Back to Journals » Clinical Epidemiology » Volume 6
Epidemiology of visceral leishmaniasis
Authors Ready P
Received 14 January 2014
Accepted for publication 4 March 2014
Published 3 May 2014 Volume 2014:6 Pages 147—154
DOI https://doi.org/10.2147/CLEP.S44267
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Paul D Ready
Disease Control Department, Faculty of Infectious and Tropical Diseases, London School of Hygiene and Tropical Medicine, London, UK
Abstract: Leishmania species are the causative agents of leishmaniasis, a neglected tropical disease. These parasitic protozoans are usually transmitted between vertebrate hosts by the bite of blood sucking female phlebotomine sand flies. This review focuses on the two parasites causing most human visceral leishmaniasis (VL), which leads to substantial health problems or death for up to 400,000 people per year. Except for travel cases, Leishmania donovani infections are restricted to the (sub-)tropics of Asia and Africa, where transmission is mostly anthroponotic, while Leishmania infantum occurs in the drier parts of Latin America as well as in the Mediterranean climate regions of the Old World, with the domestic dog serving as the main reservoir host. The prevalence of VL caused by L. infantum has been declining where living standards have improved. In contrast, infections of L. donovani continue to cause VL epidemics in rural areas on the Indian subcontinent and in East Africa. The current review compares and contrasts these continental differences and suggests priorities for basic and applied research that might improve VL control. Transmission cycles, pathogenesis, diagnosis, treatment and prognosis, prevention (including vector control), surveillance, transmission modeling, and international control efforts are all reviewed. Most case detection is passive, and so routine surveillance does not usually permit accurate assessments of any changes in the incidence of VL. Also, it is not usually possible to estimate the human inoculation rate of parasites by the sand fly vectors because of the limitations of survey methods. Consequently, transmission modeling rarely passes beyond the proof of principle stage, and yet it is required to help develop risk factor analysis for control programs. Anthroponotic VL should be susceptible to elimination by rapid case detection and treatment combined with local vector control, and one of the most important interventions may well be socioeconomic development.
Keywords: diagnosis, Leishmania donovani, Leishmania infantum, surveillance, transmission control, treatment
Introduction
Parasites, vectors, and disease forms
Species of the genus Leishmania (Kinetoplastida, Trypanosomatidae) are the causative organisms of leishmaniasis or leishmaniosis,1 and these parasitic unicellular protozoans are usually transmitted between vertebrate hosts by the bite of blood sucking female phlebotomine sand flies (Diptera, Psychodidae).2 Parasites of the subgenus Sauroleishmania are considered to be restricted to lizards, and most lizard-feeding sand flies do not usually bite humans. About 20 species of Leishmania infect mammals and many of them can cause human leishmaniasis. Motile infective forms of the parasite (metacyclic promastigotes with a long free flagellum) develop in the guts of competent sand fly vectors, which inoculate them into mammalian skin. There they are ingested by cells of the mononuclear phagocyte system (formerly the reticuloendothelial system), transform into rounded amastigote forms with the loss of the free flagellum, and then can multiply in the phagolysosomes of recruited macrophages to cause a primary lesion at the site of the sand fly bite. Infections can spread, often via the lymphatic system, to cause secondary dermal lesions with forms and tissue tropisms in humans that show some parasite species specificity. Human cutaneous leishmaniasis is caused by most Leishmania species in the subgenus Leishmania, which occur in most subtropical and tropical regions (for example Leishmania (Leishmania) major from Africa and Asia, and Leishmania (Leishmania) mexicana from Central and South America), and by many species in the subgenus Viannia, which are restricted to Latin America (for example Leishmania (Viannia) brasiliensis). Any parasite causing cutaneous leishmaniasis can visceralize (for example Leishmania (Leishmania) tropica, which normally causes Oriental sore), but only two species of the subgenus Leishmania routinely do so, and these are the causative agents of most human visceral leishmaniasis (VL) worldwide.
Objectives
This review focuses on the two most important causative agents of VL, namely Leishmania (Leishmania) donovani and Leishmania (Leishmania) infantum, the ones responsible for causing substantial health problems in up to 400,000 people and up to 40,000 deaths per year.1,3 They are not always well distinguished in the medical literature, being closely-related members of the L. donovani species complex,4,5 but this disguises some important epidemiological differences. Except for travel cases,1 L. donovani is found only in the Old World, where it is notoriously associated with VL (kala-azar or black death) of the rural poor in the northeast of the Indian subcontinent and with VL of displaced persons in East Africa.1,6 Most transmission is believed to be anthroponotic, from human to human, and this contrasts with the zoonotic transmission of L. infantum, from canine reservoir hosts to humans, not only in the Mediterranean region where it may have originated, but also in many of the drier regions of Latin America where it was probably introduced historically.5
The incidence of VL associated with the transmission of L. infantum has been declining in many foci where living standards have improved; but, in contrast, VL epidemics continue to kill thousands of people infected with L. donovani on the Indian subcontinent and in East Africa. Therefore, the main aims of the current review are to compare and contrast these continental differences, and to suggest priorities for basic and applied research that might improve VL control. Leishmaniasis is a neglected tropical disease,1,6 and it is possible that the recent increases in the direct or indirect funding of applied research will not be sustained unless research priorities are well founded.1,2
Methods: literature search
This review is based on expert knowledge, wide-ranging reviews published in 2010–2013,1,2,6 and other, sometimes more recent, literature found in PubMed by using the search terms “visceral leishmaniasis”, “kala-azar”, “Leishmania donovani”, “Leishmania infantum”, and key words in the following section titles.
Transmission cycles: geographical variation in eco-epidemiology
The key elements are set out in Table 1. VL caused by L. donovani is usually considered to be an anthroponosis,2 but this should be challenged where control fails. Zoonotic transmission has been suspected because parasites or circulating antibodies have been detected in domestic animals on the Indian subcontinent7 and in the mongoose and other wild mammals in East Africa.1 The landscape epidemiology is very different on the two continents.2 Phlebotomus (Euphlebotomus) argentipes is the vector in humid (sub-)tropical regions on the Indian subcontinent, where it is attracted to domestic cattle and water buffalo kept in and around rural villages near the flood plain of the River Ganges, in Sri Lanka and perhaps in Bhutan.8 In contrast, Phlebotomus (Larroussius) orientalis and Phlebotomus (Synphlebotomus) species are the incriminated vectors in two distinctive bioclimatic regions of East Africa,2,9 which share a savanna landscape and transmission habitats both inside and outside rural villages. Most foci on the Indian subcontinent are in long-established villages with sedentary populations, whereas migratory populations are also at high risk in East Africa, including cattle herdsmen and villagers displaced by drought and warfare. Post-kala-azar dermal leishmaniasis (PKDL) is characterized by lesions rich in parasites, and these might favor anthroponotic transmission in Africa as well as in India.3
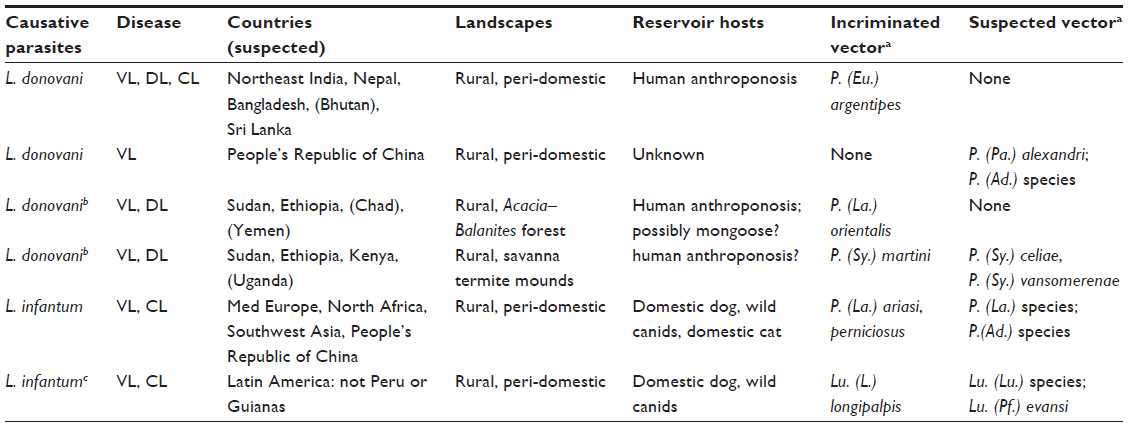 | Table 1 Disease types and transmission cycles of visceral leishmaniasis worldwide |
VL caused by L. infantum is usually considered to be a zoonosis, although congenital infections have been reported even in Europe.10 Most foci in the Old World have a Mediterranean climate and sand fly vectors that can hibernate – usually Phlebotomus (Larroussius) species – and so there is a risk of the disease spreading from southern to northern Europe with climate change.2 All vectors of Leishmania in the New World are Lutzomyia species, but only one of these is believed to be an important widespread vector of L. infantum. Diverse populations of Lutzomyia (Lutzomyia) longipalpis are abundant peridomestically in the yards of rural villages and the low-rise suburbs and shanty towns of many of the drier tropical regions of Latin America, and, unlike most sand fly species, they are competent to transmit a wide range of Leishmania species2 including L. infantum. This parasite is believed to have been introduced in domestic dogs from Portugal and Spain during the last 500 years.5
Molecular taxonomy has not identified Leishmania archibaldi as a species separate from L. donovani in Africa or Leishmania chagasi as a species separate from L. infantum in Latin America, and the diverse groupings of the strains of L. donovani and L. infantum identified by neutral (or nonadaptive) genetic markers4,5 have not been associated with phenotypes of medical importance. Similarly, there is little or no evidence that the diverse forms of the vectors P. argentipes and Lu. longipalpis, identified as cryptic sibling species based on neutral genetic markers, have distinctive epidemiological roles.2
Pathogenesis
The incubation period is usually from 2 weeks to 18 months,11 with gross inflammatory reactions within the viscera often developing 2–8 months after infection and any initial skin lesions, but VL symptoms can take years to appear.12 The disease is progressive and a symptomatic infection that is untreated is generally fatal,12 with a mortality rate of 75%–95%.11 Death usually occurs within 2 years, although spontaneous cures may occur.11 Parasites proliferate wherever there are cells of the mononuclear phagocyte system, most often in macrophages. These are most abundant in the spleen and liver and, consequently, infection leads to an enlargement of both of these organs.11–13 Bone marrow cells become infected, and patients develop pancytopenia (namely, depressed production of red blood cells, white cells, and platelets) and immunosuppression, making them susceptible to superinfections.11,13
PKDL or dermal leishmanoid11 is a condition in which dermal lesions can be heavily parasitized in a subset of patients successfully treated for VL, and it occurs mainly in India and Sudan in patients infected by L. donovani.13,14 They remain asymptomatic for months to years and then develop a progressive proliferation of parasites within the skin, giving rise to diffuse macular, maculopapular, or nodular lesions.13 Lesions appear anywhere on the body, but they often occur on the face.12
Diagnosis
Clinical symptoms and microscopic diagnosis
Most infections are diagnosed clinically. The patient has an irregular fever, anemia, and leukopenia; hepatosplenomegaly and bone marrow suppression are characteristic (see above); and HIV coinfections often produce atypical presentations.11–13 Rates of PKDL vary regionally, arising within 6 months in up to 50% of treated VL patients in parts of Sudan, but within 2–3 years in only approximately 5%–10% of patients in India.12–14
Parasites are usually identified to the genus Leishmania by light microscopy.11,12 The amastigote form is usually detected microscopically after Giemsa staining. Blood examinations are the easiest to perform, but frequently there are very few circulating parasitized cells in buffy coat films.11,12 Spleen aspirates are a rich source of parasites,11 but biopsy should be performed only by trained personnel because there is the risk of rupture or bleeding.12 Liver biopsy is safer, although finding amastigotes is certain only in very heavy infections.11,12 Bone marrow aspiration (from the sternum) is frequently used,11 but it is a specialized technique with lower sensitivity.12 Detection of parasites in nodular forms of PKDL is easier than in macular forms.13,14 Inoculation of biopsy materials into culture media or laboratory rodents can be sensitive but slow.1
Biochemical diagnosis
Until recently, isoenzyme analysis has been the gold-standard method to identify both phylogenetic species and strains (or zymodemes) of Leishmania, which usually reproduce asexually.1,5 However, it requires growing many parasites, and promastigote cultures can fail because of contaminating bacteria and fungi, or strain-specific temperature and nutrient requirements.
Serological diagnosis
Serological tests provide the most widely used indirect methods of diagnosis.1 The indirect fluorescence antibody test, the enzyme-linked immunosorbent assay, and the Western blot12 are widely used in Africa, Asia, Europe, and Latin America, but they all require equipment that has not been optimized for field conditions12 and there is often a lack of standardized protocols, antigens, and other reagents.15 The Montenegro or leishmanin skin test has the same drawbacks and may not even be specific to Leishmania species.1 It is negative in active kala-azar, and then usually becomes positive within 2 months after successful treatment.11
In contrast, two other types of tests have proved to have high sensitivity and specificity, at least regionally, and have translated to peripheral health centres.12,16 These are the direct agglutination test and immunochromatographic tests (ICTs) using the recombinant (r) K39 antigen, which is encoded by a fragment of a kinesin-related gene. The rK39 ICTs have the advantage of using convenient formats (often a dipstick) and a standardized recombinant antigen. However, the kinesin-related gene fragment derives from a Brazilian strain of L. infantum, and the kinesin genetic diversity among strains of L. donovani in East Africa and between there and Asia is extensive enough to influence the performance of the rK39 ICTs.17 Even in Brazil, the diagnostic performance of rK39 ICTs was found to be only reasonable for confirmation of infection in suspected cases of canine VL, and the sensitivity to detect infected dogs was too low for large-scale epidemiological studies and operational control programs.18
Serological tests have limitations because some antibodies are detectable years after cure.12 A latex agglutination test detecting a heat-stable carbohydrate in the urine of VL patients has promise as a field-adapted antigen test; it showed good specificity but low-moderate sensitivity.19
Molecular diagnosis
The polymerase chain reaction is more sensitive than microscopic examination and, therefore, it is often the primary test in referral hospitals and research centers, with quantitative polymerase chain reaction permitting accurate diagnosis using venous blood samples instead of bone marrow aspirates.12,16 Leishmania species and their strains causing VL can now be identified by the comparative DNA sequence analysis of five of the polymorphic metabolic enzymes routinely used for isoenzyme analysis,20 by restriction fragment length polymorphism analysis of the heat-shock 70 gene,21 and by microsatellite DNA analysis.4
Conclusion: integrated diagnosis
Molecular tests in hospitals and research centers permit strain identification and diagnosis using venous blood samples. However, many primary health teams rely on clinical symptoms together with rK39 ICTs for rapid diagnosis prior to chemotherapy. A combination of molecular and serological tests demonstrated significant numbers of asymptomatic infections in endemic areas of Bihar State, India.16 The selection of the best combinations of rapid diagnostic tests might be improved by meta-analyses of Phase IV studies performed in target populations, and Bayesian methods could improve estimates of sensitivity and specificity.22
Treatment and prognosis
Travel cases, indigenous cases, and health care
Few travel cases of VL are usually recorded in North America and Europe,1,9,23 and a recent review alerts clinicians to the range of clinical syndromes, treatments, and prognoses, including the development of PKDL seen in patients with HIV/AIDS undergoing highly active antiretroviral therapy (HAART).13 Treatment and prognosis for indigenous cases are determined by regional health care practices and budgets, with fewer uncomplicated cases being resolved in East Africa and India than in southern Europe and Brazil.1,6 Malnutrition is one of the important risk factors for VL, and a fall in disease incidence in southern Europe during the last 50 years coincided with better nutrition.1,6
Drug resistance and alternatives to pentavalent antimonials
Much research is focused on the resistance mechanism to pentavalent antimonials, which have been the standard drugs despite their toxicity.12 They are now nearly obsolete in Bihar State, India, because of drug resistance, but sodium stibogluconate (Pentostam [GlaxoSmithKline UK Ltd] and a generic brand) and meglumine antimoniate (Glucantime; Sanofi S.A., Paris, France) are useful elsewhere.
AmBisome (Gilead, Foster, CA, USA), a liposome formulation of amphotericin B, is now a standard treatment for VL, especially against L. donovani in Bihar, and a single dose treatment has been successfully trialed in rural public hospitals in Bangladesh (http://www.who.int/tdr/news/2013). However, clinical studies indicate that it is less effective against L. donovani in East Africa and L. infantum in Latin America and, therefore, further research is required.12 Similarly, paromomycin is efficacious in the Indian subcontinent, but not in East Africa.12 Miltefosine, a phospholipid derivative, provides an oral treatment for VL, and it has been used in the VL Elimination Programme for the Indian Subcontinent.12 However, it is potentially teratogenic, requires a long oral treatment (28 days) that can lead to poor compliance and relapses, and drug resistance is a concern.12
Novel drug combinations
Drug combinations can shorten therapy as well as reducing toxicities and the selection of resistant mutations, and so there have been some promising Phase III trials of combinations of AmBisome, miltefosine, and paromomycin in India and of sodium stibogluconate with paromomycin in Sudan.12 Barrett and Croft12 noted that new drugs might target parasites in the low pH phagolysosomes of macrophages.
Prevention
Vaccine development
Vaccination is probably the best way of controlling a rural vector-borne disease like leishmaniasis.1,2,24 Unfortunately, no vaccine is likely to be deployed in the short-term against VL, partly because of the complexity of the cellular immune response and the immunomodulatory effects of antigens from sand fly saliva.1,2 Salivary peptides could also be vaccine components.2,24 Vaccines against canine leishmaniasis have been trialed in Brazil and Europe with mixed results,1,10,18 and this could be an effective way of reducing the transmission of L. infantum to people.
Vector control
Unlike for mosquitoes, source reduction is not an option because the terrestrial sites of sand fly larvae are largely unknown. The spraying of residual insecticides, often dichlorodiphenyltrichloroethane (DDT), to control Anopheles mosquitoes was a key component of the malaria eradication campaign of the mid-20th century, and it is credited with having greatly depressed the transmission of L. donovani in India and L. infantum in southern Europe by the untargeted killing of peri-domestic adult sand flies.1,25 VL resurgence followed the cessation of DDT spraying, but this could have been coincidental. Since then, a range of vector control approaches have been tested, but there is little evidence that the results of experimental trials have informed public health approaches.
Indoor residual spraying with DDT or pyrethroids has been the mainstay of VL vector control on the Indian subcontinent and in Brazil,1,25,26 but it is unknown whether or not the insecticide reaches or remains for long on the surfaces favored by the adults of P. argentipes and Lu. longipalpis, many of which may spend more time on cattle or chickens, respectively, than on the inner or outer walls of houses and animal shelters.27,28 Insecticide-treated (mosquito) nets have been tested on the Indian subcontinent and in East Africa,8 occasionally as long-lasting insecticidal nets,26,29 and sometimes they have been shown to be better than indoor residual spraying for reducing the local abundance of vectors, as in Bangladesh.30 Little or no insecticide resistance has been detected in VL foci,1 and this is consistent with only a small fraction of the vector populations being exposed to insecticides.
Insecticidal sprays, roll-ons, and collars are widely used for protecting dogs in southern Europe and the wealthier areas of Latin America, but this may provide only individual protection, not the community-wide control that would reduce transmission from dogs to humans.27 N, N-diethyl-meta-toluamide (DEET) has been the repellent of choice for the individual protection of people, and cheaper natural products have started to be tested.2 Integrated control has occasionally identified avenues of applied research that should be further explored, including environmental modification by plastering the walls of village houses with mud or lime to reduce the number of sand fly resting sites in VL foci on the Indian subcontinent.25
Surveillance, transmission modeling, risk factors, and predictive modeling
Routine surveillance is usually too insensitive to assess accurately any changes in the prevalence or incidence of VL,15,27 with most case detection being passive.6 Also, it is not usually possible to estimate the human inoculation rate of parasites by the sand fly vectors2 because most sand fly surveys use light and sticky traps that do not target host-seeking female flies, and the reports often mention only the relative abundances of species without distinguishing males from females.25,26 Consequently, transmission modeling is not commonplace, although research projects have provided proofs of principle models for VL transmission in Asia31 and for canine leishmaniasis in Europe.32 Better transmission modeling is required to underpin risk factor analysis, which is more likely to be applied in control programs. Risk factor analysis has been carried out for foci on the Indian subcontinent,28 in Ethiopia,33 and in Brazil,27 frequently identifying poverty, malnutrition, and proximity of parasite or vector hosts to dwellings as high risk factors. Spatial modeling has usually only predicted the presence or absence of disease and vectors based on rather broad measures of climate and environmental variation,2,15,32 providing resolution that is little better than descriptive landscape epidemiology.
Prevalence and incidence
This topic is considered late in this review because it depends on an understanding of the different disease forms associated with geographically-isolated transmission cycles and regional differences in surveillance. The disease burden will strongly influence the regions chosen for sustainable integrated control of VL, which is discussed in the next section.
A major review of the incidence of leishmaniasis worldwide was led by the World Health Organisation (WHO) Leishmaniasis Control Programme, and the findings were published in 2012.6 This review concluded that leishmaniasis had often been ignored in discussions of tropical disease priorities because of the paucity of current incidence data. Disease burden is often expressed as disability-adjusted life years, but the accuracy of this measure depends on the reliability of the incidence, duration, severity, and mortality data for a condition, which are difficult to extrapolate from official data sources because of the focal distribution of leishmaniasis in remote locations.6 The 2012 review concluded that the majority of VL deaths are unrecorded, and case fatality rates may be 10%–20% even with treatment access. However, the authors did produce regional tables of reported and estimated VL incidence based on the expert opinions of national program managers and professionals;6 these are summarized in Table 2. Among the few published empirical assessments of under-reporting in surveillance data, two from Bihar, India, compared VL case numbers estimated by house-to-house surveys with official data, which were 4.2-fold and 8.1-fold lower.6
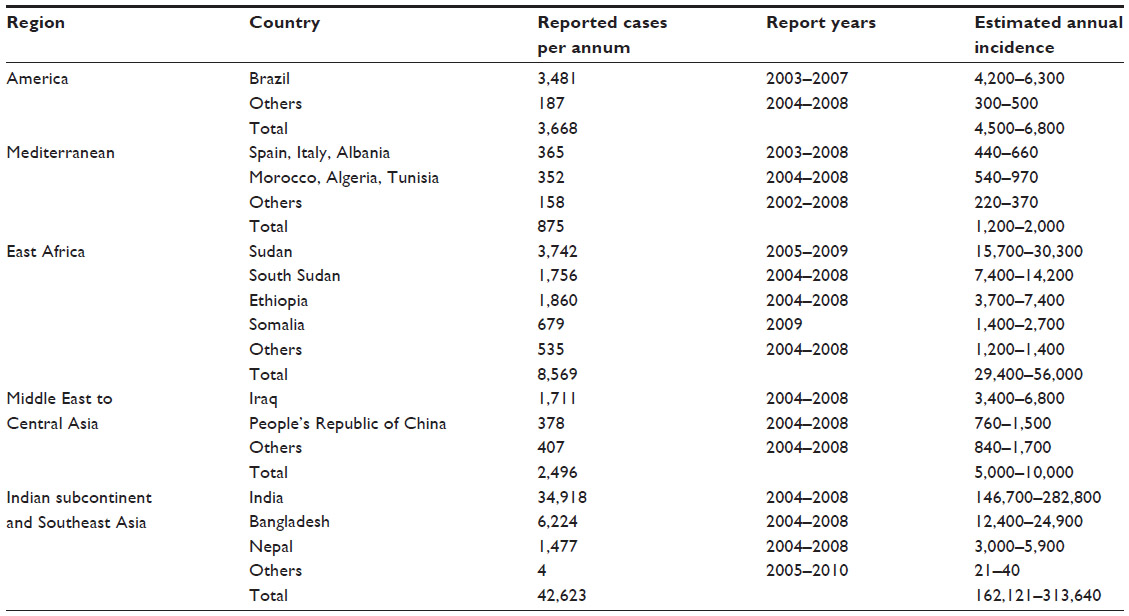 | Table 2 Reported and estimated incidence of VL worldwide |
The incidence of VL associated with the transmission of L. infantum by sand flies has been declining in many foci where living standards have improved, with better nutrition believed to mitigate the progression of debilitating or fatal infantile visceral leishmaniasis.6 Also, there has been a decline or stabilization in the incidence of VL resulting from the transmission of L. infantum by needles or blood transfusion through better awareness and drugs to combat coinfecting HIV.6
International collaboration for sustainable integrated control of VL
The paradigm for controlling vector-borne diseases should be prevention at source using an integrated set of intervention measures,34 not detection and response. There are many lessons to be learned from malaria control,35 including the realization that the aim of sand fly control should not be to reduce the relative abundance of a vector species, but to reduce the contact of its blood feeding females with humans and to reduce their lifespans so that fewer survive long enough to transmit Leishmania parasites.
The VL Elimination Programme for the Indian Subcontinent was set up as an international collaboration between Bangladesh, India, and Nepal in 2005.36 This required a clear set of aims and objectives, which prompted the standardization of experimental trials aided by European and North American researchers and funding agencies. There continues to be a need to assess by appropriate randomized trials the efficacy of some of the common intervention tools, including the residual activity of insecticides on bednets.37
Conclusion: priorities for applied and basic research
It has been argued that leishmaniasis research has focused too much on descriptive eco-epidemiology at the expense of transmission modeling and integrated control.2 For example, it does not really matter whether the causative agents of VL are classified by molecular taxonomists as two species (L. donovani and L. infantum) or as four species (with L. archibaldi and L. chagasi also recognized), if these taxa are not associated with specific sets of symptoms, treatments, and interventions in each endemic region. It is surely more important to understand what determines the distributions of parasite phenotypes of importance for treatment and control within each regional anthroponosis or zoonosis. Basic research on drug resistance and development underpins the applied aims of chemotherapy, but priorities are less clear for basic research for vector control and vaccine development. Predictive spatiotemporal modeling of VL should focus more on where and when disease incidence is likely to change, not on the presence or absence of species of parasites and vectors. A recent review of preventative measures against human leishmaniasis infection concluded that more research is needed to investigate human infection as the primary outcome measure, as opposed to intermediate surrogate markers.38
VL on the Indian subcontinent and in many African foci is believed to be an anthroponotic disease, and, therefore, it should be susceptible to elimination by rapid case detection and treatment combined with local vector control. However diagnosis of early infections can be problematic.6,17,19 The economic cost benefits of control are being explored on both continents,39,40 and one of the most important interventions against VL may well be socioeconomic development, as it is for malaria.41 Applied research on leishmaniasis must continue to be prioritized in order to ensure that it continues to benefit directly or indirectly from the support of the Bill and Melinda Gates Foundation (http://www.gatesfoundation.org), Drugs for Neglected Diseases initiative (http://www.dndi.org), Médecins Sans Frontières (http://www.msf.org), TDR-World Health Organization (http://www.who.int/tdr), and other bodies. The combat against VL might be assisted by the proposal to combine groups already working on chemotherapy into a single organization, the VL Global R&D and Access Initiative.42
Disclosure
The author reports no conflicts of interest in this work.
References
World Health Organization. Control of the leishmaniasis: report of a meeting of the WHO Expert Committee on the Control of Leishmaniases, Geneva, March 22–26, 2010. World Health Organ Tech Rep Ser. 2010;949:1–186. | |
Ready PD. Biology of phlebotomine sand flies as vectors of disease agents. Annu Rev Entomol. 2013;58:227–250. | |
Desjeux P, Ghosh RS, Dhalaria P, Strub-Wourgaft N, Zijlstra EE. Report of the Post Kala-azar Dermal Leishmaniasis (PKDL) Consortium Meeting, New Delhi, India, June 27–29, 2012. Parasit Vectors. 2013;6:196. | |
Gouzelou E, Haralambous C, Amro A, et al. Multilocus microsatellite typing (MLMT) of strains from Turkey and Cyprus reveals a novel monophyletic L. donovani sensu lato group. PLoS Negl Trop Dis. 2012;6(2):e1507. | |
Lukes J, Mauricio IL, Schönian G, et al. Evolutionary and geographical history of the Leishmania donovani complex with a revision of current taxonomy. Proc Natl Acad Sci U S A. 2007;104(22):9375–9380. | |
Alvar J, Vélez ID, Bern C, et al. Leishmaniasis worldwide and global estimates of its incidence. PLoS ONE. 2012;7:e35671. | |
Alam MS, Ghosh D, Khan MG, et al. Survey of domestic cattle for anti-Leishmania antibodies and Leishmania DNA in a visceral leishmaniasis endemic area of Bangladesh. BMC Vet Res. 2011;7:27. | |
Yangzom T, Cruz I, Bern C, et al. Endemic transmission of visceral leishmaniasis in Bhutan. Am J Trop Med Hyg. 2012;87:1028–1037. | |
Elnaiem D-EA. Ecology and control of the sand fly vectors of Leishmania donovani in East Africa, with special emphasis on Phlebotomus orientalis. J Vector Ecol. 2011;36:S23–S31. | |
Ready PD. Leishmaniasis emergence in Europe. Euro Surveill. 2010;15(10):19505. | |
John DT, Petri WA. Markell and Voge’s Medical Parasitology. 9th ed. Missouri: Saunders/Elsevier Inc.; 2006:127–139. | |
Barrett MP, Croft SL. Management of trypanosomiasis and leishmaniasis. Br Med Bull. 2012;104:175–196. | |
McGwire BS, Satoskar AR. Leishmaniasis: clinical syndromes and treatment (Sir William Osler Medicine Master Class). Q J Med. 2014;107:7–14. | |
Mondal D, Khan MG. Recent advances in post-kala-azar dermal leishmaniasis. Curr Opin Infect Dis. 2011;24:418–422. | |
Franco AO, Davies CR, Mylne A, et al. Predicting the distribution of canine leishmaniasis in western Europe based on environmental variables. Parasitology. 2011;138:1878–1891. | |
Srivastava P, Gidwani K, Picado A, et al. Molecular and serological markers of Leishmania donovani infection in healthy individuals from endemic areas of Bihar, India. Trop Med Int Health. 2013;18(5):548–554. | |
Bhattacharyya T, Boelaert, Miles MA. Comparison of visceral leishmaniasis diagnostic antigens in African and Asian Leishmania donovani reveals extensive diversity and region-specific polymorphisms. PLoS Negl Trop Dis. 2013;7(2):e2057. | |
Quinnell RJ, Carson C, Reithinger R, Garcez LM, Courtenay O. Evaluation of rK39 rapid diagnostic tests for canine visceral leishmaniasis: longitudinal study and meta-analysis. PLoS Negl Trop Dis. 2013;7(1):e1992. | |
Chappuis F, Sundar S, Hailu A, et al. Visceral leishmaniasis: what are the needs for diagnosis, treatment and control? Nature Rev Microbiol. 2007;5:S7–S16. | |
Mauricio IL, Yeo M, Baghaei M, et al. Towards multilocus sequence typing of the Leishmania donovani complex: resolving genotypes and haplotypes for five polymorphic metabolic enzymes (ASAT, GPI, NH1, NH2, PGD). Int J Parasitol. 2006;36(7):757–769. | |
Montalvo AM, Fraga J, Maes I, Dujardin JC, Van der Auwera G. Three new sensitive and specific heat-shock protein 70 PCRs for global Leishmania species identification. Eur J Clin Microbiol Infect Dis. 2012;31:1453–1461. | |
Menten J, Boelaert M, Lesaffre E. Bayesian meta-analysis of diagnostic tests allowing for imperfect reference standards. Stat Med. 2013;32(30):5398–5413. | |
Ehehalt U, Schunk M, Jensenius M, et al. Leishmaniasis acquired by travellers to endemic regions in Europe – a EuroTraveNet multicentre study. Travel Med Infect Dis. 2014;12:167–172. | |
Alvar J, Croft SL, Kaye P, Khamesipour A, Sundar S, Reed SG. Case study for a vaccine against leishmaniasis. Vaccine. 2013;31 Suppl 2: B244–B249. | |
Joshi AB, Das ML, Akhter S, et al. Chemical and environmental vector control as a contribution to the elimination of visceral leishmaniasis on the Indian subcontinent: cluster randomized controlled trials in Bangladesh, India and Nepal. BMC Med. 2009;7:54. | |
Picado A, Das ML, Kumar V, et al. Effect of village-wide use of long-lasting insecticidal nets on visceral leishmaniasis vectors in India and Nepal: a cluster randomized trial. PLoS Negl Trop Dis. 2010;4(1):e587. | |
Quinnell RJ, Courtenay O. Transmission, reservoir hosts and control of zoonotic visceral leishmaniasis. Parasitology. 2009;136:1915–1934. | |
Bern C, Courtenay O, Alvar J. Of cattle, sand flies and men: a systematic review of risk factor analyses for South Asian visceral leishmaniasis and implications for elimination. PLoS Negl Trop Dis. 2010;4:e599. | |
Picado A, Singh SP, Rijal S, et al. Longlasting insecticidal nets for prevention of Leishmania donovani infection in India and Nepal: paired cluster randomised trial. BMJ. 2010;341:c6760. | |
Chowdhury R, Dotson E, Blackstock AJ, et al. Comparison of insecticide-treated nets and indoor residual spraying to control the vector of visceral leishmaniasis in Mymensingh District, Bangladesh. Am J Trop Med Hyg. 2011;84:662–667. | |
Stauch A, Sarkar RR, Picado A, et al. Visceral leishmaniasis in the Indian subcontinent: modelling epidemiology and control. PLoS Negl Trop Dis. 2011;5(11):e1405. | |
Hartemink N, Vanwambeke SO, Heesterbeek H, et al. Integrated mapping of establishment risk for emerging vector-borne infections: a case study of canine leishmaniasis in southwest France. PLoS One. 2011;6:e20817. | |
Argaw D, Mulugeta A, Herrero M, et al. Risk factors for visceral leishmaniasis among residents and migrants in Kafta-Humera, Ethiopia. PLoS Negl Trop Dis. 2013;7(11):e2543. | |
Luckhart S, Lindsay SW, James AA, Scott TW. Reframing critical needs in vector biology and management of vector-borne disease. PLoS Negl Trop Dis. 2010;4:e566. | |
Rowland M. Malaria control – achievements and prospects. Outlooks on Pest Management. 2012;23:150–151. | |
TDR. Research to support the elimination of visceral leishmaniasis. TDR Annual report 2008, Business Line 10. 2009;1–27. | |
Picado A, Singh SP, Vanlerberghe V, et al. Residual activity and integrity of PermaNet® 2.0 after 24 months of household use in a community randomised trial of long lasting insecticidal nets against visceral leishmaniasis in India and Nepal. Trans R Soc Trop Med Hyg. 2012;106(3):150–159. | |
Stockdale L, Newton R. A review of preventative methods against human leishmaniasis infection. PLoS Negl Trop Dis. 2013;7(6):e2278. | |
Meheus F, Abuzaid AA, Baltussen R, et al. The economic burden of visceral leishmaniasis in Sudan: an assessment of provider and household costs. Am J Trop Med Hyg. 2013;89(6):1146–1153. | |
Uranw S, Meheus F, Baltussen R, Rijal S, Boelaert M. The household costs of visceral leishmaniasis care in south-eastern Nepal. PLoS Negl Trop Dis. 2013;7(2):e2062. | |
Tusting LS, Willey B, Lucas H, et al. Socioeconomic development as an intervention against malaria: a systematic review and meta-analysis. Lancet. 2013;382(9896):963–972. | |
Hayden EC. Projects set to tackle neglected diseases. Nature. 2014; 505:142. |
 © 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2014 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.