")
Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 17
Empagliflozin Induces Vascular Relaxation in Rat Coronary Artery Due to Activation of BK Channels
Authors Kong Q, Qian LL, Zhang L, Liu HH, Yang F, Zhang XL, Wang C, Zhao XX, Li KL, Wang RX
Received 7 November 2023
Accepted for publication 10 January 2024
Published 20 January 2024 Volume 2024:17 Pages 247—257
DOI https://doi.org/10.2147/DMSO.S419125
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Konstantinos Tziomalos
Qi Kong,1 Ling-ling Qian,1 Lei Zhang,1 Huan-huan Liu,1,2 Fan Yang,1 Xiao-lu Zhang,1 Chao Wang,1 Xiao-xi Zhao,1 Ku-lin Li,1 Ru-xing Wang1,2
1Department of Cardiology, The Affiliated Wuxi People’s Hospital of Nanjing Medical University, Wuxi People’s Hospital, Wuxi Medical Center, Nanjing Medical University, Wuxi, 214023, People’s Republic of China; 2Wuxi School of Medicine, Jiangnan University, Wuxi, 214122, People’s Republic of China
Correspondence: Ru-xing Wang; Ku-lin Li, Department of Cardiology, The Affiliated Wuxi People’s Hospital of Nanjing Medical University, Wuxi People’s Hospital, Wuxi Medical Center, Nanjing Medical University, Wuxi, 214023, People’s Republic of China, Email [email protected]; [email protected]
Purpose: The aim of this study was to investigate the effects and mechanisms of SGLT2 inhibitor empagliflozin on diabetic coronary function.
Methods: A rat diabetic model was established by injection of streptozotocin. Rats in the treated group were administered empagliflozin by gavage and rat coronary vascular tensions were measured after eight weeks. Large conductance calcium activated K+ channel currents were recorded using a patch clamp technique, while human coronary artery smooth muscle cells were used to explore the underlying mechanisms.
Results: After incubation with empagliflozin (10, 30, 100, 300, 1000 μmol/L), the Δ relaxation % of rat coronary arteries were 2.459 ± 1.304, 3.251 ± 1.119, 6.946 ± 3.407, 28.36 ± 11.47, 86.90 ± 3.868, respectively. Without and with empagliflozin in the bath solution, BK channel opening probabilities at a membrane potential of +60 mV were 0.0458 ± 0.0517 and 0.3413 ± 0.2047, respectively (p < 0.05, n = 4 cells). After incubation with iberiotoxin, the Δ tensions of rat coronary arteries in the control (Ctrl), untreated (DM), low empagliflozin (10 mg/kg/d)-treated (DM+L-EMPA) and high empagliflozin (30mg/kg/d)-treated (DM+H-EMPA) group were 103.20 ± 5.85, 40.37 ± 22.12, 99.47 ± 28.51, 78.06 ± 40.98, respectively (p < 0.01 vs Ctrl, n = 3– 7; p < 0.001 vs DM+L-EMPA, n = 5– 7). Empagliflozin restored high glucose-induced downregulation of Sirt1, Nrf2, and BK-β 1, while the effect of empagliflozin disappeared in the presence of EX-527, a Sirt1 selective inhibitor.
Conclusion: Empagliflozin has a vasodilation effect on the coronary arteries in a concentration-dependent manner and can activate BK channels via the Sirt1-Nrf2 mechanism.
Keywords: diabetes, SGLT2 inhibitor, vasodilatation, large conductance calcium activated potassium channel
Introduction
Diabetes mellitus (DM), as a chronic non-communicable disease, seriously harms human health. According to the World Health Organization, more than 3 million people worldwide die from diabetes mellitus and related complications each year, mainly from cardiovascular disease (CVD), especially with the most severe coronary vascular lesions.1 Therefore, exploring the pathogenesis of diabetic coronary function and improving diabetic cardiovascular lesions are of great significance.
Sodium-glucose co-transporter-2 (SGLT2) inhibitors are oral hypoglycemic drugs that reduce blood glucose level by inhibiting glucose re-absorption in the renal tubules. Representative drugs of this kind include empagliflozin (EMPA), canagliflozin, dapagliflozin, etc. In addition to glucose control, SGLT2 inhibitors have a wide range of cardiovascular benefits, including improving cardiac energy supply, reducing arterial stiffness, and inhibiting cardiac remodeling effects.2–4 Recent studies have demonstrated that SGLT2 inhibitor is of great significance in vasodilation, independently of its hypoglycemic effect, which may be associated with K+ channels on vascular smooth muscle cells (SMCs).5,6
Large conductance calcium activated K+ (BK) channel consists of four α subunits and four β subunits,7 widely distributed in all tissues and cells of the human body, especially in vascular SMCs, and is one of the main K+ channels on coronary artery SMCs.8 Our previous study showed that,9 BK channels regulate coronary artery relaxation, and BK channel function is impaired in diabetes mellitus.
In this study, we aimed to examine the vasodilation of empagliflozin and the effect of empagliflozin on BK channels, exploring new mechanisms of the cardiovascular protection of SGLT2 inhibitors and providing a new clinical strategy for the management of cardiovascular complications in diabetes mellitus.
Materials and Methods
Animal Care and Experimental Procedures
Male Sprague Dawley (SD) rats (weight 180 ± 20g) were purchased from Changzhou Carvens Laboratory Animal Co., Ltd. in China. The rats were provided food and water ad libitum. The rat diabetic model was established as previously described.10 Briefly, the rats were administered an intraperitoneal injection of streptozocin (STZ, 60 mg/kg, Sigma), and one week later, rats with a tail tip blood glucose concentration >16.7 mmol/L were enrolled in our study. Then, the diabetic rats were randomly divided into three groups: untreated (DM) group, low empagliflozin (10 mg/kg/d)-treated (DM+L-EMPA) group, high empagliflozin (30 mg/kg/d)-treated (DM+H-EMPA) group, SD rats of the same age as the control (Ctrl) group, which was based on the concentration of empagliflozin used in animal experiments as reported,11–13 rats in the treated group were administered empagliflozin by gavage for eight weeks. All experiments were conducted according to the internationally accepted principles by the ethical guidelines by the International Council for Laboratory Animal Science (ICLAS), and all animal protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of Nanjing Medical University.
Cell Culture
Human coronary smooth muscle cells (hcSMCs) were purchased from ATCC and grown in SMC basal medium (1101, ScienCell) containing 5% fetal bovine serum. The cells propagated at 37°C in a humidified atmosphere of 95% air and 5% CO2. The groups were as follows: control (Con, contain 5.5 mmol/L glucose) group, high glucose (HG, contain 33.3 mmol/L glucose) group, empagliflozin (1 μmol/L, BI10773, MedChemExpress) intervention (HG + EMPA) group, EX-527 (a Sirt1 selective inhibitor, 1 μmol/L, ab141506, Abcam) intervention (HG + EMPA + EX-527) group. Change the medium with 1% bovine serum, then add empagliflozin and glucose and EX-527 was added 15 minutes in advance. The intervention time was 14 days. Change the culture medium and drugs once in 2–3 days.
Isolation of Rat Coronary Arteries
After being anesthetized with intraperitoneal injection of pentobarbital sodium (1.5%), the rat hearts were quickly obtained and placed in 4°C physiological saline solution (in mmol/L): NaCl 145, KCl 4, CaCl2 0.05, MgCl2 1, HEPES 10, and glucose 10, and adjusted pH to 7.4 with NaOH. Then, separating connective tissue around the coronary artery under a microscope (Olympus SZX10 Stereo Microscope), the septal coronary arteries, right coronary arteries, and left coronary arteries were dissected carefully. Arteries were used for vascular tension experiments, patch clamp experiments, or preservation at −80°C.
Coronary Artery Tension Measurements
Rat coronary arteries were isolated and cut into 1.5–2.0 mm, then mounted on a four-chamber wire myograph (model 620 M, Danish Myo Technology). The coronary arteries were maintained at 37°C in physiological salt solution (PSS, in mmol/L): NaCl 130, KCl 4.7, KH2PO4 1.18, MgSO4•7H2O 1.17, NaHCO3 14.9, Glucose 5.5, EDTA 0.026, and CaCl2 1.6, at pH 7.4. Pre-warmed PSS at 37°C, gassed with 95% O2 and 5% CO2, and maintained the arteries ring at 2.5 mN. Tested the coronary artery tension with KCl (60 mmol/L) and referenced as the maximum contraction for contractile capacity. Then, we performed the following procedures: 1) After the vascular tension rises to equilibrium, wash it three times with PSS. When the vessel tension returns to the basal value, add iberiotoxin (IBTX, 100 nmol/L, I5904, Sigma) or 4-Aminopyridine (4-AP, 10 mmol/L, 275875, Sigma) to the bath until the vessel tension reaches the maximum value and acts for 1 hour. The ratio of the amplitude of vascular tension change of IBTX or 4-AP to KCl, namely Δ tensions (% Max) of rat coronary arteries can be obtained. 2) After the vascular tension rises to equilibrium, empagliflozin (10 μmol/L, 30 μmol/L, 100 μmol/L, 300 μmol/L, 1000 μmol/L) was added to observe the effect of vasodilation. After the experiment, KCl was used again to verify the vascular activity.
Single Channel Electrophysiology
Inside-out single BK currents were recorded from freshly isolated rat coronary artery SMCs as we have previously described14 using an Axopatch 200B Amplifier (Molecular Devices, Inc., Sunnyvale, CA, USA), filtered at 2 kHz, and sampled at 50 kHz. All experiments were performed at room temperature (22–24°C). Inside-out single BK channel currents were elicited at +60 mV before and after perfusing EMPA. Channel open probabilities (NP0) were calculated using Clampfit 10.4 software (Axon Instruments, Foster City, CA, USA). Pipette resistance was 5–10 MΩ when filled with the pipette solution that contained (in mmol/L) KCl 140, CaCl2 1, MgCl2 1, HEPES 10, and EGTA 1 with a pH of 7.4. The bath solution contained (in mmol/L) KCl 140, MgCl2 1, EGTA 1, HEPES 10, CaCl2 0.816 (1 μmol/L free Ca2+) at pH 7.35.
Western Blot Analysis
The rat coronary arterial fragments or hcSMCs were lysed in RIPA buffer (89901, Thermo scientific), which contained protease inhibitors (B14001, Bimake). The lysates were fractionated using SDS-PAGE and transferred onto polyvinylidene di-fluoride (PVDF) membranes (Millipore). The PVDF membranes were incubated with specific primary antibodies, with β-actin (ab20272, Abcam, 1:5000), BK-α (APC-021, Alomone Labs, 1:500), BK-β1 (APC-036, Alomone Labs, 1:1000), Sirt1 (ab110304, Abcam, 1:2000) and Nrf2 (SAB5700720, Sigma, 1:1000). Western blot bands were quantified using densitometry and ImageJ software. The density was normalized to the control treatment level, and the relative folds were normalized to the β-actin level.
Statistical Analysis
Statistical analysis was performed by GraphPad Prism 8 and SPSS 22.4 software. Data are expressed as average ± SEM. Student’s t-test was used to compare data between two groups. One-way ANOVA with posthoc LSD analysis was used to compare data between multiple groups. Statistical significance was defined as a p value of < 0.05.
Results
Effects of Empagliflozin on Coronary Artery Function
In order to investigate the direct effect of empagliflozin on arterial constriction, we examined vascular tone in rat coronary arteries. Under the continuous action with KCl, the blood vessel constriction reached its peak and then empagliflozin was added successively with concentrations of 10 μmol/L, 30 μmol/L, 100 μmol/L, 300 μmol/L, and 1000 μmol/L. The results showed that the relaxation % were 2.459 ± 1.304, 3.251 ± 1.119, 6.946 ± 3.407, 28.36 ± 11.47, 86.90 ± 3.868, respectively (n = 8 arteries from 4 rats) (Figure 1a and b). These data demonstrated that empagliflozin has a vasodilation effect on normal rat coronary vessels in a concentration-dependent manner.
 |
Figure 1 Empagliflozin induces vasodilation in KCl-constricted coronary arteries. Abbreviation: EMPA, empagliflozin. Notes: (a) An original trace showing concentration-dependent inhibition of 60K-induced constriction in a coronary artery by empagliflozin (10 to 1000 μmol/L). (b) Mean data for relaxation % of vasodilation induced by empagliflozin, n = 8. |
Effects of Empagliflozin on Single BK Channel Currents
Arterial SMCs express various K+ channels, among which BK channels are one of the most important K+ channels in coronary SMCs, and play a crucial role in the regulation of coronary artery function.15 Therefore, we sought to examine the hypothesis that empagliflozin may directly activate BK channels to dilate coronary arteries.
Single BK channel currents were recorded from freshly isolated rat coronary artery SMCs in an inside-out configuration. In the presence of 1 μmol/L free calcium, without and with 1 μmol/L EMPA in the bath solution, BK channel opening probabilities at a membrane potential of +60 mV were 0.0458 ± 0.0517 and 0.3413 ± 0.2047, respectively (p < 0.05, n = 4 cells from 4 rats). The result suggests that empagliflozin activates BK channels in normal rats and increases the open probability of BK channels (Figure 2a and b).
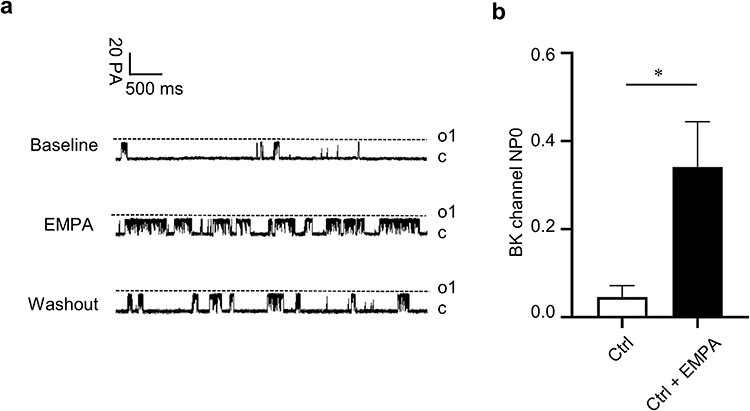 |
Figure 2 Empagliflozin activates BK channels. Abbreviation: EMPA, empagliflozin. Notes: (a) The single-channel patch-clamp experimental technique was used to record the BK currents before and after empagliflozin (1µM) in coronary SMCs of normal rats at 60 mV stimulation voltage. (b) The statistical diagram. *p < 0.05 vs no EMPA, n = 4 cells. |
Effects of Chronic Treatment of Empagliflozin on Blood Glucose and Body Weight in Rats
To explore the effects of chronic treatment of empagliflozin in diabetic rats, we selected two doses of empagliflozin to treat DM rat for 8 weeks, and a total of 34 rats were eventually included. The blood glucose levels and the body weights of four groups of rats are shown in Figure 3. The blood glucose of the Ctrl group was kept at around 6 mmol/L, while the glucose level of the DM was maintained >25 mmol/L, and empagliflozin intervention failed to improve the blood glucose status of the rats (Figure 3a). The Ctrl rats showed a gradual increase in body weight and the other three groups showed a slight increase in the early stage and then decreased slightly (Figure 3b).
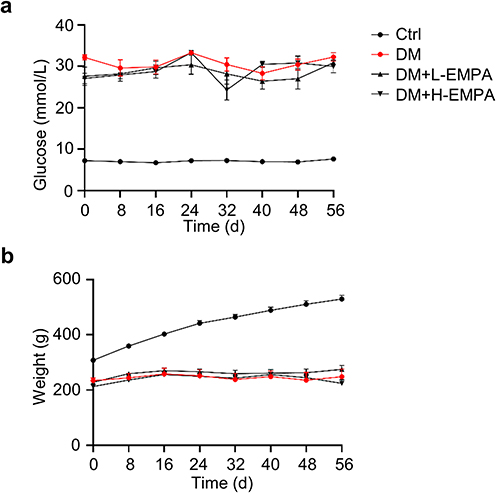 |
Figure 3 Effects of streptozotocin and empagliflozin on blood glucose and body weight in rats. Abbreviation: EMPA, empagliflozin. Notes: (a) Blood glucose and (b) Body weight of Ctrl, DM, DM+L-EMPA, and DM+H-EMPA group at 8 weeks after successful diabetic modeling. Ctrl: the controlled group; DM: untreated group; DM+L-EMPA: low empagliflozin (10 mg/kg/d)-treated group; DM+H-EMPA: high empagliflozin (30 mg/kg/d)-treated group. |
Effects of Chronic Treatment of Empagliflozin on IBTX-Induced Vasoconstriction in Coronary Arteries
Our previous studies have shown that coronary artery SMCs have impaired BK channel function, decreased BK currents, and increased vascular tone in diabetes mellitus.9 In this study, our data showed that the application of IBTX, a selective BK blocker, induced coronary vasoconstriction in four groups. The Δ tensions (% Max) were 103.20 ± 5.85, 40.37 ± 22.12, 99.47 ± 28.51, and 78.06 ± 40.98, respectively (p < 0.01 vs Ctrl, n = 3–7 arteries from 3 to 4 rats; p < 0.001 vs DM+L-EMPA, n = 5–7 arteries from 3 to 4 rats) (Figure 4). Compared with the Ctrl, rats in DM had impaired BK channel function of mediating coronary artery relaxation (Figure 4a and b); low-dose empagliflozin treatment significantly improved BK channel function and restored its function of mediating coronary artery relaxation (Figure 4c), while high-dose empagliflozin was less effective (Figure 4d). The results suggested that chronic empagliflozin therapy improves BK channels function in diabetic rats, which may correlate with the concentrations.
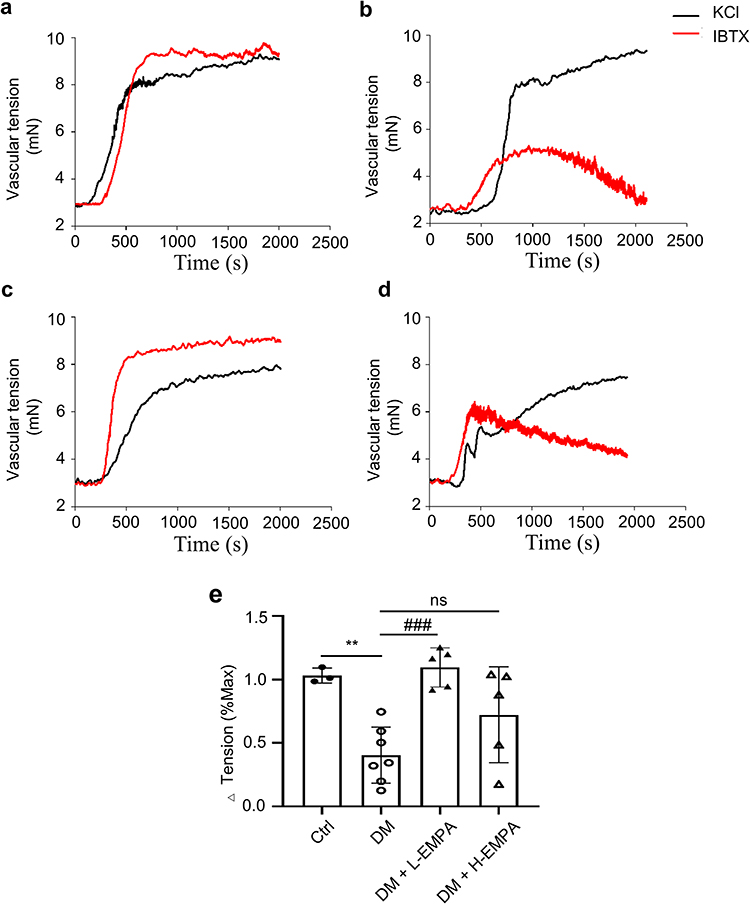 |
Figure 4 Effect of chronic treatment of empagliflozin on IBTX-induced vasoconstriction in coronary arteries. Abbreviation: EMPA, empagliflozin. Notes: (a–d) Original traces of KCl and IBTX induced coronary contractions in four groups. (e) Mean data for the ratio of vascular tension induced by IBTX to KCl in four groups. Vascular tensions in four groups increased after IBTX treatment, but the maximal vascular tension of the DM group was lower compare to Ctrl, DM+L-EMPA, and DM+H-EMPA groups. Ctrl: the controlled group; DM: untreated group; DM+L-EMPA: low empagliflozin (10 mg/kg/d)-treated group; DM+H-EMPA: high empagliflozin (30 mg/kg/d)-treated group. **p < 0.01 vs Ctrl, n = 3–7 arteries from 3 to 4 rats; ###p < 0.001 vs DM+L-EMPA, n = 5–7 arteries from 3 to 4 rats. |
Effects of Chronic Treatment of Empagliflozin on 4-AP-Induced Vasoconstriction in Coronary Arteries
Arterial SMCs also express voltage-gated K+ (KV) channels that, when activated, lead to arterial hyperpolarization and vasodilation.16 In this study, we also added 4-AP, a non-selective Kv blocker, to observe the coronary vasoconstriction response. Our results show that 4-AP induced coronary vasoconstriction in Ctrl, DM, DM+L-EMPA, and DM+H-EMPA rats. The Δ tensions (% Max) were 104.56 ± 15.30, 82.57 ± 17.66, 85.53 ± 15.15, 93.30 ± 23.35, respectively (p < 0.01 vs DM, n = 7–10 arteries from 4 rats) (Figure 5); Compared with the Ctrl, rats in DM had impaired Kv channel function, weakening their function of mediating coronary artery relaxation (Figure 5a and b); low-dose empagliflozin treatment failed to improve the impaired coronary Kv channel function caused by diabetes (Figure 5c), and high-dose empagliflozin treatment slightly improved the impaired Kv channel-mediated coronary artery function, but the difference was not statistically significant (Figure 5d).
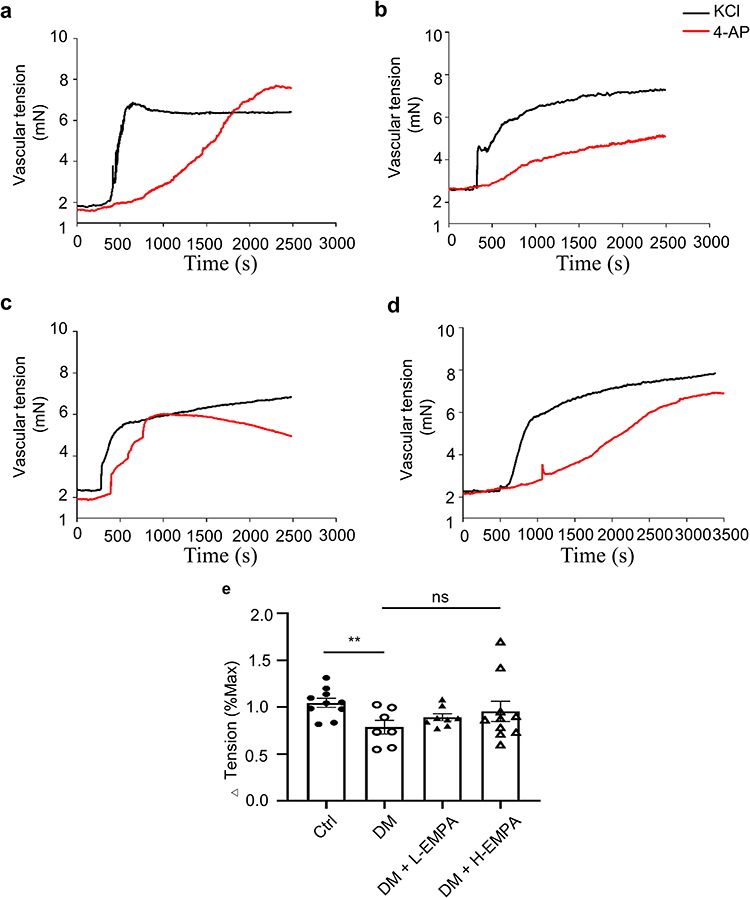 |
Figure 5 Effects of chronic treatment of empagliflozin on 4-AP-induced vasoconstriction in coronary arteries. Abbreviation: EMPA, empagliflozin. Notes: (a–d) Original traces of KCl and 4-AP induced coronary contractions in four groups. (e) Mean data for the ratio of vascular tension induced by 4-AP to KCl in four groups. Vascular tensions in four groups increased after 4-AP treatment, but the maximal vascular tension of the DM group was lower compared to the Ctrl and DM+H-EMPA groups. Ctrl: the controlled group; DM: untreated group; DM+L-EMPA: low empagliflozin (10mg/kg/d)-treated group; DM+H-EMPA: high empagliflozin (30 mg/kg/d)-treated group. **p < 0.01 vs DM, n = 7–10 arteries from four rats. |
Effects of Chronic Empagliflozin Treatment on BK-β Protein Expression
Western blot analysis showed that the expression of BK-β1 subunits in DM rats was significantly decreased compared to Ctrl rats; low-dose empagliflozin treatment for 8 weeks increased BK-β1 subunit expression compared to the DM, all the differences were statistically significant (p < 0.05 vs Ctrl, p < 0.05 vs DM+L-EMPA, n = 7), while high-dose has less effect (Figure 6). As what we expected, the Western blot data were consistent with the coronary vascular tension data.
 |
Figure 6 Western blots for BK protein expression in four groups. Abbreviation: EMPA, empagliflozin. Notes: (a) Representative images of Western blot analysis of BK-β1 and BK-α subunit; β-actin was used as internal control; (b and c) Mean data for relative protein expression of BK-β1 and BK-α. Ctrl: the controlled group; DM: untreated group; DM+L-EMPA: low empagliflozin (10mg/kg/d)-treated group; DM+H-EMPA: high empagliflozin (30 mg/kg/d)-treated group. *p < 0.05 vs Ctrl, #p < 0.05 vs DM+L-EMPA, n = 7. |
Empagliflozin Activates BK Channels Through Sirt1-Nrf2 Pathway in hcSMCs Chronically Exposed to High Glucose
We hypothesized that empagliflozin could activate BK channels through Sirt1-Nrf2 pathway. We performed in vitro studies on hcSMCs chronically exposed to high glucose levels. Consistent with in vivo experiments, hcSMCs treated with 1 μmol/L EMPA showed that the inhibition of Sirt1 and Nrf2 induced by high glucose promoted the decrease of BK-β1 expression, applying of empagliflozin increases BK-β1 expression, while the effect of empagliflozin disappeared in the presence of EX527, a Sirt1 selective inhibitor (Figure 7). Taken together, these data demonstrated that empagliflozin activates BK channels through Sirt1-Nrf2 pathway.
 |
Figure 7 Empagliflozin activates BK channel through Sirt1-Nrf2 pathway in hcSMCs chronically exposed to high glucose. Abbreviation: EMPA, empagliflozin. Notes: (a) Representative images of Western blot analysis of Sirt1, Nrf2, BK-β1, and BK-α, β-actin was used as internal control; (b–e) Mean data for relative protein expression of Sirt1, Nrf2, BK-β1, and BK-α. Con: control group (5.5mM glucose); HG: high glucose (33.3mM glucose) group; HG + EMPA: empagliflozin intervention group; HG + EMPA + EX-527: EX-527 intervention group. *p < 0.05, **p < 0.01 vs HG; #p < 0.05 vs HG+EMPA; $p < 0.05 vs HG+EMPA+EX-527; $$$p < 0.001 vs HG+EMPA+EX-527; n = 3–8. |
Discussion
In this study, we have made several main findings. First, empagliflozin had a vasodilation effect on the coronary arteries of normal rats in a concentration-dependent manner. Second, empagliflozin activates BK channels, and its function in vasodilation is related to BK channels. Third, empagliflozin increases BK channel protein expression in diabetic rats and restores its function in mediating vasodilation. Fourth, empagliflozin activates BK channels via the Sirt1-Nrf2 signal pathway.
SGLT2 inhibitors are commonly used in the clinical practice, and their cardiovascular benefits are widely recognized. Many studies have demonstrated that SGLT2 inhibitors play an important role in vasodilation, independently of its hypoglycemic effect. Consistent with our study, De Stefano et al17 demonstrated that empagliflozin induces vasodilation in rabbit thoracic aorta in a concentration-dependent manner. Moreover, Ahasanul et al5,6,18 investigated the effects of empagliflozin, dapagliflozin, and canagliflozin on normal rat mesenteric arteries and found that all three SGLT2 inhibitors relax rat mesenteric arteries in a concentration-dependent manner.
A recent study19 suggested that both canagliflozin and phlorizin (natural SGLT2 inhibitors) relaxed pulmonary arteries in a dose-dependent manner, while SGLT2 inhibitors (including canagliflozin) have small relaxation effects on coronary arteries. They used 10 μmol/L as the maximum concentration of dapagliflozin in the coronary group, while our study found empagliflozin had significant relaxation on normal rat coronary arteries in 300 μmol/L. This indicates that the sensitivity of vessels to SGLT2 inhibitors is different, although it is different in SGLT2 inhibitors of the two studies.
One of the important questions was whether SGLT2 inhibitors exert vasodilatory effects by activating BK channels. Our data indicate that 1 μmol/L empagliflozin activated BK channels; however, existing studies showed favorable effects of Kv channels in vasodilation of SGLT2 inhibitors, and the isoforms involving Kv channels are not entirely consistent. This may be due to the difference in type and concentration of SGLT2 inhibitors and the difference in the proportion of K+ channels in various arteries. Chronic treatment with empagliflozin fails to improve the glucose status in the rats, which excludes additional effects of the hypoglycemic effect of SGLT2 inhibitors. We found that only low-dose empagliflozin treatment restored the function of BK channel-mediated vasodilation; high-dose empagliflozin treatment slightly improved the function of Kv channels-mediated vasodilation, but the difference was not statistically significant. We were unable to determine the causes for the different results between the low and high concentrations, there might be several possibilities to support our results: 1) Intervention form: Most of the existing studies with SGLT2 inhibitors and ion channel inhibitors are in vitro acute experiments, and the results of acute experiment and chronic experiment are not completely consistent. We did not perform in vitro experiments with ion channel inhibitors and detect total K+ channel-mediated vasodilation function. 2) K+ channel proportion: BK channels are one of the major K+ channels on coronary artery smooth muscle cells and account for about 65% of the total K+ channel currents in rat coronary artery smooth muscle cells.8 This could explain the appropriate concentration of empagliflozin acting primarily through BK channels. 3) Drug concentrations: Our study showed that empagliflozin activates BK at low concentrations, and its effect is not obvious at high concentrations, while we can find a slight improvement of empagliflozin on Kv-mediated coronary vasodilation; thus, we make a bold and reasonable conjecture that SGLT2 inhibitors may activate vasodilation through multiple K+ channels, which may depend on drug concentrations, intervention time, and vessel type. This requires more studies to verify.
The Sirt1-Nrf2 signaling pathway is widely involved in oxidative stress, inflammation, and apoptosis, and has positive implications for organs such as heart, brain, eye, and kidney,20,21 but its role in the diabetic coronary arteries is still unclear. Recent studies have shown that SGLT2 inhibitors can upregulate Sirt1 to achieve their cardiovascular beneficial effects.22–24 In this study, we propose that SGLT2 inhibitors activate BK channels through the Sirt1-Nrf2 pathway. To explore the role of Sirt1-Nrf2 signaling in the up regulation of BK-β1 subunits by empagliflozin, we performed in vitro studies using hcSMCs. The results prompted that empagliflozin activates BK channels through the Sirt1-Nrf2 signaling pathway.
Conclusion
To summarize, our results demonstrated that empagliflozin has a vasodilation effect on the coronary arteries in a concentration-dependent manner. Empagliflozin-induced coronary arteries vasodilation at low concentrations is mediated through BK channels, while empagliflozin at high concentrations still needs further investigation. In addition, empagliflozin can activate BK channels via the Sirt1-Nrf2 mechanism. We explored new mechanisms of the cardiovascular protective effects of SGLT2 inhibitors and provide a new clinical strategy for the management of cardiovascular complications in diabetes mellitus.
Funding
This work was supported in part by grants from the National Natural Science Foundation of China (grant numbers: 81770331) and Top Talent Support Program for Young and Middle-aged People of Wuxi Health Committee (No. BJ2020018).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Preis SR, Hwang SJ, Coady S, et al. Trends in all-cause and cardiovascular disease mortality among women and men with and without diabetes mellitus in the Framingham heart study, 1950 to 2005. Circulation. 2009;119(13):1728–1735. doi:10.1161/CIRCULATIONAHA.108.829176
2. Santos-Gallego CG, Requena-Ibanez JA, San Antonio R, et al. Empagliflozin ameliorates adverse left ventricular remodeling in nondiabetic heart failure by enhancing myocardial energetics. J Am Coll Cardiol. 2019;73:1931–1944. doi:10.1016/j.jacc.2019.01.056
3. Zanchi A, Burnier M, Muller ME, et al. Acute and chronic effects of SGLT2 inhibitor empagliflozin on renal oxygenation and blood pressure control in nondiabetic normotensive subjects: a randomized, placebo-controlled trial. J Am Heart Assoc. 2020;9(13):e016173. doi:10.1161/JAHA.119.016173
4. Gaspari T, Spizzo I, Liu H, et al. Dapagliflozin attenuates human vascular endothelial cell activation and induces vasorelaxation: a potential mechanism for inhibition of atherogenesis. Diab Vasc Dis Res. 2018;15(1):64–73. doi:10.1177/1479164117733626
5. Hasan A, Hasan R. Empagliflozin relaxes resistance mesenteric arteries by stimulating multiple smooth muscle cell voltage-gated K+ (KV) Channels. Int J Mol Sci. 2021;22(19):10842. doi:10.3390/ijms221910842
6. Hasan A, Zerin F, Menon SN, Alam MA, Hasan R. Mechanism of canagliflozin-induced vasodilation in resistance mesenteric arteries and the regulation of systemic blood pressure. J Pharmacol Sci. 2022;150(4):211–222. doi:10.1016/j.jphs.2022.09.004
7. Salkoff L, Butler A, Ferreira G, Santi C, Wei A. High-conductance potassium channels of the SLO family. Nat Rev Neurosci. 2006;7(12):921–931. doi:10.1038/nrn1992
8. Wang RX, Chai Q, Lu T, Lee HC. Activation of vascular BK channels by docosahexaenoic acid is dependent on cytochrome P450 epoxygenase activity. Cardiovasc Res. 2011;90(2):344–352. doi:10.1093/cvr/cvq411
9. Sun MQ, Qian LL, Miao LF, et al. Impairment of TRPC1-BK complex in diabetic rat coronary artery. Chin Med J. 2022;135(15):1873–1875. doi:10.1097/CM9.0000000000001904
10. Tang X, Qian LL, Wang RX, et al. Regulation of coronary arterial large conductance Ca2+-Activated K+ channel protein expression and function by n-3 polyunsaturated fatty acids in diabetic rats. J Vasc Res. 2017;54(6):329–343. doi:10.1159/000479870
11. Shao Q, Meng L, Lee S, et al. Empagliflozin, a sodium glucose co-transporter-2 inhibitor, alleviates atrial remodeling and improves mitochondrial function in high-fat diet/streptozotocin-induced diabetic rats. Cardiovasc Diabetol. 2019;18(1):165. doi:10.1186/s12933-019-0964-4
12. Park SH, Farooq MA, Gaertner S, et al. Empagliflozin improved systolic blood pressure, endothelial dysfunction and heart remodeling in the metabolic syndrome ZSF1 rat. Cardiovasc Diabetol. 2020;19(1):19. doi:10.1186/s12933-020-00997-7
13. Oelze M, Kröller-Schön S, Welschof P, et al. The sodium-glucose co-transporter 2 inhibitor empagliflozin improves diabetes-induced vascular dysfunction in the streptozotocin diabetes rat model by interfering with oxidative stress and glucotoxicity. PLoS One. 2014;9(11):e112394. doi:10.1371/journal.pone.0112394
14. Wang RX, Shi HF, Chai Q, et al. Molecular mechanisms of diabetic coronary dysfunction due to large conductance Ca2⁺-activated K⁺ channel impairment. Chin Med J. 2012;125(14):2548–2555.
15. Qian LL, Sun MQ, Wang RX, et al. Mechanisms of BK channel activation by docosahexaenoic acid in rat coronary arterial smooth muscle cells. Front Pharmacol. 2018;9:223. doi:10.3389/fphar.2018.00223
16. Jackson WF. KV channels and the regulation of vascular smooth muscle tone. Microcirculation. 2018;25(1):
17. Seo MS, Jung HS, An JR, et al. Empagliflozin dilates the rabbit aorta by activating PKG and voltage-dependent K+ channels. Toxicol. Appl Pharmacol. 2020;403:115153. doi:10.1016/j.taap.2020.115153
18. Hasan A, Menon SN, Zerin F, Hasan R. Dapagliflozin induces vasodilation in resistance-size mesenteric arteries by stimulating smooth muscle cell KV7 ion channels. Heliyon. 2022;8(5):e09503. doi:10.1016/j.heliyon.2022.e09503
19. Han Y, Cho YE, Ayon R, et al. SGLT inhibitors attenuate NO-dependent vascular relaxation in the pulmonary artery but not in the coronary artery. Am J Physiol Lung Cell Mol Physiol. 2015;309(9):L1027–L1036. doi:10.1152/ajplung.00167.2015
20. Arab HH, Ashour AM, Eid AH, Arafa EA, Al Khabbaz HJ, Abd El-Aal SA. Targeting oxidative stress, apoptosis, and autophagy by galangin mitigates cadmium-induced renal damage: role of SIRT1/Nrf2 and AMPK/mTOR pathways. Life Sci. 2022;291:120300. doi:10.1016/j.lfs.2021.120300
21. Jin Q, Zhu Q, Wang K, Chen M, Li X. Allisartan isoproxil attenuates oxidative stress and inflammation through the SIRT1/Nrf2/NF-κB signalling pathway in diabetic cardiomyopathy rats. Mol Med Rep. 2021;23(3):215. doi:10.3892/mmr.2021.11854
22. Ren FF, Xie ZY, Jiang YN, et al. Dapagliflozin attenuates pressure overload-induced myocardial remodeling in mice via activating SIRT1 and inhibiting endoplasmic reticulum stress. Acta Pharmacol Sin. 2022;43(7):1721–1732. doi:10.1038/s41401-021-00805-2
23. Fang R, Chen J, Long J, et al. Empagliflozin improves kidney senescence induced by D-galactose by reducing sirt1-mediated oxidative stress. Biogerontology. 2023;24(5):771–782. doi:10.1007/s10522-023-10038-x
24. Zaghloul MS, Elshal M, Abdelmageed ME. Preventive empagliflozin activity on acute acetic acid-induced ulcerative colitis in rats via modulation of SIRT-1/PI3K/AKT pathway and improving colon barrier. Environ Toxicol Pharmacol. 2022;91:103833. doi:10.1016/j.etap.2022.103833
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.