Back to Journals » Infection and Drug Resistance » Volume 16
Escherichia coli from Human Wounds: Analysis of Resistance to β-Lactams and Expression of RND Efflux Pumps
Authors Rihacek M ![]() , Kuthanova M, Splichal Z, Adam V
, Kuthanova M, Splichal Z, Adam V ![]() , Hrazdilova K
, Hrazdilova K ![]() , Vesely R, Zurek L, Cihalova K
, Vesely R, Zurek L, Cihalova K ![]()
Received 15 August 2023
Accepted for publication 4 November 2023
Published 29 November 2023 Volume 2023:16 Pages 7365—7375
DOI https://doi.org/10.2147/IDR.S435622
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Martin Rihacek,1 Michaela Kuthanova,1 Zbynek Splichal,1 Vojtech Adam,1 Kristyna Hrazdilova,1 Radek Vesely,2,3 Ludek Zurek,1 Kristyna Cihalova1
1Department of Chemistry and Biochemistry, Mendel University in Brno, Brno, Czech Republic; 2Department of Traumatology at the Medical Faculty, Masaryk University and Trauma Hospital Brno, Brno, Czech Republic; 3Department of Traumatology, Faculty of Medicine, Masaryk University, Brno, Czech Republic
Correspondence: Kristyna Cihalova, Department of Chemistry and Biochemistry, Mendel University in Brno, Zemědělská 1, Brno, CZ-613 00, Czech Republic, Tel +420-5-4513-3350, Fax +420-5-4521-2044, Email [email protected]
Purpose: Resistance of pathogenic strains of Escherichia coli to β-lactams, particularly to ampicillin, is on the rise and it is attributed to intrinsic and acquired mechanisms. One important factor contributing to resistance, together with primarily resistance mechanisms, is a mutation and/or an over-expression of the intrinsic efflux pumps in the resistance-nodulation-division (RND) superfamily. Among these efflux pumps, AcrA, AcrB, TolC, and AcrD play an important role in antimicrobial co-resistance, including resistance to β-lactams.
Materials and Methods: Twelve E. coli isolates obtained from patients’ wounds and the control strain of E. coli ATCC 25922 were analyzed. The phenotypic resistance of these isolates to selected β-lactams was assessed by determination of the minimal inhibitory concentration. Additionally, the prevalence of β-lactamase genes (blaTEM, blaCTX-M, blaSHV, and blaAmpC) was screened by PCR. Real-time qPCR was used to determine the expression of the selected efflux pumps acrA, acrB, tolC, and acrD and the repressor acrR after the exposure of E. coli to ampicillin.
Results: Phenotypic resistance to β-lactams was detected in seven isolates, mainly to ampicillin and piperacillin. This was corroborated by the presence of at least one acquired bla gene in each of these isolates. Although E. coli strains varied in the expression of RND-family efflux pumps after the ampicillin exposure, their gene expression indicated that these pumps did not play a major role in the phenotypic resistance to ampicillin.
Conclusion: Each E. coli isolate displayed unique characteristics, differing in minimum inhibitory concentration (MIC) values, prevalence of acquired blaTEM and blaCTX-M genes, and expression of the RND-family pumps. This together demonstrates that these clinical isolates employed distinct intrinsic or acquired resistance pathways for their defense against ampicillin. The prevalence and spread of ampicillin resistant E. coli has to be monitored and the search for ampicillin alternatives is needed.
Keywords: clinical isolates, ampicillin resistance, resistance mechanisms, antibiotics
Introduction
The emergence of resistant pathogenic bacterial strains is a significant concern worldwide. Antibiotics in the β-lactams family are among the most commonly prescribed drugs with many clinical FDA (Food and Drug Administration)-approved indications and off-label clinical uses.1 They have highly reactive β-lactam ring inhibiting penicillin-binding proteins responsible for the cell wall synthesis.2 Among these, ampicillin (AMP) is commonly used as a drug of the first choice for treatments of respiratory, gastrointestinal, and urinary tract infections as well as meningitis caused by Escherichia coli, Proteus mirabilis, Salmonella species, enterococci, staphylococci, and other bacterial taxa. It is also used as prophylaxis in neonatal group B streptococcal infections and in surgeries.3,4 However, an extensive use of AMP has led to the development of bacterial resistance in many bacteria, including E. coli,5 resulting in higher antibiotic dosages for an effective treatment.6
Mechanisms of resistance to β-lactams, including ampicillin, are numerous and complex. Resistance to β-lactams is primarily mediated by β-lactamases that hydrolyze the β-lactam ring in the periplasmic space, rendering the antibiotic inactive.7 This effect can be achieved through the production of an intrinsic cephalosporinase AmpC, encoded by the blaAmpC gene and activated by a variety of β-lactams.8 In addition to the intrinsic resistance, many acquired β-lactamases have been described. The most common plasmid-mediated β-lactamases in Gram-negative bacteria are TEM and SHV enzymes, encoded by blaTEM and blaSHV with different protein variants, respectively.2 Moreover, in E. coli, CTX-M β-lactamase is also commonly present.9 Other resistance factors to β-lactams include a mutation of β-lactam targets and overexpression of efflux pumps.10,11 Efflux pumps play an important role in resistance as they export antibiotics, including β-lactams, out of bacterial cells,12,13 which leads to multidrug resistance (MDR).11,14–17 Generally, bacterial MDR efflux pumps are categorized into five families.18,19 Clinically significant in antibiotic transport in Gram-negative bacteria are efflux pumps from the resistance-nodulation-cell division (RND) superfamily,20 that includes AcrA, AcrB, TolC, and AcrD.21 So far, only the pump AcrB has been directly linked to AMP resistance.22
This study focused on the phenotypic and genotypic characterization of the clinical isolates of E. coli from the infected wounds with emphasis on the resistance to β-lactams. Furthermore, the gene expression of RND family efflux pumps (acrA, acrB, acrD, tolC) of E. coli under AMP pressure was investigated to elucidate their role in AMP resistance.
Materials and Methods
Sample Collection
Escherichia coli isolates were obtained in 2018–2019 from the long-lasting purulent wounds at different stages and with varying levels of infection severity. The collection was carried out with the agreement of 91 patients attending the Outpatient Department of Chronic Wounds at the Trauma Hospital in Brno, Czech Republic, and approved by the Ethics Committee of the hospital. After the bacterial collection by a sterile swab, swabs were immediately placed in the Amies transport medium with charcoal (Med-Lab trade, Vranovice, Czech Republic) and transferred to a laboratory. From identified isolates, the study focused only on identified E. coli isolates.
Cultivation and Identification of Isolates
For bacterial identification, the swabs were streaked on Endo agar (St. Anne´s Hospital, Brno) and incubated at 37°C overnight. After 24 h, morphologically different individual colonies were harvested from each plate and stored in 0.5 mL of 80% glycerol at −80°C. From individual colonies, bacterial strains were identified by the matrix-assisted laser desorption/ionization time-of-flight mass spectrometer (MALDI-TOF MS) Bruker ultrafleXtreme (Bruker Daltonik GmbH, Bremen, Germany). The identification was also confirmed by the 16S rRNA gene amplification and sequencing. The total DNA from each bacterial isolate was extracted with the NucleoSpin Microbial DNA kit (Macherey Nagel, Germany). The 16S rRNA gene was amplified by PCR in the 25 µL reaction mixture as follows: 0.5 µL of dNTP, 0.2 µL of Taq DNA polymerase (5000 units/mL), 2.5 µL of 10X standard Taq reaction buffer (New England Biolabs, Ipswich, MA, USA), 0.5 µL of the forward primer (10 µM) 8F (5’-AGAGTTTGATCCTGGCTCAG-3’), 0.5 µL of the reverse primer (10 µM) 1492R (5’-GGTTACCTTGTTACGACTT-3’) [23], 15.8 µL of deionized water and 5.0 µL of the template DNA (10 µg/mL~50 ng). Amplification was done on the Mastercycler nexus (Eppendorf, Germany) with cycling conditions: 90°C for 4 min., 30 cycles of 90°C for 30 sec., 58°C for 30 sec., 68°C for 105 sec., followed by 68°C extension for 10 min. and 4°C hold. For sequencing, the amplified fragments of DNA were purified by the MinElute PCR Purification Kit (Qiagen, Hilden, Germany) and sent for sequencing with amplification primers for commercial sequencing service (SeqME, Dobris, Czech Republic). Obtained sequences were assembled and compared against the GenBank database at NCBI.
Antibiotic Susceptibility Testing and Screening for the Resistance Genes
Phenotype
Antibiotic susceptibility testing (AST) was performed using the BD PhoenixTM M50 instrument (Franklin Lakes, NJ, USA). Five concentrations of each antibiotic including the breakpoint were measured. Bacterial samples were prepared according to the manufacturer guidelines. Tested β-lactams and their combinations with β-lactamase inhibitors with concentrations are listed in Table 1.
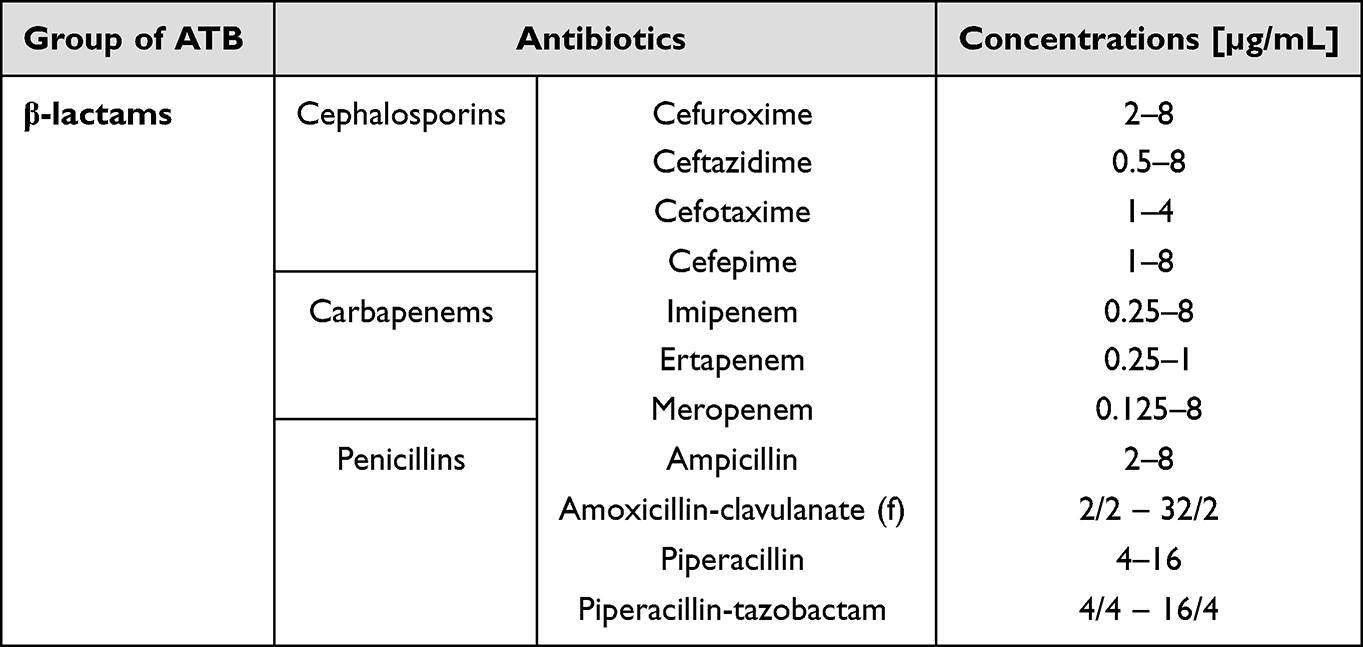 |
Table 1 Selected β-Lactams and Their Concentration Range Used for Determination of the Minimal Inhibitory Concentration |
Phenotype of Ampicillin Susceptibility by Minimum Inhibitory Concentration Determination
All E. coli isolates and the negative control strain ATCC 25922 were cultivated overnight on 5% Columbia blood agar (37°C). Overnight cultures were diluted in double concentrated Mueller-Hinton broth to OD600 0.1 AU, 100× dilution was used for MIC determination using serial dilution of ampicillin in the 96-well plates. Plates were incubated for 24 hrs at 37°C and with shaking. The final absorbance was measured (λ = 600 nm) by the MultiScan EX Microplate Photometer (Thermo Fisher, Germany). Minimum inhibitory concentration (MIC) was measured in technical duplicates.
Genotype
Preparation of the Lysates
Isolates of E. coli were grown on 5% Columbia blood agar overnight at 37°C. Five bacterial colonies of each isolate were placed in 200 µL of sterile water (LC-MS grade, LiChrosolv®, Sigma-Aldrich), re-suspended and vortexed. Bacterial solution was incubated for 10 min at 100°C and then immediately put on ice for 5 min to cause heat shock. The lysate was centrifuged (35,000 rcf, 2 min, 4°C) and supernatant containing DNA was stored at −20°C until further use.
Screening for Resistance Genes
Based on the phenotype determined by the BD PhoenixTM M50 instrument, four bla genes (Table 2), together with acrA, acrB, tolC, acrD and acrR were selected for PCR identification. The primers for acrD and acrR were designed based on the sequence of E. coli strain K-12 (NC_000913.3) using the PrimerQuest online tool of Integrated DNA Technologies (www.eu.idtdna.com) (Table 3). Escherichia coli ATCC 25922 was used as a negative control and E. coli ATCC 35218 was used as a positive control. 10 µL of the Hot Start Taq 2× Master Mix (New England BioLabs) was mixed with 0.5 μL of each primer (10 μM), 3 μL of bacterial lysate and filled up to 20 μL with sterile water. The following conditions were used for amplification: 95°C for 3 min followed by 30 cycles at 95°C for 15 sec, 60°C for 30 sec and 72°C for 60 sec with final extension 72°C for 5 min. PCR products were analyzed on 1% agarose gel, stained with MIDORIGREEN (0.06 µg/mL) and visualized by the Azure Biosystems C600 Multifunction Imaging System (Azure Biosystems, USA) under the UV light.
 |
Table 2 Primers and the Expected Size of PCR Products for Detection of Genes Encoding β-Lactamases |
 |
Table 3 Primers and the Expected Size of PCR Products of Genes Encoding the Efflux Pumps and the Repressor from RND Family Superfamily and the Reference Gene rpsL |
Expression of RND Efflux Pumps After Ampicillin Treatment
Escherichia coli was cultivated overnight on 5% Columbia blood agar at 37°C. Based on the ampicillin MIC results, one concentration of ampicillin (1.875 μg/mL) was selected to treat all isolates. The next day, three colonies from each isolate (biological triplicate) were put in Luria-Bertani broth adjusted to pH 7 with and without ampicillin in 15 mL sterile falcon tubes and incubated at 37°C for 16 h with shaking. Bacteria in the stationary phase were harvested and washed once with phosphate-buffered saline (6440 rcf, 10 min, 4°C).
RNA was isolated by the TRIzol Reagent® according to manufacturer instructions (TRIzol Reagent, Invitrogen, Carlsbad, CA) with the modification of the lysis step after homogenization, when the samples were incubated at −20°C for 10 min. After the isolation, RNA was purified by the ethanol precipitation protocol according to manufacturer instructions28 with modification of incubation time and temperature. Instead of −20°C for overnight, we used −80°C for one hour and the remaining DNA was removed by the TURBO DNA-freeTM Kit (Ambion®, Life Technologies Europe BV) using 5.0 μg of the total RNA. The concentration and purity of RNA was determined by the NanoDropTM OneC Spectrophotometer (Thermo Fischer Scientific, Waltham, MA, USA). RNA integrity was verified by gel electrophoresis in 2% agarose gel (90V/30min) supplemented with 1% of bleach and stained by MIDORIGREEN (0.06 μg/mL). The gel was visualized by Azure c600 from Azure Biosystems (Dublin, California, USA). The RNA samples were aliquoted and stored in −80°C until further analyses.
cDNA of bacterial isolates was prepared by the first strand cDNA synthesis kit for RT-PCR (Roche, Germany, Mannheim), using random hexamers as primers, according to manufacturer instructions. Five hundred nanograms of RNA after the TURBO DNase treatment were used as a template for reverse transcription. Prepared cDNA (20 μL) was diluted in UltraPureTM Dnase/Rnase-Free Distilled Water to a total volume 200 μL.
All bacterial isolates were tested for the presence of MDR efflux pumps genes acrA, acrB, acrD, tolC, repressor acrR and the reference gene rpsL (Table 3) by PCR. If the genes were detected in E. coli isolates, transcriptional expression of these pumps by real-time qPCR was determined. Analysis was performed using the qTOWER3 system (Analytik Jena, Jena, Germany). For each reaction, 10 μL of diluted reverse-transcribed cDNA (25 ng/rxn) was subjected to PCR amplification in a 20 μL final volume containing 9 μL of Luna® Universal qPCR Master Mix (New England Biolabs) and 0.5 μL (10 μM) of each primer. The following conditions were used for amplification: 95°C for 3 min, 30 cycles at 95°C for 15 sec, 60°C for 30 sec and 72°C for 30 sec with final extension at 72°C for 2 min. To ensure the specificity of the amplified PCR products, a melting curve analysis was performed.
The relative quantities of mRNA of each gene of interest were calculated by ΔΔCT method. First of all, the expression of each gene of interest (GOI) was normalized to the reference gene (REF) rpsL (ΔCt = CtREF – CtGOI), then the differences in relative gene expression between treated and untreated isolates were calculated (ΔΔCt = ΔCtTreated – ΔCtUntreated). Results were presented as mean of ΔΔCt (~Log2 Fold Change) from three biological replicates and ± SEM calculated by GraphPad Prism 8.0.1. (GraphPad Software, CA, USA). This program was also used to perform an unpaired t-test between untreated and treated isolate from ΔCt values of biological triplicate to determine the significance (p < 0.05) of the obtained differences.27
Results
Isolates Quantification
A total of 219 bacterial isolates were collected from 91 clinical patients with different types of infected wounds. In this study, we specifically focused on up to four E. coli isolates from each patient (twelve in total) (Table S1).
Phenotypic Determination of E. coli Resistance to β-Lactams
The MIC of all tested β-lactams and their combination with inhibitors is shown in Table S2. Four out of twelve isolates were susceptible to all tested β-lactams. One isolate (38B1a) showed resistance only to ertapenem and remaining seven isolates were resistant to two or more of tested β-lactams (Table S2). From all E. coli isolates, 58.3% showed resistance to ampicillin and piperacillin respectively, and 33.3% to amoxicillin-clavulanate (Table 4). Based on these findings, we focused on ampicillin resistance exclusively. In this study, the MIC of ampicillin was assessed for all isolates, including the control strain E. coli ATCC 25922. The categorization of isolates as either sensitive or resistant to AMP was based on the MIC values according to the EUCAST guidelines for the year 202229 (Table 4). EUCAST considers an MIC value above 8.0 µg/mL as indicative of resistance to AMP.23–26
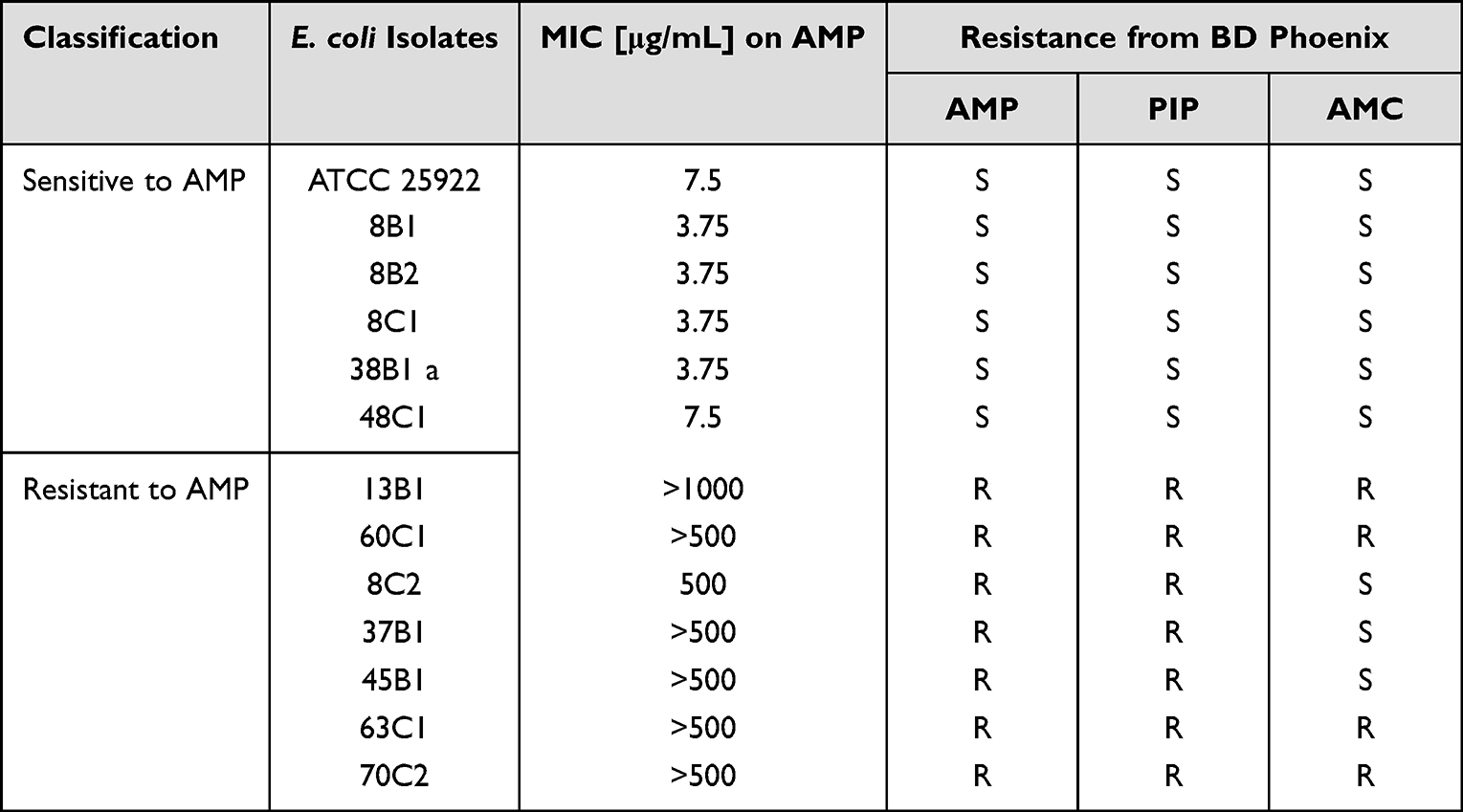 |
Table 4 Classification of E. coli Isolates and the Control Strain ATCC 25922 Based on the Minimal Inhibitory Concentration (MIC) to Ampicillin (AMP). The BD Phoenix System Classification of Isolates as Susceptible (S) and Resistant (R) to Ampicillin (AMP), Piperacillin (PIP) and Amoxicillin-Clavulanic Acid (AMC) |
Genotypic Determination of bla Genes
Bacterial isolates were examined for the presence of four bla genes. Escherichia coli isolates sensitive to AMP did not carry any acquired bla genes (blaTEM, blaCTX-M, blaSHV) and only the intrinsic blaAmpC gene was detected. In contrast, in strains resistant to AMP, in addition to blaAmpC, at least one acquired bla gene was found. The blaTEM gene was present in 6 out 7 strains and the strain without blaTEM and one more isolate were positive for blaCTX-M (Table 5).
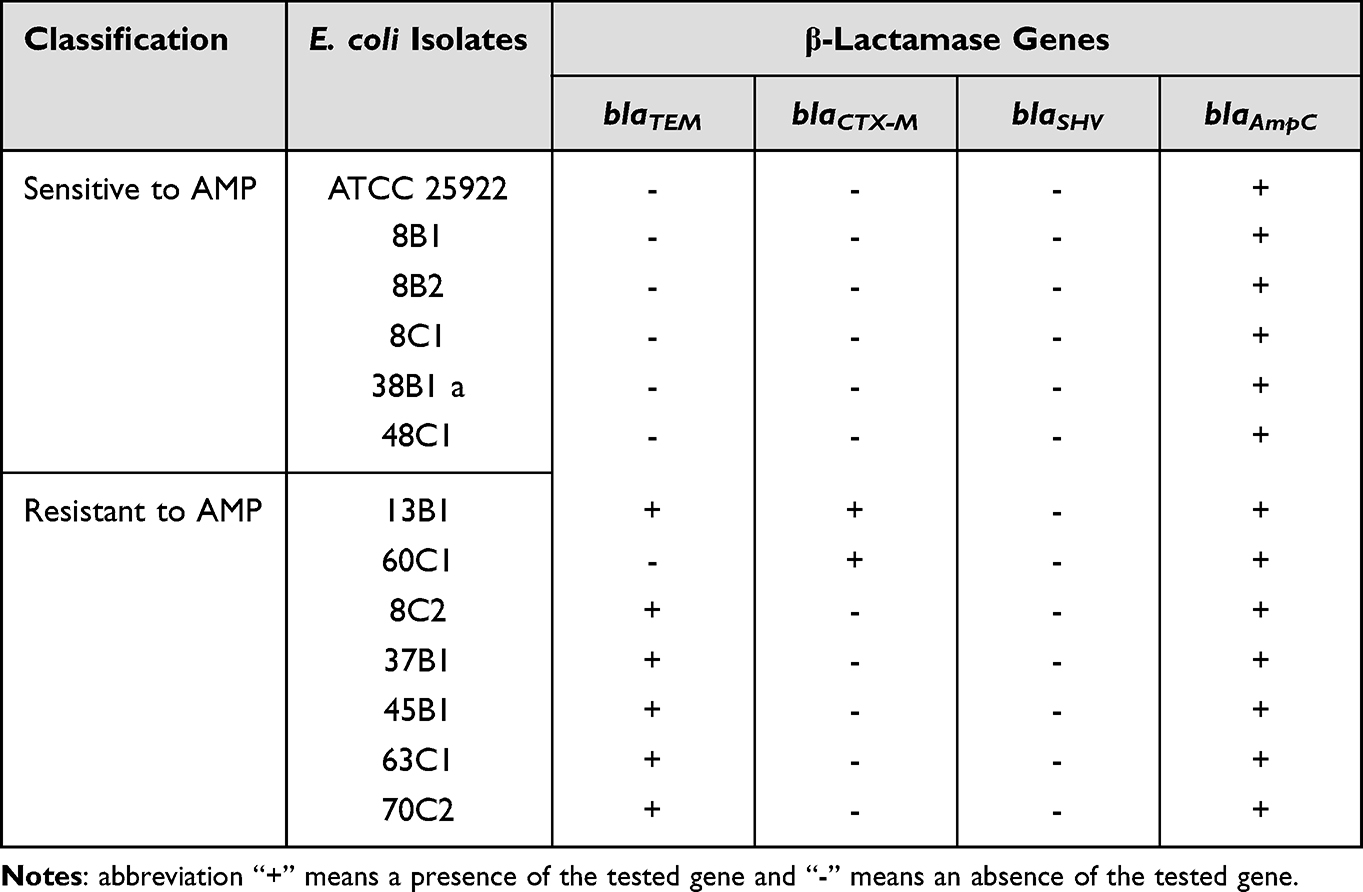 |
Table 5 Prevalence of β-Lactam Resistance Genes in E. coli Isolates |
Effect of Ampicillin on Expression of RND-Family Efflux Pumps Genes in E. coli
Before conducting the expression analyses, the presence of MDR efflux pump genes (acrA, acrB, acrD, tolC), as well as the acrR gene coding for the transcriptional regulatory protein, was confirmed in all isolates using PCR.
Tested E. coli differed in the expression of tested genes after ampicillin exposure (Figure 1). Among susceptible strains with no acquired bla genes, a significant up-regulation of acrB was found in the control strain (2.1× higher) and the isolate 48C1, where the expression level was 5.6× higher. In contrast, a significant down-regulation of acrA (2.8× decreased in 8B1 and 3.7× decreased for 8C1) and acrD (15.9× decreased for 8B1 and 18.2× decreased for 8C1) was detected in isolates 8B1 and 8C1. In the control strain, down-regulation of acrD (15.3× decreased) and acrR (4.6× decreased) was found as well and no significant changes in the expression of these genes were observed in all other susceptible isolates (Figure 1). In resistant isolates, the significant changes in expression were seen in 13B1, 37B1, 63C1 and 70C2 isolates. Among them, 13B1 and 37B1 isolates with the blaTEM gene, had up-regulated acrA. A significant down-regulation of acrB (2.7× decreased for 63C1 and 3.4× decreased for 70C2) was observed in 70C2 and 63C1 isolates, containing the blaTEM gene. Furthermore, in the isolate 70C2, the acrD gene was also down-regulated (20.7× decreased). No significant changes in expression of tolC were detected in any of our isolates (Figure 1).
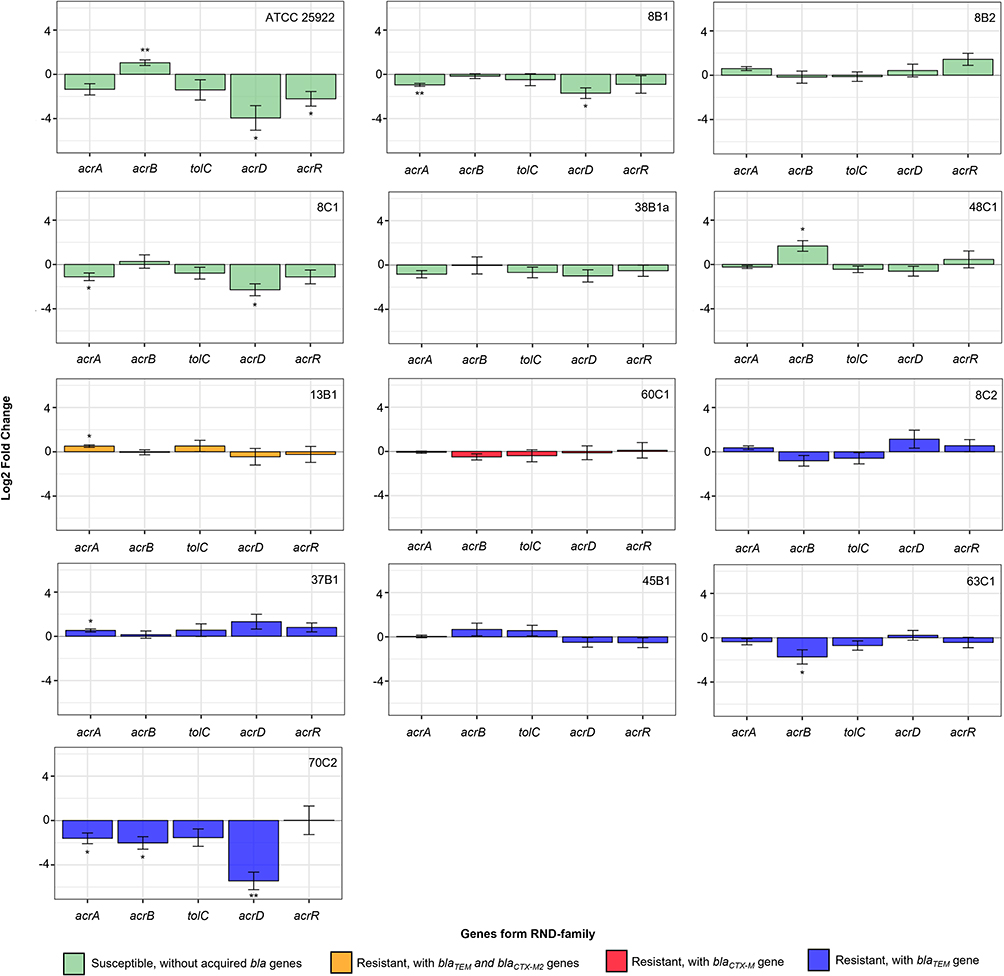 |
Figure 1 Expression of tested genes acrA, acrB, tolC, acrD and acrR from RND family in E. coli isolates: ATCC 25922, 8B1, 8B2, 8C1, 38B1a, 48C1, 13B1, 60C1, 8C2, 37B1, 45B1, 63C1, 70C2 after AMP treatment normalized to gene rpsL. Untreated and treated isolate after AMP exposure were compared and significance was calculated by unpaired t-test from ΔCt values. The significance is represented as stars according to the p-value (* p-value ≤ 0.05, ** p-value ≤ 0.01) above or below the column. Green coloured columns represent susceptible isolates, Orange isolates containing two acquired bla genes, blue and red isolates containing only one type of bla gene. |
Discussion
Resistance of clinical isolates of E. coli to β-lactams, especially to ampicillin, is on a rise.30–34 About twenty years ago, the prevalence of ampicillin resistance in the human clinical isolates was only around 8.2%.35,36 Even when the resistance occurred, the MIC was slightly above the FDA breakpoint (>8 µg/mL) only, with rare exceptions, when the MIC to ampicillin reached >236 µg/mL.37,38 Nowadays the ampicillin resistance in E. coli isolates from human infections has increased to nearly 85% with MIC values commonly exceeding >250 µg/mL.39–44 In our study, MIC values of E. coli resistant isolates have been even much higher and exceeded 500 µg/mL.5,41,42 The staggering ampicillin MIC is likely due to combination of several β-lactam resistance mechanisms.5,41–43
Resistance to β-lactams is primarily mediated by β-lactamases.7 Presence of the AmpC cephalosporinase in all E. coli results in resistance to most penicillins.44 In the presence of extended spectrum of β-lactamases (ESBLs) from the acquired plasmids, the expression of blaAmpC gene is either poorly expressed or not expressed at all.8 In our study, both susceptible and resistant E. coli strains carried the blaAmpC gene. Within susceptible isolates, this gene ostensibly contributes to a marginally elevated ampicillin MIC that does not surpass the FDA breakpoint.8 Conversely, in resistant strains featuring ESBLs, the presence of this gene appears to have no influence on the heightened MIC against ampicillin. Notably, the elevated MIC levels towards ampicillin are likely attributed to the presence of acquired bla genes. Noteworthy, among the β-lactamases in E. coli are TEM, CTX-M, and SHV, with the lion’s share of ampicillin resistance being attributed to TEM.45 While CTX-M and SHV β-lactamases have been linked to ampicillin resistance in Klebsiella pneumoniae clinical isolates,46–48 their link to ampicillin resistance of E. coli has not been established.49,50 In our study, as in other clinical studies, blaTEM was found to be the most common gene encoding β-lactamases followed by blaCTX-M and blaSHV.5,51–54 In the presence of both blaTEM and blaCTX-M, the MIC for ampicillin in E. coli clinical isolates was in range 32.0 to 256.0 µg/mL.54,55 Comparing to the upper MIC levels in the previous studies, in our study, E. coli isolates with blaTEM or blaCTX-M had the MIC for ampicillin four times higher (>1000 µg/mL), and two times higher (>500 µg/mL). Overall, ESBLs aside, E. coli probably employs a variety of resistance mechanisms against β-lactams, which are needed to elevate the MIC for ampicillin.
Escherichia coli commonly employs a diverse set of efflux pumps from the RND-superfamily resulting in resistance to antibiotics. Among these pumps, only the AcrB has been directly linked to ampicillin resistance.22 Additionally, the AcrA pump, working together with AcrB, is responsible for intrinsic resistance to fluoroquinolones and aminoglycosides.27 Both their genes, acrA and acrB, are located on the same operon, therefore their expression levels should correlate56 as confirmed in several studies;27,57 however, other studies reported an opposite regulation.57–59 Both, the same and opposite, expressions were found in our E. coli isolates after ampicillin treatment as well. Expression of acrA and acrB genes is influenced by the repressor protein AcrR,60 thus the overexpression of acrAB would correlate with down-regulation of the acrR gene. In our study, this trend was seen in the control strain ATCC 25922. In other isolates, the expression of acrR remained unchanged, even when acrA or acrB showed different expressions. This might be due to a mutation in the acrR gene or the involvement of other acrAB repressors with higher affinity to acrAB protomor, such as AcrS. Furthermore, the binding affinity of AcrR in the acrAB promotor is lower than that of AcrS.61 The pump AcrA and AcrB can assemble with other pumps from the RND-family and create a tripartite system spanning both the inner and the outer membrane, leading to multidrug resistance.62,63 While the primary tripartite system is composed of AcrAB and the TolC pump, when the AcrD pump is overexpressed, it can replace AcrB and this newly formed system can confer resistance to antibiotics as well.64,65 Significant down-regulation for the acrD gene was observed in several E. coli isolates after ampicillin treatment but no significant changes were detected in tolC. Based on these results, it is likely that the assembly of the tripartite system does not take place under ampicillin pressure, as the AcrD pump is primarily linked to aminoglycosides resistance and may not be necessary for survival in the presence of ampicillin. Resistance to ampicillin in E. coli is commonly caused by a combination of several resistance mechanisms. In our study, we primarily detected the presence of ESBLs genes. The up-regulation of the acrB gene was only observed in two out of six sensitive E. coli strains, suggesting it may have some protective effect against ampicillin. Overall, the resistance mechanisms in clinical isolates of E. coli to ampicillin are complex and numerous and further studies analysing more isolates are needed to better understand all mechanisms E. coli employs for protection to ampicillin and other β-lactams.
Conclusions
This pilot study aimed to investigate ampicillin resistance and its mechanisms in E. coli isolates from human wounds. Phenotype and genotype screening for β-lactam resistance genes and the expression of efflux pumps from the RND-family under ampicillin stress were performed. Remarkably, each E. coli isolate displayed unique characteristics, differing in the MIC values to ampicillin as well as in the prevalence of acquired blaTEM and blaCTX-M genes and in the expression level of the RND-family pumps. This diversity indicates that these clinical isolates employed distinct intrinsic or acquired resistance pathways for their defence against ampicillin. The prevalence and spread of ampicillin resistance in clinical as well as non-clinical isolates should therefore be monitored and new antimicrobial compounds developed to replace ampicillin in the future.
Abbreviations
AMP, ampicillin; MDR, multi-drug resistance; MIC, minimum inhibitory concentration; RND, resistance-nodulation-division.
Ethics Statement
The study was approved by the Ethics Committee of the Trauma Hospital in Brno. The relevant guidelines and regulations provided in the Declaration of Helsinki were followed for handling human samples. Informed consent was waived by the Ethics Committee of the Trauma Hospital in Brno since this study did not involve any interventions. Patients were treated anonymously.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
The study was financially supported by EDRF “Multidisciplinary research to increase application potential of nanomaterials in agricultural practise” (No. CZ.03.1.01/0.0/0.0/16_025/0007314).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Pandey N, Cascella M. Beta lactam antibiotics. 2019.
2. Goussard S, Courvalin P. Updated sequence information for TEM β-lactamase genes. Antimicrob Agents Chemother. 1999;43(2):367–370. doi:10.1128/AAC.43.2.367
3. Osmon DR, Berbari EF, Berendt AR, et al. Executive summary: diagnosis and management of prosthetic joint infection: clinical practice guidelines by the Infectious Diseases Society of America. Clin Infect Dis. 2013;56(1):1–10. doi:10.1093/cid/cis966
4. Bratzler DW, Dellinger EP, Olsen KM, et al. Clinical practice guidelines for antimicrobial prophylaxis in surgery. Surg Infect (Larchmt). 2013;14(1):73–156. doi:10.1089/sur.2013.9999
5. Li M, Liu Q, Teng Y, et al. The resistance mechanism of Escherichia coli induced by ampicillin in laboratory. Infect Drug Resist. 2019;12:2853. doi:10.2147/IDR.S221212
6. Katzung BG, Masters SB, Trevor AJ. Basic & Clinical Pharmacology. Ch. 8. New York: McGraw-Hill; 2012.
7. Livermore DM. Beta-lactamases in laboratory and clinical resistance. Clin Microbiol Rev. 1995;8(4):557–584. doi:10.1128/CMR.8.4.557
8. Jacoby GA. AmpC β-lactamases. Clin Microbiol Rev. 2009;22(1):161–182. doi:10.1128/CMR.00036-08
9. Tan P, Wang Z, Sun A, Yan J, Zhao J. Predominant β-lactamase genotypes of Escherichia coli isolates and induction and inhibition mechanisms of β-lactamase gene expression. Zhonghua liu Xing Bing xue za zhi. 2015;36(5):484–490.
10. Alcalde-Rico M, Hernando-Amado S, Blanco P, Martínez JL. Multidrug efflux pumps at the crossroad between antibiotic resistance and bacterial virulence. Front Microbiol. 2016;7:1483. doi:10.3389/fmicb.2016.01483
11. Du D, Wang-Kan X, Neuberger A, et al. Multidrug efflux pumps: structure, function and regulation. Nat Rev Microbiol. 2018;16(9):523–539. doi:10.1038/s41579-018-0048-6
12. Blanco P, Hernando-Amado S, Reales-Calderon JA, et al. Bacterial multidrug efflux pumps: much more than antibiotic resistance determinants. Microorganisms. 2016;4(1):14. doi:10.3390/microorganisms4010014
13. Hinchliffe P, Symmons MF, Hughes C, Koronakis V. Structure and operation of bacterial tripartite pumps. Annu Rev Microbiol. 2013;67:221–242. doi:10.1146/annurev-micro-092412-155718
14. Yasufuku T, Shigemura K, Shirakawa T, et al. Correlation of overexpression of efflux pump genes with antibiotic resistance in Escherichia coli strains clinically isolated from urinary tract infection patients. J Clin Microbiol. 2011;49(1):189–194. doi:10.1128/JCM.00827-10
15. Yang S, Clayton SR, Zechiedrich EL. Relative contributions of the AcrAB, MdfA and NorE efflux pumps to quinolone resistance in Escherichia coli. J Antimicrob Chemother. 2003;51(3):545–556. doi:10.1093/jac/dkg126
16. Li X-Z, Plésiat P, Nikaido H. The challenge of efflux-mediated antibiotic resistance in Gram-negative bacteria. Clin Microbiol Rev. 2015;28(2):337–418. doi:10.1128/CMR.00117-14
17. Falagas ME, Koletsi PK, Bliziotis IA. The diversity of definitions of multidrug-resistant (MDR) and pandrug-resistant (PDR) Acinetobacter baumannii and Pseudomonas aeruginosa. J Med Microbiol. 2006;55(12):1619–1629. doi:10.1099/jmm.0.46747-0
18. Delmar JA, Yu EW. The A bg T family: a novel class of antimetabolite transporters. Prot Sci. 2016;25(2):322–337. doi:10.1002/pro.2820
19. Hassan KA, Elbourne LD, Li L, et al. An ace up their sleeve: a transcriptomic approach exposes the AceI efflux protein of Acinetobacter baumannii and reveals the drug efflux potential hidden in many microbial pathogens. Front Microbiol. 2015;6:333. doi:10.3389/fmicb.2015.00333
20. Nikaido H. Efflux-mediated drug resistance in bacteria: an update. Drugs. 2009;69(12):1555–1623. doi:10.2165/11317030-000000000-00000
21. Elkins CA, Nikaido H. Substrate specificity of the RND-type multidrug efflux pumps AcrB and AcrD of Escherichia coli is determined predominately by two large periplasmic loops. J Bacteriol. 2002;184(23):6490–6498. doi:10.1128/JB.184.23.6490-6499.2002
22. Wang Z, Zhang P, Ding X, et al. Co-delivery of ampicillin and β-lactamase inhibitor by selenium nanocomposite to achieve synergistic anti-infective efficiency through overcoming multidrug resistance. Chem Eng J. 2021;414:128908. doi:10.1016/j.cej.2021.128908
23. Zheng B, Zhang J, Ji J, et al. Emergence of Raoultella ornithinolytica coproducing IMP-4 and KPC-2 carbapenemases in China. Antimicrob Agents Chemother. 2015;59(11):7086–7089. doi:10.1128/AAC.01363-15
24. Feizabadi MM, Delfani S, Raji N, et al. Distribution of bla TEM, bla SHV, bla CTX-M genes among clinical isolates of Klebsiella pneumoniae at Labbafinejad Hospital, Tehran, Iran. Microbial Drug Resist. 2010;16(1):49–53. doi:10.1089/mdr.2009.0096
25. Belaaouaj A, Lapoumeroulie C, Caniça MM, et al. Nucleotide sequences of the genes coding for the TEM-like β-lactamases IRT-1 and IRT-2 (formerly called TRI-1 and TRI-2). FEMS Microbiol Lett. 1994;120(1–2):75–80. doi:10.1111/j.1574-6968.1994.tb07010.x
26. Féria C, Ferreira E, Correia JD, Gonçalves J, Caniça M. Patterns and mechanisms of resistance to β-lactams and β-lactamase inhibitors in uropathogenic Escherichia coli isolated from dogs in Portugal. J Antimicrob Chemother. 2002;49(1):77–85. doi:10.1093/jac/49.1.77
27. Swick MC, Morgan-Linnell SK, Carlson KM, Zechiedrich L. Expression of multidrug efflux pump genes acrAB-tolC, mdfA, and norE in Escherichia coli clinical isolates as a function of fluoroquinolone and multidrug resistance. Antimicrob Agents Chemother. 2011;55(2):921–924. doi:10.1128/AAC.00996-10
28. Scientific T. Sodium acetate/ethanol precipitation protocol. ThermoFisher Scientific; 2023. Available from: https://www.thermofisher.com/cz/en/home/references/protocols/nucleic-acid-purification-and-analysis/dna-protocol/sodium-acetate-precipitation-of-small-nucleic-acids.html.
29. Testing, E.C.o.A.S. Breakpoint tables for interpretation of MICs and zone diameters; 2021.
30. Tewari R, Ganaie F, Venugopal N, Mitra S, Shome R, Shome BR. Occurrence and characterization of genetic determinants of β-lactam-resistance in Escherichia coli clinical isolates. Infect Genet Evol. 2022;100:105257. doi:10.1016/j.meegid.2022.105257
31. Röderova M, Halova D, Papousek I, et al. Characteristics of quinolone resistance in Escherichia coli isolates from humans, animals, and the environment in the Czech Republic. Front Microbiol. 2017;7:2147. doi:10.3389/fmicb.2016.02147
32. Pandit R, Awal B, Shrestha SS, Joshi G, Rijal BP, Parajuli NP. Extended-spectrum β-lactamase (ESBL) genotypes among multidrug-resistant uropathogenic Escherichia coli clinical isolates from a teaching hospital of Nepal. Interdiscip Perspect Infect Dis. 2020;2020. doi:10.1155/2020/6525826
33. Oberoi L, Singh N, Sharma P, Aggarwal A. ESBL, MBL and Ampc β lactamases producing superbugs–Havoc in the Intensive Care Units of Punjab India. J Clin Diagn Res. 2013;7(1):70. doi:10.7860/JCDR/2012/5016.2673
34. Jafari SA, Mobaiyen H. Frequency and resistance patterns in clinical isolates of Escherichia coli Extended Spectrum Beta Lactamase producing treatment Centers in Marand city, Iran; 2017.
35. Mesa-Varona O, Kaspar H, Grobbel M, Tenhagen B-A. Phenotypical antimicrobial resistance data of clinical and non-clinical Escherichia coli from poultry in Germany between 2014 and 2017. PLoS One. 2020;15(12):e0243772. doi:10.1371/journal.pone.0243772
36. Sáenz Y, Zarazaga M, Briñas L, Lantero M, Ruiz-Larrea F, Torres C. Antibiotic resistance in Escherichia coli isolates obtained from animals, foods and humans in Spain. Int J Antimicrob Agents. 2001;18(4):353–358. doi:10.1016/S0924-8579(01)00422-8
37. Maynard C, Bekal S, Sanschagrin F, et al. Heterogeneity among virulence and antimicrobial resistance gene profiles of extraintestinal Escherichia coli isolates of animal and human origin. J Clin Microbiol. 2004;42(12):5444–5452. doi:10.1128/JCM.42.12.5444-5452.2004
38. Brinas L, Zarazaga M, Sáenz Y, Ruiz-Larrea F, Torres C. β-Lactamases in ampicillin-resistant Escherichia coli isolates from foods, humans, and healthy animals. Antimicrob Agents Chemother. 2002;46(10):3156–3163. doi:10.1128/AAC.46.10.3156-3163.2002
39. Alharbi NS, Khaled JM, Kadaikunnan S, et al. Prevalence of Escherichia coli strains resistance to antibiotics in wound infections and raw milk. Saudi J Biol Sci. 2019;26(7):1557–1562. doi:10.1016/j.sjbs.2018.11.016
40. Naik TB, Mane V, Biradar AJP. Occurrence of extended-spectrum beta-lactamases and the Antibiogram Pattern of Escherichia coli Isolates from Pus and Urine Samples. J Pure Appl Microbiol. 2022;15(26).
41. Yenn TW, Khan MA, Syuhada NA, Ring LC, Ibrahim D, Tan W-N. Stigmasterol: an adjuvant for beta lactam antibiotics against beta-lactamase positive clinical isolates. Steroids. 2017;128:68–71. doi:10.1016/j.steroids.2017.10.016
42. Sjölund M, Bengtsson S, Bonnedahl J, Hernandez J, Olsen B, Kahlmeter G. Antimicrobial susceptibility in Escherichia coli of human and avian origin—a comparison of wild-type distributions. Clin Microbiol Infect. 2009;15(5):461–465. doi:10.1111/j.1469-0691.2009.02705.x
43. Rodríguez-Villodres Á, Gil-Marqués ML, Álvarez-Marín R, et al. Extended-spectrum resistance to β-lactams/β-lactamase inhibitors (ESRI) evolved from low-level resistant Escherichia coli. J Antimicrob Chemother. 2020;75(1):77–85. doi:10.1093/jac/dkz393
44. Deshpande LM, Jones RN, Fritsche TR, Sader HS. Occurrence of plasmidic AmpC type β-lactamase-mediated resistance in Escherichia coli: report from the SENTRY Antimicrobial Surveillance Program (North America, 2004). Int J Antimicrob Agents. 2006;28(6):578–581. doi:10.1016/j.ijantimicag.2006.07.025
45. Tadesse DA, Zhao S, Tong E, et al. Antimicrobial drug resistance in Escherichia coli from humans and food animals, United States, 1950–2002. Emerg Infect Dis. 2012;18(5):741. doi:10.3201/eid1805.111153
46. Bora A, Hazarika NK, Shukla SK, Prasad KN, Sarma JB, Ahmed G. Prevalence of blaTEM, blaSHV and blaCTX-M genes in clinical isolates of Escherichia coli and Klebsiella pneumoniae from Northeast India. Indian J Pathol Microbiol. 2014;57(2):249. doi:10.4103/0377-4929.134698
47. Ojdana D, Sacha P, Wieczorek P, et al. The occurrence of blaCTX-M, blaSHV, and blaTEM genes in extended-spectrum β-lactamase-positive strains of Klebsiella pneumoniae, Escherichia coli, and Proteus mirabilis in Poland. Int J Antibiot. 2014;2014:1–7. doi:10.1155/2014/935842
48. Olesen I, Hasman H, Møller Aarestrup F. Prevalence of β-lactamases among ampicillin-resistant Escherichia coli and Salmonella isolated from food animals in Denmark. Microbial Drug Resist. 2004;10(4):334–340. doi:10.1089/mdr.2004.10.334
49. Fu Y, Zhang F, Zhang W, et al. Differential expression of blaSHV related to susceptibility to ampicillin in Klebsiella pneumoniae. Int J Antimicrob Agents. 2007;29(3):344–347. doi:10.1016/j.ijantimicag.2006.10.015
50. Chen Y, Zhao J, Ding F, et al. The blaCTX-M gene independently enhances drug resistance level to ampicillin in clinical isolates of Klebsiella pneumoniae. J Antibiot (Tokyo). 2012;65(9):479–481. doi:10.1038/ja.2012.44
51. Lesani SS, Soleimani M, Shakib P, Zolfaghari MR. Prevalence of blaCTX-M, blaSHV, and blaTEM genes in Escherichia coli strains isolated from urinary tract infection samples of patients in the Intensive Care Unit in Qom, Iran. Gene Cell Tissue. 2020;7(2). doi:10.5812/gct.102700
52. Pishtiwan AH, Khadija KM. Prevalence of blaTEM, blaSHV, and blaCTX-M genes among ESBL-producing Klebsiella pneumoniae and Escherichia coli isolated from thalassemia patients in Erbil, Iraq. Mediterr J Hematol Infect Dis. 2019;11(1). doi:10.4084/MJHID.2019.041
53. Ugbo E, Anyamene C, Moses I, et al. Prevalence of blaTEM, blaSHV, and blaCTX-M genes among extended spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae of clinical origin. Gene Rep. 2020;21:100909. doi:10.1016/j.genrep.2020.100909
54. Nawaz M, Khan A, Khan S, et al. Identification of novel plasmid replicons harboring β-lactamase resistant genes in ampicillin-resistant uropathogenic Escherichia coli. J Bacteriol Mycol. 2019;6:1102.
55. Soares CRP, Magalhães V, de Araújo PSR. Coexistence of blaTEM, blaCTX, blaKPC, blaNDM, blaSIM e blaOXA-48 in polymicrobial bloodstream isolates from a patient with acute myeloid leukemia. Res Soc Dev. 2021;10(5):e39310514985–e39310514985. doi:10.33448/rsd-v10i5.14985
56. Grkovic S, Brown MH, Skurray RA. Regulation of bacterial drug export systems. Microbiol Mol Biol Rev. 2002;66(4):671–701. doi:10.1128/MMBR.66.4.671-701.2002
57. Mazzariol A, Tokue Y, Kanegawa TM, Cornaglia G, Nikaido H. High-level fluoroquinolone-resistant clinical isolates of Escherichia coli overproduce multidrug efflux protein AcrA. Antimicrob Agents Chemother. 2000;44(12):3441–3443. doi:10.1128/AAC.44.12.3441-3443.2000
58. Chetri S, Bhowmik D, Paul D, et al. AcrAB-TolC efflux pump system plays a role in carbapenem non-susceptibility in Escherichia coli. BMC Microbiol. 2019;19(1):1–7. doi:10.1186/s12866-019-1589-1
59. Atac N, Kurt-Azap O, Dolapci I, et al. The role of AcrAB–TolC efflux pumps on quinolone resistance of E. coli ST131. Curr Microbiol. 2018;75(12):1661–1666. doi:10.1007/s00284-018-1577-y
60. Ma D, Alberti M, Lynch C, Nikaido H, Hearst JE. The local repressor AcrR plays a modulating role in the regulation of acrAB genes of Escherichia coli by global stress signals. Mol Microbiol. 1996;19(1):101–112. doi:10.1046/j.1365-2958.1996.357881.x
61. Li M, Gu R, Su -C-C, et al. Crystal structure of the transcriptional regulator AcrR from Escherichia coli. J Mol Biol. 2007;374(3):591–603. doi:10.1016/j.jmb.2007.09.064
62. Blair JM, Richmond GE, Piddock LJ. Multidrug efflux pumps in Gram-negative bacteria and their role in antibiotic resistance. Future Microbiol. 2014;9(10):1165–1177. doi:10.2217/fmb.14.66
63. Anes J, McCusker MP, Fanning S, Martins M. The ins and outs of RND efflux pumps in Escherichia coli. Front Microbiol. 2015;6:587. doi:10.3389/fmicb.2015.00587
64. Aires JR, Nikaido H. Aminoglycosides are captured from both periplasm and cytoplasm by the AcrD multidrug efflux transporter of Escherichia coli. J Bacteriol. 2005;187(6):1923–1929. doi:10.1128/JB.187.6.1923-1929.2005
65. Sano N, Kim J-S, Onda Y, et al. RNA-Seq using bulked recombinant inbred line populations uncovers the importance of brassinosteroid for seed longevity after priming treatments. Sci Rep. 2017;7(1):8095. doi:10.1038/s41598-017-08116-5
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.