Back to Journals » Infection and Drug Resistance » Volume 16
ESBL-Producing and Non-ESBL-Producing Escherichia coli Isolates from Urinary Tract Differ in Clonal Distribution, Virulence Gene Content and Phylogenetic Group
Authors Fang Y ![]() , Tao S
, Tao S ![]() , Chen H, Xu Y, Chen L, Liang W
, Chen H, Xu Y, Chen L, Liang W
Received 23 June 2023
Accepted for publication 15 August 2023
Published 23 August 2023 Volume 2023:16 Pages 5563—5571
DOI https://doi.org/10.2147/IDR.S423860
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Sandip Patil
Yewei Fang,1 Shuan Tao,2 Huimin Chen,2 Yao Xu,3 Luyan Chen,4 Wei Liang1
1Department of Clinical Laboratory, The First Affiliated Hospital of Ningbo University, Ningbo, People’s Republic of China; 2School of Medicine, Jiangsu University, Zhejiang, People’s Republic of China; 3School of Medicine, Ningbo University, Ningbo, People’s Republic of China; 4Department of Blood Transfusion, The First Affiliated Hospital of Ningbo University, Ningbo, People’s Republic of China
Correspondence: Wei Liang, Department of Clinical Laboratory, The First Affiliated Hospital of Ningbo University, No. 59. Liuting Road, Haishu District, Zhejiang, 315010, People’s Republic of China, Tel +86 15351883016, Fax +86-51885213100, Email [email protected]
Purpose: The objectives of this study are to determine the differences in clonality, virulence gene (VG) content and phylogenetic group between non extended-spectrum beta-lactamase-producing E. coli (non-ESBL-EC) and ESBL-EC isolates from urine.
Patients and Methods: This study characterized a total of 100 clinical E. coli isolates consecutively obtained from the inpatients hospitalized in The First Affiliated Hospital of Ningbo University in China by polymerase-chain reaction (PCR).
Results: Phylogenetic group B2 was found to be the most prevalent in both ESBL-EC and non-ESBL-EC group. Among 100 clinical isolates, the count of acquired virulence genes in group B2 was found to be significantly higher than that in group A, B1, and D (p < 0.001). Additionally, the presence of content within virulence genes (the total number of virulence genes detected per isolate) in B2 of non-ESBL-EC and ESBL-EC showed a significant difference (p< 0.001). ST131 was detected exclusively in ESBL-EC, while ST95 and ST73 were the main sequence types in non-ESBL-EC.
Conclusion: Our study demonstrated the different distribution of MLST, phylogenetic group in ESBL-EC and non-ESBL-EC group. The inverse association between beta-lactamase resistance and VG content performed in this study should get a lot more attention. At the same time, we should also be wary of the appearance of non-ESBL-EC isolates of group B2 harboring more virulence genes which will lead to high pathogenicity.
Keywords: E. coli, clonal structure, ST73, ST95, ST131, ST1193, virulence genes
Introduction
Escherichia coli is the primary cause of urinary tract infections (UTI).1,2 Extended-spectrum beta-lactamases (ESBL)-producing strains, which are resistant to extended-spectrum cephalosporins and monobactams3,4, play an important role in UTI.
The prevalence of ESBL-producing E. coli (ESBL-EC) is on the rise in the United States,5 Africa,6 and Vietnam regions,7 despite variations in geography and population. In Jordan, the prevalence of ESBL-EC reached 62% in hospitalized patients with UTIs,8,9 which is a concerning trend. Similarly, China has shown a high prevalence of ESBL-EC isolates in urinary tract infections.10 Unfortunately, the common treatment for ESBL-EC involves parenteral aminoglycosides or carbapenems, which require hospitalization11 and increase financial burden.12 Therefore, it is of great significance to understand the clinical characteristics of E. coli causing UTI episodes, as they represent different target populations for prevention. In order to develop effective preventive strategies for UTIs caused by both non-ESBL-EC and ESBL-EC, it is important to gain thorough insight into their molecular epidemiology and clinical characteristics. These information can guide the development of targeted interventions, including vaccines currently in development.13 Currently, there is limited evidence available on the differences in molecular epidemiology between ESBL-EC and non-ESBL-EC isolates from urine in China.
The present study provides data on the clonal distribution, virulence genes and phylogenetic groups distribution displayed by fifty ESBL-EC and fifty non-ESBL-EC strains.
Materials and Methods
Bacterial Isolates
A total of 100 non-duplicate isolates (one isolate per patient) randomly selected from the patients suffered UTI during 2022 in The First Affiliated Hospital of Ningbo University. These collections of isolates all came from urine and were identified by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) (BioMérieux, Marcy l’Etoile, FRANCE). And a random selection among them for that fifty isolates were positive for beta-lactamase production and fifty were negative.
Phenotypic Detection of ESBLs in Escherichia coli
The difference of phenotype between ESBL-EC and non-ESBL-EC was were detected by use of the double-disk synergy test in accordance with the phenotypic confirmation test recommended by Clinical and Laboratory Standards Institute (CLSI, 2019) (https://www.clsi.org). The confirmatory analysis for ESBL was determined as the zone of inhibition of cefotaxime and/or ceftazidime to ≥5mm from that of cefotaxime/clavulanicacid and/or ceftazidime/clavulanic acid. Klebsiella pneumoniae ATCC 700603 and Escherichia coli ATCC 25922 were used as positive and negative control strains for ESBLs detection, respectively.
Detection of Phylogenetic Group
Whole-cell DNA of uropathogenic Escherichia coli (UPEC) colonies was extracted by boiling lysis method at 90°C for 10 minutes. Polymerase Chain Reactions were performed to detect the gadA, chuA, and yjaA genes for the classification of main phylogenetic groups (A, B1, B2, D). All primers14 used in this study are shown in Table S1.
The PCR reaction volumes consisted of 0.5ul forward primer, 0.5ul reverse primer, 5ul of reaction mixture reagent (Yeasen Biotechnology Inc., Shanghai, China), 5ul of distilled water, and 1ul of DNA product extracted by boiling, for a total of 12ul. Thermal cycling was performed in GeneAmp® PCR System 9700 (Applied Biosystems Inc., Foster City, CA, USA) under the following conditions: denaturation for 5 minutes at 95°C; 30 amplification cycles of 1 minute at 95°C, 1 minute at 60°C, and 5 minutes at 72°C; and a final extension of 10 minutes at 72°C. Expected amplicon sizes for gadA, chuA, yjaA and TSPE4.C2 was 373-, 281-, 216- and 158-base-pair (bp) respectively. Finally, PCR products were subjected to electrophoresis and photography under ultraviolet trans-illumination.
The results of PCRs that chuA–TSPE4.C2+, chuA+yjaA+, chuA–TSPE4.C2–, chuA+yjaA– were identified to group B1, B2, A and D respectively. The function of the gadA gene was a control for internal amplification.
Multilocus Sequence Typing
Based on a previously standardized protocol about multilocus sequence typing (MLST) of E. coli,15 seven housekeeping genes were analyzed from 100 strains of UPEC by PCR reactions. And the amplification products were sequenced using the BigDye Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems Inc., Foster City, CA, USA) with forward primer. The sequencing results were submitted to a web database (http://mlst.ucc.ie/mlst/dbs/Ecoli) and compared on line to obtain the allelic profile so that the result sequence type were obtained. The PCR amplification settings and materials were identical to those described above for the phylogenetic group assay and the primer sequences utilized for MLST were listed in Table S1.
Detection of Virulence Genes
In this work, 19 virulence genes were analyzed by PCR to investigate the adhesion, protection, toxin, and iron uptake-related capabilities of UPEC. The virulence gene count was calculated by summing the number of virulence genes found in each isolate. The primers of detecting virulence genes were listed in Table S2.
Statistical Analysis
The data were analyzed by using the one-way ANOVA test and the non-parametric Mann Whitney test with SPSS for Windows version 21.0 (SPSS Inc., Chicago, IL, USA). P values <0.05 were considered statistically significant.
Results
Detection of Phylogenetic Groups
The main phylogenetic group of the two groups was B2 (52%-non-ESBL-EC vs 56%-ESBL-EC) came after other three groups: B1 (12% vs 14%), D (14% vs 28%), and A (22% vs 2%). Besides, group D was found to be more prevalent in the non-ESBL-EC group, and group A in the ESBL-EC was more common. The differences were statistically significant (Figure 1).
 |
Figure 1 Comparison of the distribution of phylogenetic groups between non-ESBL-EC and ESBL-EC. Abbreviations: ESBL, extended-spectrum beta-lactamase; ESBL-EC, ESBL-producingE. coli; non-ESBL-EC, non-ESBL-producingE. coli; phylogroup, phylogenetic group. |
Sequence Types
Forty-nine STs were identified among the 100 studied isolates, 68% of the ESBL-EC group isolates corresponded to the following STs: ST1193 (24%), ST131 (14%), ST648 (8%), ST38 (8%), ST69 (6%), ST75 (4%) and ST1011 (4%). And 48% of the non ESBL-EC group isolates were ST73 (20%), ST95 (14%), ST216 (6%), ST409 (4%) and ST58 (4%). The main sequence types of the two groups were totally different (Figure 2).
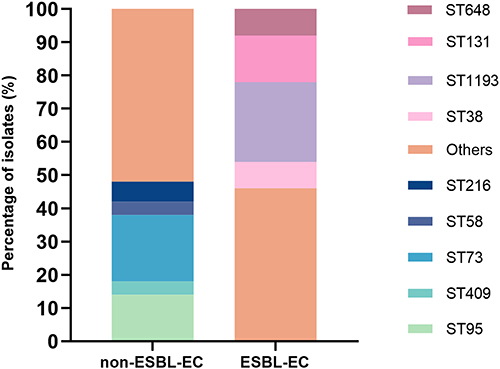 |
Figure 2 Comparison of the distribution of sequence types between non-ESBL-EC and ESBL-EC. Others, STs that occurred <5% with non-ESBL-EC or ESBL-EC were categorized as “others”. Abbreviations: ESBL, extended-spectrum beta-lactamase; ESBL-EC, ESBL-producingE. coli; non-ESBL-EC, non-ESBL-producingE. coli; ST, sequence type. |
Virulence Genes
Of the 19 virulence genes were detected, the virulence genes (VG) count ranged from 2 to 16. The median of VG among non-ESBL-EC (11, IQR 6–15) were higher compared to ESBL-EC (10, IQR 8–12) (Figure 3). From the graph (Figure 4), VG count varied between different phylogenetic groups for that the highest in group B2 and the lowest in group A. Specifically, the average of VG belonging to group B2, B1, A and D were 12.35, 7.31, 6.58 and 8.57, respectively. The distribution of virulence genes in group B2 differed significantly from that in groups B1, A, and D, respectively (p< 0.001). What ‘s more, ESBL-negative ST73 (median 15, IQR 14–15) and ST95 (median 13, IQR 10–16) isolates had higher average VG count and ESBL-positive ST 38 isolates had the lowest count (median 8, IQR 7.5–8) (Figure 5). Additionally, in group B2 which most strains belonged to, average VG count of non-ESBL-EC was higher than ESBL-EC and the difference between the B2 group made a great sense (Figure 6).
 |
Figure 3 Comparison of the total number of VG detected per isolates between non-ESBL-EC and ESBL-EC. Every dot represents a single isolate. Abbreviations: ESBL, extended-spectrum beta-lactamase; ESBL-EC, ESBL-producingE. coli; non-ESBL-EC, non-ESBL-producingE. coli; VG count, the number of virulence genes. |
 |
Figure 4 Comparison of the distribution of virulence factors in four main phylogenetic groups. B2, phylogenetic group B2; B1, phylogenetic group B1; A, phylogenetic group A; D, phylogenetic group D; VG count, the number of virulence genes; every dot represents a single isolate; p value of comparison about the distribution of virulence factors in four main phylogenetic groups, calculated with analysis of ANOVA, one-way analysis of variance, SPSS 21.0. P <0.0001 (marked as ****) was defined as extremely significant. |
 |
Figure 5 UPEC-associated VG count in different STs, stratified for ESBL-positivitya. aESBL-positivity based on the phenotypic ESBL production; Boxplots display median VG count and inter quartile range (IQR); every dot represents a single isolate; only STs that occurred >5% with non-ESBL-EC or ESBL-EC were grouped into main groups, the rest were categorized as “Others”. Abbreviations: ESBL, extended-spectrum beta-lactamase; ESBL-EC, ESBL-producingE. coli; non-ESBL-EC, non-ESBL-producingE. coli; ST, sequence type; VG, virulence gene. |
 |
Figure 6 Comparison of acquired virulence genes count per phylogenetic group. Every dot represents a single isolate; B2, phylogenetic group B2; B1, phylogenetic group B1; A, phylogenetic group A; D, phylogenetic group D. P <0.001 (marked as***) was defined as extremely significant. Abbreviations: VG count, the number of virulence genes; ESBL, extended-spectrum beta-lactamase; ESBL-EC, ESBL-producingE. coli; non-ESBL-EC, non-ESBL-producingE. coli. |
There was an inverse correlation between β-lactamase producing and the number of VG count. Wherein, eleven types of virulence genes were more prevalent in isolates from non-ESBL-EC group. Statistical significance was observed in the genes for hlyA, papC, papGII, cnf1, iroN, vat and sfa (p<0.05) (Table 1).
 |
Table 1 Virulence Genes Distribution Between ESBL-EC and Non-ESBL-EC Group of UPEC Isolates |
Discussion
In this study, we have noted discernible distinctions with regard to clonal distribution, virulence gene count, and phylogenetic group between ESBL-EC and non-ESBL-EC strains that were isolated from urine.
Compared to prior analogous investigations on E. coli isolates from bloodstream16–18 the clonal distribution exhibited by ESBL-EC urine isolates stands in stark contrast to their non-ESBL-EC counterparts. As per a recent investigation, the two most prevalent sequence types of E. coli urine isolates tested for the presence of ESBL genes via PCR were ST131 and ST1193,19 a result that aligns with our own research findings. The observed discrepancy in clonal distribution between ESBL-positive and non-ESBL E. coli urine isolates may be primarily attributed to the presence of multidrug resistance (MDR) in a subset of the former isolates. In recent times, the ST131 clone has emerged as the foremost and highly potent multi-drug resistant (MDR) clone on a global scale, serving as a major etiological factor for community-acquired urinary and bacteremic infections.20,21 ST1193 clone follows closely due to its prevalent overuse of Fluoroquinolone (FQ) and third-generation cephalosporin.22 Birgy et al23 revealed that ST1193 constituted one of the most prevalent clones during the final study phase, among 218 ESBL-producing E. coli infections that resulted in febrile urinary tract infections in children between 2014 and 2017.
In our study, ST73 and ST95 were absent in ESBL-EC group, and it is established that ST73 is vulnerable to antibiotics.24 The ST distribution phenomena shows that in UPEC, the typically antimicrobial-susceptible STs: ST73 and ST95 have been replaced by strong antibiotic pressure. Most striking was that ST73 and ST95 which were both negative in producing extended beta-lactamase had the highest VG content among other STs. Besides, the average of VG content related to group B2 of ESBL-EC was also lower than non-ESBL-EC. The result ties well with a former study performed by Horcajada et al.25 These findings were unexpected and reflect the inverse association previously described26 similarly between antimicrobial resistance and VG content. As for the other A, B1 and D groups poor in inverse association, we speculate that the limited strains is responsible for this result. Notably, the four primary sequence classifications identified in this investigation, namely ST131, ST1193, ST73, and ST95, happen to coincide with the sequence types previously demonstrated to disseminate from UTI to E. coli Bacteremia.19 Such a coincidence may be due to the phenomenon that the aforementioned prevailing clones were renowned UPEC clones.27
Studies undertaken in South Korea,28 Ethiopia,29 and Mexico30 revealed similar distributions, which are consistent with the majority of findings on the phylogenetic grouping of UPEC. These studies have shown that the phylogenetic group B2 primarily encompasses the bulk of E. coli isolates. In this study, we have also discovered that B2 was prevalent in both ESBL-EC and non-ESBL-EC patients. This emphasizes the need of addressing the appearance of isolates linked to this group. The information shown in Figure 4 supports this and indicates that group B2 demonstrates a high number of virulence determinants. Genetically stable isolates with much lower recombination rates make up the commensal assemblages of E. coli. This trait gives the population a clonal structure and allows the major phylogenetic groups to be classified.31 Commensal E. coli isolates make up the majority of phylogenetic group A, in light of numerous past investigations. This observation remains true for both human commensal E. coli and UPEC strains, based on the reports of Duriez et al32 and Khairy et al,33 where phylogenetic group A accounts for the most significant fraction of phylogenetic groups. Depending on several reports, phylogenetic group A is the most prevalent phylogenetic group among UPEC isolates.33–36 According to our research, almost 22% of non-ESBL-EC isolates fell into phylogenetic group A, which is much higher than the similar number reported in earlier studies carried out in South Korea (3.44%).28 The aforementioned finding suggests that the gastrointestinal system is the primary source of ESBL negative strains that colonize the urinary tracts since phylogenetic group A, which is normally associated with commensal infections, is prevalent among UPEC isolates.37
Additionally, these phylogenetic groupings have a noteworthy characteristic as a result of their comparatively increased quantity of virulence variables, making them extremely virulent clinical isolates and complicating efforts to cure them. Compared to other phylogenetic groups, the group B2 showed a significantly higher prevalence of virulence genes. Specifically, the isolates belonging to group B2 in non-ESBL-EC group harbored more virulence genes. One plausible justification for this observation could be the co-localization of the extended-spectrum beta-lactamase-producing gene and certain virulence genes on the same plasmids or chromosomes. This is consistent with the findings of earlier research from Denmark,38 Pakistan, Ethiopia,39 Mexico,30 and Poland.40
This study focused on the clonal distribution, phylogenetic group, and virulence profiling of UPEC isolates based on their beta-lactamase production status. According to the findings above, more attention should be paid not only to phylogenetic group B2 that most isolates belonged to, but also non-β-lactamase producing strains harboring high virulence genes, which will lead to high pathogenicity.
It is noteworthy to mention that a limited number of E. coli isolates were selectively obtained from a singular tertiary-care hospital. Consequently, the findings are not generalizable to all UPEC isolates. Accordingly, it is our hope to conduct a wide-ranging molecular epidemiological study utilizing a multicenter approach in future endeavors.
In conclusion, our research indicates that there is relevant evidence between the molecular characteristics of E. coli isolates and the phenotype of beta-lactamase synthesis. Here, we found significant differences in the clonal distribution, virulence gene content, and phylogenetic groups of non-ESBL-EC and ESBL-EC urine isolates that outweigh their phenotypic ESBL-positivity. To avoid attributing resistant E. coli features to the overall E. coli population, future multicenter studies attempting to explain the molecular epidemiology of UTI should instead focus on E. coli without ESBL-positivity.
Conclusion
Non-ESBL-EC and ESBL-EC isolates from The First Affiliated Hospital of Ningbo University differed in clonal distribution, phylogenetic group and VG content. The inverse association between beta-lactamase resistance and VG content should get a lot more attention. At the same time, we should also be wary of the appearance of non-ESBL-EC isolates belonging to group B2.
Ethics Approval and Consent to Participate
Our research was reviewed and approved by Medical Ethics Committee of The First Affiliated Hospital of Ningbo University (Approval NO:2023053RS). The patient who is going to be hospitalized will sign an informed consent which implies that biological samples taken may be used for medical teaching and research. In this study, the sample were obtained from inpatients who freely and voluntarily signed the informed consent under the full explanation of the clinician. And for the patients who were actively screened, oral consent was also obtained.
Acknowledgments
The authors would like to thank the staff of the Laboratory Department at The First Affiliated Hospital of Ningbo University for their devoted technical support. The study was approved by the Ethics Committee of the First Affiliated Hospital of Ningbo University.
Disclosure
The authors declare that there is no conflict of interest in the publication of this paper.
References
1. Foxman B. The epidemiology of urinary tract infection. Nat Rev Urol. 2010;7(12):653–660. doi:10.1038/nrurol.2010.190
2. Flores-Mireles AL, Walker JN, Caparon M, Hultgren SJ. Urinary tract infections: epidemiology, mechanisms of infection and treatment options. Nat Rev Microbiol. 2015;13(5):269–284. doi:10.1038/nrmicro3432
3. Mazzariol A, Bazaj A, Cornaglia G. Multi-drug-resistant Gram-negative bacteria causing urinary tract infections: a review. J Chemother. 2017;29(sup1):2–9. doi:10.1080/1120009X.2017.1380395
4. Bader MS, Loeb M, Leto D, Brooks AA. Treatment of urinary tract infections in the era of antimicrobial resistance and new antimicrobial agents. Postgrad Med. 2020;132(3):234–250. doi:10.1080/00325481.2019.1680052
5. McDanel J, Schweizer M, Crabb V, et al. Incidence of Extended-Spectrum β-Lactamase (ESBL)-producing Escherichia coli and Klebsiella infections in the United States: a systematic literature review. Infect Contr Hospital Epidemiol. 2017;38(10):1209–1215. doi:10.1017/ice.2017.156
6. Tansarli GS, Poulikakos P, Kapaskelis A, Falagas ME. Proportion of extended-spectrum beta-lactamase (ESBL)-producing isolates among Enterobacteriaceae in Africa: evaluation of the evidence--systematic review. J Antimicrob Chemother. 2014;69(5):1177–1184. doi:10.1093/jac/dkt500
7. Yamasaki S, Le TD, Vien MQ, Van Dang C, Yamamoto Y. Prevalence of extended-spectrum β-lactamase-producing Escherichia coli and residual antimicrobials in the environment in Vietnam. Anim Health Res Rev. 2017;18(2):128–135. doi:10.1017/S1466252317000160
8. Hayajneh WA, Hajj A, Hulliel F, et al. Susceptibility trends and molecular characterization of Gram-negative bacilli associated with urinary tract and intra-abdominal infections in Jordan and Lebanon: SMART 2011-2013. Int J Infect Dis. 2015;35:56–61. doi:10.1016/j.ijid.2015.04.011
9. Al-Jamei SA, Albsoul AY, Bakri FG, Al-Bakri AG. Extended-spectrum β-lactamase producing E. coli in urinary tract infections: a two-center, cross-sectional study of prevalence, genotypes and risk factors in Amman, Jordan. J Infect Public Health. 2019;12(1):21–25. doi:10.1016/j.jiph.2018.07.011
10. Jia P, Zhu Y, Li X, et al. High prevalence of extended-spectrum beta-lactamases in Escherichia coli strains collected from strictly defined community-acquired urinary tract infections in adults in China: a multicenter prospective clinical microbiological and molecular study. Front Microbiol. 2021;12:663033. doi:10.3389/fmicb.2021.663033
11. Pilmis B, Parize P, Zahar JR, Lortholary O. Alternatives to carbapenems for infections caused by ESBL-producing Enterobacteriaceae. Eur J Clin Microbiol Infect Dis. 2014;33(8):1263–1265. doi:10.1007/s10096-014-2094-y
12. Liu X, Cui D, Li H, et al. Direct medical burden of antimicrobial-resistant healthcare-associated infections: empirical evidence from China. J Hosp Infect. 2020;105(2):295–305. doi:10.1016/j.jhin.2020.01.003
13. Huttner A, Hatz C, van den Dobbelsteen G, et al. Safety, immunogenicity, and preliminary clinical efficacy of a vaccine against extraintestinal pathogenic Escherichia coli in women with a history of recurrent urinary tract infection: a randomised, single-blind, placebo-controlled phase 1b trial. Lancet Infect Dis. 2017;17(5):528–537. doi:10.1016/S1473-3099(17)30108-1
14. Yun KW, Kim DS, Kim W, Lim IS. Molecular typing of uropathogenic Escherichia coli isolated from Korean children with urinary tract infection. Korean J Pediatr. 2015;58(1):20–27. doi:10.3345/kjp.2015.58.1.20
15. Tartof SY, Solberg OD, Manges AR, Riley LW. Analysis of a uropathogenic Escherichia coli clonal group by multilocus sequence typing. J Clin Microbiol. 2005;43(12):5860–5864. doi:10.1128/JCM.43.12.5860-5864.2005
16. Mathers AJ, Peirano G, Pitout JD. The role of epidemic resistance plasmids and international high-risk clones in the spread of multidrug-resistant Enterobacteriaceae. Clin Microbiol Rev. 2015;28(3):565–591. doi:10.1128/CMR.00116-14
17. Hertz FB, Nielsen JB, Schønning K, et al. ”Population structure of drug-susceptible,-resistant and ESBL-producing Escherichia coli from community-acquired urinary tract”. BMC Microbiol. 2016;16:63. doi:10.1186/s12866-016-0681-z
18. Manges AR, Geum HM, Guo A, Edens TJ, Fibke CD, Pitout JDD. Global extraintestinal pathogenic Escherichia coli (ExPEC) lineages. Clin Microbiol Rev. 2019;32(3). doi:10.1128/CMR.00135-18
19. Choi HJ, Jeong SH, Shin KS, et al. Characteristics of Escherichia coli urine isolates and risk factors for secondary bloodstream infections in patients with urinary tract infections. Microbiol Spectr. 2022;10(4):e0166022. doi:10.1128/spectrum.01660-22
20. Jafari A, Falahatkar S, Delpasand K, Sabati H, Sedigh Ebrahim-Saraie H. Emergence of Escherichia coli ST131 causing urinary tract infection in western Asia: a systematic review and meta-analysis. Microb Drug Resist. 2020;26(11):1357–1364. doi:10.1089/mdr.2019.0312
21. Xiao S, Tang C, Zeng Q, et al. Antimicrobial resistance and molecular epidemiology of Escherichia coli from bloodstream infection in Shanghai, China, 2016–2019. Front Med. 2022;8. doi:10.3389/fmed.2021.803837
22. Fukushima Y, Sato T, Tsukamoto N, et al. Clonal/subclonal changes and accumulation of CTX-M-type β-lactamase genes in fluoroquinolone-resistant Escherichia coli ST131 and ST1193 strains isolated during the past 12 years, Japan. J Global Antimicrob Resist. 2021;27:150–155. doi:10.1016/j.jgar.2021.08.015
23. Birgy A, Madhi F, Jung C, et al. Diversity and trends in population structure of ESBL-producing Enterobacteriaceae in febrile urinary tract infections in children in France from 2014 to 2017. J Antimicrob Chemother. 2020;75(1):96–105. doi:10.1093/jac/dkz423
24. Kallonen T, Brodrick HJ, Harris SR, et al. Systematic longitudinal survey of invasive Escherichia coli in England demonstrates a stable population structure only transiently disturbed by the emergence of ST131. Genome Res. 2017;27(8):1437–1449. doi:10.1101/gr.216606.116
25. Horcajada JP, Soto S, Gajewski A, et al. Quinolone-resistant uropathogenic Escherichia coli strains from phylogenetic group B2 have fewer virulence factors than their susceptible counterparts. J Clin Microbiol. 2005;43(6):2962–2964. doi:10.1128/JCM.43.6.2962-2964.2005
26. Moreno E, Prats G, Sabaté M, Pérez T, Johnson JR, Andreu A. Quinolone, fluoroquinolone and trimethoprim/sulfamethoxazole resistance in relation to virulence determinants and phylogenetic background among uropathogenic Escherichia coli. J Antimicrob Chemother. 2006;57(2):204–211. doi:10.1093/jac/dki468
27. Matsukawa M, Igarashi M, Watanabe H, et al. Epidemiology and genotypic characterisation of dissemination patterns of uropathogenic Escherichia coli in a community. Epidemiol Infect. 2019;147:e148. doi:10.1017/S0950268819000426
28. Lee JH, Subhadra B, Son YJ, et al. Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea. Lett Appl Microbiol. 2016;62(1):84–90. doi:10.1111/lam.12517
29. Dadi BR, Abebe T, Zhang L, Mihret A, Abebe W, Amogne W. Distribution of virulence genes and phylogenetics of uropathogenic Escherichia coli among urinary tract infection patients in Addis Ababa, Ethiopia. BMC Infect Dis. 2020;20(1):108. doi:10.1186/s12879-020-4844-z
30. Paniagua-Contreras GL, Monroy-Pérez E, Rodríguez-Moctezuma JR, Domínguez-Trejo P, Vaca-Paniagua F, Vaca S. Virulence factors, antibiotic resistance phenotypes and O-serogroups of Escherichia coli strains isolated from community-acquired urinary tract infection patients in Mexico. J Microbiol Immunol Infect. 2017;50(4):478–485. doi:10.1016/j.jmii.2015.08.005
31. Stoppe NC, Silva JS, Carlos C, et al. Worldwide Phylogenetic Group Patterns of Escherichia coli from commensal human and wastewater treatment plant isolates. Front Microbiol. 2017;8:2512. doi:10.3389/fmicb.2017.02512
32. Duriez P, Clermont O, Bonacorsi S, et al. Commensal Escherichia coli isolates are phylogenetically distributed among geographically distinct human populations. Microbiology. 2001;147(Pt 6):1671–1676. doi:10.1099/00221287-147-6-1671
33. Khairy RM, Mohamed ES, Abdel Ghany HM, Abdelrahim SS. Phylogenic classification and virulence genes profiles of uropathogenic E. coli and diarrhegenic E. coli strains isolated from community acquired infections. PLoS One. 2019;14(9):e0222441. doi:10.1371/journal.pone.0222441
34. Grude N, Potaturkina-Nesterova NI, Jenkins A, et al. A comparison of phylogenetic group, virulence factors and antibiotic resistance in Russian and Norwegian isolates of Escherichia coli from urinary tract infection. Clin Microbiol Infect. 2007;13(2):208–211. doi:10.1111/j.1469-0691.2006.01584.x
35. Ramos NL, Dzung DT, Stopsack K, et al. Characterisation of uropathogenic Escherichia coli from children with urinary tract infection in different countries. Eur J Clin Microbiol Infect Dis. 2011;30(12):1587–1593. doi:10.1007/s10096-011-1264-4
36. Derakhshandeh A, Firouzi R, Motamedifar M, et al. Virulence characteristics and antibiotic resistance patterns among various phylogenetic groups of uropathogenic Escherichia coli isolates. Jpn J Infect Dis. 2015;68(5):428–431. doi:10.7883/yoken.JJID.2014.327
37. Moreno E, Andreu A, Pérez T, Sabaté M, Johnson JR, Prats G. Relationship between Escherichia coli strains causing urinary tract infection in women and the dominant faecal flora of the same hosts. Epidemiol Infect. 2006;134(5):1015–1023. doi:10.1017/S0950268806005917
38. Ejrnæs K, Stegger M, Reisner A, et al. Characteristics of Escherichia coli causing persistence or relapse of urinary tract infections: phylogenetic groups, virulence factors and biofilm formation. Virulence. 2011;2(6):528–537. doi:10.4161/viru.2.6.18189
39. Bashir S, Haque A, Sarwar Y, Ali A, Anwar MI. Virulence profile of different phylogenetic groups of locally isolated community acquired uropathogenic E. coli from Faisalabad region of Pakistan. Ann Clin Microbiol Antimicrob. 2012;11:23. doi:10.1186/1476-0711-11-23
40. Kot B, Wicha J, Grużewska A, Piechota M, Wolska K, Obrębska M. Virulence factors, biofilm-forming ability, and antimicrobial resistance of urinary Escherichia coli strains isolated from hospitalized patients. Turkish J Med Sci. 2016;46(6):1908–1914. doi:10.3906/sag-1508-105
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.