Back to Journals » Clinical, Cosmetic and Investigational Dermatology » Volume 19
Exosomes in Psoriasis: From Pathogenic Mechanisms to Therapeutic Innovations
Authors Xu Q, Liu W, Zhang S, Zhou L
Received 4 March 2026
Accepted for publication 4 June 2026
Published 15 June 2026 Volume 2026:19 606845
DOI https://doi.org/10.2147/CCID.S606845
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Prof. Dr. Rungsima Wanitphakdeedecha
Qi Xu,1,* Wenting Liu,2,* Siqing Zhang,3 Lin Zhou1
1Department of Dermatology, The Traditional Chinese Medicine Hospital of Longquanyi, Chengdu, Sichuan Province, 610100, People’s Republic of China; 2Department of Intensive Care Unit, Foshan Hospital of Traditional Chinese Medicine, Foshan, Guangdong Province, 528099, People’s Republic of China; 3Shishan Community Health Service Center, Foshan City, Guangdong Province, 528234, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Lin Zhou, Department of Dermatology, The Traditional Chinese Medicine Hospital of Longquanyi, Chengdu, Sichuan Province, 610100, People’s Republic of China, Email [email protected]
Abstract: Psoriasis is a chronic recurrent inflammatory disease driven by keratinocytes and immune cells. Exosomes, as key mediators of intercellular communication, play multidimensional roles in this disease. In terms of pathogenic mechanisms, exosomes released from psoriatic lesions carry non‑coding RNAs and proteins that program T‑cell polarization, drive M1 macrophage activation, and amplify keratinocyte inflammation, thereby sustaining the IL‑23/Th17 immune axis. Translational breakthroughs have repurposed these same vesicles into diagnostic and therapeutic tools. Circulating exosomal fingerprints offer non‑invasive biomarkers for disease activity and psoriatic arthritis differentiation. Leveraging their biocompatibility and low immunogenicity, exosomes from mesenchymal stem cells, plants, and microbes serve as cell‑free platforms achieving immune regulation, antioxidant effects, and microecological repair. Engineering strategies—including cargo loading, membrane surface modification and intelligent microneedle delivery systems—further enhance targeting and efficacy. Despite these advances, clinical translation faces fundamental challenges: lack of production standardization, undefined core active components and insufficient high‑level clinical evidence. Future efforts should prioritize international standards, rational design, and rigorous trials to accelerate exosome‑based precision medicine for psoriasis.
Keywords: psoriasis, exosomes, biomarkers, cell-free therapy, immunomodulation
Introduction
Psoriasis is a chronic recurrent inflammatory disease characterized by skin erythema, scales, and immune infiltration.1,2 Its pathological essence is far from simple skin barrier dysfunction but rather an imbalance in a complex network formed by keratinocytes, T cells, dendritic cells, and other immune cells, along with the cytokines they secrete, triggered by environmental factors in genetically susceptible individuals.3,4 This persistent inflammatory microenvironment not only drives the stubbornness of skin lesions but also forms the basis for systemic comorbidities such as psoriatic arthritis, leading to the chronic and relapse-prone nature of the disease. Currently, the core pathological mechanism of this disease has been clearly focused on the abnormal activation of the interleukin-23 (IL-23)/T helper 17 (Th17) axis.5,6 With the innovation of treatment concepts, some perspectives further propose that early precise intervention combined with advanced therapies to reshape immune homeostasis holds promise for achieving a clinical cure, and this has now become a groundbreaking research direction.7
Exosomes, a class of extracellular vesicles actively secreted by cells with a diameter of approximately 30–150 nanometers and abundant bioactive molecules, have garnered unprecedented attention because of their central role in intercellular communication.8 The discovery of exosomes as intercellular messengers dates back to the 1980s, but their potential as disease biomarkers and therapeutic tools accelerated notably after the 2013 Nobel Prize in Physiology or Medicine, and since 2021, research on exosomes in psoriasis therapy has witnessed explosive growth.9,10 In the complex pathological environment of psoriasis, exosomes play multiple and seemingly contradictory dual roles: on the one hand, exosomes derived from lesional keratinocytes or activated immune cells can act as precise messengers of inflammatory signals, delivering specific inflammatory instructions accurately to neighboring and distal target cells and participating in and amplifying the disease-specific immune imbalance network; on the other hand, this natural delivery system, owing to its excellent biocompatibility, low immunogenicity, and efficient targeting capabilities, is regarded as a highly promising “cell-free therapy” carrier and a diagnostic tool for liquid biopsy.11 Exosomes also carry cytokines, as evidenced by elevated circulating exosomal IL‑23 and TNF‑α mRNAs12 and increased IL‑17A exosome levels13 in psoriasis, directly implicating the IL‑23/Th17 axis.
Moreover, the translational potential of exosomes in the fields of diagnosis and treatment is increasingly evident. At the diagnostic level, the specific “molecular fingerprints” carried by exosomes in patient circulation, such as specific phospholipid profiles, differentially expressed miRNAs, and disease‑associated protein cargo, demonstrate the potential to assess disease activity, subtypes, and treatment responses. This facilitates noninvasive monitoring.14,15 At the therapeutic level, exosomes derived from mesenchymal stem cells, plants, and even probiotics, which leverage their natural immunomodulatory and tissue-repairing properties, are considered promising cell-free therapeutic agents. They can regulate the Th17/Treg balance by delivering microRNA molecules while intervening in the IL-23/Th17 axis.16,17 Furthermore, through engineered modifications to equip exosomes with targeting ligands or therapeutic payloads, their precision therapeutic efficacy can be significantly enhanced, highlighting their immense potential for development into intelligent delivery systems.17
Currently, research in this field has progressed from phenomenological descriptions to functional molecular analysis and animal model validation, with preliminary clinical exploration initiated, positioning it at a critical juncture transitioning toward clinical translation and application. A recent review18 covered exosome roles in psoriasis pathogenesis, drug delivery, and therapy, but several gaps remain: the diagnostic potential of exosomal cargoes as liquid biopsy biomarkers is underexplored, engineering strategies for targeted therapy lack systematic comparison, and an integrated framework connecting pathogenic mediators to therapeutic vectors is missing. Therefore, systematically organizing the multidimensional roles and translational logic of exosomes in psoriasis is crucial. This review focuses on interconnected areas: (1) pathogenic mechanisms (exosomal cytokines and non-coding RNAs driving T‑cell polarization, innate immune amplification, and tissue remodeling); (2) non‑invasive biomarkers (exosomal cargoes for severity assessment, treatment monitoring, and psoriatic arthritis differentiation); and (3) therapeutic applications (diverse natural sources and engineering/delivery strategies). By bridging pathogenesis, diagnosis, and therapy, this review provides a framework for exosome‑based precision medicine in psoriasis.
Pathogenic Mechanisms of Exosomes in Psoriasis
Psoriatic skin lesions represent a pathological ecosystem with highly active intercellular communication. Exosomes serve as core signaling vehicles, delivering bioactive molecules (proteins, nucleic acids) that precisely regulate immune responses and tissue structure. The pathogenic role of these exosomes is characterized by multidimensionality and cascade amplification (Figure 1). Among the various pathways, three interconnected themes emerge as dominant: T‑cell polarization (core adaptive event), the keratinocyte–neutrophil amplification loop (key innate amplifier), and tissue remodeling (downstream effector).
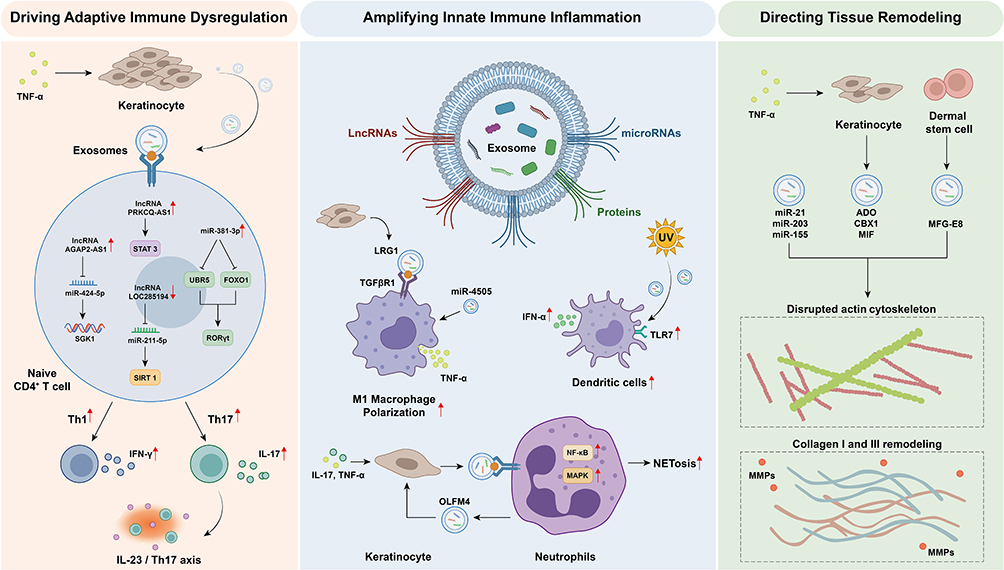 |
Figure 1 Exosome-mediated pathogenic mechanisms in psoriasis. Activated keratinocytes and immune cells release exosomes carrying diverse bioactive molecules, driving pathogenesis through three core pathways: 1) Adaptive immune dysregulation: Keratinocyte-derived exosomes deliver regulatory non-coding RNAs to CD4+ T cells, programming their differentiation into pro-inflammatory Th1/Th17 subsets. 2) Innate immune amplification: Exosome-mediated crosstalk between keratinocytes and neutrophils establishes a positive inflammatory feedback loop, promoting macrophage M1 polarization, neutrophil extracellular trap formation, and sustained keratinocyte hyperactivation. 3) Tissue remodeling: Exosomal cargo disrupts cytoskeletal dynamics and extracellular matrix homeostasis in resident skin cells, driving epidermal hyperplasia and dermal disorganization. Red arrows pointing upward indicate upregulation, while red arrows pointing downward indicate downregulation. In this figure, red upward arrows (↑) indicate upregulation or activation, red downward arrows (↓) indicate downregulation or inhibition, and black arrows (→) denote the direction of processes or interactions without implying quantitative change. |
Exosomes Participate in Regulating Adaptive Immune Responses
The abnormal differentiation of CD4+ T cells into Th1 and Th17 cells is a primary driver of psoriasis. Among the numerous exosomal cargoes identified, non‑coding RNAs delivered by keratinocyte (KC)‑derived exosomes have been most consistently shown to reprogram CD4+ T cells in vitro and in animal models. This process begins with the establishment of an inflammatory microenvironment: key cytokines such as IL-17A can significantly alter the release kinetics and cargo composition of KC-derived extracellular vesicles, preparing the carrier for subsequent communication.19
When these exosomes are taken up by CD4+ T cells, the specific lncRNA they carry initiates precise posttranscriptional regulation. For example, under TNF-α stimulation, the lncRNA AGAP2-AS1 enriched in KC exosomes can act as a “molecular sponge” to adsorb miR-424-5p, thereby relieving its inhibition of the downstream target gene SGK1, driving the differentiation of naive CD4+ T cells into Th1/Th17 cells.20 Another lncRNA, PRKCQ-AS1, directly activates the STAT3 signaling pathway in T cells, promoting Th17 differentiation and IL-17 secretion.21 Moreover, miR-381-3p in KC exosomes simultaneously targets and inhibits UBR5 and FOXO1, synergistically stabilizing RORγt and increasing the suppression of the Th1/Th17 transcriptional program, thereby promoting the production of IFN-γ and IL-17A/F.22 All three mechanisms converge on the IL‑23/Th17 axis – the central hub of psoriasis – but operate through distinct molecular routes: lncRNA sponging, direct STAT3 activation, and miRNA‑mediated stabilization of RORγt. Among them, direct STAT3 activation by PRKCQ‑AS1 likely represents the most efficient route, though all contribute to Th17 skewing. Additionally, environmental factors influence this programming process. After stimulating cells, environmental pollutants such as benzo[a]pyrene release exosomes that activate the aryl hydrocarbon receptor signaling pathway, inducing Th1/Th17 differentiation in immune cells and mediating the long-term programming effects of environmental exposure on the immune system.23
The loss of protective regulatory mechanisms also exacerbates immune imbalance. In psoriatic patients, the downregulation of lncRNA LOC285194 expression in KC exosomes leads to the loss of its protective function in suppressing Th17 differentiation by adsorbing miR-211–5p to maintain the expression of the deacetylase SIRT1, which is equivalent to removing an intrinsic “immune checkpoint”24 Furthermore, bioinformatics analysis revealed a regulatory network formed by exosome-related genes, supporting the central role of exosomes in adaptive immune dysregulation at the systemic level. For example, the exosome-related gene CD274 (encoding PD-L1) regulates T-cell activation through the PD-1/PD-L1 axis, CXCL13 mediates inflammatory cell recruitment, and BIRC5 participates in pathological hyperplasia by inhibiting keratinocyte apoptosis.25 Taken together, the evidence strongly supports that KC‑derived exosomal non‑coding RNAs are dominant upstream regulators of Th17 polarization – arguably the most critical pathogenic axis in psoriasis.
Exosomes Participate in Regulating Innate Immune Responses
Abnormal activation and sustained amplification of the innate immune system constitute the cornerstone of psoriasis inflammation. Exosomes act as key messengers for intercellular inflammatory signal transmission and form a vicious positive feedback loop. Among the various innate pathways, the bidirectional crosstalk between KCs and neutrophils stands out as the most powerful amplifier, while macrophage and dendritic cell modulation play secondary supporting roles.
First, KC exosomes directly regulate the functional state of innate immune cells, primarily by reprogramming macrophages and activating dendritic cells. For example, under inflammatory stimulation, KCs release extracellular vesicles rich in leucine-rich alpha-2-glycoprotein 1 (LRG1), which drive macrophages toward the proinflammatory M1 phenotype by binding to TGFβR1 on their surface.26 Vitamin D receptor-deficient KC-derived exosomes promote M1 polarization of macrophages by delivering miR-4505.27 Ultraviolet irradiation systematically modifies the miRNA cargo of KC exosomes, which, after being taken up by plasmacytoid dendritic cells, activate Toll-like receptor 7 and strongly induce type I interferon production, thereby initiating inflammatory responses.28 While these pathways contribute to the inflammatory milieu, they are likely secondary amplifiers rather than primary initiators, as their blockade does not completely abrogate psoriatic inflammation in animal models.
Second, a robust positive inflammatory feedback loop is formed between KCs and neutrophils through exosomes, which is a crucial mechanism for the acute exacerbation of skin lesions. On the one hand, exosomes released by KCs stimulated by IL-17A and TNF-α can activate the NF-κB/p38 MAPK signaling pathway within neutrophils, inducing the formation of neutrophil extracellular traps (NETs) and the release of inflammatory factors, directly leading to tissue damage.29 On the other hand, activated neutrophils (especially in generalized pustular psoriasis) release exosomes rich in olfactomedin-4 (OLFM4), which can “counterattack” KCs and activate their NF-κB, p38 MAPK, and ERK1/2 pathways, leading to the overexpression of IL-36γ, TNF-α, and other factors in KCs. This recruits more neutrophils, forming a self-amplifying inflammatory vortex.30 This KC–neutrophil loop is self‑sustaining and directly linked to acute exacerbations; therefore, it is prioritized as the most critical innate amplifier. This dysregulation also has systemic effects: elevated levels of platelet- and erythrocyte-derived microvesicles in the patient’s circulation can induce endothelial inflammation and activate the coagulation system, collectively shaping a systemic prothrombotic and proinflammatory state.31
Exosomes Participating in the Cytoskeleton and Matrix Remodeling
In addition to immune regulation, exosomes are also deeply involved in the pathological destruction and abnormal reconstruction of local tissue structures in psoriatic skin lesions, synergistically mediating abnormal epidermal hyperplasia and dermal connective tissue disorganization. Unlike immune‑driven pathways, these tissue remodeling events are largely downstream consequences of inflammation, although they feed back into disease chronicity and clinical severity.
Exosomes directly influence the cytoskeletal dynamics and extracellular matrix metabolic balance of cells by delivering specific regulatory molecules. For example, specific miRNAs (such as miR-21, miR-203, and miR-155) are transferred across cells via exosomes. After entering target cells, they precisely regulate downstream target genes, affecting processes such as actin polymerization during cytoskeletal reorganization, collagen synthesis, degradation, and the expression of matrix metalloproteinases.32 Environmental factors such as TNF-α disrupt KC homeostasis and trigger the secretion of exosomes carrying specific proteins (eg, ADO, which is involved in angiogenesis; the epigenetic regulator CBX1; and the proinflammatory factor MIF). These vesicles act as signaling carriers that drive early tissue alterations.33
Moreover, the dysfunction of other cells in the local microenvironment of skin lesions also contributes to tissue remodeling through exosomes. For example, the function of dermal mesenchymal stem cells may be altered, and the expression of proteins related to skin barrier function (such as milk fat globule epidermal growth factor 8) in the exosomes they secrete is downregulated, thereby disrupting the microenvironment that maintains skin homeostasis.34 Mesenchymal stem cells and the vesicles they secrete can also directly affect the expression of proliferation-related markers in KCs by delivering bioactive molecules, contributing to the disruption of tissue homeostasis.35 Overall, as multifunctional signaling platforms, exosomes carry diverse cargoes that collectively form a complex regulatory network. While exacerbating immune inflammation, they also directly instruct structural lesions in local tissues, reflecting their multidimensional role throughout the pathogenesis of psoriasis.36 In the pathogenic hierarchy, this tissue remodeling layer is positioned as an effector consequence – essential for clinical manifestations (hyperplasia, scaling) but not a primary driver; thus, therapeutic strategies may prioritize upstream immune targets.
Circulating Exosomes as Biomarkers for the Diagnosis and Subtyping of Psoriasis
Protected by their lipid bilayer, circulating exosomes can stably carry molecular fingerprints, such as proteins and nucleic acids derived from parent cells, providing an ideal “liquid biopsy” window for the dynamic monitoring of psoriasis. Their noninvasive and repeatable acquisition characteristics make it possible to assess disease activity in real time, monitor treatment response, and achieve precise clinical subtype differentiation. Routine clinical assessment of psoriasis still depends on the Psoriasis Area and Severity Index (PASI) and body surface area (BSA) – scores that vary considerably between observers. Skin biopsy, although definitive, is invasive and unsuitable for frequent monitoring. Serum markers such as C‑reactive protein (CRP) lack disease specificity. Exosome‑based miRNA profiling offers a quantitative, easily repeatable, and patient‑friendly strategy that addresses many of these gaps. Currently, research focuses primarily on the analysis of specific miRNA profiles within exosomes. These molecules are not only closely associated with disease states but also reveal underlying pathological mechanisms through their regulatory pathways.
Practical Workflow for Exosome‑based Clinical Diagnostics
A clinically reliable exosome‑based diagnostic workflow must integrate robust isolation, standardized characterization, and clear criteria for result interpretation. For tissue‑derived EVs, a validated protocol37 involves gentle mincing of tissues into 2×2×2 mm pieces, followed by enzymatic digestion with collagenase D and DNase I to release EVs trapped in the extracellular space. Subsequent differential ultracentrifugation separates large and small EVs, which are then resolved by iodixanol density gradient into six subpopulations: large/small low‑density (LD) and high‑density (HD) EVs. For liquid biopsies (eg, blood, urine), size‑exclusion chromatography or ultracentrifugation combined with density gradients is recommended to remove highly abundant lipoproteins and protein aggregates. Following MISEV2023 guidelines,38 each preparation should be characterized by orthogonal methods: particle concentration (nanoparticle tracking analysis), EV marker proteins including transmembrane (eg, CD9, CD63, CD81) and cytosolic markers (eg, ALIX, TSG101), and exclusion of non‑vesicular contaminants (lipoproteins, Tamm‑Horsfall protein). The ten‑year evolution of MISEV39 emphasizes transparent reporting of pre‑analytical variables, separation yields and detection limits – all essential for clinical translation.
To implement exosomes as a routine diagnostic tool, we envisage a stepwise pipeline (Figure 2): (1) sample collection (blood, urine or tissue) under standardized pre‑analytical conditions; (2) EV separation using a clinically compatible method (eg, automated SEC or density gradient); (3) quality control (particle count, protein/RNA yield, marker identification); (4) molecular profiling (eg, qPCR for IL‑23/TNF‑α mRNA or proteomics); and (5) algorithm‑based scoring (eg, machine‑learning) to discriminate patients from healthy controls. Despite current challenges – such as the 22‑h processing time for tissue EVs, limited applicability to frozen samples, and potential surface protein alteration by enzymatic treatment – adherence to MISEV recommendations and further technological automation will pave the way for exosome‑based liquid biopsy in psoriasis management.
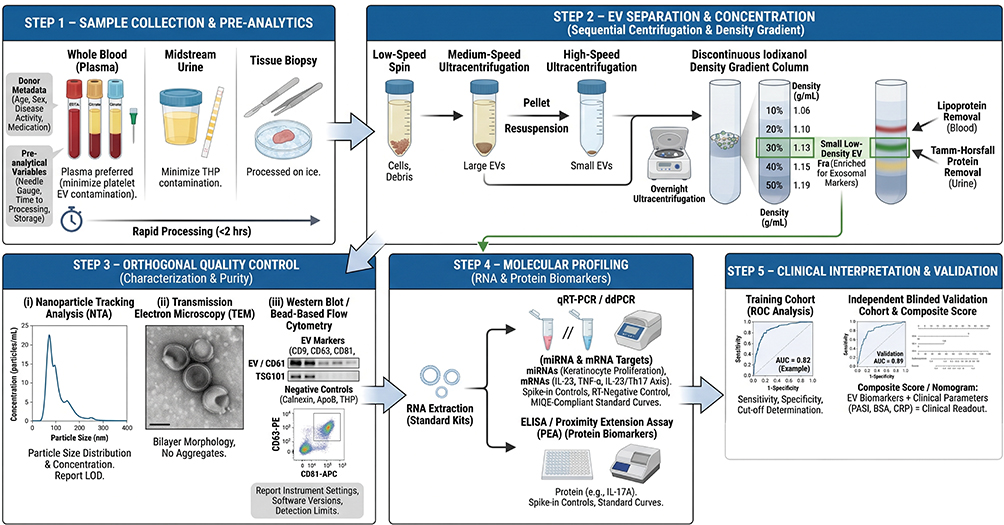 |
Figure 2 Proposed workflow for exosome‑based clinical diagnostics in psoriasis. The workflow follows MISEV2023 guidelines and includes (1) sample collection (plasma, urine or tissue, processed on ice within 1 h); (2) EV separation (differential ultracentrifugation plus iodixanol density gradient); (3) orthogonal quality control (NTA, TEM, EV markers CD9/CD63/CD81/ALIX/TSG101, and exclusion of non‑EV contaminants such as calnexin); (4) molecular profiling (qRT‑PCR or ddPCR for exosomal miRNAs or mRNAs); and (5) clinical interpretation (ROC‑derived cut‑offs, composite scores with PASI/CRP, independent validation). No exosome‑based psoriasis test has yet received regulatory clearance. |
Disease Severity Assessment and Treatment Efficacy Monitoring
The correlation between circulating exosomal miRNA profiles and the clinical severity of psoriasis has been confirmed by multiple studies, with some molecules demonstrating exceptional diagnostic efficacy and dynamic monitoring potential. A study involving 63 patients revealed that the level of miR-625-3p in keratinocyte-derived extracellular vesicles was strongly positively correlated with the PASI and BSA scores, with an area under the curve (AUC) as high as 0.9515 for distinguishing mild, moderate, and severe patients. Mechanistically, this miRNA activates the IGF-1/Akt pro-proliferative pathway in keratinocytes by targeting and inhibiting insulin-like growth factor-binding protein 3 (IGFBP3). Notably, its levels are significantly decreased in treatment responders, suggesting its potential as a sensitive biomarker for monitoring therapeutic efficacy.40 Another case‒control study of 40 psoriasis patients and 43 healthy controls revealed that the expression of miR-16-5p, miR-21-5p, and miR-155-5p in total circulating vesicles of patients was generally downregulated. Among these, miR-21-5p exhibited the best diagnostic performance (AUC=0.7059), while the level of miR-16-5p and its ratio to miR-21-5p were positively correlated with the PASI score. The target genes coregulated by these differentially expressed miRNAs were enriched in key pathways, such as the TNF and PI3K-Akt pathways.41 Importantly, some exosomal miRNAs may exert protective feedback effects. For example, a clinical study of 15 psoriasis patients and 10 healthy controls found that serum exosomal miR‑6785‑5p was significantly upregulated.42 This miRNA can be taken up by KCs, where it targets MNK2 and inhibits its downstream phosphorylated eukaryotic translation initiation factor 4E (p-eIF4E) axis, thereby suppressing abnormal proliferation and inflammation. Collectively, these studies demonstrate that circulating exosomal miRNAs are sensitive indicators reflecting disease severity and that their dynamic changes may encompass rich information ranging from pathogenic drivers to protective feedback mechanisms.
Differential Diagnosis of Psoriatic Arthritis
Accurate differentiation between cutaneous-only psoriasis (PsO) and psoriatic arthritis (PsA) is crucial for clinical decision-making, and circulating exosomes provide a highly promising noninvasive tool for this purpose. The current diagnosis of PsA relies on the CASPAR criteria and imaging techniques (ultrasound and MRI) that are often insensitive to subclinical joint inflammation. A blood‑based exosomal miRNA panel could identify individuals at risk before erosive damage occurs.
Studies have revealed systematic differences in the molecular profiles of plasma exosomes between PsA patients and PsO patients. In the Han Chinese population, a study of 42 patients (25 PsO, 17 PsA) identified significantly reduced plasma extracellular vesicle miR‑218‑5p in PsA versus PsO via next‑generation sequencing.43 ROC analysis showed its diagnostic value (AUC=0.758, 95% CI: 0.535–0.982, P=0.023; sensitivity 73.3%, specificity 87.5%). Western blotting confirmed elevated ADP-ribosylation factor 6 (ARF6) and tumor necrosis factor alpha-induced protein 3 (TNFAIP3) in PsA, with ARF6 positively correlated with TNFAIP3 and inversely related to miR‑218‑5p. Both genes are involved in the regulation of inflammatory pathways such as NF-κB, suggesting that this axis may play a role in PsA-specific joint inflammation. Broader omics studies have confirmed the universality of these differences. In 12 psoriasis vulgaris (PsV), 12 PsA, and 12 healthy controls, serum EV miRNAs were profiled by RNA sequencing after size‑exclusion chromatography. Compared with PsV, PsA had eight differentially enriched miRNAs (FDR ≤ 0.1), including upregulated miR‑33a‑5p (1.82‑fold) and downregulated miR‑671‑3p (0.43‑fold); miR‑10b‑5p was also lower in PsA versus controls (0.46‑fold). Pathway analysis of target genes implicated arthritis‑related processes (methyl CpG binding protein 2, circadian rhythm, dopamine release cycle), suggesting that circulating EV miRNAs reflect joint‑specific pathology in PsA.44 Additionally, studies indicate that the expression of let-7b-5p and miR-30e-5p in the plasma exosomes of PsA patients is significantly lower than that in the plasma exosomes of PsO patients.45 The former can target and inhibit the key proinflammatory factor IL-6, while the latter targets genes such as BMI1, an activator of the NF-κB pathway. Their downregulation may promote joint inflammation.46 These findings collectively highlight that circulating exosomes carry molecular information reflecting systemic disease involvement. By analyzing their specific miRNA profiles, efficient and noninvasive differentiation between PsA and PsO may be achieved, guiding early therapeutic interventions. However, direct comparisons with established imaging or serological biomarkers (eg, ultrasound‑detected enthesitis, CRP) are still lacking, and such validation studies should be a priority for translation.
Exosomes as a Promising Therapeutic Strategy
Therapeutic exosomes, which leverage their inherent biocompatibility, low immunogenicity, and targeting capabilities, are emerging as a cutting-edge approach in the field of cell-free therapy, with a trend toward diversified sources. Current research not only systematically elucidates the core effector molecules and signaling pathways of exosomes from various sources but also enhances their functions through engineering strategies, revealing a developmental trajectory from natural biological agents to intelligent drug systems.
Mesenchymal Stem Cell-Derived Exosomes
Mesenchymal stem cell-derived exosomes (MSC-Exos) have emerged as the most promising therapeutic carriers because of their ability to inherive the immunomodulatory and tissue repair properties of MSCs while avoiding the risks associated with live cell transplantation, such as potential tumorigenesis and embolism. Their efficacy has been extensively validated in preclinical studies, with meta-analyses confirming their ability to significantly reduce the PASI score and epidermal thickness, as well as downregulate key inflammatory cytokines such as TNF-α and IL-17A.47
The efficacy of these methods is closely related to the tissue source and pretreatment strategy. A comparative study revealed that human umbilical cord-derived MSCs (hUCMSC-Exos) are superior to adipose-derived exosomes in reversing epidermal acanthosis and reducing CD3+ T-cell and CD68+ macrophage infiltration without the common side effects of topical corticosteroids, such as skin thinning. Their superiority may be associated with the enrichment of hepatocyte growth factor (HGF) revealed by proteomics.48 More innovatively, the “inflammatory cytokine pretreatment” strategy involves stimulating MSCs with a mixture of IL-17, IL-22, and TNF-α (10–50 ng/mL for 24 h), after which the secreted exosomes (MSC-Exo 3C) exhibit a qualitative improvement in efficacy, even surpassing that of the parent cells. In an imiquimod‑induced psoriasis‑like mouse model, subcutaneous injection of MSC‑Exo 3C on days 1 and 4 significantly reduced the cumulative PASI score from 11.5 (control) to 4.0 on day 8 (65.2% reduction), outperforming intact hUCB‑MSCs (PASI 7.0) and the parent cells. The molecular mechanism involves the “reprogramming” of exosomes by pretreatment, enabling them to carry more targeted immunomodulatory signals, such as increasing the secretion of regulatory cytokines such as transforming growth factor-β1 (TGF-β1), thereby more effectively correcting the Th17/Treg imbalance in vivo.49
Analysis of its active components revealed precise molecular targets. miR-100-5p in human placental MSC-Exos is one of the core effector molecules. It directly targets the messenger RNA of the mammalian target of rapamycin (mTOR), inhibiting the phosphorylation and activation of mTOR and its downstream ribosomal protein S6 kinase 1 (S6K1). Topical MSC‑Exos reduced epidermal thickness by ≈50% and the PASI score from 7.83 to 2.00 on day 4. The suppression of this pathway blocks keratinocyte hyperproliferation and alleviates inflammation, achieving dual anti‑proliferative and anti‑inflammatory effects.50 Additionally, MSC-Exos have been shown to induce the expression of transforming growth factor-β2 (TGF-β2), which interferes with another critical pathological process of psoriasis—angiogenesis—by inhibiting the abnormal proliferation of dermal microvascular endothelial cells.51
Research on the molecular mechanisms of topical application provides insights for clinical positioning. Studies indicate that topically applied MSC-Exos localize primarily to the stratum corneum of the skin and that the surface marker CD59 effectively inhibits the assembly of the terminal complement complex C5b-9. Moreover, C5b-9 is a key trigger for inducing neutrophils to undergo neutrophil extracellular traps (NETosis) and release IL-17. Topical MSC-Exos significantly reduced IL-17 and C5b-9 levels in the skin. Therefore, MSC-Exos exert anti-inflammatory effects by inhibiting complement activation, reducing NETosis, and lowering IL-17 levels, which is particularly suitable for mild psoriatic phenotypes.52 This mechanism also partially explains its limited efficacy in deep or severe inflammation—due to insufficient transdermal delivery depth. To repair more extensive cellular damage, ADSC-Exos have been shown to reverse keratinocyte dysfunction induced by serum exosomes (Ps-Exos) from psoriasis patients. The mechanism involves the following mechanisms for restoring multidimensional homeostasis: reinstating autophagy function to clear damaged organelles; alleviating oxidative stress by downregulating NADPH oxidase 2/4 and upregulating nuclear factor erythroid 2-related factor 2 (NRF2); and simultaneously inhibiting the NF-κB and p38 MAPK signaling pathways activated by Ps-Exo.53
Encouragingly, clinical translation has achieved positive progress. In terms of safety, a Phase I open-label trial confirmed that topical MSC exosome ointment was well tolerated, with no treatment-related adverse events or abnormal laboratory indicators observed.54 In terms of efficacy, a randomized, double-blind, controlled study in 12 patients with bilateral psoriasis utilized Wharton’s jelly MSC secretome (exosome-enriched) combined with hyaluronic acid in a lyophilized sponge for topical treatment over 30 days. The results revealed an average 33% reduction in the modified PASI (mPASI) score, a 41% decrease in the target plaque area, and improvements in skin barrier function (transepidermal water loss) and elasticity.55 A dose‑finding study in 12 patients with mild‑to‑moderate plaque psoriasis (divided into three groups receiving single intradermal injections of 50, 100, or 200 μg/cm² autologous ADSC‑exosomes) identified 200 μg/cm² as the optimal regimen, which reduced lesional IL‑17 and CD3 expression and increased FOXP3, with no severe adverse events.56 A systematic review of multiple clinical studies (totaling >30 patients with moderate‑to‑severe psoriasis) revealed that ADSCs and their exosomes exert therapeutic effects through multiple pathways, including secretion of IL-10, prostaglandin E2 (PGE2), and other molecules, as well as direct regulation of keratinocyte autophagy and oxidative stress; clinical improvements included significant PASI reductions (eg, from 24.0 to 8.3, or complete resolution of plaques) with no serious adverse events reported.57
Plant-Derived Nanovesicles
Plant-derived exosome-like nanovesicles (PDNVs) are emerging as a highly promising direction in the field of psoriasis treatment because of their unique advantages of wide availability, low cost, and rich natural bioactive components. These natural nanostructures derived from plants demonstrate therapeutic potential comparable to or even superior to that of some traditional drugs by regulating key inflammatory pathways and oxidative stress responses in psoriasis, and they have garnered significant attention for their excellent biocompatibility.
Various plant-derived PDNVs have been demonstrated to be effective through different molecular mechanisms. For example, PDNVs extracted from garlic and shallot significantly alleviated psoriasis symptoms. The mechanism involves dual synergistic effects: inhibition of IL‑17 signaling (downregulating downstream proinflammatory cytokines such as TNF‑α) and activation of the NRF2 antioxidant pathway (inducing protective proteins such as HMOX1). This dual regulatory effect makes their therapeutic efficacy comparable to that of topical glucocorticoids while avoiding the associated side effects.58
Another groundbreaking study revealed the astonishing cross-kingdom regulatory capabilities between plants and mammals. Extracellular vesicle-like particles (PLEVPs) derived from Perilla frutescens leaves carry the unique plant microRNA Pab-miR-396a-5p. When delivered to psoriatic skin lesions, Pab‑miR‑396a‑5p crosses species barriers, is taken up by keratinocytes, and directly targets heat shock protein 90 (HSP90), inhibiting its expression. This suppresses the NF‑κB and JAK‑STAT pathways, blocks IL‑17 signaling, and reduces epidermal thickness and PASI scores, with efficacy even surpassing that of topical tacrolimus.59
Furthermore, rose hip-derived nanoparticles (RNPs) provide another mechanism to directly combat oxidative stress in psoriasis. Unlike those that activate the endogenous NRF2 pathway, RNPs possess a strong ability to scavenge reactive oxygen species (ROS) directly. After entering keratinocytes through clathrin-mediated endocytosis and other pathways, RNPs can directly eliminate excess ROS, alleviating oxidative damage. Simultaneously, they inhibit the phosphorylation of Akt, thereby interfering with the cell cycle and suppressing the abnormal proliferation of keratinocytes.60
Probiotic/Microbial-Derived Vesicles
With a focus on the “gut‒skin axis” and the regulation of local skin microecology, the use of probiotics and symbiotic microbe-derived vesicles provides a novel approach for modulating systemic and local immunity in psoriasis. They circumvent the potential colonization and infection risks associated with live bacterial therapies by carrying specific bacteria-derived active metabolites, precisely regulating the host immune response and microenvironment homeostasis. In a study by Xie et al61 using an IMQ-induced psoriasis mouse model (n=5/group), oral administration of Lactobacillus plantarum-derived cytoplasmic membrane vesicles (CMVs) reduced PASI scores by approximately 73–78% and decreased epidermal thickness by 56% compared to the IMQ group. The therapeutic effect was attributed to the enrichment of the endogenous cannabinoid ligand anandamide (AEA) within CMVs. Despite these promising preclinical results, the clinical translation of L. plantarum-derived CMVs requires further investigation into optimal dosing, long-term safety, and scalable production.
The outer membrane vesicles of the gut commensal bacterium Parabacteroides goldsteinii (Pg OMVs) exert systemic regulatory effects through their ability to carry the active lipid pentadecanoic acid (PEA). Pg OMVs can be administered via two routes, oral (acting on the gut) or local subcutaneous injection, both of which effectively inhibit psoriasis-like skin inflammation, reducing PASI scores and epidermal thickness. At the molecular level, PEA effectively inhibits the activation of NF-κB and STAT3, two core inflammatory transcription factors in psoriasis, thereby downregulating the expression of downstream cytokines such as IL-17 and IL-23 and restoring the Th17/Treg balance.62 Multiomics analysis revealed its dual regulatory mechanisms: direct action on the skin to inhibit the local mTOR/NF-κB pathway; oral administration also reshaped the disordered gut microbiota, increased the abundance of beneficial Bacteroidetes, and alleviated systemic inflammation.63 Despite these preclinical efficacy data, clinical translation of Pg OMVs requires further evaluation of long-term safety and optimal dosing. These exosomes, which have diverse origins and mechanisms, significantly expand the diversity and precision of psoriasis treatment strategies. To clearly outline the main strategies and molecular mechanisms of therapeutic exosomes, key points are summarized in Table 1.
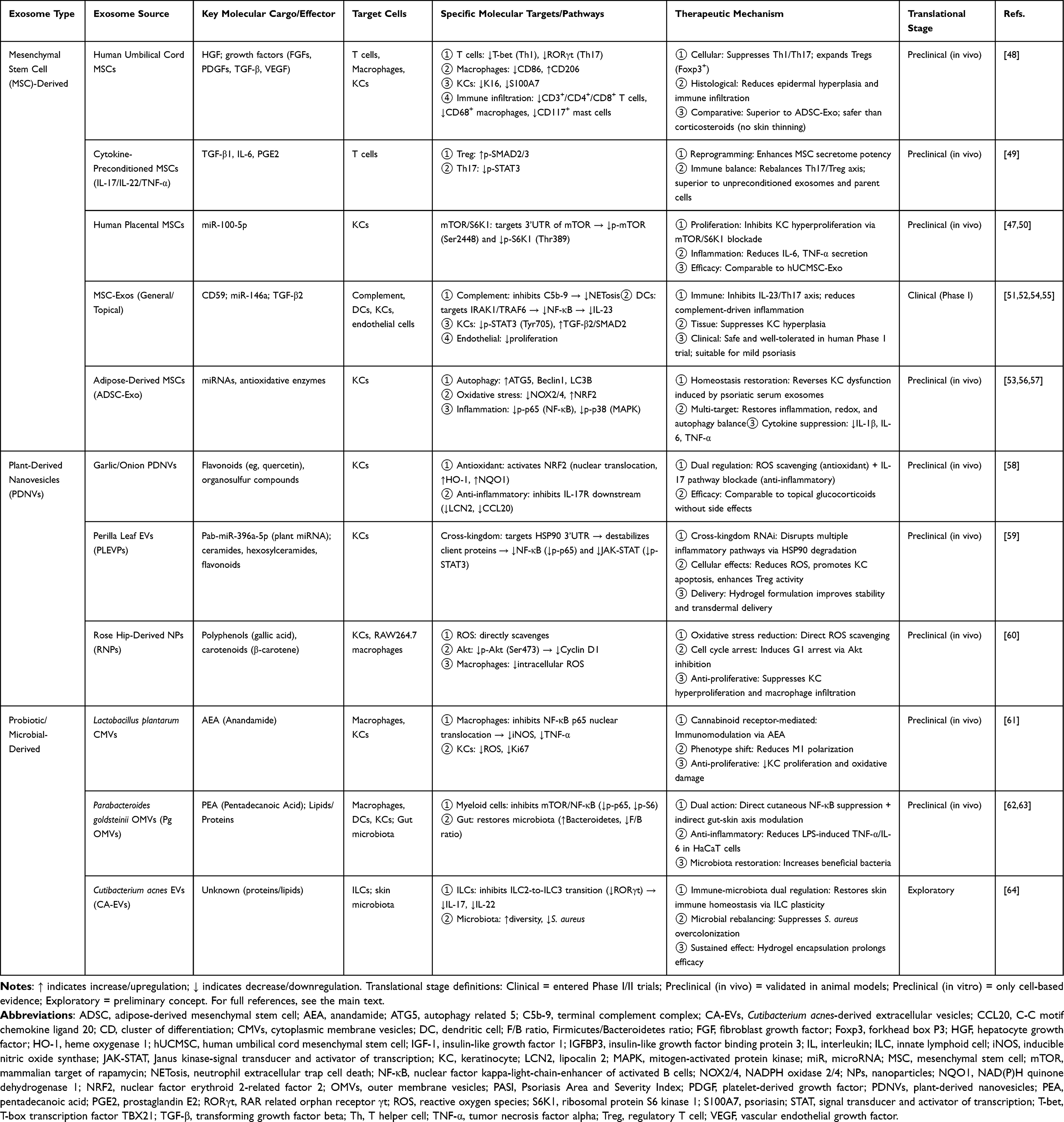 |
Table 1 Therapeutic Strategies of Diverse Exosomes in Psoriasis |
Engineering and Delivery Strategies for Exosomes
To overcome the limitations of natural exosomes in targeting, efficacy, and stability, the integration of engineered modifications and innovative delivery technologies is driving their evolution into intelligent nanomedicines that can be designed on demand and precisely regulated.
Engineering Modifications of Exosomes
Engineering modifications aim to endow exosomes with novel functions through rational design, with core strategies including loading therapeutic molecules, modifying membrane properties and engineering parent cells. Loading strategies focus on encapsulating exogenous active substances into exosomes to achieve precise intervention in specific pathological processes. For example, nor@MSC-EVs were constructed by loading the arginase 1 (Arg1) inhibitor nor-NOHA into MSC-derived vesicles and effectively inhibited Arg1 activity and reduced polyamine production upon uptake by keratinocytes, thereby blocking the key step of protein citrullination in autoantigen formation and subsequently suppressing downstream inflammatory pathways such as NF-κB activation.65 Its efficacy in animal models even surpasses that of clinical anti-interleukin-17A antibodies. Similar loading strategies have been applied to improve existing drugs, such as TFC-Exos, which are formed by loading the JAK inhibitor tofacitinib (TFC) into keratinocyte-derived exosomes via probe sonication.66 This system leverages the natural skin affinity of exosomes to significantly increase drug accumulation and cellular uptake in the skin layer. By more efficiently inhibiting the JAK-STAT pathway, it achieves robust downregulation of inflammatory factors such as TNF-α and IL-23, while its sustained release reduces its cytotoxicity. However, a limitation of this study is the use of exosomes derived from A-431 cancer cells, which may carry potential safety risks for clinical translation. Additionally, exosomes loaded with naturally active ingredients such as green tea polyphenol epigallocatechin-3-gallate nanoparticles (EGN-Exos) can synergistically downregulate NF-κB inflammatory signaling and modulate apoptosis balance (reducing Bax and increasing Bcl-2), demonstrating multitarget synergistic advantages.67
Membrane modification and fusion strategies endow exosomes with new capabilities, such as active targeting by altering their surface properties or integrating other membrane structures. Inspired by tumor immune evasion mechanisms, melanoma-derived exosomes (B16-Exos) were found to possess unique transdermal capabilities (dependent on transcytosis mediated by the membrane protein RhoA) and immunosuppressive cargo (mmu-miR-320-3p targeting IL-17a mRNA). The biomimetic formulation ExoLipo, which uses B16-Exo as a template, incorporates a RhoA membrane protein and is loaded with therapeutic miRNA, not only inheriting the dual functions of natural exosomes—transdermal delivery and immune regulation—but also effectively avoiding the potential safety risks associated with the direct use of tumor-derived exosomes.68 Another approach involves membrane fusion technology, where T-cell exosomes overexpressing the anti-inflammatory protein annexin A1 (ANXA1) are fused with M2 macrophage membranes, enabling the resulting engineered vesicles to simultaneously possess the direct anti-inflammatory activity of ANXA1 and the inherent natural homing-targeting ability of M2 cell membranes. In an IMQ-induced psoriasis mouse model, subcutaneous injection of these engineered EVs enhances the polarization of local macrophages toward the M2 anti-inflammatory phenotype, downregulating M1 markers such as CD86 and IL-6, thereby significantly alleviating skin inflammation in psoriasis animal models and demonstrating superior stability compared with nonfused vesicles.69 A more innovative strategy is cross-species fusion, such as fusing grapefruit exosomes (GEVs) with mesenchymal stem cell membrane nanovesicles (CCR6-NVs) that overexpress the chemokine receptor CCR6 on their surface. The resulting intelligent hybrid vesicles can actively target inflammatory sites via the CCR6‒CCL20 axis and synergistically load the cell cycle inhibitor CX5461 while retaining the plant-derived miR-159a with regulatory functions. This enables cross-kingdom suppression of inflammatory factors such as IL-6 and TNF-α, promotes regulatory T-cell infiltration, and polarizes macrophages toward the M2 anti-inflammatory phenotype, achieving multitarget synergistic therapy.70 The use of Cutibacterium acnes-derived extracellular vesicles encapsulated in sustained-release hydrogel microspheres (CA-EVs@GHMs) represents another functionalized delivery system. They enable sustained vesicle release, inhibiting the conversion of skin innate lymphoid cells (ILCs) into pathogenic ILC3 subtypes and restoring skin microbiota diversity, thereby suppressing the overcolonization of harmful bacteria such as Staphylococcus aureus. This system exerts a dual-target “immune microbiota” regulatory effect.64
The parental cell engineering strategy achieves precise intervention in diseases by accurately manipulating the source cells that secrete exosomes, thereby directionally regulating the cargo composition and function of exosomes. The most representative method involves genetic engineering to overexpress specific therapeutic molecules in parental cells. For example, lentiviral vectors can be used to induce MSCs to overexpress the immune checkpoint protein programmed death ligand 1 (PD-L1). The engineered small extracellular vesicles (MSC-sEVs-PD-L1) secreted by these cells consequently exhibit high PD-L1 expression on their surfaces. These vesicles can target psoriatic inflammatory sites and, by binding to programmed death receptor 1 (PD-1) on activated T cells, efficiently activate the PD-1/PD-L1 inhibitory signaling pathway. This leads to more effective suppression of pathogenic Th1 and Th17 cell proliferation, promotes the differentiation of Treg cells, and inhibits dendritic cell maturation. The in vivo therapeutic efficacy of these vesicles even surpasses that of PD-L1-overexpressing MSCs themselves while avoiding the risk of pulmonary embolism associated with cell infusion.71 Additionally, physical or chemical pretreatment of parental cells serves as an efficient nongenetic modification approach. Studies have shown that exposing keratinocytes to UVB irradiation can “reprogram” their secreted exosomes (UV KEV) into potent immunomodulatory carriers. The mechanism involves UVB-induced enrichment of platelet-activating factor (PAF) in exosomes, which, via its receptor pathway, remotely promotes mast cell migration to lymph nodes and the secretion of IL-10. This systemically expands the Treg cell population and decreases mature dendritic cell numbers, inducing systemic immune tolerance.72 A codelivery system (J@EV) was developed by loading UV-KEV with the metabolic inhibitor JPH203. This system synergistically combines the anti-inflammatory effects of the IL-1RA/NF-κB axis with the antimetabolic proliferation effects of the JPH203/mTORC1 axis.73 Another parental cell strategy leverages the natural “organelle transfer” capability of platelet-derived extracellular vesicles (PEVs). PEVs can directly transfer their intact, functional mitochondria to macrophages, fundamentally reprogramming the latter’s energy metabolism from proinflammatory glycolysis to anti-inflammatory, reparative oxidative phosphorylation.74 This profound metabolic reprogramming is a key mechanism driving macrophage polarization toward the M2 anti-inflammatory phenotype. In summary, through genetic modification, physical stimulation, or the utilization of inherent properties of parental cells, engineered exosomes carrying specific therapeutic cargos can be directionally produced, significantly enhancing the precision of exosome-based strategies for psoriasis treatment.
Exosome Local Delivery System
Efficient local delivery is the key to the therapeutic efficacy of exosome therapy. Microneedle-based delivery systems overcome the skin barrier by physically creating micron-sized channels, providing a minimally invasive and efficient solution. The basic microneedle platform can serve as a carrier for codelivery, such as loading Treg cell-derived exosomes (rExos) with the anti-inflammatory drug dimethyl fumarate (DMF) into hyaluronic acid microneedle patches (rExo@DMF MNs). Among these, rExos can induce the generation of tolerogenic dendritic cells and Tregs, whereas DMF enhances anti-inflammatory signaling and activates the NRF2 antioxidant pathway. The two synergistically inhibit NF-κB, thereby locally remodeling the immune microenvironment.75 Another study addressed the clinical translation bottleneck of storage stability for exosome formulations. Researchers have developed microneedles composed of hyaluronic acid and the protective agent trehalose for delivering natural killer cell-derived extracellular vesicles (NK-EVs). These microneedles not only effectively treated psoriasis-like skin lesions but also demonstrated a prominent advantage: the addition of trehalose enabled the loaded vesicles to retain their anti-inflammatory activity after room temperature storage for up to 6 months, offering a highly promising solution for the long-term storage and transportation of exosome formulations.76
Intelligent responsive microneedle systems represent a more advanced direction, achieving integrated diagnosis and treatment through the integration of environmentally responsive components. For example, a dual-segment subcutaneous light-responsive enhanced microneedle (SLE MN) system features an upper section composed of polymethyl methacrylate optical fibers, which efficiently transmit near-infrared light irradiated on the surface to the epidermal basal layer. The lower drug-loaded matrix encapsulates M2 macrophage-derived exosomes and the photothermal conversion material MXene. Under near-infrared light irradiation, MXene generates localized mild photothermal effects, directly inducing apoptosis in hyperproliferative basal layer keratinocytes. Simultaneously, the photothermal trigger enables the rapid release of exosomes, thereby regulating the polarization of local macrophages toward the M2 anti-inflammatory phenotype.77
In summary, through the deep integration of diverse engineering modification strategies and advanced delivery systems, exosomes have evolved from natural messengers to programmable, targetable, and controllable intelligent therapeutic platforms, opening a promising new pathway for the precise treatment of psoriasis. The main approaches and characteristics are summarized in Table 2.
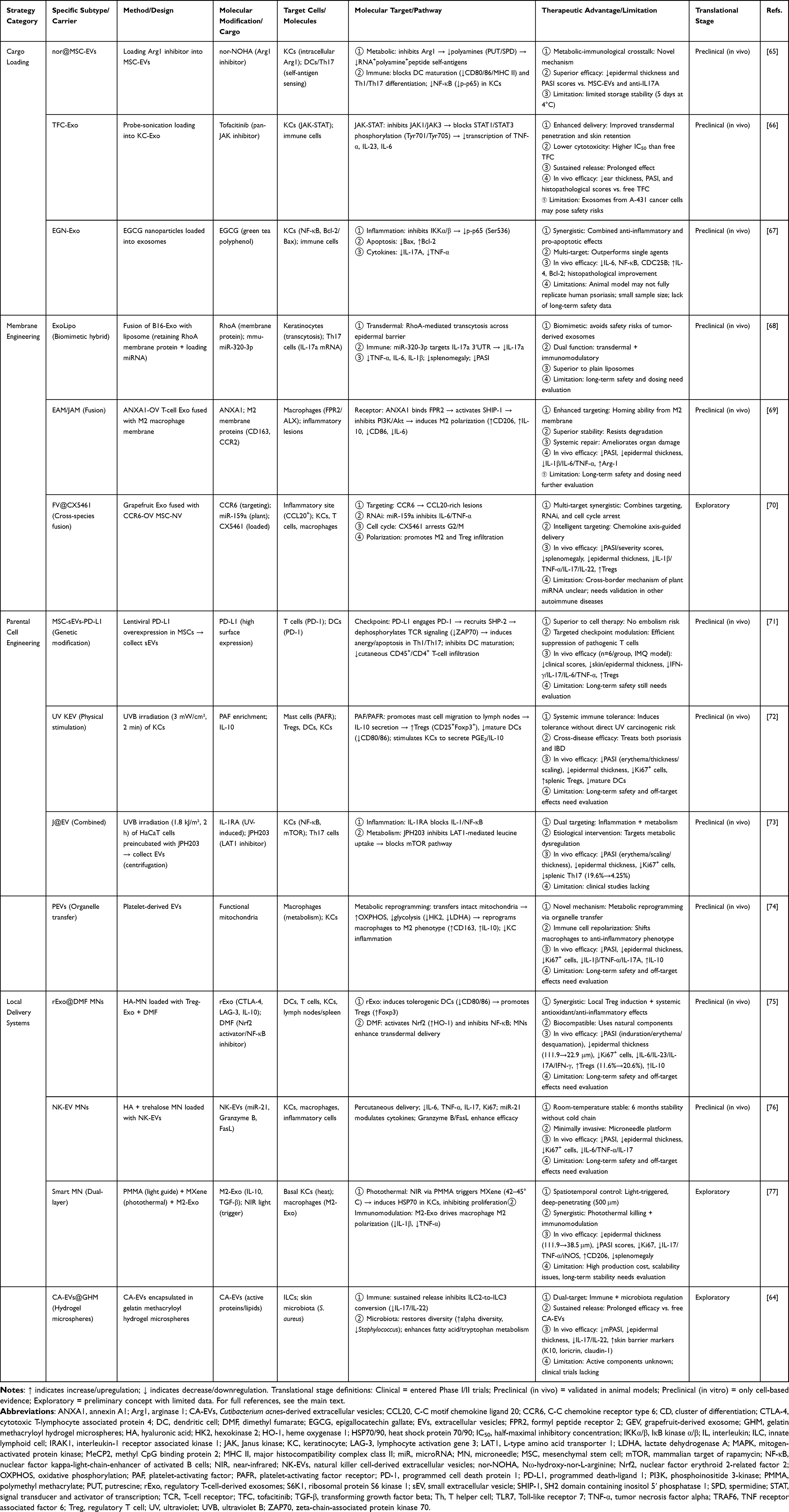 |
Table 2 Engineering and Delivery Strategies for Exosome-Based Psoriasis Therapy |
Challenges and Future Prospects
Although exosomes have demonstrated disruptive potential in mechanistic research and therapeutic exploration of psoriasis, their translation from basic discoveries to clinical routines still faces a series of fundamental scientific and translational challenges. The essence of these obstacles largely stems from the inherent complexity of exosomes as endogenous nanoscale vesicles and the vast gap between this complexity and the rigor required to reshape them into standardized “pharmaceutical products”.
The core challenge stems from the dual dilemma of standardization and mechanistic understanding. The primary bottleneck is the lack of standardization in production processes and quality control. Currently, there is no global consensus on isolation and purification methods for obtaining exosomes from different cell sources, leading to significant variations in product purity, subgroup heterogeneity, and biological activity. Large-scale, high-purity, low-cost manufacturing processes compliant with good manufacturing practices are lacking, and their industrial pathways, along with strict quality control systems encompassing attributes such as potency, purity, and sterility, urgently need to be established, which directly hinders their clinical translation. A deeper challenge lies in the “black box of effector components” for therapeutic exosomes. Although a few studies have identified active molecules such as miR-100-5p, AEA, and PEA, for the vast majority of exosomes, it remains unclear whether their therapeutic effects are attributable to a single star molecule, specific molecular combinations, or the synergistic effects of the vesicle as a whole. This mechanistic ambiguity hinders rational process design on the basis of quality attributes, precise potency evaluation, and targeted engineering modifications.
The depth and breadth of clinical translation evidence have significant limitations. Current human trials are generally small in scale and short in duration and lack reliable long-term safety and sustained efficacy data. Study designs predominantly use open-label or single-arm formats, and there is an urgent need for randomized, double-blind, head‒to-head superiority comparisons with existing standard therapies to clarify their therapeutic position. The optimal dosing regimen (formulation, dose, and course) has not undergone systematic pharmacokinetic/pharmacodynamic optimization. Additionally, the potential immunogenicity, off-target effects, and long-term biodistribution risks of allogeneic or engineered exosomes must be evaluated through more rigorous preclinical and clinical monitoring.
Future breakthroughs rely on technological innovation and paradigm shifts. Advancing the field requires focusing on several key directions: 1) Establishing standards and regulatory consensus: Through international collaboration, developing guiding principles for critical quality attributes of therapeutic exosomes, standardized testing methods, and regulatory pathways. 2) Deepening mechanism analysis and rational design: Single-vesicle analysis, spatial omics, and gene editing technologies can be utilized to clarify the causal contributions of functional components at the donor cell level, and predictable, modular engineering platforms can be developed to achieve “on-demand customization”. 3) Innovative production paradigms: Exploring cell-free synthetic biology or robust microbial expression systems to achieve truly standardized and scalable production. 4) Conduct high-level clinical validation: Design large-scale, multicenter, randomized controlled trials and conduct comparative or combination therapy studies with existing standard treatments (eg, biologics) while systematically establishing pharmacokinetic/pharmacodynamic models. 5) Promotion of integrated diagnosis and treatment: Accelerate the validation of circulating exosome biomarkers; develop companion diagnostic tools for patient stratification, efficacy prediction, and dynamic monitoring; and advance toward personalized precision medicine.
Beyond these technical and regulatory priorities, several biologically intriguing questions merit future investigation. Although genetic association studies have linked IL19/IL20/IL24 polymorphisms to psoriasis susceptibility,78,79 whether exosomal cargo of these cytokines contributes to subtype‑specific diagnosis (eg, palmoplantar pustulosis vs. psoriasis vulgaris) remains unknown. Transcriptomic analyses have revealed differential expression of repetitive elements (REs), such as LINE‑1, Alu, and HERV‑K, in psoriatic skin,80 but whether these REs are selectively packaged into exosomes and contribute to disease pathogenesis is an open question. Likewise, exosomes can carry functional apoptosis‑inducing factors, as shown by placental exosomes expressing oligomerized FasL and TRAIL.81 In psoriasis, TRAIL and its death receptors DR4/DR5 are upregulated in lesional skin,82 raising the possibility that psoriatic exosomes may similarly transport these molecules and modulate immune cell survival or keratinocyte apoptosis. Addressing these questions will help bridge the gap between basic exosome biology and clinical application.
Only through deep interdisciplinary integration and systematic resolution of these scientific challenges—from molecular mechanisms to industrial implementation—can exosomes transform from a broad research blueprint into tangible clinical breakthroughs benefiting psoriasis patients.
Conclusion
Exosomes play dual roles in psoriasis as both core pathogenic mediators and versatile translational tools. Mechanistically, diseased cells release exosomes that carry not only non-coding RNAs but also proteins, lipids, and metabolites, which collectively program CD4+ T cells toward Th17 polarization, drive M1 macrophage activation, and establish a positive inflammatory feedback loop with neutrophils, thereby amplifying the IL‑23/Th17 axis. Therapeutically, this natural delivery system has been successfully repurposed: circulating exosomal fingerprints enable non‑invasive diagnosis and subtype differentiation, while exosomes derived from mesenchymal stem cells, plants, and microorganisms offer cell‑free, multi‑mechanistic synergy (anti‑inflammatory, immunoregulatory, antioxidant, and microecological repair). Engineering modifications and intelligent delivery platforms, particularly microneedle‑based systems, are further advancing exosomes into targeted, smart nanomedicines. What is most promising today is the clinical translation of mesenchymal stem cell‑derived exosomes, which have shown safety and preliminary efficacy in early‑stage trials, and microneedle‑mediated local delivery, which overcomes skin barriers and enhances retention. The key remaining barriers to clinical adoption are standardized good manufacturing practice production, identification of core effector components, and rigorous Phase II/III validation against current standards (eg, biologics). Overcoming these challenges will transform exosome‑based strategies into precision tools for psoriasis management.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Boehncke WH, Schön MP. Psoriasis. Lancet. 2015;386(9997):983–22. doi:10.1016/S0140-6736(14)61909-7
2. Pan Y, Zou J, Hu T, et al. Psoriasis: a multidimensional review of onset, progression, treatment, and the evolution of disease models. Mol Diagn Ther. 2025;29(3):345–366. doi:10.1007/s40291-025-00776-8
3. Armstrong AW, Read C. Pathophysiology, clinical presentation, and treatment of psoriasis: a review. JAMA. 2020;323(19):1945–1960. doi:10.1001/jama.2020.4006
4. Lee HJ, Kim M. Challenges and future trends in the treatment of psoriasis. Int J Mol Sci. 2023;24(17):13313.
5. Prema SS, Shanmugamprema D. Systemic psoriasis: from molecular mechanisms to global management strategies. Clin Rev Allergy Immunol. 2025;68(1):79. doi:10.1007/s12016-025-09089-4
6. Blauvelt A, Chiricozzi A. The immunologic role of IL-17 in psoriasis and psoriatic arthritis pathogenesis. Clin Rev Allergy Immunol. 2018;55(3):379–390. doi:10.1007/s12016-018-8702-3
7. Lwin SM, Azrielant S, He J, Griffiths CEM. Curing psoriasis. J Investig Dermatol. 2024;144(12):2645–2649. doi:10.1016/j.jid.2024.09.012
8. Yu H, Feng H, Zeng H, et al. Exosomes: the emerging mechanisms and potential clinical applications in dermatology. Int J Biol Sci. 2024;20(5):1778–1795. doi:10.7150/ijbs.92897
9. Raposo G, Stoorvogel W. Extracellular vesicles: exosomes, microvesicles, and friends. J Cell Biol. 2013;200(4):373–383. doi:10.1083/jcb.201211138
10. Elliott RO, He M. Unlocking the power of exosomes for crossing biological barriers in drug delivery. Pharmaceutics. 2021;13(1):122. doi:10.3390/pharmaceutics13010122
11. Bin Dayel S, Hussein RS. Exosomes in dermatology: emerging roles in skin health and disease. Pharmaceutics. 2025;17(5):600. doi:10.3390/pharmaceutics17050600
12. Sawamura S, Tselmeg Mijiddorj M, Kajihara I, et al. Clinical significance of circulating exosomal interleukin-23 and tumour necrosis factor-α messenger RNA in psoriasis. J Eur Acad Dermatol Venereol. 2023;37(6):e815–e817. doi:10.1111/jdv.18945
13. Jacquin-Porretaz C, Cordonnier M, Nardin C, et al. Increased levels of interleukin-17A exosomes in psoriasis. Acta Dermato-Venereologica. 2019;99(12):1143–1147. doi:10.2340/00015555-3300
14. Tang B, Bi Y, Zheng X, et al. The role of extracellular vesicles in the development and treatment of psoriasis: narrative review. Pharmaceutics. 2024;16(12):1586. doi:10.3390/pharmaceutics16121586
15. Nahm WJ, Nikas C, Goldust M, et al. Exosomes in dermatology: a comprehensive review of current applications, clinical evidence, and future directions. Int J Dermatol. 2025;64(11):1995–2010. doi:10.1111/ijd.17903
16. Abdal Dayem A, Song M, Park J, et al. The therapeutic potential of extracellular vesicles in psoriasis treatment: mechanisms, applications, and prospects. Int J Mol Sci. 2025;26(21):10297. doi:10.3390/ijms262110297
17. Li W, Li H, Fan L, et al. The potential application of exosomes as therapeutic agents, carriers, and biomarkers in skin diseases. Int J Nanomed. 2025;20:12627–12658. doi:10.2147/IJN.S547871
18. Chen Y, Liu H, He Y, Yang B, Lu W, Dai Z. Roles for exosomes in the pathogenesis, drug delivery and therapy of psoriasis. Pharmaceutics. 2025;17(1):51. doi:10.3390/pharmaceutics17010051
19. Mangino G, Iuliano M, Carlomagno S, et al. Interleukin-17A affects extracellular vesicles release and cargo in human keratinocytes. Exp Dermatol. 2019;28(9):1066–1073. doi:10.1111/exd.14015
20. Yuan Z, Zeng X, Zhang X, Xia C, Peng X. Exosomes containing long non-coding RNA AGAP2-AS1 promote the differentiation of CD4(+) T cells through the miR-424-5p/SGK1 axis in psoriasis. Front Genet. 2025;16:1521470. doi:10.3389/fgene.2025.1521470
21. Gao P, Gao X, Lin L, et al. Identification of PRKCQ-AS1 as a keratinocyte-derived exosomal lncRNA that promotes Th17 differentiation and IL-17 secretion in psoriasis through bioinformatics, machine learning algorithms, and cell experiments. J Inflamm Res. 2025;18:6557–6582. doi:10.2147/JIR.S521553
22. Jiang M, Fang H, Dang E, et al. Small extracellular vesicles containing miR-381-3p from keratinocytes promote T helper type 1 and T helper type 17 polarization in psoriasis. J Investig Dermatol. 2021;141(3):563–574. doi:10.1016/j.jid.2020.07.009
23. Kim HR, Lee SY, You GE, Park CW, Kim HO, Chung BY. Exosomes released by environmental pollutant-stimulated Keratinocytes/PBMCs can trigger psoriatic inflammation in recipient cells via the AhR signaling pathway. Front Mol Biosci. 2023;10:1324692. doi:10.3389/fmolb.2023.1324692
24. Lin J, Cao Y, Ma L, Tao M, Yang X. Keratinocyte exosomal LOC285194 ameliorates psoriasis by inhibiting the differentiation of CD4(+)T cells to Th17 cells through regulating miR-211-5p/SIRT1 axis. IUBMB Life. 2025;77(1):e2935. doi:10.1002/iub.2935
25. Wang Z, Luo F. Integrated analysis of exosome-related genes and their role in psoriasis pathogenesis. Front Immunol. 2025;16:1492012. doi:10.3389/fimmu.2025.1492012
26. Jiang W, Zhang T, Qiu Y, et al. Keratinocyte-to-macrophage communication exacerbate psoriasiform dermatitis via LRG1-enriched extracellular vesicles. Theranostics. 2024;14(3):1049–1064. doi:10.7150/thno.89180
27. Sun W, Chen J, Li J, et al. Vitamin D receptor-deficient keratinocytes-derived exosomal miR-4505 promotes the macrophage polarization towards the M1 phenotype. PeerJ. 2023;11:e15798. doi:10.7717/peerj.15798
28. Salvi V, Gaudenzi C, Mariotti B, et al. Cell damage shifts the microRNA content of small extracellular vesicles into a Toll-like receptor 7-activating cargo capable to propagate inflammation and immunity. Cell Commun Signaling. 2024;22(1):536. doi:10.1186/s12964-024-01924-z
29. Jiang M, Fang H, Shao S, et al. Keratinocyte exosomes activate neutrophils and enhance skin inflammation in psoriasis. FASEB J. 2019;33(12):13241–13253. doi:10.1096/fj.201900642R
30. Shao S, Fang H, Zhang J, et al. Neutrophil exosomes enhance the skin autoinflammation in generalized pustular psoriasis via activating keratinocytes. FASEB J. 2019;33(6):6813–6828. doi:10.1096/fj.201802090RR
31. Margouta A, Anyfanti P, Lazaridis A, et al. Assessment of microvesicles from different cell origins in patients with psoriasis: evidence of thrombogenic, proinflammatory microenvironment in the absence of established cardiovascular disease. J Human Hypertens. 2023;37(10):925–930. doi:10.1038/s41371-022-00787-6
32. Li S, Chik Z, Faruqu FN, et al. miRNAs and exosomes in psoriasis: coordinating cytoskeleton dynamics and extracellular matrix remodeling. Front Cell Develop Biol. 2025;13:1608902. doi:10.3389/fcell.2025.1608902
33. Zhang B, Wu F. Proteomic identification of exosomes derived from psoriasis cells using data-independent acquisition mass spectrometry. Arch Dermatolog Res. 2024;316(6):224. doi:10.1007/s00403-024-02984-4
34. Wang S, Han Q, Wang J, Peng X, Zhou L, Niu X. Proteomic analysis of mesenchymal stem cell-derived exosomes in psoriasis. Current Mol Med. 2025;25(10):1282–1289. doi:10.2174/0115665240304551241017050628
35. Dashti M, Mohammadi M, Dehnavi S, Sadeghi M. Immunomodulatory interactions between mesenchymal stromal/stem cells and immune cells in psoriasis: therapeutic potential and challenges. Stem Cell Res Ther. 2025;16(1):241. doi:10.1186/s13287-025-04375-6
36. Iuliano M, Grimaldi L, Rosa P, et al. Extracellular vescicles in psoriasis: from pathogenesis to possible roles in therapy. Front Immunol. 2024;15:1360618. doi:10.3389/fimmu.2024.1360618
37. Crescitelli R, Lässer C, Lötvall J. Isolation and characterization of extracellular vesicle subpopulations from tissues. Nat Prot. 2021;16(3):1548–1580. doi:10.1038/s41596-020-00466-1
38. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13(2):e12404. doi:10.1002/jev2.12404
39. Zhang Y, Lan M, Chen Y. Minimal Information for studies of extracellular vesicles (MISEV): ten-year evolution (2014-2023). Pharmaceutics. 2024;16(11):1394. doi:10.3390/pharmaceutics16111394
40. Park YJ, Kim DC, Lee SJ, et al. Keratinocyte-derived circulating microRNAs in extracellular vesicles: a novel biomarker of psoriasis severity and potential therapeutic target. J Transl Med. 2024;22(1):235. doi:10.1186/s12967-024-05030-z
41. Guzmán-Martín CA, Jiménez-Ortega RF, Ortega-Springall MF, et al. miR-16-5p, miR-21-5p, and miR-155-5p in circulating vesicles as psoriasis biomarkers. Sci Rep. 2025;15(1):6971. doi:10.1038/s41598-025-91532-9
42. Wang R, Huang Y, Shao K, Yan J, Sun Q. High expression of miR-6785-5p in the serum exosomes of psoriasis patients alleviates psoriasis-like skin damage by interfering with the MNK2/p-eIF4E axis in keratinocytes. Inflammation. 2024;47(5):1585–1599. doi:10.1007/s10753-024-01995-7
43. Yan K, Zhu J, Zhang M, et al. Differential expression of plasma extracellular vesicle miRNAs as biomarkers for distinguishing psoriatic arthritis from psoriasis. Chin Med J. 2025;138(2):219–221. doi:10.1097/CM9.0000000000003288
44. Lättekivi F, Guljavina I, Midekessa G, et al. Profiling blood serum extracellular vesicles in plaque psoriasis and psoriatic arthritis patients reveals potential disease biomarkers. Int J Mol Sci. 2022;23(7):4005. doi:10.3390/ijms23074005
45. Born LJ, Khachemoune A. Extracellular vesicles: a comprehensive review of their roles as biomarkers and potential therapeutics in psoriasis and psoriatic arthritis. Clin Exp Dermatol. 2023;48(4):310–318. doi:10.1093/ced/llac108
46. Pasquali L, Svedbom A, Srivastava A, et al. Circulating microRNAs in extracellular vesicles as potential biomarkers for psoriatic arthritis in patients with psoriasis. J Eur Acad Dermatol Venereol. 2020;34(6):1248–1256. doi:10.1111/jdv.16203
47. Huang YC, Chang CY, Huang CJ. Effectiveness of exosomes from different mesenchymal stem cells in the treatment of psoriasis: a murine study and meta-analysis of experimental studies. Biomedicines. 2025;13(9):2093. doi:10.3390/biomedicines13092093
48. Siriphanit R, Thanasarnaksorn W, Boonpethkaew S, et al. Exosome-based therapy: a comparative study of adipose- and umbilical cord-derived mesenchymal stem cells-derived exosomes in psoriatic mouse model. Exp Dermatol. 2025;34(11):e70176. doi:10.1111/exd.70176
49. Dairov A, Issabekova A, Sarsenova M, et al. Study of the therapeutic effect of cytokine-preconditioned mesenchymal stem cells and their exosomes in a mouse model of psoriasis. Biology. 2025;14(8):1033. doi:10.3390/biology14081033
50. Huang YC, Chang CY, Huang CJ. MicroRNA-100-5p in exosomes from human placental mesenchymal stem cells: a key modulator of anti-psoriatic effects through mTOR/S6 K1 pathway. J Mol Med. 2025;103(8):951–961. doi:10.1007/s00109-025-02560-x
51. Abed ZI, Arianejad M, Azizi Z. Mesenchymal stem cell-derived exosomes decrease Hyperplasia in Psoriasis by inducing transforming growth factor β2 (TGF-β2). Mol Biol Rep. 2024;51(1):635. doi:10.1007/s11033-024-09337-4
52. Zhang B, Lai RC, Sim WK, Choo ABH, Lane EB, Lim SK. Topical application of mesenchymal stem cell exosomes alleviates the imiquimod induced psoriasis-like inflammation. Int J Mol Sci. 2021;22(2):720.
53. Kim HR, Lee SY, You GE, Kim HO, Park CW, Chung BY. Adipose-derived stem cell exosomes alleviate psoriasis serum exosomes-induced inflammation by regulating autophagy and redox status in keratinocytes. Clin Cosmetic Investig Dermatol. 2023;16:3699–3711. doi:10.2147/CCID.S439760
54. Chandran NS, Bhupendrabhai MN, Tan TT, et al. A phase 1, open-label study to determine safety and tolerability of the topical application of mesenchymal stem/stromal cell (MSC) exosome ointment to treat psoriasis in healthy volunteers. Cytotherapy. 2025;27(5):633–641. doi:10.1016/j.jcyt.2025.01.007
55. Elgueta E, Prieto CP, Hartmann D, et al. Novel sponge formulation of mesenchymal stem cell secretome and hyaluronic acid: a safe and effective topical therapy for Psoriasis vulgaris. Stem Cell Res Ther. 2025;16(1):348. doi:10.1186/s13287-025-04415-1
56. Mohseni Meybodi MA, Nilforoushzadeh MA, KhandanDezfully N, Mansouri P. The safety and efficacy of adipose tissue-derived exosomes in treating mild to moderate plaque psoriasis: a clinical study. Life Sci. 2024;353:122915. doi:10.1016/j.lfs.2024.122915
57. Matwiejuk M, Mikłosz A, Myśliwiec H, Chabowski A, Flisiak I. Adipose-derived mesenchymal stem cells and their derivatives in inflammatory skin diseases: a systematic review. Front Immunol. 2025;16:1617157. doi:10.3389/fimmu.2025.1617157
58. Kalarikkal SP, Kumar MN, Rajendran S, et al. Natural plant-derived nanovesicles for effective psoriasis therapy via dual modulation of IL-17 and NRF2 pathway. iScience. 2025;28(6):112556. doi:10.1016/j.isci.2025.112556
59. Liu Y, Tao S, Zhang Z, et al. Perilla frutescens leaf-derived extracellular vesicle-like particles carry Pab-miR-396a-5p to alleviate psoriasis by modulating IL-17 signaling. Research. 2025;DC(8):0675. doi:10.1126/science.aec7378
60. Hashimoto M, Itakura S, Kusamori K, et al. Therapeutic potential of rose hip-derived nanoparticles for psoriatic skin inflammation. ACS Biomater Sci Eng. 2025;11(10):5938–5951. doi:10.1021/acsbiomaterials.5c00826
61. Xie Y, Lv G, Su D, et al. Lactobacillus plantarum-derived cytoplasmic membrane vesicles as novel anti-inflammatory nanotherapeutics for psoriasis management. Front Immunol. 2025;16:1647466. doi:10.3389/fimmu.2025.1647466
62. Su D, Li M, Xie Y, et al. Gut commensal bacteria Parabacteroides goldsteinii-derived outer membrane vesicles suppress skin inflammation in psoriasis. J Controlled Release. 2025;377:127–145. doi:10.1016/j.jconrel.2024.11.014
63. Du A, Chen J, Xie Z, et al. Multi-omics analysis of outer membrane vesicles from P. goldsteinii in a psoriasis mouse model. Life Sci. 2025;377:123776. doi:10.1016/j.lfs.2025.123776
64. Xu Y, Gan Y, Qi F, et al. Innate lymphoid cell-based immunomodulatory hydrogel microspheres containing Cutibacterium acnes extracellular vesicles for the treatment of psoriasis. Acta Biomater. 2024;184:296–312. doi:10.1016/j.actbio.2024.06.006
65. Zhou X, Tang B, Huang Q, et al. Engineered mesenchymal stem cell-derived extracellular vesicles scavenge self-antigens for psoriasis therapy via modulating metabolic and immunological disorders. Adv Sci. 2025;12(6):e2410067. doi:10.1002/advs.202410067
66. Dehghani P, Varshosaz J, Mirian M, Minaiyan M, Kazemi M, Bodaghi M. Keratinocyte exosomes for topical delivery of tofacitinib in treatment of psoriasis: an in vitro/in vivo Study in Animal Model of Psoriasis. Pharmaceut Res. 2024;41(2):263–279.
67. Kishta MS, Abd-Rabou AA, Sarkissian GK, et al. Exosome mediated delivery of Epigallocatechin 3 gallate as a novel approach to alleviate psoriasis symptoms through cytokine and apoptotic pathway modulation. Sci Rep. 2025;15(1):30013. doi:10.1038/s41598-025-10886-2
68. Yang J, Zhu J, Lu S, Qin H, Zhou W. Transdermal psoriasis treatment inspired by tumor microenvironment-mediated immunomodulation and advanced by exosomal engineering. J Controlled Release. 2025;382:113664. doi:10.1016/j.jconrel.2025.113664
69. Wang Z, Qin Z, Wang J, et al. Engineering extracellular vesicles with macrophage membrane fusion for ameliorating imiquimod-induced psoriatic skin inflammation. J Dermatolog Treatment. 2023;34(1):2220445. doi:10.1080/09546634.2023.2220445
70. Huang R, Jia B, Su D, et al. Plant exosomes fused with engineered mesenchymal stem cell-derived nanovesicles for synergistic therapy of autoimmune skin disorders. J Extracell Vesicles. 2023;12(10):e12361. doi:10.1002/jev2.12361
71. Xu F, Fei Z, Dai H, et al. Mesenchymal stem cell-derived extracellular vesicles with high PD-L1 expression for autoimmune diseases treatment. Adv Mat. 2022;34(1):e2106265. doi:10.1002/adma.202106265
72. Liu L, Yang D, Ji J, et al. Harnessing the immunomodulation of UV-exposed keratinocyte extracellular vesicles for inflammatory disorder treatment. Adv Sci. 2025;12(36):e01517. doi:10.1002/advs.202501517
73. Jiang X, Jiang Z, Huang S, et al. Ultraviolet B radiation-induced JPH203-loaded keratinocyte extracellular vesicles exert etiological interventions for psoriasis therapy. J Controlled Release. 2023;362:468–478. doi:10.1016/j.jconrel.2023.08.059
74. Wang B, Li D, Pan C, et al. Platelet-derived extracellular vesicles alleviate psoriatic inflammation via mitochondrial transfer to macrophages. Exp Dermatol. 2025;34(8):e70152. doi:10.1111/exd.70152
75. Ou M, Cao J, Luo R, et al. Drug-loaded microneedle patches containing regulatory T cell-derived exosomes for psoriasis treatment. Acta Biomater. 2025;198:452–466. doi:10.1016/j.actbio.2025.04.015
76. Lim SA, Chen S, Suzuki I, et al. Harnessing microneedles for delivery and preservation of natural killer cell-derived extracellular vesicles. ACS Biomater Sci Eng. 2025;11(7):4266–4278. doi:10.1021/acsbiomaterials.5c00760
77. Zhao ZQ, Zhang SL, Yu R, et al. Optical microneedle-enhanced transdermal light scattering for in situ photothermal therapy targeting basal-layer psoriasis. ACS Appl Mat Interfaces. 2025;17(13):19446–19458. doi:10.1021/acsami.4c23014
78. Kingo K, Kõks S, Nikopensius T, Silm H, Vasar E. Polymorphisms in the interleukin-20 gene: relationships to plaque-type psoriasis. Genes Immun. 2004;5(2):117–121. doi:10.1038/sj.gene.6364046
79. Kingo K, Mössner R, Kõks S, et al. Association analysis of IL19, IL20 and IL24 genes in palmoplantar pustulosis. Brit J Dermatol. 2007;156(4):646–652. doi:10.1111/j.1365-2133.2006.07731.x
80. Krishnan VS, Kõks S. Transcriptional landscape of repetitive elements in psoriatic skin from large cohort studies: relevance to psoriasis pathophysiology. Int J Mol Sci. 2023;24(23):16725. doi:10.3390/ijms242316725
81. Stenqvist AC, Nagaeva O, Baranov V, Mincheva-Nilsson L. Exosomes secreted by human placenta carry functional Fas ligand and TRAIL molecules and convey apoptosis in activated immune cells, suggesting exosome-mediated immune privilege of the fetus. J Immunol. 2013;191(11):5515–5523. doi:10.4049/jimmunol.1301885
82. Peternel S, Prpić-Massari L, Manestar-Blažić T, Brajac I, Kaštelan M. Increased expression of TRAIL and its death receptors DR4 and DR5 in plaque psoriasis. Arch Dermatolog Res. 2011;303(6):389–397. doi:10.1007/s00403-011-1125-0
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Current Status, Opportunities, and Challenges of Exosomes in Oral Cancer Diagnosis and Treatment
Liu H, Huang Y, Huang M, Huang Z, Wang Q, Qing L, Li L, Xu S, Jia B
International Journal of Nanomedicine 2022, 17:2679-2705
Published Date: 16 June 2022
The Combination of IL-6, PLR and Nail Psoriasis: Screen for the Early Diagnosis of Psoriatic Arthritis
Liu X, Zhao Y, Mu Z, Jia Y, Liu C, Zhang J, Cai L
Clinical, Cosmetic and Investigational Dermatology 2023, 16:1703-1713
Published Date: 28 June 2023
Adipose-Derived Stem Cell Exosomes Alleviate Psoriasis Serum Exosomes-Induced Inflammation by Regulating Autophagy and Redox Status in Keratinocytes
Kim HR, Lee SY, You GE, Kim HO, Park CW, Chung BY
Clinical, Cosmetic and Investigational Dermatology 2023, 16:3699-3711
Published Date: 23 December 2023
Mesenchymal Stem Cell–Derived Exosomes in Various Chronic Liver Diseases: Hype or Hope?
Zhu L, Wang Q, Guo M, Fang H, Li T, Zhu Y, Jiang H, Xiao P, Hu M
Journal of Inflammation Research 2024, 17:171-189
Published Date: 10 January 2024
Exosomes in Systemic Autoimmune Diseases: Recent Advances in Diagnostic Biomarkers and Therapeutic Applications
Lv X, Liu W, Zhou X, Yang Y, Zhao W, Meng L, Mu F, Zhang Z, Zhu S, Zhang S, Wang Y
International Journal of Nanomedicine 2025, 20:5137-5160
Published Date: 21 April 2025