Clin Pract. 2009;83(1):54–60. doi:10.1016/j.diabres.2008.09.044
101. Zhang Z, Yan X, Wu C, et al. Adding vitamin D3 to the dipeptidyl peptidase‐4 inhibitor saxagliptin has the potential to protect β‐cell function in LADA patients: a 1‐year pilot study. Diabetes Metab Res Rev. 2020;36(5):e3298. doi:10.1002/dmrr.3298
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.