Back to Journals » Infection and Drug Resistance » Volume 16
Emergence of ST1193 Clone in Maternal and Neonatal ESBL-Producing E. coli Isolates
Authors Wei L, Chen Q, Yang L, Ji T, Peng W, Shao B, Li H
Received 24 April 2023
Accepted for publication 14 September 2023
Published 13 October 2023 Volume 2023:16 Pages 6681—6689
DOI https://doi.org/10.2147/IDR.S418455
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Ling Wei,1,* Qiyan Chen,2,3,* Lu Yang,2,3 Tongzhen Ji,4 Wenjing Peng,5 Bing Shao,2 Hui Li2
1Department of Obstetrics, Beijing Obstetrics and Gynecology Hospital, Capital Medical University, Beijing Maternal and Child Health Care Hospital, Beijing, People’s Republic of China; 2Beijing Key Laboratory of Diagnostic and Traceability Technologies for Food Poisoning, Beijing Center for Disease Prevention and Control, Beijing, People’s Republic of China; 3National Center for Veterinary Drug Safety Evaluation, College of Veterinary Medicine, China Agricultural University, Beijing, People’s Republic of China; 4Department of Clinical Laboratory, Beijing Obstetrics and Gynecology Hospital, Capital Medical University, Beijing Maternal and Child Health Care Hospital, Beijing, People’s Republic of China; 5College of Animal Husbandry and Veterinary Medicine, Jinzhou Medical University, Jinzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Hui Li; Bing Shao, Email [email protected]; [email protected]
Objective: The emerging epidemic of extended-spectrum β-lactamase-producing E. coli (ESBL-EC) is a global public health crisis. ESBL-EC infections are increasing worldwide and contribute to morbidity and mortality among newborn infants. However, the antimicrobial resistance characteristics and clonal transmission of maternal and neonatal ESBL-EC isolates need to be further deciphered.
Materials and Methods: We performed phenotypic and genotypic characterization of 33 ESBL-EC isolates from pregnant women and newborn during 2019– 2020.
Results: Minimum inhibitory concentrations of 17 antimicrobial agents showed that all isolates were multidrug-resistant (MDR) and had a resistance rate of 100% to ampicillin, and mild resistance to florfenicol, gentamicin, ceftazidime, and amoxicillin-clavulanate. Additionally, imipenem, meropenem, polymyxin, and tigecycline exhibited good activity against the tested ESBL-EC isolates with low MIC50 (0.06– 1 μg/mL) and MIC90 (0.06– 1 μg/mL). Whole genome sequencing indicated that ESBL-EC isolates contained diverse antimicrobial resistant genes (blaCTX-M, blaTEM, blaSHV, tetA, etc.) and toxin genes (ompA, csg, fimH, hybtA, etc.). blaCTX-M genes were the main ESBL genotype. ST1193 (18.2%) was the second most abundant ST among the ESBL-EC isolates (ST131 was the most common, with 30.3%), and this is the first report of its mother-to-infant colonization transmission in China.
Conclusion: These findings revealed the occurrence of high-risk ST1193 clone among ESBL-EC isolates from pregnant women and newborn colonization in China. Further national or regional multicenter studies are needed to assess the dissemination and evolution of ESBL-EC ST1193 clone as a nosocomial pathogen in China.
Keywords: ESBL, E. coli, antibiotic resistance, colonization transmission, ST1193 clone
Introduction
Extended-spectrum β-lactamase Enterobacteriaceae is an important public health threat of widespread clinical concern, with increasing infection rates resulting in increased neonatal mortality.1 Extended-spectrum β-lactamase-producing Escherichia coli (ESBL-EC) are the largest group of multidrug-resistant pathogens in obstetric bacteraemia.2,3 ESBL-EC infections are responsible for an estimated 14,000 healthcare-associated infections annually in the United States.4 ESBL-EC infections may lead to poor treatment outcomes, lower rates of clinical and microbiological response, prolonged hospitalization, and higher hospital costs.5,6
Since ESBL-EC can spread antimicrobial resistance among different bacteria in many ways, it is easy to cause the epidemic of antimicrobial resistant bacteria. Neonatal infection by ESBL-EC may be associated with higher fatality rates, colonized mothers may be a source of neonatal transmission.7 Maternal colonization with ESBL-EC is considered as a significant risk factor for the acquisition of neonatal infection.8,9 Fecal carriage of ESBL-EC is an important reservoir for the transmission and dissemination of infection with resistant strains.10 Previous studies on clinical patients found that fecal carriage appeared to be common several months after ESBL-EC infection.11,12 In Europe, fecal carriage rate of AmpC β-lactamase genes (blaCMY-2 and blaDHA-1) in the community was approximately 10%.13 Due to the high number of maternal and infant colonization associated with infections caused by these bacteria, the World Health Organization classifies Enterobacteriaceae as priority organisms for which new antimicrobials drugs are urgently needed.14 β-lactam antimicrobials have become the first-line antibacterial drugs for humans because of their high potency, broad anti-bacterial spectrum, and minimal side effects.15 ESBL-producing microorganisms exhibit co-resistance to many other classes of antibiotics and carry additional resistance genes, which limit therapeutic options. Inevitably, the problem of ESBL-producing antimicrobial resistance poses great threat to the treatment of clinical infections.
ESBLs are plasmid-encoded enzymes produced by Enterobacteriaceae that induce bacterial resistance by hydrolyzing β-lactam antibiotics.16,17 Currently, hundreds of ESBLs have been identified. blaTEM, blaSHV, and blaCTX-M are the common genotypes. CTX-M ESBLs are predominantly found in isolates from community-acquired infections,18 where the CTX-M-15 variant is the most common enzyme. The widespread use of third-generation cephalosporins and aztreonam is believed to be the major cause of mutations in these enzymes which has led to the emergence of the ESBLs. In addition, sequence type 131 (ST131) has been shown to contribute to the global spread of CTX-M-producing E. coli.19 Recently, ST1193 constituted an emerging fluoroquinolone-resistant E. coli clone in clinic.20,21 It is well known that the epidemiology of maternal-neonatal colonization has found that certain species or ST groups are more likely to harbor resistance or virulence determinants.22,23 Therefore, a comprehensive analysis of antibiotic resistome of ESBL-EC isolated from pregnant women and newborn could decipher the spread of molecular transmission. To investigate the prevalence and molecular characteristics of maternal and neonatal ESBL-EC isolates in Beijing, China, we evaluated the molecular profiles, including STs, phylogroups, and antibiotic resistome of ESBL-EC strains. We clarify the associations between phenotypic and genotypic data and demonstrate the presence of diverse antibiotic resistome in maternal and neonatal ESBL-EC.
Materials and Methods
Bacteria Isolate
We studied maternal and neonatal samples collected from the Beijing Obstetrics and Gynecology Hospital affiliated to Capital Medical University in Beijing, China (detailed descriptions of the patients were given in Table S1). All clinical samples (throat swab, placenta, perineum incision, blood, urine, or cervical secretion) were inoculated onto selective chromogenic medium ESBL-E (CHROMagarTM ESBL, France) and incubated in ambient air at 37°C for 24 h. Individual colonies were then sub-cultured onto blood agar plates. Species confirmation was performed using an API20E system (bioMerieux) and matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF/MS) (Bruker, Germany). ESBL production was confirmed using the combined disc diffusion method according to the Clinical and Laboratory Standard (CLSI) guidelines.24 The production of ESBL was confirmed by an increase in the diameter of the zone tested with cefotaxime/clavulanate or ceftazidime/clavulanate relative to when tested alone by ≥5 mm.25
Antimicrobial Susceptibility Test
A total of 33 well-grown maternal and neonatal ESBL-EC isolates were selected for antimicrobial susceptibility test. The agar dilution method was used to determine their susceptibility to 17 antimicrobial agents, including imipenem, meropenem, cefepime, cefotaxime, ceftazidime, cefoxitin, tigecycline, tetracycline, polymyxin B, colistin, ampicillin, amoxicillin-clavulanate, amikacin, kanamycin, gentamicin, ciprofloxacin, and florfenicol. Antimicrobial susceptibility test was performed in accordance with the guideline of CLSI documents M100-S30.24 E. coli ATCC 25922 was used as a quality control. Minimum inhibitory concentrations (MICs) for susceptibility (S), intermediate susceptibility (I) and resistance (R) were defined according to the recommendations of the European Committee on Antimicrobial Susceptibility Testing (EUCAST).25
Whole Genome Sequencing
Genomic DNA was extracted from ESBL-EC strains using the Wizard Genomic DNA Purification Kit (Promega, Madison, WI, US). Then, DNA libraries were constructed using the KAPA Hyper Prep Kit (Roche, Basel, Switzerland). Genomic sequencing was performed using the Illumina Novaseq 6000 platform (Illumina, San Diego, CA), which generated 150-bp paired-end reads from the libraries with an average insert size of 350 bp. For each isolate, >550 M bp of high-quality clean reads were obtained and assembled de novo using SPAdes v.3.1026 by default settings and k-mer sizes ranging from 21 to 127. Multi-locus sequences were identified using the pMLST 2.0 database. Antimicrobial resistance and toxin genes were identified using the SRST2 Toolkit (version 0.2.0, University of Melbourne, http://katholt.github.io/srst2/). All genome drafts were used to construct a phylogenetic tree using the parsnp software,27 and the trees were finally visualized using the online tool iTOL (http://itol.embl.de/).
Results
Epidemiology of Isolates
We included all 33 ESBL-EC isolates in this study and the isolates were submitted from maternal and newborn patients in Beijing. The isolates were collected from a variety of sources: 9 from neonatal carriage (7 isolates from throat swabs and 2 isolate from neonatal bloodstream infection), 24 from pregnant women (16 isolates from placenta, 3 isolates from perineum incision, 3 isolates from blood, 1 strain from cervical secretions, and 1 isolate from urine).
Phenotypic Characterization of ESBL-EC Isolates
The distributions of MICs and test concentrations of different antibiotics against the tested 33 isolates are summarized in Table 1. Among the tested antibiotics, high proportion of resistance to ampicillin (n = 33, 100.0%), cefotaxime (n = 30, 90.9%), tetracycline (n = 28, 84.85%) was observed, with MIC90 in all above the dilution ranges (>128 μg/mL, >16 μg/mL, >64 μg/mL, respectively). Mild resistance was observed to gentamicin (21.21%), florfenicol (18.18%), ceftazidime (30.3%), and amoxicillin-clavulanate (12.12%). However, almost all the tested ESBL-EC isolates exhibited low-level resistance to kanamycin (3.03%), cefepime (3.03%), and cefoxitin (9.09%). We found that colistin, polymyxin B, tigecycline, amikacin, imipenem and meropenem showed good antimicrobial activity against these ESBL-EC isolates with low MIC50 (0.06–1 μg/mL) and MIC90 (0.06–1 μg/mL) (Figure 1A). Overall, ampicillin-tetracycline-cefotaxime was the most frequently observed antimicrobial resistance profile.
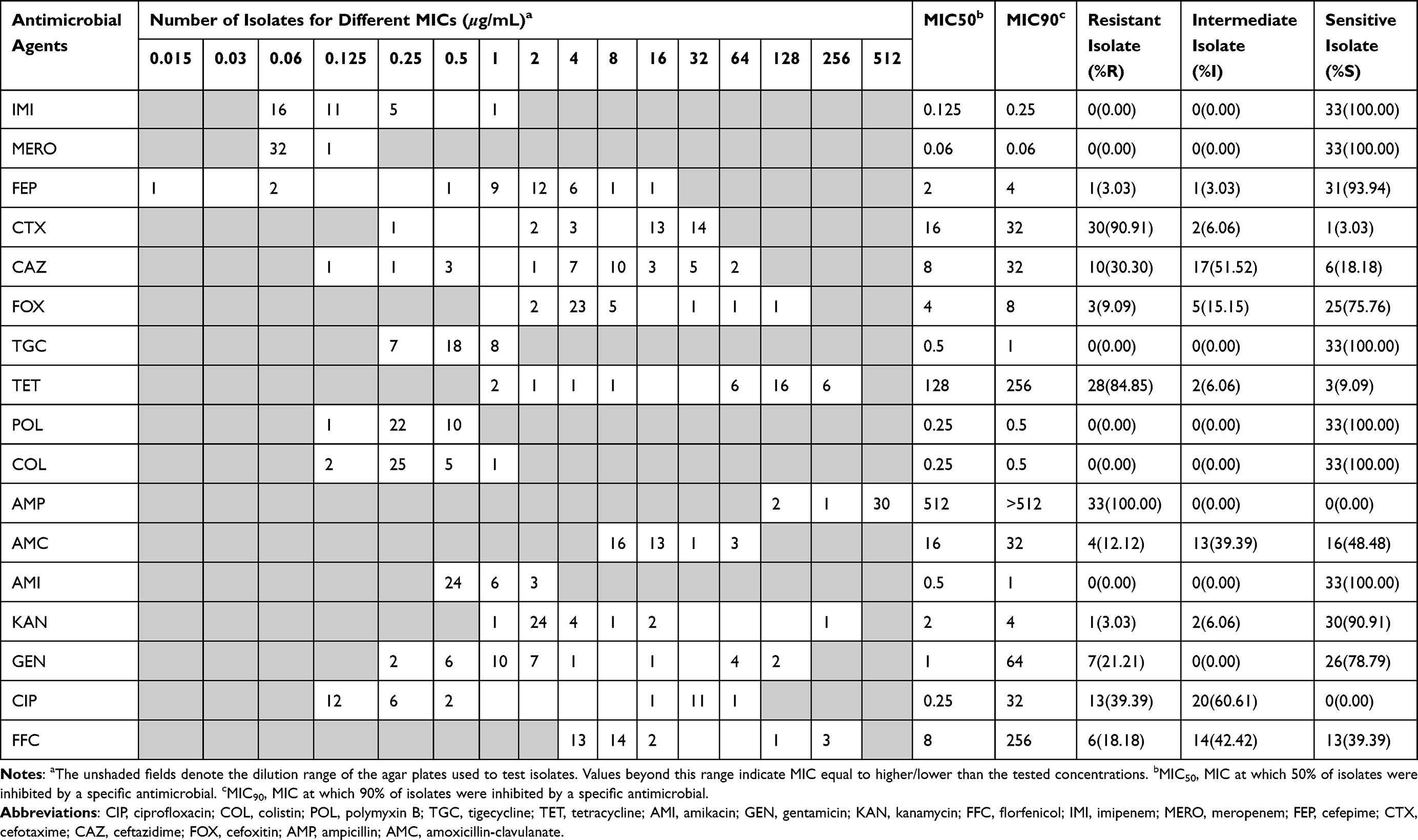 |
Table 1 MIC Distribution of 33 ESBL-Producing E. coli Isolates Against 17 Antimicrobial Agents |
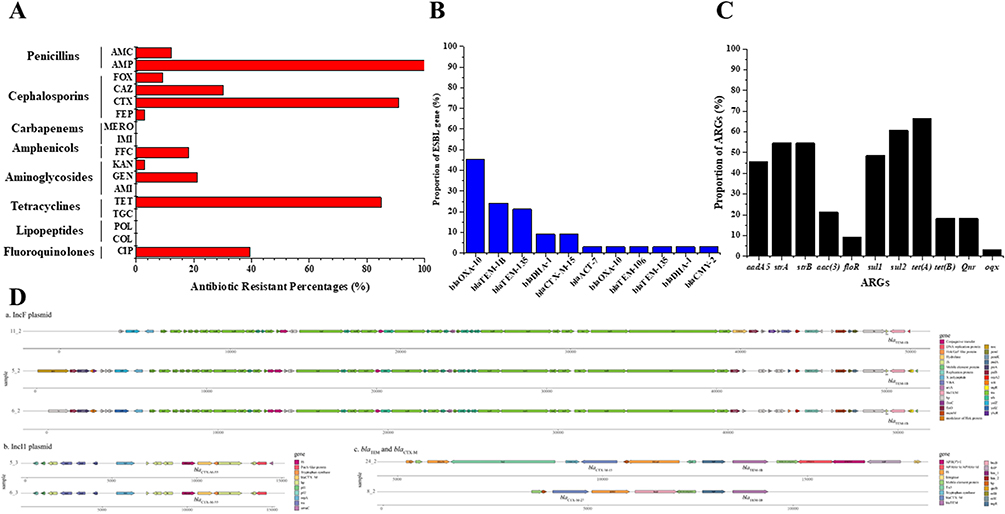 |
Figure 1 Antimicrobial resistance genes and mobile genetic elements in ESBL-EC isolates. (A) Proportions of antimicrobial resistance genes (ARGs) and (B) Proportions of mobile genetic elements (MGEs); (C) Schematic representation of ARGs and MGEs in five isolates. (D) Genetic context of typical ESBL-EC isolates. |
Antibiotic Resistome Characterization of ESBL-EC Isolates
We confirmed the presence of the ESBL genes in each isolate by using WGS. β-lactam resistance genes were observed in all isolates, which verified our ESBL phenotypic screen (Figure 2). Among the 33 ESBL-producing E. coli strains, 28 strains (84.85%) carried the blaCTX-M genes, 10 strains (30.30%) carried the blaTEM genes, and 4 strains (12.12%) carried the blaSHV genes. In addition to the three most common β-lactamase genes, blaCMY-2, blaOXA, and blaDHA were also observed. The most prevalent blaCTX-M genes detected were blaCTX-M-27 (15/33, 45.45%), blaCTM-M-14 (7/33, 21.21%), blaCTM-M-55 (3/33, 9.09%), and blaCTX-M-15 (3/33, 9.09%) (Figure 1B). The ESBL-EC isolates were classified into 14 different STs, with the predominant STs being ST131 (n = 10, 30.3%) ST1193 (n = 6, 18.2%), followed by ST69 (n = 3, 9.1%). The rest (11 STs) occurred in one isolate each. Interestingly, a new globally emerging clone ST1193 in ESBL-EC was identified in 6 maternal and neonatal isolates. Among the ST1193 isolates, the most common resistance genes included blaCTX-M-27 (β-lactam resistance) and dfrA17 (trimethoprim resistance). Notably, we found that maternal-neonatal colonization transmission was found in the ST1193 clone strains.
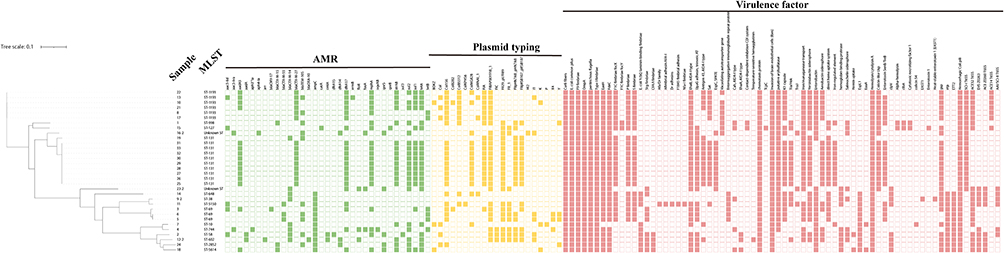 |
Figure 2 Distribution of STs, antimicrobial resistance genes, plasmid typing, and virulence factors in the phylogenetic tree of 33 maternal and neonatal ESBL-EC isolates. Colored strips indicate areas corresponding to the isolates. Green colored cells represent the presence of the genes and white cells represent the absence of the genes. Brown colored cells represent the presence of plasmid typing and white cells represent the absence of the plasmids. |
A total of 11 additional resistant genes were detected followed by the β-lactam resistance genes. The detection rate of tetracycline-resistant gene tetA presented was the highest (66.67%), followed by another tetracycline-resistant gene tetB (18.18%). Furthermore, sulfonamide-resistant genes sul1 and sul2 were conserved in 48.48% and 60.61% of the isolates, respectively. The most common aminoglycoside-resistant genes were aadA5 (45.45%), strA (54.55%), strB (54.55%), and aac(3) (21.21%), respectively. Notably, several isolates were found to carry the amphenicol-resistant gene floR (n = 3), fluoroquinolone resistant genes qnr (n = 6) and oqx (n = 1) (Figure 1C).
Then we analyzed the genetic context of ESBL-EC isolates by examining the contigs to determine the chromosomal or plasmid environment. Isolates 5, 6, and 11 (carrying blaTEM-1B) were located on the IncF plasmid, and the regulatory genetic element IS1 was located the downstream of blaTEM-1B. Isolates 5 and 6 (carrying blaCTX-M-55) also contained the IncI1 plasmid and ISEcp1 was located the upstream of blaCTX-M-55 (Figure 1D).
Plasmid Typing
Plasmid typing revealed that seven plasmid incompatibility (Inc) groups (IncF1A, IncFIB, Inc FIB, IncCol156, IncColBS512, IncColR, IncFII) were common in these isolates. All ST1193 isolates contained the four incompatibility groups IncCol156, IncColBS512, IncFIA, IncFIB (Figure 2). All ST131 isolates also contained 4 incompatibility groups such as IncFIA, IncFIB, IncFII29, IncFIIP.
Virulence Factors
In order to determine the toxin-producing of these isolates, we analyzed virulence factors at genetic level, including membrane protein gene ompA (33/33, 100%), enterobactin biosynthetic enzymes gene ent (32/33, 97.0%), curli nucleator gene csg (32/33, 97.0%), adherence gene fimH, yag, papB, sfa (31/33, 93.9%; 31/33, 93.9%; 22/33, 66.7%; 2/33, 6.0%), high-pathogenicity island gene ybtA (27/33, 81.8%), iron repressible protein irp (26/33, 78.8%), heme absorption related genes chu (25/33, 75.8%), plasmid-carried enterotoxin gene senB (19/33, 57.6%), secreted autotransporter gene sat (16/33, 48.5%), T3SS effectors esp (12/33, 36.4%), vacuolating autotransporter gene vat (9/33, 27.3%), and hemolysin gene hly (2/33, 6.0%) (Table 2 and Figure 2). Importantly, the high prevalence of ybtA among all ST1193 isolates was interesting, because this gene is correlated with the high-pathogenicity island on virulence.
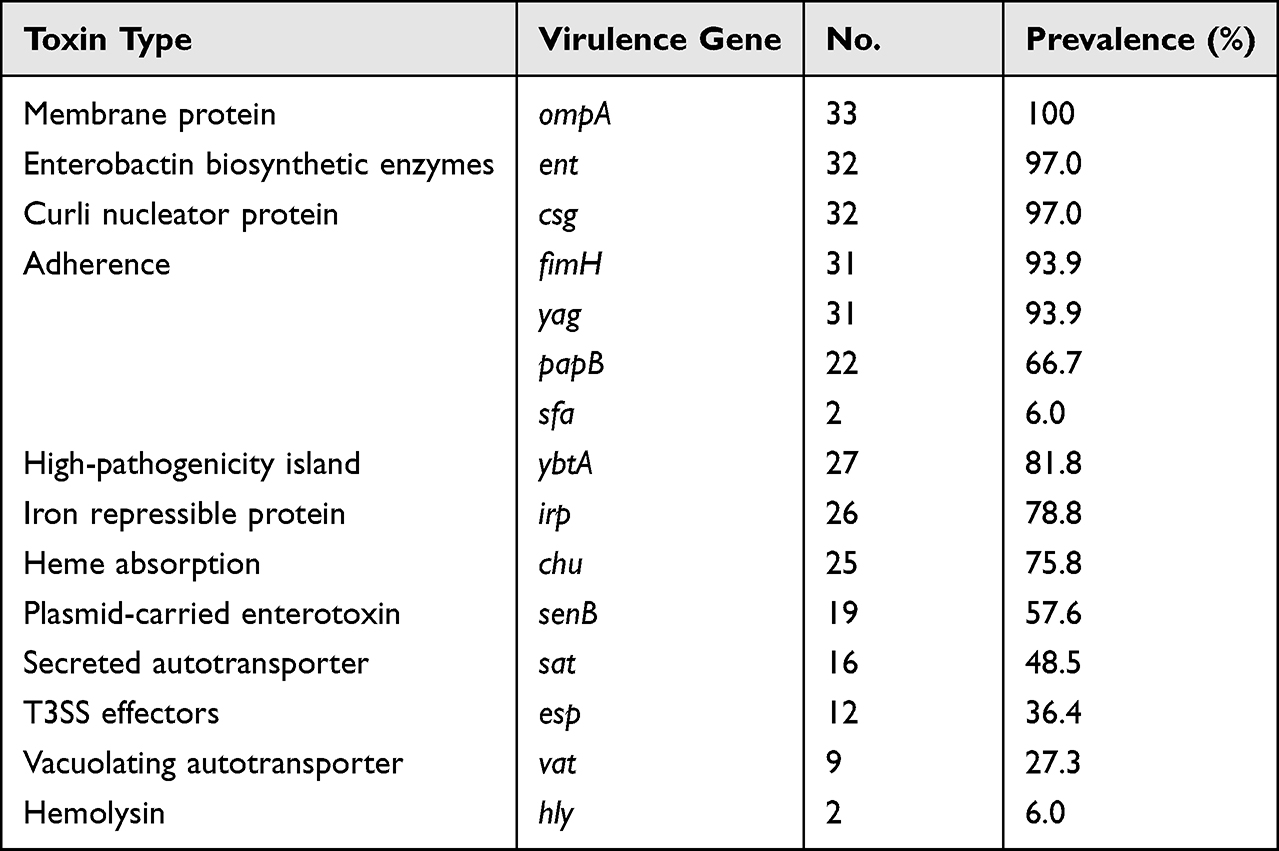 |
Table 2 Virulence Genes Carried by All the 33 ESBL-EC Isolates |
Discussion
The increasing prevalence of ESBL in China poses great challenges to patients, clinicians, and public health.28,29 Over time, the rapid spread of antimicrobial resistance could lead to the emergence and evolution of dominant cloned strains. E. coli ST131 is a classic example of a successful human antimicrobial resistance (AMR) super-clone.30 However, recent studies confirm that the emergence of ST1193 represents the only known large clonal expansion of E. coli isolates apart from the global spread of ST131 over the last two decades.20,31–33 This study reports the isolation of the ST1193 clone in ESBL-EC from pregnant women and newborn in China. Typically, ST131 is associated with blaCTX-M-15, which is currently disseminated globally and identified as the most widespread CTX-M ESBL enzyme worldwide.34 Unlike ST131, the six ST1193 clone from mothers and newborns all carried blaCTX-M-27, which suggest that there might be a relationship of clonal propagation between them.
Phenotypic correlations are generally considered to reflect genotypic correlations. The association between the phenotype and genotype is important because it links together the effects of natural selection. Correlation analysis between genotype and phenotype is the main approach to explain the development of antimicrobial resistance. In this study, all isolates were phenotypically resistant to β-lactams and exhibited various resistant profiles, with all of them being resistant to at least one more antibiotic group, such as aminoglycosides, quinolones, and sulfonamides. The resistance rates of ESBL-EC strains were greater than 80% against the third-generation cephalosporins, cefotaxime, and tetracyclines (Figure 1A). It has been suggested that cefotaxime and tetracycline are not considered effective choices for the treatment of maternal and neonatal infections caused by ESBL-EC. As expected, the ESBL-EC had a high susceptibility rate to imipenem, meropenem, polymyxin, and tigecycline (100%), indicating that carbapenems and polymyxin are considered as last resort antibiotics for the treatment of ESBL-EC infections in China. Notably, the clinical use of colistin should be recommended as an alternative for the treatment ESBL-producing Enterobacteriaceae only when carbapenems cannot be utilized, but this recommendation must be considered carefully in order to avoid the development of bacterial resistance.35,36
We identified 18 STs among 33 ESBL-EC isolates. The most abundant ST type was ST131 (30.3%), followed by ST1193 (18.2%). A recent study of major MLSTs associated with ESBL-EC harboring CTX-M-14, CTX-M-55, or CTX-M-15 genes in Chinese hospitals showed that the most common STs were ST131 (14.6%), ST1193 (7.9%), ST405 (6.6%), ST69 (6.3%), and ST648 (6.3%).33,37 The ST1193 isolates was 50% resistant to ciprofloxacin. All ST1193 isolates possess an IncF plasmid that is apparently deficient in plasmid transfer due to complete deletion of its transfer locus, which is consistent with the report by Johnson et al.31
The most common virulence factors in our study were ompA, fimH, and ybtA. ompA and fimH are relevant to the pathogenesis of neonatal meningitis by promoting the adhesion and invasion of human brain microvascular endothelial cells. Therefore, they may also play a role in the early stage of neonatal bacteremia pathogenesis, which likely involve in bacterial adhesion and invasion of epithelial barriers. Importantly, all ST1193 isolates contained the high pathogenicity island (HPI) virulence factor ybtA, a transcriptional regulator of the AraC-like family, and the iron master regulator Fur that controlled the transcriptional regulation of the four operons located within the HPI.38 Furthermore, HPI was responsible for the synthesis, secretion, and uptake of the siderophore Yersinia, which was shown to be an important virulence determinant.
The present study has some limitations. Firstly, the study was conducted in a single center in Beijing, China, which may not be representative of the ESBL-EC situation in other regions or countries. Secondly, the sample size is relatively small (33 isolates), which may limit the generalizability of the findings. Thirdly, the study only focused on ESBL-EC isolates from pregnant women and infants and did not include other types of patients or bacteria. Lastly, the study did not investigate the clinical outcomes of the patients infected with ESBL-EC, which may be important for understanding the impact of this pathogen on public health.
Conclusions
This study revealed for the first time the occurrence of ST1193 and mother-to-infant colonization transmission in maternal and neonatal ESBL-EC isolates. There was a high ampicillin-tetracycline-cefotaxime resistance among these ESBL-EC isolates. The blaCTX-M genes were the main ESBL genotype. All ST1193 clones carried blaCTX-M-27 and were loaded as IncF-type plasmids. In addition, our study demonstrated that the whole genome sequencing could be used to accurately predict antimicrobial resistance determinants and monitor the epidemiology of clinically significant nosocomial clones. The relationship of clonal propagation between blaCTX-M-27 and ST1193 clone remains to be further studied.
Ethical Approval
The studies involving human participants have been reviewed and ethical approval was approved by the Ethics Committee of the Beijing Obstetrics and Gynecology Hospital of Capital Medical University (No. 2023-KY-002-01). Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements. In this study, we investigated the transmission characteristics and antimicrobial resistance of the ESBL-EC strains isolated from patient samples (blood, urine, or incision secretions). The samples could not identify the patient’s identity information, and did not involve the patient’s personal privacy or commercial interests. The content of the article only analyzed the situation of the strain (including drug resistance and genotyping), and did not analyze the situation related to the patient. The risk of this study is minimal, and it is an investigation of the antimicrobial resistance of pregnant women and newborns infected with ESBL-EC strains. We believe that informed consent can be waived, and we also explained it in the ethical review report.
Funding
This work was supported by grants from the Key Research and Development Program of Beijing (Z211100007021009) and Beijing Natural Science and Foundation (Grant No. 6222017).
Disclosure
The authors declare that they have no conflict of interest.
References
1. Madec JY, Haenni M, Nordmann P, Poirel L. Extended-spectrum beta-lactamase/AmpC- and carbapenemase-producing Enterobacteriaceae in animals: a threat for humans? Clin Microbiol Infect. 2017;23:826–833. doi:10.1016/j.cmi.2017.01.013
2. Lukac PJ, Bonomo RA, Logan LK. Extended-spectrum β-lactamase-producing Enterobacteriaceae in children old foe, emerging threat. Clin Infect Dis. 2015;60:1389–1397. doi:10.1093/cid/civ020
3. Neemann K, Olateju EK, Izevbigie N, et al. Neonatal outcomes associated with maternal recto-vaginal colonization with extended-spectrum beta-lactamase producing Enterobacteriaceae in Nigeria: a prospective, cross-sectional study. Clin Microbiol Infect. 2020;26:463–469. doi:10.1016/j.cmi.2019.07.013
4. Birgy A, Mariani-Kurkdjian P, Bidet P, et al. Characterization of extended-spectrum-beta-lactamase-producing Escherichia coli strains involved in maternal-fetal colonization: prevalence of E. coli ST131. J Clin Microbiol. 2013;51:1727–1732. doi:10.1128/JCM.03255-12
5. Day MJ, Hopkins KL, Wareham DW, et al. Extended-spectrum β-lactamase-producing Escherichia coli in human-derived and foodchain-derived samples from England, Wales, and Scotland: an epidemiological surveillance and typing study. Lancet Infect Dis. 2019;19:1325–1335. doi:10.1016/s1473-3099(19)30273-7
6. Chong Y, Shimoda S, Shimono N. Current epidemiology, genetic evolution and clinical impact of extended-spectrum beta-lactamase-producing Escherichia coli and Klebsiella pneumoniae. Infect Genet Evol. 2018;61:185–188. doi:10.1016/j.meegid.2018.04.005
7. Danino D, Melamed R, Sterer B, et al. Mother-to-child transmission of extended-spectrum-beta-lactamase-producing Enterobacteriaceae. J Hosp Infect. 2018;100:40–46. doi:10.1016/j.jhin.2017.12.024
8. Bulabula ANH, Dramowski A, Mehtar S. Maternal colonization or infection with extended-spectrum beta-lactamase-producing Enterobacteriaceae in Africa: a systematic review and meta-analysis. Int J Infect Dis. 2017;64:58–66. doi:10.1016/j.ijid.2017.08.015
9. Jalilian N, Kooshkiforooshani M, Ahmadi S, Nankali A. Colonisation with extended-spectrum beta-lactamase-producing Enterobacteriaceae in pregnant/post-partum women: systematic review and meta-analysis. J Glob Antimicrob Resist. 2019;19:338–347. doi:10.1016/j.jgar.2019.06.010
10. Karanika S, Karantanos T, Arvanitis M, Grigoras C, Mylonakis E. Fecal colonization with extended-spectrum beta-lactamase-producing Enterobacteriaceae and risk factors among healthy individuals: a systematic review and metaanalysis. Clin Infect Dis. 2016;63:310–318. doi:10.1093/cid/ciw283
11. Titelman E, Hasan CM, Iversen A, et al. Faecal carriage of extended-spectrum beta-lactamase-producing Enterobacteriaceae is common 12 months after infection and is related to strain factors. Clin Microbiol Infect. 2014;20:O508–515. doi:10.1111/1469-0691.12559
12. Overdevest I, Haverkate M, Veenemans J, et al. Prolonged colonisation with Escherichia coli O25:ST131 versus other extended-spectrum beta-lactamase-producing E. coli in a long-term care facility with high endemic level of rectal colonisation, the Netherlands, 2013 to 2014. Euro Surveill. 2016;21. doi:10.2807/1560-7917.ES.2016.21.42.30376
13. van den Bunt G, Liakopoulos A, Mevius DJ, et al. ESBL/AmpC-producing Enterobacteriaceae in households with children of preschool age: prevalence, risk factors and co-carriage. J Antimicrob Chemother. 2017;72:589–595. doi:10.1093/jac/dkw443
14. Tacconelli E, Carrara E, Savoldi A, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. 2018;18:318–327. doi:10.1016/s1473-3099(17)30753-3
15. Miao Z, Li S, Wang L, Song W, Zhou Y. Antimicrobial resistance and molecular epidemiology of esbl-producing Escherichia coli isolated from outpatients in town hospitals of Shandong Province, China. Front Microbiol. 2017;8:63. doi:10.3389/fmicb.2017.00063
16. Coque TM, Baquero F, Canton R. Increasing prevalence of ESBL-producing Enterobacteriaceae in Europe. Euro Surveill. 2008;13:19044.
17. Bush K, Jacoby GA. Updated functional classification of beta-lactamases. Antimicrob Agents Chemother. 2010;54:969–976. doi:10.1128/AAC.01009-09
18. Ben-Ami R, Rodríguez‐Baño J, Arslan H, et al. A multinational survey of risk factors for infection with extended-spectrum beta-lactamase-producing Enterobacteriaceae in nonhospitalized patients. Clin Infect Dis. 2009;49:682–690. doi:10.1086/604713
19. Rogers BA, Sidjabat HE, Paterson DL. Escherichia coli O25b-ST131: a pandemic, multiresistant, community-associated strain. J Antimicrob Chemother. 2011;66:1–14. doi:10.1093/jac/dkq415
20. Tchesnokova VL, Rechkina E, Larson L, et al. Rapid and extensive expansion in the United States of a new multidrug-resistant Escherichia coli clonal group, sequence type 1193. Clin Infect Dis. 2019;68:334–337. doi:10.1093/cid/ciy525
21. Platell JL, Trott DJ, Johnson JR, et al. Prominence of an O75 clonal group (clonal complex 14) among non-ST131 fluoroquinolone-resistant Escherichia coli causing extraintestinal infections in humans and dogs in Australia. Antimicrob Agents Chemother. 2012;56:3898–3904. doi:10.1128/AAC.06120-11
22. Sands K, Carvalho MJ, Portal E, et al. Characterization of antimicrobial-resistant Gram-negative bacteria that cause neonatal sepsis in seven low- and middle-income countries. Nat Microbiol. 2021;6:512–523. doi:10.1038/s41564-021-00870-7
23. Solomon S, Akeju O, Odumade OA, et al. Prevalence and risk factors for antimicrobial resistance among newborns with gram-negative sepsis. PLoS One. 2021;16:e0255410. doi:10.1371/journal.pone.0255410
24. CLSI. Performance Standards for Antimicrobial Susceptibility Testing. In: CLSI Supplement M100.
25. European Committee on Antimicrobial Susceptibility Testing. Routine and extended internal quality control for MIC determination and disk diffusion as recommended by EUCAST. Version 7.0. 2017.
26. Bankevich A, Nurk S, Antipov D, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19:455–477. doi:10.1089/cmb.2012.0021
27. Treangen TJ, Ondov BD, Koren S, Phillippy AM. The Harvest suite for rapid core-genome alignment and visualization of thousands of intraspecific microbial genomes. Genome Biol. 2014;15:524. doi:10.1186/s13059-014-0524-x
28. Jones RN, Castanheira M, Hu B, et al. Update of contemporary antimicrobial resistance rates across China: reference testing results for 12 medical centers. Diagn Microbiol Infect Dis. 2011;77:258–266. doi:10.1016/j.diagmicrobio.2013.07.003
29. Quan J, Zhao D, Liu L, et al. High prevalence of ESBL-producing Escherichia coli and Klebsiella pneumoniae in community-onset bloodstream infections in China. J Antimicrob Chemother. 2017;72:273–280. doi:10.1093/jac/dkw372
30. Nicolas-Chanoine MH, Bertrand X, Madec JY. Escherichia coli ST131, an intriguing clonal group. Clin Microbiol Rev. 2014;27:543–574. doi:10.1128/CMR.00125-13
31. Johnson TJ, Elnekave E, Miller EA, et al. Phylogenomic analysis of extraintestinal pathogenic Escherichia coli sequence type 1193, an emerging multidrug-resistant clonal group. Antimicrob Agents Chemother. 2019;63. doi:10.1128/AAC.01913-18
32. Ding Y, Zhang J, Yao K, Gao W, Wang Y. Molecular characteristics of the new emerging global clone ST1193 among clinical isolates of Escherichia coli from neonatal invasive infections in China. Eur J Clin Microbiol Infect Dis. 2021;40:833–840. doi:10.1007/s10096-020-04079-0
33. Xia S, Fan X, Huang Z, et al. Dominance of CTX-M-type extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli isolated from patients with community-onset and hospital-onset infection in China. PLoS One. 2014;9:e100707. doi:10.1371/journal.pone.0100707
34. Overdevest I, Willemsen I, Rijnsburger M, et al. Extended-spectrum beta-lactamase genes of Escherichia coli in chicken meat and humans, The Netherlands. Emerg Infect Dis. 2011;17:1216–1222. doi:10.3201/eid1707.110209
35. Katip W, Yoodee J, Uitrakul S, Oberdorfer P. Efficacy of loading dose colistin versus carbapenems for treatment of extended spectrum beta lactamase producing Enterobacteriaceae. Sci Rep. 2021;11:18. doi:10.1038/s41598-020-78098-4
36. Katip W, Rayanakorn A, Oberdorfer P, Taruangsri P, Nampuan T. Short versus long course of colistin treatment for carbapenem-resistant A. baumannii in critically ill patients: a propensity score matching study. J Infect Public Health. 2023;16:1249–1255. doi:10.1016/j.jiph.2023.05.024
37. Gu S, Lai J, Kang W, et al. Drug resistance characteristics and molecular typing of Escherichia coli isolates from neonates in class A tertiary hospitals A multicentre study across China. J Infect. 2022;85:499–506. doi:10.1016/j.jinf.2022.09.014
38. Anisimov R, Brem D, Heesemann J, Rakin A. Transcriptional regulation of high pathogenicity island iron uptake genes by YbtA. Int J Med Microbiol. 2005;295:19–28. doi:10.1016/j.ijmm.2004.11.007
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Prevalence of Extended-Spectrum β-Lactamase and Carbapenemase Producers of Gram-Negative Bacteria, and Methicillin-Resistant Staphylococcus aureus in Isolates from Diabetic Foot Ulcer Patients in Ethiopia
Woldeteklie AA, Kebede HB, Abdela AA, Woldeamanuel Y
Infection and Drug Resistance 2022, 15:4435-4441
Published Date: 11 August 2022
The Epidemiology, Associated Factors and Bacterial Profile of Asymptomatic Bacteriuria in Pregnant Women: A Retrospective Chart Review Study in Saudi Arabia
AlShamlan NA, AlOmar RS, Aldossary R, Alahmari M, Alghamdi A, AlGhamdi M, Alkanaan N, AlReedy AH, AlOtaibi AS, Alghamdi NS
International Journal of Women's Health 2022, 14:1749-1759
Published Date: 16 December 2022
Antibiograms of Gut Flora of Poultry Farms Workers Reveal Higher Resistance Levels as Compared to Non-Workers
Saeed MQ, Aziz M, Afzal RK, Hussain A, Manzoor H, Rasul S, Ruby T, Akrem A, Masud S, Arshad A, Latif A, Yousif M, Aali H, Fatima M, Bhutta MM, Khan AA
Infection and Drug Resistance 2022, 15:7699-7705
Published Date: 29 December 2022
Comparison of Epidemiological Characteristics Between ESBL and Non-ESBL Isolates of Clinically Isolated Escherichia coli from 2014 to 2022: A Single-Center Study

Zhang W, Wang Q, Zhang L, Wu J, Liu J, Lu C, Wang X, Zhang Z
Infection and Drug Resistance 2023, 16:5185-5195
Published Date: 9 August 2023
A Systematic review on Prevalence, Serotypes and Antibiotic resistance of Salmonella in Ethiopia, 2010–2022
Kahsay AG, Dejene TA, Kassaye E
Infection and Drug Resistance 2023, 16:6703-6715
Published Date: 13 October 2023