Back to Journals » Infection and Drug Resistance » Volume 12
Emergence of novel methicillin-resistant Staphylococcus aureus strains in a tertiary care facility in Riyadh, Saudi Arabia
Authors Senok A ![]() , Somily AM
, Somily AM ![]() , Nassar R, Garaween G, Kim Sing G, Müller E, Reissig A, Gawlik D, Ehricht R, Monecke S
, Nassar R, Garaween G, Kim Sing G, Müller E, Reissig A, Gawlik D, Ehricht R, Monecke S
Received 9 June 2019
Accepted for publication 10 August 2019
Published 3 September 2019 Volume 2019:12 Pages 2739—2746
DOI https://doi.org/10.2147/IDR.S218870
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eric Nulens
Abiola Senok1, Ali M Somily2, Rania Nassar1, Ghada Garaween3, Garwin Kim Sing3, Elke Müller4,5, Annett Reissig4,5, Darius Gawlik6, Ralf Ehricht4,5, Stefan Monecke4,5,7
1Department of Basic Medical Sciences, College of Medicine, Mohammed Bin Rashid University of Medicine and Health Sciences, Dubai, United Arab Emirates; 2Department of Pathology and Laboratory Medicine, College of Medicine, King Khalid University Hospital and King Saud University, Riyadh, Saudi Arabia; 3Department of Microbiology and Immunology, College of Medicine, Alfaisal University, Riyadh, Saudi Arabia; 4InfectoGnostics Research Campus Jena, Jena, Germany; 5Leibniz Institute of Photonic Technology (IPHT), Jena, Germany; 6PTC - Phage Technology Center GmbH, Bönen, Germany; 7Medical Faculty “Carl Gustav Carus”, Institute for Medical Microbiology and Hygiene, Technische Universität Dresden, Dresden, Germany
Correspondence: Abiola Senok
College of Medicine, Mohammed Bin Rashid University of Medicine and Health Sciences, P.O. Box 505055, Dubai, United Arab Emirates
Tel +971 4 383 8717
Email [email protected]
Purpose: There is a need for continuous surveillance of methicillin-resistant Staphylococcus aureus (MRSA) to identify emergence of new strains. We hypothesize that MRSA strains are evolving with ongoing acquisition of SCCmec elements. This study was carried out to evaluate the evolution of MRSA at a tertiary care facility in Saudi Arabia.
Methods: MRSA isolates associated with invasive clinical infection, which were identified in 2017 at the microbiology laboratory, King Khalid University Hospital (KKUH) in Riyadh, Saudi Arabia, were studied. The molecular characterization of isolates was carried out using StaphyType DNA microarray (Alere Technologies GmbH/Abbott, Jena, Germany).
Results: The 125 MRSA isolates studied belonged to 18 clonal complexes (CC) which were distributed into 32 strain assignments. The predominant CC were CC5 (n=30), CC6 (n=17), CC80 (n=13), CC22 (n=12), CC361 (n=12). The findings demonstrated the first identification of CC152, CC361 and CC1153 MRSA as well as ST5-MRSA-[I+fus], “Geraldine Clone”, CC6-MRSA-IV (PVL+) and CC88-MRSA-V (PVL+), WA MRSA-117 in Saudi Arabia. Four novel variants were identified: CC5-MRSA-[VI+fus+tirS], CC22-MRSA-[V/VT+fus](PVL+), CC152-MRSA-[V+fus](PVL+) and CC361-MRSA-[VT+fus]. Fifty-four isolates (n/N=54/125; 43.2%) including the novel strains carried the Q6GD50 SCCfusC gene while the Panton-Valentine leukocidin genes were present in 30.4% (n/N=38/125).
Conclusion: The findings demonstrate an expanding MRSA repertoire in our setting including emergence of previously unreported clonal complexes and novel strains. The high carriage of fusC gene suggests a role for fusidic acid misuse in driving the evolution of the MRSA genome and underscores the need for increased monitoring of antibiotic use.
Keywords: DNA microarray, fusidic acid, clonal complex, panton-valentine leukocidin
Introduction
Methicillin-resistant S. aureus (MRSA) which was first described in the 1960s remains a major etiological agent of community-acquired and nosocomial infections. MRSA infections contribute to the burden of health care accounting for significant morbidity, mortality with significant economic consequences.1 The epidemiology of MRSA is dynamic and ever evolving with community-acquired MRSA lineages (CA-MRSA) being identified as increasingly relevant also in the health care sector.2,3
Antibiotic resistance in MRSA is mediated by the presence of modified penicillin-binding protein (PBP2a), which is encoded for by alleles of the mecA gene located on SCCmec (staphylococcal cassette chromosome mec) element. In addition to the mecA gene, this large, genetic element also harbors regulatory genes, recombinase genes and a variety of accessory genes. The nomenclature of the SCCmec elements is based on the composition of the regulatory and recombinase genes as well as those of additional genes within the mec complex which encode for other resistance mechanisms or virulence determinants.2,4 Pseudo-SCCmec elements which are truncated with absence of the ccr recombinase genes as well as SCC elements lacking mec genes but harboring other markers such as the fusidic acid resistance gene fusC have been described in the literature.5,6 High-resolution typing utilizing newly developed probes been used to differentiate SCCmec types into various subtypes.3,6
In Saudi Arabia, MRSA infections contribute significantly to the burden of health care delivery with increasing occurrence of CA-MRSA lineages causing nosocomial infections.7,8 Recently reported work showed the presence of a high degree of diversity and an emergence of both pandemic and rare MRSA strains among isolates obtained from 2009 to 2015 at King Khaled University Hospital (KKUH), Riyadh, Saudi Arabia.8 Further work also showed that MRSA colonizing health care workers at the facility were of similar population structure as those identified in patients.9
The changing epidemiology of MRSA necessitates the continuous surveillance particularly in Saudi Arabia and the Arabian Gulf region where a dynamic population movement exists. The large expatriate population, increasing numbers of tourists and, specifically in Saudi Arabia, the annual influx of millions of Hajj pilgrims makes it plausible that the Arabian Gulf region might be pivotal in the global transmission of MRSA clones. This need for surveillance is underscored by the recent report of the diversity of CC22-MRSA-IV strains across the Arabian Gulf region.3 Using high-resolution typing, at least six CC22-MRSA-IV strains differing in SCCmec subtypes and toxin carriage were identified from three countries in the region.3 We speculate that new MRSA strains might continuously evolve in our health care facilities and the acquisition of SCCmec elements might occur more commonly than appreciated. The present study was carried out to evaluate the evolution of MRSA at a tertiary care center in the capital of Saudi Arabia, Riyadh, and findings demonstrate the emergence of novel variants and previously unidentified MRSA strains.
Materials and methods
Specimen collection and bacterial strains
The study was carried out at the King Khalid University Hospital (KKUH) in Riyadh, Saudi Arabia. Ethical approval was obtained from the hospital ethics committee. MRSA isolates associated with invasive clinical infection identified in 2017 were studied. Bacterial identification, confirmation of methicillin resistance and antibiotic susceptibility testing were performed in accordance with Clinical and Laboratory Standards Institute guidelines as previously described.8
DNA microarray for molecular characterization
All isolates were characterized using microarray assays for S. aureus typing and characterization of SCC elements (Alere Technologies GmbH/Abbott, Jena, Germany). The probes, primers and procedures used in the assay for detection of species markers, virulence genes, resistance genes and SCCmec subtyping have been described previously.6,10 Assays were carried out in accordance with manufacturer provided protocols. The analysis of presence or absence of target gene, strain and clonal complex assignment and SCCmec subtype was carried out as previously described.6,10
Results
A total of 125 MRSA isolates associated with clinical infections obtained in 2017 at KKUH microbiology laboratory were studied. The isolates were obtained from blood (n=25), respiratory aspirates (n=20) and wound swabs (n=80). Based on DNA microarray analysis, these 125 MRSA isolates were grouped into 18 clonal complexes (CC) which were distributed into 32 strain assignments based on CC affiliation, toxin gene carriage and SCCmec subtype (Table 1). The predominant CC were CC5 (n=30), CC6 (n=17), CC80 (n=13), CC22 (n=12), CC361 (n=12) (Table 1). The findings demonstrated the first identification of CC152, CC361 and CC1153 MRSA in Saudi Arabia. Furthermore, we report the first identification of ST5-MRSA-[I+fus], “Geraldine Clone”, CC6-MRSA-IV (PVL+) and CC88-MRSA-V (PVL+), WA MRSA-117 in Saudi Arabia (Table 1). Three isolates identified as MRSA phenotypically were genotypically characterized as CC2250 S. argenteus.
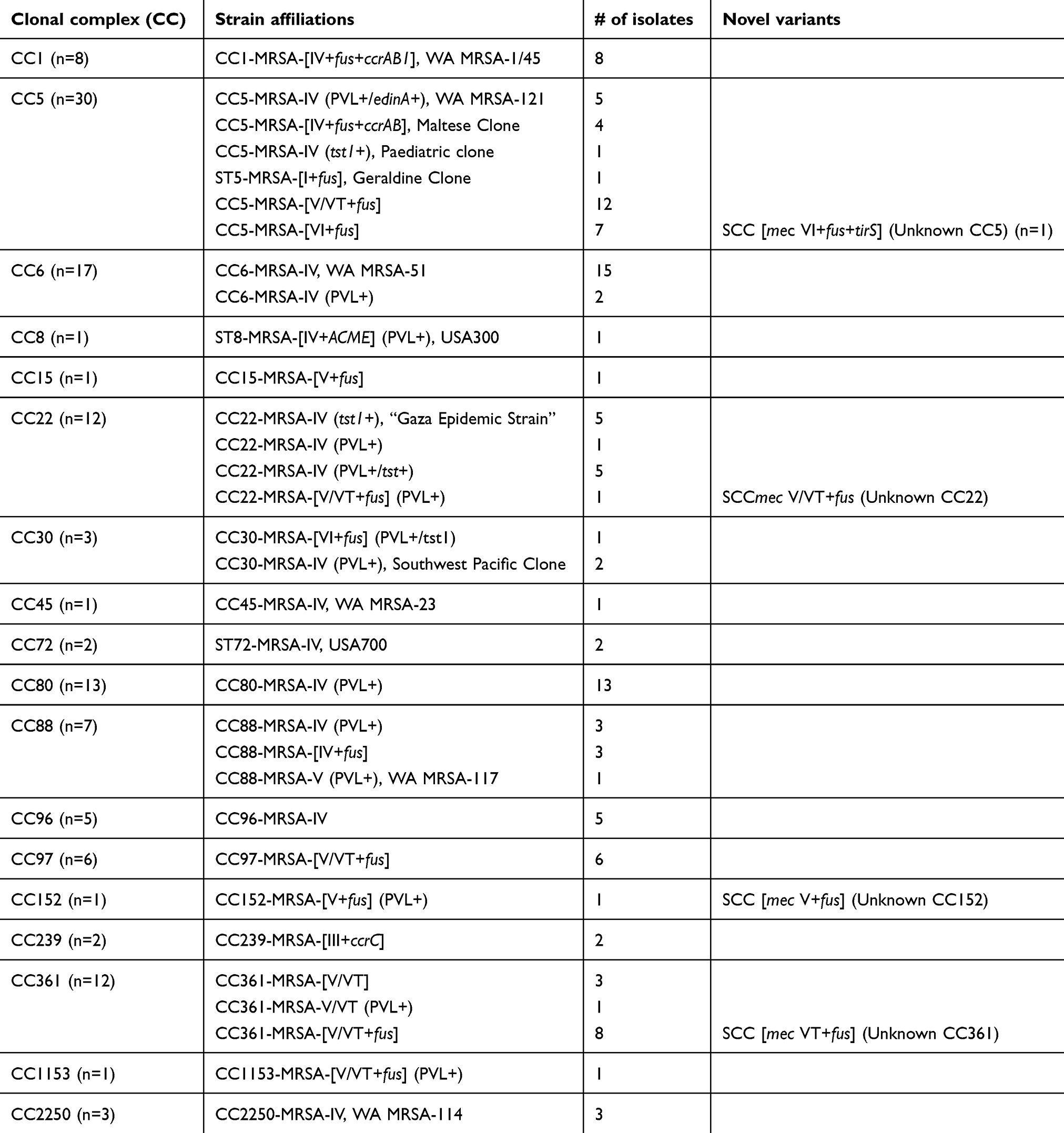 |
Table 1 Distribution of clonal complex and strain affiliations |
Four novel variant of MRSA strains were identified. These strains belong to CC5, CC22, CC152 and CC361. Table 2 shows the molecular characterization of these four MRSA strains. Except for the CC5 strain with SCCmecVI, all others harbored SCCmecV. Almost half of the total isolates (n/N=54/125; 43.2%) including all the novel strains carried the Q6GD50 (fusC, SCC-borne fusidic acid resistance) (Table 2). None of the isolates harbored the vancomycin or mupirocin resistance genes. Table 3 shows the distribution of carriage of virulence and antibiotic resistance genes among all 125 isolates. The Panton-Valentine leukocidin (pvl) genes were found in 30.4% (n/N=38/125) of isolates. The predominant virulence genes identified were sak and scn which were present in 96% of isolates.
 |
Table 2 Characterization of novel variants of methicillin-resistant Staphylococcus aureus strains |
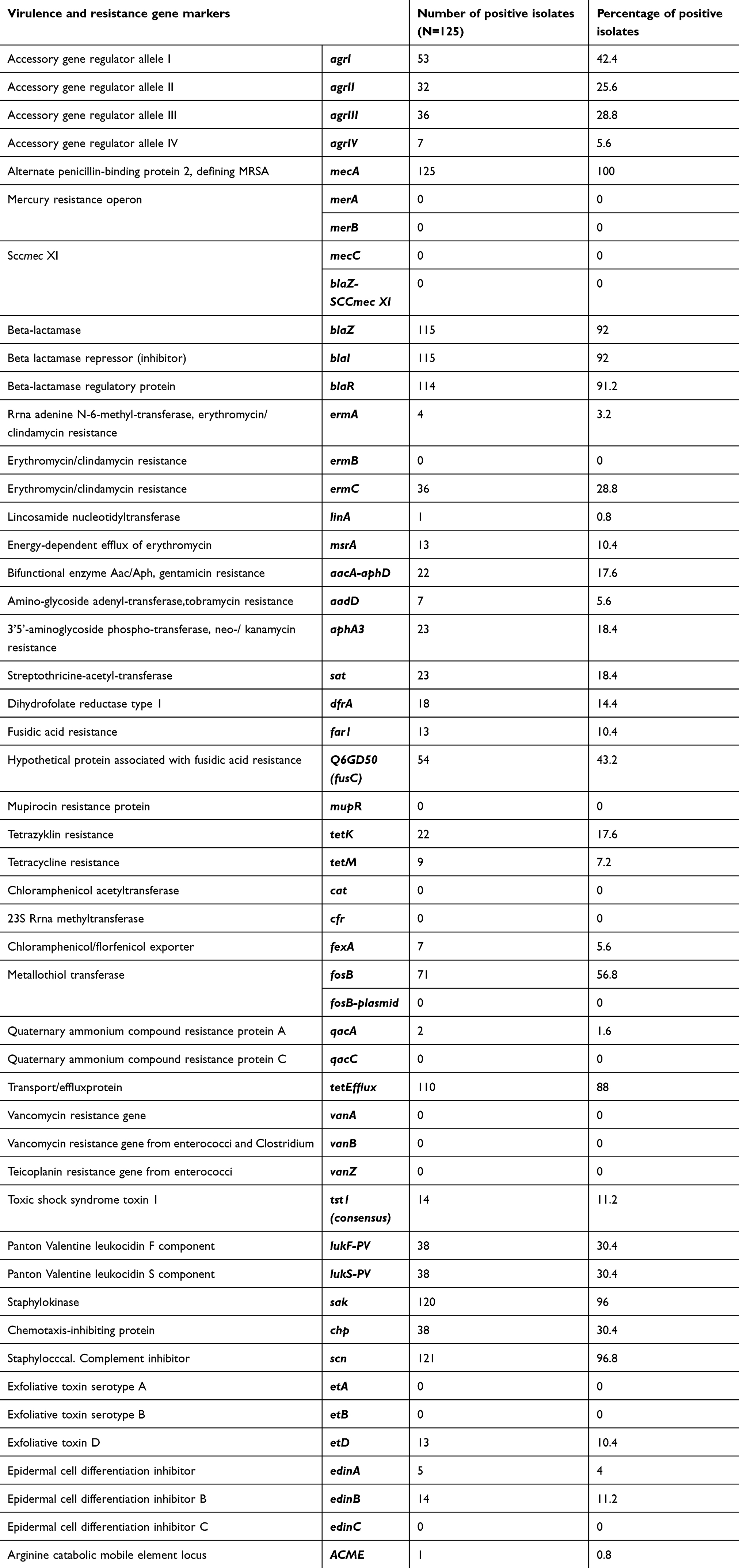 |
Table 3 Distribution of virulence and resistance gene markers |
Discussion
The identification of MRSA strains which have not been previously reported from Saudi Arabia and novel variants of MRSA strains demonstrate the continuing evolution of MRSA in our setting. Over 40% of the MRSA isolates identified harbored the fusidic acid resistance gene fusC presumably as additional payload on the SCCmec genetic element. A similar high occurrence of co-carriage of fusC and SCCmec elements has been shown in MRSA isolates from Kuwait.5 It is of interest that all the novel variants identified harbored the SCCmec/fusC element. This evolution of SCCmec/fusC composite element is probably due to a survival benefit for the strains driven in part by misuse of fusidic acid given its availability in our setting as an over-the-counter medication in the community. The presence of composite SCCmec/fusC element and chimeras of multiple progenitor cassettes has been demonstrated in fusidic-acid-resistant MRSA strains in the United Kingdom.11 It was suggested that the occurrence of strains harboring genetic elements which include SCCmec, antibiotic resistance and virulence genes may be indicative of novel adaptive mechanisms in MRSA.11 Indeed, this adaptive mechanism could drive the emergence of new MRSA strains such of the novel variants of MRSA strains shown in this study.
CC5-MRSA is a common clonal complex which encompasses a large variety of MRSA strains.10 CC5-MRSA is prevalent in the Arabian Gulf region with CC5-MRSA-IV (PVL+/edinA+), WA MRSA-121, CC5-MRSA-[IV+fus+ccrAB], “Maltese Clone” and CC5-MRSA-IV (tst1+), “Pediatric clone” previously identified from Saudi Arabia (including from KKUH), Kuwait and Qatar.5,8,12,13 Although CC5-MRSA-V has been identified in the UAE and Saudi Arabia, the CC5-MRSA-[V/VT+fus] reported in this study had only been described from Kuwait.5,8,13 Our findings demonstrate the emergence of CC5-MRSA-VI in the Arabian Gulf region. CC5-MRSA-VI was first described in Portugal and has been reported in France, Colombia, Argentina and the USA while the PVL-positive CC5-MRSA-VI has been reported from Switzerland.10,14–16 All the CC5-MRSA-VI identified in this study harbored the SCCmec/fusC genetic element. In addition, we describe a novel variant with SCC [mec VI+fus+tirS] harboring the Staphylococcal TIR-protein-binding protein (tirS) as additional payload. The presence of the tirS gene in S. aureus has been associated with enhanced bacterial survival in the host as well as increased virulence through attenuation of the TLR2 immune response.17 The horizontal transfer of this gene among MRSA isolates circulating in our setting is probable as it is localized on a mobile genetic element. This scenario might drive the evolution of MRSA toward emergence of strains with increased bacterial fitness and virulence.
A single isolate of the ST5-MRSA-[I+fus] “Geraldine Clone” was identified and to the best of our knowledge, this is the first description of this strain in the Arabian Gulf region. The ST5-MRSA-[I+fus] “Geraldine Clone” was first described in 2003 in France where it is a prevalent clone associated with outbreaks, particularly in the neonatal setting.18,19 In 2012, a single ST5-MRSA-[I+fus] “Geraldine Clone” was reported in Saxony, Germany, in a patient with a history of foreign travel.6 A travel link could also explain the emergence of this strain in our setting. Although verification of a travel history in this instance was not possible due to lack of access to patient data, the large expatriate population and common occurrences of travel for medical reasons in our setting makes this highly plausible.
CC152-MRSA-V has been reported from Germany, Australia, Sweden, Switzerland and Kuwait.5,10 In keeping with reported literature on CC152-MRSA-V, our isolate harbored the pvl and edinB genes, and lacked enterotoxin genes.10 The CC152-MRSA-[V+fus] (PVL+) identified in this study is very similar to that recently reported as a novel variant with the carriage of the SCCfusC element. However, our isolate had the fibronectin-binding protein genes (fnbA, fnbB) which was not present in the previously described isolates.5
The findings in this study demonstrate the first identification of CC361 strains in Saudi Arabia. CC361-MRSA-V has been described in the UAE and recently CC361-MRSA-[V/VT+fus] was reported in Kuwait.5,10 The CC361-MRSA-[V/VT+fus] harboring the SCCmec VT+fus genetic element identified in this study is similar to the novel variant recently described from Kuwait, thus indicating its second description in our region.5
We had previously reported CC1153-MSSA as nasal colonizer in health care workers at KKUH but this is the first identification of CC1153-MRSA in this facility.9 In our region, the two CC1153-MRSA isolates reported have been from Kuwait.5,20 Both isolates were pvl+ while one carried the SCCmec-V and the other had SCCmec I+fus. Similar to these two, our isolate is pvl+. Thus, all three CC1153 strains identified so far in the region show variability in their SCCmec types/ subtypes. It has been suggested that emergence of novel strains as reported for CC5-MRSA-IV, CC22-MRSA-IV and PVL-positive CC30-MRSA-IV might be due to the independent acquisition of SCCmec types/subtypes by the parental MSSA strains.6 This observation with CC1153-MRSA supports this premise for the emergence of novel MRSA strains.
CC22 is a widespread clonal group and six distinguishable strains based on SCCmec IV subtypes and virulence factors have been reported from this region.3 In this study, three of the previously reported CC22-MRS-IV strains were identified as well as a novel variant of CC22-MRSA-V characterized by carriage of the SCCmec V/VT+fus and pvl+. Previously reported CC22-MRSA-[V+fus] from Saxony, Germany, were negative for pvl gene.6 In Saudi Arabia, a single isolate of CC22-MRSA-V which did not carry the pvl gene was reported as a nasal colonizer.21 The finding of this novel CC22-MRSA variant suggests the expanding repertoire of CC22 strains in this region.
S. argenteus is a newly identified staphylococcus species previously misidentified as S. aureus.22 The isolates identified in this study were phenotypically classified as S. aureus but found to be CC2250 S. argenteus on genotyping. This represents the second report of S. argenteus CC2250-MRSA-IV, WA MRSA-114 in Saudi Arabia having been previously identified from clinical infection in this health care facility.8 Indeed, S. argenteus has been shown to be of clinical relevance and isolates belonging to CC75, CC1233, CC2198, CC2250 and CC2854 have been described.22,23
Conclusion
Our findings demonstrate the emergence of novel variants of MRSA strains belonging to CC5, CC22, CC152 and CC361 as well as the first identification of other MRSA strains in Saudi Arabia. These findings are significant and indicative of an expanding repertoire of MRSA strains in our setting. This continued evolution of the MRSA genome arising from acquisition of resistance and virulence genes poses significant challenges for treatment and infection control. The role of fusidic acid misuse in driving this evolution underscores the need for increased monitoring of antibiotic use.
Acknowledgment
This study was partially funded by Strategic Research Grant no. 307191502153 from Alfaisal University, Riyadh, Saudi Arabia.
Disclosure
EM, AR, DG, RE, SM were employees at Alere Technologies GmbH/Abbott, Jena, Germany, at the time the study was carried out. The authors report no other conflicts of interest in this work.
References
1. Reddy PN, Srirama K, Dirisala VR. An update on clinical burden, diagnostic tools, and therapeutic options of Staphylococcus aureus. Infect Dis (Auckl). 2017;10:1179916117703999. doi:10.1177/1179916117703999
2. Deurenberg RH, Stobberingh EE. The molecular evolution of hospital- and community-associated methicillin-resistant Staphylococcus aureus. Curr Mol Med. 2009;9(2):100–115.
3. Senok A, Somily A, Raji A, et al. Diversity of methicillin-resistant Staphylococcus aureus CC22-MRSA-IV from Saudi Arabia and the Gulf region. Int J Infect Dis. 2016;51:31–35. doi:10.1016/j.ijid.2016.08.016
4. Deurenberg RH, Vink C, Kalenic S, Friedrich AW, Bruggeman CA, Stobberingh EE. The molecular evolution of methicillin-resistant Staphylococcus aureus. Clin Microbiol Infect. 2007;13(3):222–235. doi:10.1111/j.1469-0691.2006.01573.x
5. Boswihi SS, Udo EE, Monecke S, et al. Emerging variants of methicillin-resistant Staphylococcus aureus genotypes in Kuwait hospitals. PLoS One. 2018;13(4):e0195933. doi:10.1371/journal.pone.0195933
6. Monecke S, Jatzwauk L, Muller E, et al. Diversity of SCCmec elements in Staphylococcus aureus as observed in South-Eastern Germany. PLoS One. 2016;11(9):e0162654. doi:10.1371/journal.pone.0162654
7. Bukharie HA. Increasing threat of community-acquired methicillin-resistant Staphylococcus aureus. Am J Med Sci. 2010;340(5):378–381. doi:10.1097/MAJ.0b013e3181e95fdc
8. Senok A, Ehricht R, Monecke S, Al-Saedan R, Somily A. Molecular characterization of methicillin-resistant Staphylococcus aureus in nosocomial infections in a tertiary-care facility: emergence of new clonal complexes in Saudi Arabia. New Microbes New Infect. 2016;14:13–18. doi:10.1016/j.nmni.2016.07.009
9. Senok A, Somily A, Raji M, et al. Genotyping of Staphylococcus aureus associated with nasal colonization among healthcare workers using DNA microarray. J Infect Dev Ctries. 2018;12(5):321–325. doi:10.3855/jidc.10328
10. Monecke S, Coombs G, Shore AC, et al. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS One. 2011;6(4):e17936. doi:10.1371/journal.pone.0017936
11. Ellington MJ, Reuter S, Harris SR, et al. Emergent and evolving antimicrobial resistance cassettes in community-associated fusidic acid and meticillin-resistant Staphylococcus aureus. Int J Antimicrob Agents. 2015;45(5):477–484. doi:10.1016/j.ijantimicag.2015.01.009
12. El-Mahdy TS, El-Ahmady M, Goering RV. Molecular characterization of methicillin-resistant Staphylococcus aureus isolated over a 2-year period in a Qatari hospital from multinational patients. Clin Microbiol Infect. 2014;20(2):169–173. doi:10.1111/1469-0691.12240
13. Monecke S, Skakni L, Hasan R, et al. Characterisation of MRSA strains isolated from patients in a hospital in Riyadh, Kingdom of Saudi Arabia. BMC Microbiol. 2012;12:146. doi:10.1186/1471-2180-12-146
14. Conceicao T, Tavares A, Miragaia M, Hyde K, Aires-de-Sousa M, de Lencastre H. Prevalence and clonality of methicillin-resistant Staphylococcus aureus (MRSA) in the Atlantic Azores islands: predominance of SCCmec types IV, V and VI. Eur J Clin Microbiol Infect Dis. 2010;29(5):543–550. doi:10.1007/s10096-010-0892-4
15. Dauwalder O, Lina G, Durand G, et al. Epidemiology of invasive methicillin-resistant Staphylococcus aureus clones collected in France in 2006 and 2007. J Clin Microbiol. 2008;46(10):3454–3458. doi:10.1128/JCM.01050-08
16. Sa-Leao R, Santos Sanches I, Dias D, Peres I, Barros RM, de Lencastre H. Detection of an archaic clone of Staphylococcus aureus with low-level resistance to methicillin in a pediatric hospital in Portugal and in international samples: relics of a formerly widely disseminated strain? J Clin Microbiol. 1999;37(6):1913–1920.
17. Askarian F, van Sorge NM, Sangvik M, et al. A Staphylococcus aureus TIR domain protein virulence factor blocks TLR2-mediated NF-kappaB signaling. J Innate Immun. 2014;6(4):485–498. doi:10.1159/000357618
18. Durand G, Bes M, Meugnier H, et al. Detection of new methicillin-resistant Staphylococcus aureus clones containing the toxic shock syndrome toxin 1 gene responsible for hospital- and community-acquired infections in France. J Clin Microbiol. 2006;44(3):847–853. doi:10.1128/JCM.44.3.847-853.2006
19. Leroyer C, Lehours P, Tristan A, et al. Outbreak in newborns of methicillin-resistant Staphylococcus aureus related to the sequence type 5 Geraldine clone. Am J Infect Control. 2016;44(2):e9–e11. doi:10.1016/j.ajic.2015.09.020
20. Udo EE, Boswihi SS, Dimitrov TI, Noronha B, Mathew B. T. V. Characterization of a CC1153 PVL-producing community-acquired methicillin-resistant Staphylococcus aureus from a dog bite wound. J Infect Dev Ctries. 2017;11(6):513–516. doi:10.3855/jidc.8954
21. Abou Shady HM, Bakr AE, Hashad ME, Alzohairy MA. Staphylococcus aureus nasal carriage among outpatients attending primary health care centers: a comparative study of two cities in Saudi Arabia and Egypt. Braz J Infect Dis. 2015;19(1):68–76. doi:10.1016/j.bjid.2014.09.005
22. Hansen TA, Bartels MD, Hogh SV, et al. Whole genome sequencing of Danish Staphylococcus argenteus reveals a genetically diverse collection with clear separation from Staphylococcus aureus. Front Microbiol. 2017;8:1512. doi:10.3389/fmicb.2017.01512
23. Chantratita N, Wikraiphat C, Tandhavanant S, et al. Comparison of community-onset Staphylococcus argenteus and Staphylococcus aureus sepsis in Thailand: a prospective multicentre observational study. Clin Microbiol Infect. 2016;22(5):458 e411–458 e459. doi:10.1016/j.cmi.2016.01.008
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.