Back to Journals » Infection and Drug Resistance » Volume 16
Effect of Three Different Amino Acids Plus Gentamicin Against Methicillin-Resistant Staphylococcus aureus
Authors Guo J, Pan Z, Fan L, Zhong Y, Pang R, Su Y
Received 29 March 2023
Accepted for publication 5 July 2023
Published 20 July 2023 Volume 2023:16 Pages 4741—4754
DOI https://doi.org/10.2147/IDR.S411658
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Juan Guo,1,* Zhiyu Pan,1,* Lvyuan Fan,1 Yilin Zhong,1 Rui Pang,2 Yubin Su1
1Department of Cell Biology & Institute of Biomedicine, National Engineering Research Center of Genetic Medicine, MOE Key Laboratory of Tumor Molecular Biology, Guangdong Provincial Key Laboratory of Bioengineering Medicine, College of Life Science and Technology, Jinan University, Guangzhou, 510632, People’s Republic of China; 2Guangdong Provincial Key Laboratory of Microbial Safety and Health, State Key Laboratory of Applied Microbiology Southern China, Guangdong Institute of Microbiology, Guangdong Academy of Sciences, Guangzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Yubin Su, Email [email protected]
Background: The issue of methicillin-resistant Staphylococcus aureus (MRSA) resistant to many antibiotics and causing serious infectious diseases is a growing healthcare concern.
Purpose: In recent years, exogenous administration of metabolites in combination with antibiotics can re-sensitize resistant bacteria to antibiotics; however, their effects vary, and their underlying mechanism of action remains elusive.
Methods: We assessed the bactericidal effects of the three amino acids in combination with gentamicin in vitro and in vivo. Subsequently, we explored the role of these amino acids on the metabolomics of MRSA using Liquid chromatography-tandem mass spectrometry (LC-MS/MS). Furthermore, we performed the downstream analyses using MetaboAnalyst and Interactive Pathways Explorer.
Results: Exogenous threonine showed the best bactericidal efficacy with gentamicin, followed by glycine, wherein serine had no effect. Amino acid treatments mainly up-regulated the metabolites, increased the amino acid abundance, and significantly activated metabolisms; these effects were consistent with the bactericidal efficacy of the three amino acids. Most amino acids participated in the tricarboxylic acid cycle, and threonine supplementation increased the activities of citrate synthase, isocitrate dehydrogenase and α-ketoglutarate dehydrogenase, whereas glycine increased activities of citrate synthase and α-ketoglutarate dehydrogenase, and serine did not affect the activities of any of the three key enzymes. We identified 24 biomarkers in the three groups, among which glutamic acid and cysteine showed a gradient decrease and increase, respectively. Subsequent analyses revealed that glutamic acid but not cysteine promoted the bactericidal effect of gentamicin synergistically.
Conclusion: Threonine has the best synergistic effect in reversing bacterial resistance compared to glycine and serine. We show that different amino acids combined with an antibiotic mainly affect amino acid metabolism and act via different metabolic regulatory mechanisms, which could help develop effective strategies for tackling MRSA infections.
Keywords: methicillin-resistant Staphylococcus aureus, metabolomics, gentamicin, amino acid, metabolic regulation
Graphical Abstract:

Introduction
Methicillin-resistant Staphylococcus aureus (MRSA) had been described in 1960 as an infection caused by a species of Staphylococcus. MRSA is one of the most dangerous clinical pathogens, which leads to prolonged hospital stays and increased mortality, thus posing a great threat to clinical antimicrobial therapy.1–3 MRSA is highly resistant to cefoxitin, ciprofloxacin, gentamicin, and other antibiotics.4,5 The most commonly used treatments for MRSA infection include radiation,6 antibiotic,7,8 surfactant molecular,9 and nanomaterial10 therapies; among these, antibiotic therapy is the first choice in many cases. However, antibiotic resistance remains one of the leading causes of death worldwide.11 Nearly 4.95 million deaths were estimated to be associated with antimicrobial resistance (AMR), of which 1.27 million deaths were the direct result of AMR in 2019, especially MRSA caused more than 100, 000 deaths.12 Therefore, it is essential to tackle the problems associated with MRSA resistance.
Recent studies have explored the efficiency of reprogramming metabolomics that alters the bacterial metabolic mechanism by adding exogenous metabolites to enhance the killing efficacy of antibiotics and assist with overcoming antibiotic resistance.13–18 Several studies have explored that amino acids, as important metabolites involved in metabolism, have been used to enhance the bactericidal efficiency of antibiotics. For instance, glycine, serine, and threonine have been shown to interfere with the serum-resistant bacteria metabolomes, thereby enhancing the bactericidal ability of serum against Escherichia coli.19 Similarly, glycine, serine, and threonine potentiated the kanamycin-mediated killing of Edwardsiella piscicida.20 A recent report showed that exogenous glycine restored serum sensitivity to clinical isolates of Vibrio alginolyticus and E. coli by compensating defects in glycine, serine, and threonine metabolism related to serum resistance.21
Aminoglycoside antibiotics were among the early discovered and widely used antibiotics; however, their clinical use has diminished because of their strong toxic side effects.22,23 Studies have shown that both Gram-positive such as MRSA and Enterococcus faecalis and Gram-negative bacteria such as Acinetobacter were highly resistant to aminoglycosides.24,25 To improve the bactericidal ability of traditional antibiotics and avoid the side effects caused by excessive use of antibiotics, many studies have demonstrated the potential of exogenous substances to enhance the killing of bacteria by aminoglycoside antibiotics and reduce the dosage of antibiotics. For example, n-butanol enhanced the bactericidal effect of aminoglycosides on MRSA;26 sanguinarine effectively enhanced the killing effect of aminoglycosides on a variety of bacterial pathogens;27 uracil restored the sensitivity of MRSA to aminoglycosides through metabolic programming.28 These studies showed that restoring sensitivity to existing antibiotics has received great attention. Although gentamicin alone has not been widely reported for the treatment of MRSA infection, in recent years, many studies have demonstrated that gentamicin can act synergistically with substances against clinically isolated MRSA.26,28–30 Although some studies have explored the mechanisms involved in reversing aminoglycoside resistance and improving the efficiency of antibiotic-mediated killing,17,31 knowledge of detailed mechanisms is still not comprehensive. Therefore, identifying new mechanisms to reverse bacterial drug resistance could reinvigorate the strategies for tackling multidrug-resistant pathogen.
In recent years, metabolomics can use multivariate statistics to extract meaningful information from metabolomics data.32 So, using metabolomics to crack the mystery of bacterial drug resistance has become one of the main means.33,34 The present study aimed to explore the bactericidal effects of threonine, glycine, and serine in combination with gentamicin and analyze the alterations in the metabolome of MRSA treated with either of these three amino acids with an LC-MS/MS-based untargeted metabolomics. The study will help explore the hidden mechanism underlying the differences in the efficacies of different amino acids against MRSA. In addition, it will provide insight into the metabolomic changes induced by the combination approach and help develop metabolism-targeted treatment strategies to reverse bacterial drug resistance.
Materials and Methods
Bacterial Strain and Growth Conditions
Methicillin-resistant S. aureus (MRSA; USA300_FPR3757) in this study was obtained by Dr. Hua Zhou from Zhejiang University (Hangzhou, China). The MRSA was incubated for 12 h at 37 °C with shaking device at 220rpm. Bacterial cells were harvested by centrifugation at 8000 rpm for 5 min. The cells were washed twice with 0.85% sterile saline, re-suspended in M9 minimal media containing 10 mM acetate, 1 mM magnesium sulfate, and 100 μM calcium chloride, diluted to a final density of OD600 of 1.0. The test and control groups with or without added glycine, serine, or threonine (200μM), were then incubated at 37 °C, with shaking at 220 rpm, for 6 h. Cultures were collected for metabolomics detection and enzyme activity analysis.
Minimum Inhibitory Concentration (MIC) Determination
Single colonies were picked from Luria-Bertani (LB) medium and cultured until saturation. The cells were then transferred at a ratio of 1:100 and cultured until the logarithmic growth phase. During this period, bacteria were used for MIC determination in a 96-well plate. The first column of the 96-well plate received 160 μL of medium, and each of the remaining columns 90 μL. Afterward, 20 μL of antibiotics was immediately added to the first column and mixed well, then 90 μL of the mixture was pipetted to the second column, and this operation was repeated in each column so that each antibiotic was in a state of doubling dilution. Finally, 10 μL of 100-fold-diluted bacterial solution was added to each well and mixed, and the plate was incubated for 16 h to observe colony deposition in the wells and judge the MIC of each antibiotic.
Chequerboard Method
Single colonies were inoculated into LB medium to saturation, transferred to a 1:100 dilution, and cultured to the logarithmic growth phase, at which point bacteria were taken for determination in a 96-well plate. The first column of the 96-well plate received 160 μL of medium, and each of the remaining columns 90 μL. Afterward, 20 μL of gentamicin was immediately added to the first column and mixed well, then 90 μL of the mixture was pipetted to the second column, and this operation was repeated in each column so that the antibiotics were in a state of doubling dilution. Finally, 10 μL of 100-fold-diluted bacteria solution or/and 10 μL of each of the three amino acids (200μM) were added to each well, mixed, and incubated for 16 h to observe colony deposition in the wells and judge the MIC values.
Bacterial Survival Assays
Bacterial survival assays were performed as described previously.13 In brief, the centrifuged bacterial precipitate was washed three times with 0.85% sterilized saline, resuspended in M9 minimal medium, and diluted to OD600 = 0.2. Three biological replicates were performed according to the above steps, and 5 mL of OD600 = 0.2 bacterial solution was pipetted into a test tube for each biological replicate. The experimental groups were as follows: blank, amino acid alone, antibiotic alone, and amino acid and antibiotic. The amino acids (threonine, glycine, and serine were all applied according to the same concentration gradient: 0, 100, 150, 200, 250, and 300 μM; cysteine and glutamic acid were applied according to the same concentration gradient: 0, 5, 10, 20, 40, and 80 μM) and antibiotics (gentamicin concentration of 200 μg/mL, methicillin concentrations of 0, 0.0625, 0.125, 0.25, 0.5, and 1 μg/mL) were added to the 5 mL system. The tubes were then incubated on a shaker (6 h, 37 °C, 220 rpm). Serial dilutions of 100 μL cultures were used to determine the number of bacteria. Aliquots (10 μL) of each dilution were inoculated onto LB agar plates and incubated at 37 °C for 12 h. Only plates with 20–200 colonies were counted, and colony-forming units per milliliter (CFU/mL) were calculated.
Quantitative Analysis by Liquid Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
Sample Preparation
Sample preparation was performed as previously described.13 In brief, 10 mL of cells with an OD600 of 1.0 were incubated in M9 minimal medium for 6 h, with or without threonine, glycine or serine (200μM). Equivalent amounts of the cells were collected by centrifugation and quenched using liquid nitrogen. The bacterial precipitate was resuspended in a cold extraction solution (2:2:1 acetonitrile:methanol:water, containing 2 µg/mL of L-2-chlorophenylalanine as an internal standard) and sonicated for 15 min on ice. The supernatant was collected after centrifugation and incubated for 1h at −20 °C, then centrifuged again for 15 min at 12,000 × g at 4 °C. The cell suspension was transferred to a new centrifuge tube and concentrated under vacuum, and then 150 μL of solvent (acetonitrile:water = 1:1) was added for re-dissolution. The samples were then centrifuged for 15 min at 12,000 × g and 4 °C), and the supernatant was used for subsequent analysis by LC-MS/MS.
LC-MS/MS Analysis
Briefly, LC-MS/MS analysis was performed using an UltiMate 3000 Rapid Separation Liquid System (Thermo Fisher Scientific, Waltham, MA, USA). Extracted metabolite samples were loaded into a Waters ACQUITY UPLC HSS T3 column (2.1 mm × 100 mm, 1.8 μm particle size; Waters Co., Milford, MA, USA) with mobile phase A containing 0.1% formic acid with 5 mmol/L ammonium acetate and linear gradient elution of mobile phase B (acetonitrile). The gradient elution conditions of mobile phase chromatography were as follows: 1% (v/v) B over 0–1.0 min; 1–99% (v/v) B over 1.0–8.0 min; 99% (v/v) B over 8.0–10.0 min; 99–1% (v/v) B over 10.0–10.1 min; 1% (v/v) B over 10.1–12 min. The mobile phase flow rate was set at 500 μL/min and analysed at a column temperature of 35 °C. Xcalibur 4.0.27 (Thermo Fisher Scientific) software was used for the acquisition of primary and secondary mass spectrometry data. Data were collected using previously described settings.35
Metabolome Data Analysis
Data analysis was performed using IBM SPSS Statistics 22 software (IBM, Armonk, NY, USA), and values of p < 0.05 were regarded as significant differences. Hierarchical clustering was performed using the R programming language with the g-plots package. Z-scores were used to determine the normalised peak areas of each differential metabolite. Multivariate statistical approaches, including principal component analysis (PCA) and S-plots, were performed using the software program Simca–P + 12.0.1 (Umetrics AB, Umea, Sweden). MetaboAnalyst 5.0 was used to perform enrichment of important metabolic pathways.36
Enzyme Assay
Isocitrate dehydrogenase (ICDH) and α-ketoglutarate dehydrogenase (OGDH) were detected as previously described.37 The carefully prepared bacterial precipitate was resuspended in phosphate-buffered saline (PBS) and sonicated (200 W total power with 60% output, 2 s pulse, and 3 s pause) over ice for 40 min. The reagents required for the ICDH enzyme activity assay included 0.15 mM 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT), 0.5 mM phenazine methosulfate (PMS), 2.5 mM MgCl2, 1M Tris-HCl, and 70 mM sodium isocitrate, with water added to produce a final volume of 200 μL. The OGDH assay used 0.15 mM MTT, 0.5 mM PMS, 0.2 mM thiamine PPi (TPP), 2.5 mM MgCl2, 50 mM potassium phosphate buffer, and 80 mM α-ketoglutaric acid, with water added to produce a final volume of 200 μL. The reaction mixtures were incubated in the dark at 37 °C for 30 min. Finally, colorimetric assay readings were measured at 566 nm. The activity of citrate synthase (CS) was assayed using a commercial chemical assay kit (Keming Biotechnology Co., Ltd., Suzhou, China). The initial absorbance at 412 nm was recorded after adding the sample to the working solution, and the absorbance at 412 nm was recorded again after 2 min in a water bath at 25 °C.
Mouse Infection Assay
The experiment was conducted as described previously.28 BALB/c mice (8 weeks old, weighing 20 ± 2 g) were purchased from Zhuhai BesTest Bio-Tech Co., Ltd. (SCXK (G) 2020–0051; Zhuhai, China). The mice were reared in cages, fed sterile water and dry pellet diets, and acclimatised for one week. Bacteria (USA300) grown in LB medium to an OD600 of 1.0 were harvested by centrifugation and washed twice in PBS. Subsequently, the bacterial precipitate was resuspended in PBS, and the concentration was adjusted to 1 × 109 CFU/mL. Each mouse was injected with 100 μL cell suspension containing 1 × 109 CFU/mL of bacteria through the tail vein; control mice were injected with sterile PBS instead. After 6 h, the mice that received bacteria injections were randomly and equally divided into eight groups (n = 8 per group) and injected with PBS, glycine (500 mg kg −1), serine (500 mg kg −1), threonine (500 mg kg −1), gentamicin (5 mg kg −1), gentamicin + glycine, gentamicin + serine, or gentamicin + threonine once a day. The control group was a bacterial non-injection group (sterile PBS injection only). The survival of all mice was observed for seven days, and observations were made and recorded daily. Animal experiment was approved by the Institutional Animal Care and Use Committee at the Guangdong Institute of Microbiology (Animal Welfare Assurance Number GT-IACUC201907031).
Results
Comparison of Bacteriostatic and Bactericidal Effects of Gentamicin on MRSA
To determine the sensitivity of MRSA to antibiotics, we used the MIC method to determine the inhibitory effects of nine common clinical antibiotics. The results showed that the MIC values of these antibiotics toward MRSA were between 0.078125 and 5 μg/mL, and the MIC value of gentamicin was 0.3125 μg/mL (Figure S1A). This indicated that MRSA is extremely sensitive to gentamicin and other antibiotics, which means that low antibiotic concentrations could inhibit its growth; however, to determine whether low concentrations of antibiotics had a bactericidal effect, we further determined the bactericidal concentration of gentamicin on MRSA. The results showed a significant bactericidal effect when gentamicin concentration reached 800 μg/mL (2560 MIC), that is, the MRSA survival rate dropped to approximately 30% (Figure S1B). These results suggest that although the MIC value of gentamicin for MRSA is very low, a high concentration of gentamicin should be used to kill MRSA. Therefore, it is necessary to explore whether other substances have synergistic bactericidal effects when applied alongside gentamicin.
Efficacy of Threonine, Glycine, and Serine with Gentamicin in Killing MRSA
To determine the synergistic effect of amino acids on gentamicin-induced killing of MRSA, we investigated the bactericidal efficacy of either threonine, glycine, or serine in combination with gentamicin in vitro. Results showed that cell survival decreased with increasing concentrations of threonine (Figure 1A) and glycine (Figure 1B) in the presence of a fixed gentamicin concentration. In comparison, we found that for a given gentamicin concentration, the bactericidal effect of threonine was approximately 100-fold stronger than that of glycine. However, serine did not show any synergistic bactericidal effects (Figure 1C). We further determined whether there were differences in the synergistic bacteriostatic effects of the amino acids combined with gentamicin on MRSA. We used the checkerboard method and found that both threonine and glycine reduced the MIC values twofold; however, serine had no effect on the MIC value of gentamicin against MRSA (Figure S1C). Next, we explored the effects of threonine, glycine, and serine in a mouse model of MRSA infection. While the mice in the bacterial non-injection group had a 100% survival rate, the mortality rate of the mice in the bacterial injection group (bacteria suspended in PBS) reached 70% (Table S1), indicating that MRSA infection led to the death of the mice. In contrast to the bacterial injection group, the survival rates of the mice in the treatment groups increased by 50%, 40%, and 20% after the injection of gentamicin plus threonine, glycine, and serine, respectively. However, only the combination of gentamicin and threonine resulted in a significant difference (Figure 1D and Table S1). Taken together, both in vitro and in vivo results show that the synergistic effect of the three amino acids with gentamicin on MRSA varied in the following order: threonine > glycine > serine.
 |
Figure 1 Effects of threonine, glycine and serine synergistically with gentamicin on the survival rate of MRSA and the survival rate of mice. (A–C) gentamicin (200μg/mL) combined with different concentrations of threonine (Thr), glycine (Gly) and serine (Ser) killing MRSA. (D) Percent survival of mice infected by S. aureus, USA300 during 7 days. All data are displayed as mean ± SEM. ns, no significant difference, *p < 0.05, **p < 0.01, ***p < 0.001, determined by Student’s t test. |
Threonine, Glycine, and Serine Impact Metabolic Profile
To explore the effect of threonine, glycine, and serine on the metabolic regulation of MRSA, we treated MRSA with the three amino acids at the same concentration. We analyzed their effects on metabolic alterations using LC-MS/MS-based metabolomics approach. Sixty-five differential metabolites were identified in bacteria treated with threonine, glycine, and serine (Figure 2A). Z-scores of the intensity area of differential metabolites varied from −50 to 150. By contrast with the control group, abundance of 40 metabolites were significantly increased, while 25 metabolites were significantly decreased in the threonine group (Figure 2B); 35 metabolites were significantly increased, and 30 metabolites were significantly decreased in the glycine group (Figure 2C); only 29 metabolites were significantly increased, whereas 36 metabolites were significantly decreased in the serine group (Figure 2D). These findings suggest that three amino acids have different effects on metabolic profiles of MRSA, which may be related to the varying degree of synergistic effects with gentamicin in eliminating MRSA.
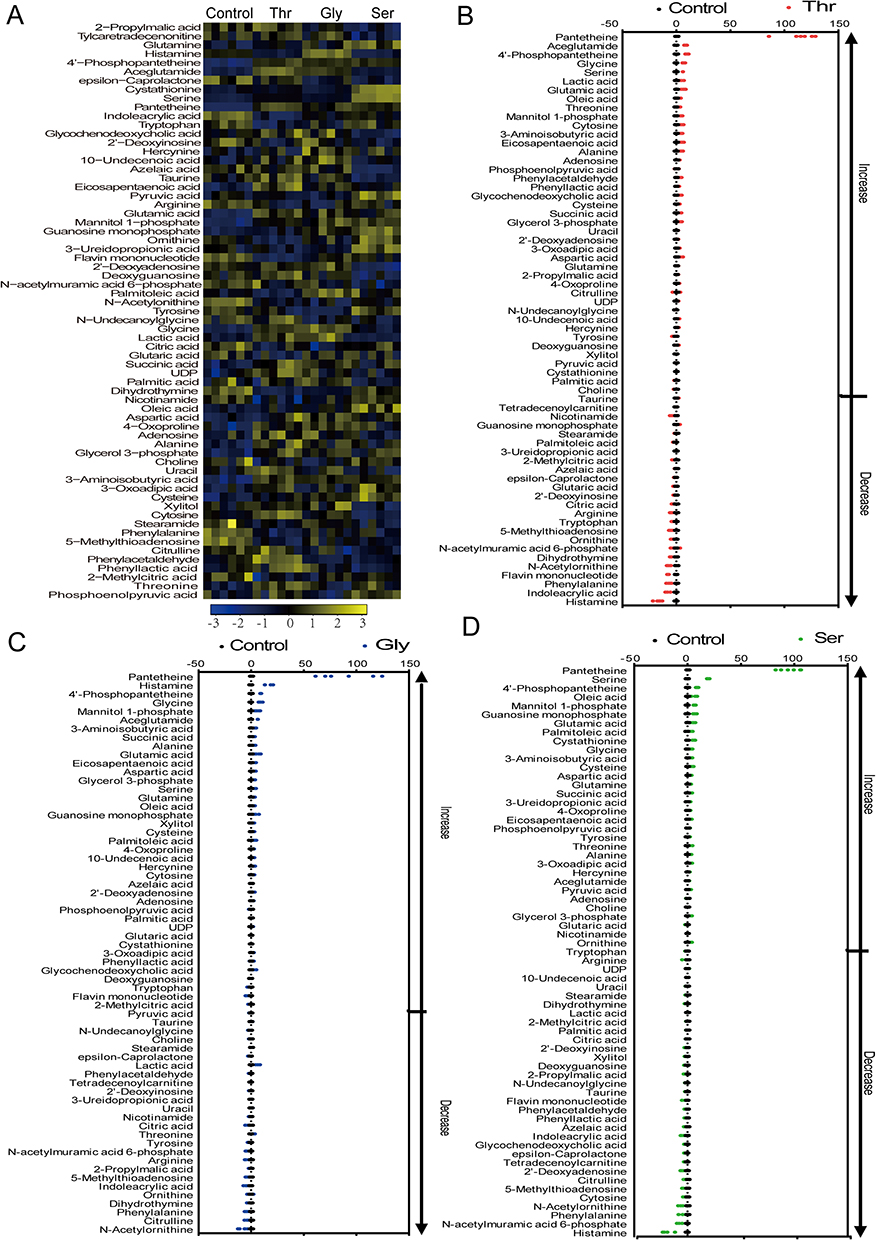 |
Figure 2 Differential metabolomics of USA300 in response to threonine, glycine and serine. (A) The heatmap showing the relative abundance of metabolites in the control group, Thr group, Gly group, and Ser group. Differential metabolites were detected through the Mann–Whitney U-Test by SPSS analysis software, with the asymptotically significant p < 0.05. Scales are shown at the bottom, where blue to yellow indicate low to high abundance. (B–D) Control-based differential metabolite Z-score plots for threonine, glycine, and serine groups, respectively. |
Analysis of Differential Metabolite Pathways After Treatment with Threonine, Glycine, and Serine
Pathway enrichment analysis using the MetaboAnalyst platform can identify possible bio-perturbed metabolic pathways, which is important for analyzing metabolic states affected by different substances. Pathway enrichment analysis using the MetaboAnalyst identified 14 significantly involved metabolic pathways (p < 0.05; Figure 3A). In contrast to the control group, among a lot of differential metabolites, the relative abundance of amino acids in enriched metabolic pathways mostly increased in the following order: threonine group > glycine group > serine group (Figure 3B). Further analysis showed that these amino acids (ie, aspartate, glutamate, glutamine, and alanine) were closely associated with the tricarboxylic acid (TCA) cycle. Although the TCA cycle was not identified as a significantly enriched pathway, the increased abundance of these amino acids may affect the TCA cycle. Furthermore, the activities of the three key enzymes in the TCA cycle, including citrate synthase (CS), isocitrate dehydrogenase (ICDH) and α-ketoglutarate dehydrogenase (OGDH), were elevated in the threonine group. By contrast, only CS and OGDH in the glycine group were increased, wherein no significant effect on the three enzyme activities was observed in the serine group (Figures 3C–E). In addition, it should be noted that citrulline was elevated in the threonine group, whereas in the glycine and serine groups were decreased (Figure 3B). These results suggest that the supply of exogenous three amino acids mainly affects amino acid metabolism, wherein only threonine can enhance the TCA cycle.
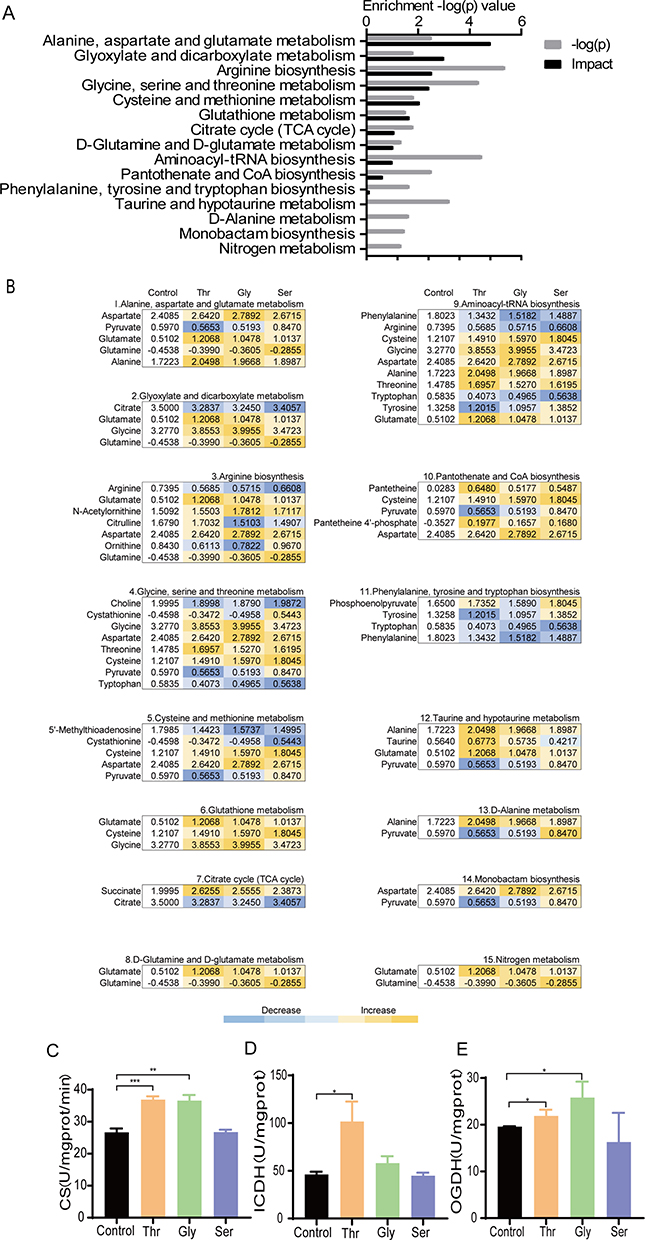 |
Figure 3 Threonine, glycine and serine groups have some differences in the relative abundance of metabolites and the TCA cycle was more active in the threonine and glycine groups than in the serine group. (A) Enriched metabolic pathways shared by Thr group, Gly group, and Ser group. Select and analyze differentially abundant metabolites to enrich pathways in MetaboAnalyst 5.0 (https://pathways.embl.de). The fifteen pathways that had significant difference (p < 0.05) were enriched and sorted by their weights (impact). (B) Differential metabolites in enriched pathways. Scales are shown in the lower right, where blue indicates decreased relative abundance of differential metabolites and yellow indicates increased relative abundance of differential metabolites. Lighter colors represent less increase or decrease, darker colors represent more increase or decrease. (C–E) Activity of citrate synthase (CS), isocitrate dehydrogenase (ICDH) and α-ketoglutarate dehydrogenase (OGDH) were detected with 200 μM the Thr, Gly, and Ser, respectively. All data are displayed as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001, determined by Student’s t test. |
Effects of Threonine, Glycine, and Serine on the Global Metabolic Network of MRSA
Visualization of the pathway diagram using the Interactive Pathways Explorer (ipath) 3.0 revealed shared as well as unique metabolic networks in the three groups treated with threonine, glycine, and serine (Figures 4A–C). Most metabolic pathways, containing lipid, amino acid, energy metabolism, and other amino acid metabolisms, were up-regulated in the three amino acid groups. On the contrary, the carbohydrate metabolism pathway was down-regulated in the threonine and glycine groups but was up-regulated in the serine group. These results demonstrate that the global metabolic regulation of MRSA is similar between the threonine and glycine groups but different in the serine group.
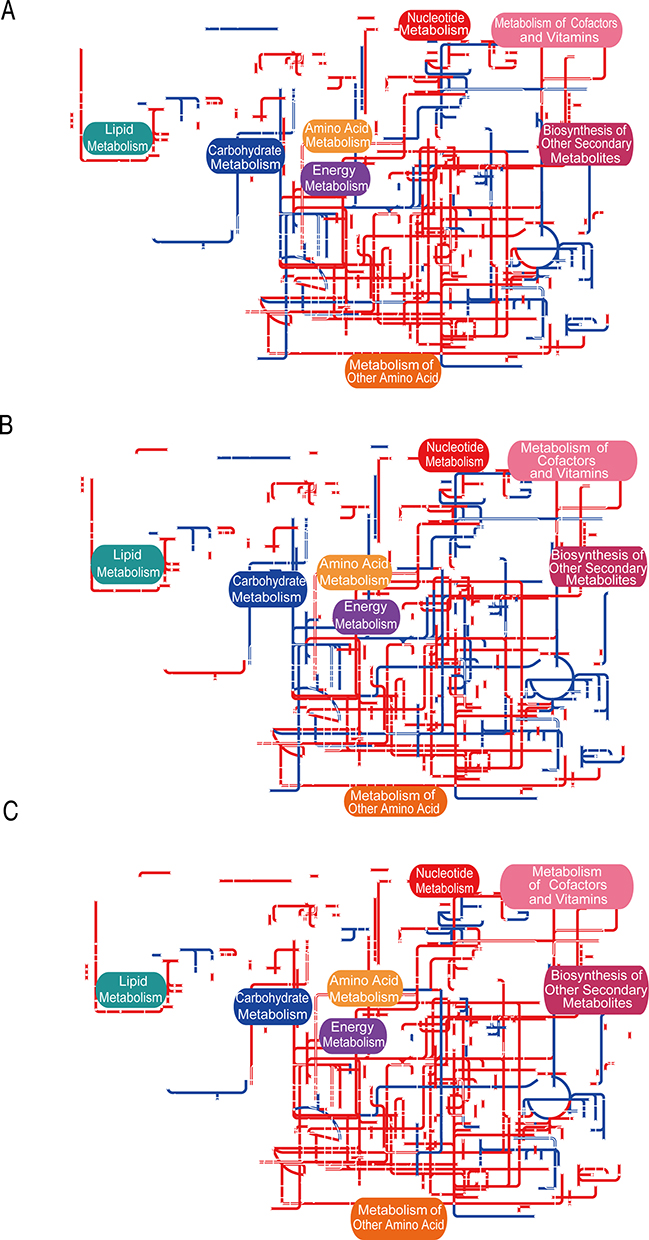 |
Figure 4 IPath integrated analysis of changes in the metabolic pathways. (A–C) Global metabolic network of the Thr, Gly, and Ser groups, respectively. Red lines represent up-regulation of metabolic pathways, blue lines represent down-regulation of metabolic pathways. |
Crucial Biomarkers Responsible for the Susceptibility of MRSA to Gentamicin
Metabolites distinguishing the three amino acid groups were performed using orthogonal partial least squares-discriminant analysis (OPLS-DA) model. As shown in Figure 5A, t[1] separated the control group from the serine and other two groups one by one, and t[2] separated the control and serine groups from the other two groups. The threonine and glycine groups were relatively close, indicating relatively small metabolome differences between the threonine and glycine groups. Next, we used S-plots to deliver both the covariance and correlation between the endogenous metabolites and the modeled class designation names to identify crucial biomarkers. The cut-off values were set for the covariance of p ≥ |0.05| and for the correlation of p (corr) ≥ |0.5|.38 We identified 24 biomarkers in the three groups (Figure 5B), and nine biomarkers showed gradient changes in those groups (Figure 5C). In contrast with the control group, the content of cysteine in the serine group was the highest, followed by the glycine group, and the lowest in the threonine group. The glutamic acid content was the highest in the threonine group, followed by those in the glycine and serine groups. Together, these findings indicate that cysteine and glutamic acid production might be related to antibiotic-mediated killing.
 |
Figure 5 Identification and verification of crucial metabolites. (A) Principal component analysis of control group, Thr group, Gly group, and Ser group. Each point represents a technical replicate analysis of the sample in the graph. (B) S-plot of OPLS-DA. Triangle represents individual metabolite. Red indicates the potential biomarkers, which is greater than or equal to 0.05 and 0.5 for absolute value of covariance p [1] and correlation p (corr) [1], respectively. (C) Abundance of biomarkers in control, Thr, Gly, and Ser groups. (D) Different concentrations of glutamic acid synergistically sterilize with 200μg/mL gentamicin. (E) Different concentrations of cysteine synergistically sterilize with 200μg/mL gentamicin. All data are displayed as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001. All date determined by Student’s t test. |
Next, to explore whether glutamic acid and cysteine affect the synergistic bactericidal effect, we assessed the effects of different concentrations of glutamic acid and cysteine in addition to gentamicin against MRSA. Our result found that the synergistic bactericidal effect of glutamic acid plus gentamicin enhanced with the increased concentration of glutamic acid (Figure 5D). In contrast, cysteine and gentamicin had no synergistic bactericidal effect (Figure 5E). These results suggest that the crucial biomarkers of glutamic acid and cysteine affect the synergistic bactericidal action caused by the three amino acids of different metabolic reactions.
Discussion
Several studies have demonstrated that amino acids combined with aminoglycoside antibiotics can synergistically reverse bacterial resistance.39,40 Consistently, in this study, we showed that threonine had the best killing efficacy with gentamicin, wherein the efficacy of glycine was moderate, and serine had no effect. But, interestingly, threonine or glycine in combination with methicillin could not re-sensitize resistant bacteria (Figure S2). In animal experiments, we found that the injection of threonine in combination with gentamicin could significantly increase survival rate of mice, but glycine or serine plus gentamicin had no effect. Notably, doses of gentamicin (5 mg/kg/day) combined with threonine improved the survival rates of the mice, even though this concentration was high (7 mg/kg/day is recommended to achieve the maximum concentration) and may produce certain toxic effects. Therefore, it is necessary to explore more suitable combinations of antibiotic and substance concentrations. Moreover, it would have been valuable to look for the presence of bacteria after the mice died at the end of the experiment. Further studies are needed to explore which tissues and organs harbour bacteria. We performed metabolome analysis, which revealed different numbers of differentially up-regulated metabolites among the three amino acid-treated groups; threonine up-regulated the highest, wherein serine up-regulated the least. Therefore, we speculate that threonine, the substance with the best synergistic bactericidal effect, may increase bacterial metabolic flux and accelerate bacterial metabolism, thereby making it easier to reverse bacterial resistance.
Consistent with a previous study,20 the metabolic pathway analysis in this study showed that the addition of exogenous three amino acids mainly affected the amino acid metabolic pathways. Our results also demonstrated that most up-regulated amino acids are involved in the TCA cycle. Consistent with the metabolomics results, the accumulation of amino acids associated with the TCA cycle was the highest levels of the threonine group, followed by the glycine and serine groups. These findings suggest different TCA cycle pathway activities in the three groups. TCA cycle is an important metabolic pathway of bacteria and has been shown to be associated with the reversal of drug resistance of bacteria. For example, alanine restored biotic sensitivity in Xanthomonas oryzae by promoting the TCA cycle;41 baicalin and ethylenediaminetetraacetic acid (EDTA) increased the antimicrobial effect of colistin against colistin-resistant Salmonella by accelerating the TCA cycle;42 zeolitic imidazolate framework-8 accelerated MRSA death by disrupting the TCA cycle.43 Therefore, to gain further insights, we detected the activities of three key enzymes belonging to the TCA cycle. In contrast with the control group, the activities of three key enzymes (CS, ICDH and OGDH) in the threonine group and two (CS and OGDH) in the glycine group were markedly increased. In contrast, the activities of three key enzymes did not change in the serine group. These results provided evidence that the addition of threonine or glycine activated the TCA cycle and enhanced the bactericidal action of aminoglycoside antibiotics synergistically. It had been reported that nitric oxide enhanced the bactericidal activity of antibiotics.44–46 Citrulline, act as an antioxidant and participates in the process of arginine-mediated nitric oxide synthesis. Therefore, we speculated that the best effect of threonine in reversing drug resistance compared with those of glycine and serine could be associated with citrulline generation in the threonine group.
Furthermore, iPath analysis revealed that the activation of metabolism was dominant significantly, and carbohydrate metabolism was down-regulated in the threonine and glycine groups but up-regulated in the serine group. This could be explained by the fact that the key pathways, such as glycolysis or other metabolic pathways connecting the carbohydrate, fat, and amino acid metabolisms, diverge the metabolites towards carbohydrate metabolism in the serine group. This coincides with the result that only threonine and glycine groups promote the TCA cycle, whereas serine does not. However, this still needs to be further explored. Finally, we identified nine crucial biomarkers that showed gradient changes across the three groups and found that serine was more readily converted to cysteine, wherein glycine and threonine were converted to glutamic acid, consistent with previous findings.21 Glutamic acid reverses the drug resistance of Gram-negative and Gram-positive bacteria.47,48 Our data, consistent with these studies, showed that cysteine did not help reverse bacterial resistance, while glutamic acid could. These findings, at least partially, explain the lack of synergistic bactericidal effect of serine compared to those of threonine and glycine.
Conclusion
This study compared and analyzed the metabolome characteristics of MRSA strains stimulated by threonine, glycine, and serine to explore the hidden reversing drug resistance mechanism. Compared with serine, threonine and glycine had a synergistic bactericidal effect, among which threonine performed better than glycine. Based on the results of this study, we hypothesized two mechanisms underlying the synergistic effect of threonine and glycine in promoting the bactericidal effect of gentamicin. First, the TCA cycle pathways of these two groups are more active, and the increase in metabolic flux promotes bacterial metabolism, thereby reversing bacterial drug resistance. Second, adding threonine or glycine promotes more conversion into glutamic acid, whereas serine promotes conversion into cysteine. Furthermore, our results indicated that the better synergistic bactericidal effect of threonine than that of glycine could be because threonine promoted citrulline production and increased the amount of glutamic acid accumulated in bacteria. Overall, our study reveals that different amino acids have different mechanisms of metabolic regulation for MRSA and suggests that threonine has the best synergistic effect in reversing bacterial resistance compared to glycine and serine.
Ethical Approval
Animal research was approved by Guangdong Institute of Microbiology of Institutional Animal Care and Use Committee (Animal Welfare Assurance No. is GT-IACUC201907031). All procedures related to animals were performed in accordance with the ethical and welfare guidelines of Guangdong Institute of Microbiology.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was sponsored by National Natural Science Foundation of China (42076095), Project of Guangzhou Science and Technology Planning (202102021206) and Guangdong Province Natural Science Foundation (2022A1515010408).
Disclosure
The authors declare they have no competing interests.
References
1. Mickymaray S, Alturaiki W, Al-Aboody MS, et al. Anti-bacterial efficacy of bacteriocin produced by marine Bacillus subtilis against clinically important extended spectrum beta-lactamase strains and methicillin-resistant Staphylococcus aureus. Int J Med Res Health Sci. 2018;7(2):75–83.
2. Tasneem U, Mehmood K, Majid M, Ullah SR, Andleeb S. Methicillin resistant Staphylococcus aureus: a brief review of virulence and resistance. J Pak Med Assoc. 2022;72(3):509–515. doi:10.47391/JPMA.0504
3. Wang X, Qiu H, Yang N, et al. Fascaplysin derivatives binding to DNA via unique cationic five-ring coplanar backbone showed potent antimicrobial/antibiofilm activity against MRSA in vitro and in vivo. Eur J Med Chem. 2022;230:114099. doi:10.1016/j.ejmech.2021.114099
4. mLynarczyk-Bonikowska B, Kowalewski C, Krolak-Ulinska A, Marusza W. Molecular mechanisms of drug resistance in Staphylococcus aureus. Int J Mol Sci. 2022;23(15):8088. doi:10.3390/ijms23158088
5. Moazen J, Riyahi Zaniani F, Hallaj Asghar B. Characterization of virulence genes and antibiotic resistance of methicillin-resistant Staphylococcus aureus (MRSA) and methicillin-susceptible Staphylococcus aureus (MSSA) isolates in intensive care unit (ICU) and non-ICU wards. Trends Med Sci. 2022;2(2):
6. Dong PT, Mohammad H, Hui J, et al. Photolysis of staphyloxanthin in methicillin-resistant Staphylococcus aureus potentiates killing by reactive oxygen species. Adv Sci. 2019;6(11):1900030. doi:10.1002/advs.201900030
7. Tsai CY, Lee CH, Chien CC, Chen IL. Impact of teicoplanin maintenance dose and MIC values on the clinical outcomes of patients treated for methicillin-resistant Staphylococcus aureus bacteremia. Infect Drug Resist. 2018;11:1205–1217. doi:10.2147/IDR.S171236
8. Wilcox MH, Tack KJ, Bouza E, et al. Complicated skin and skin-structure infections and catheter-related bloodstream infections: noninferiority of linezolid in a Phase 3 study. Clin Infect Dis. 2009;48(2):203–212. doi:10.1086/595686
9. Tancer RJ, Baynes K, Wiedman GR. Synergy among humimycins against methicillin‐resistant Staphylococcus aureus. Peptide Science. 2021;113:e24197. doi:10.1002/pep2.24197
10. Hassan D, Omolo CA, Fasiku VO, et al. Formulation of pH-responsive quatsomes from quaternary bicephalic surfactants and cholesterol for enhanced delivery of vancomycin against methicillin resistant Staphylococcus aureus. Pharmaceutics. 2020;12(11):1093. doi:10.3390/pharmaceutics12111093
11. World Health Organization. Ten threats to global heath in 2019. Available from: https://www.who.int/news-room/spotlight/ten-threats-to-global-health-in-2019.
12. Antimicrobial Resistance C. Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet. 2022;399(10325):629–655. doi:10.1016/S0140-6736(21)02724-0
13. Peng B, Su YB, Li H, et al. Exogenous alanine and/or glucose plus kanamycin kills antibiotic-resistant bacteria. Cell Metab. 2015;21(2):249–262. doi:10.1016/j.cmet.2015.01.008
14. Su YB, Peng B, Han Y, Li H, Peng XX. Fructose restores susceptibility of multidrug-resistant Edwardsiella tarda to kanamycin. J Proteome Res. 2015;14(3):1612–1620. doi:10.1021/pr501285f
15. Su YB, Peng B, Li H, et al. Pyruvate cycle increases aminoglycoside efficacy and provides respiratory energy in bacteria. Proc Natl Acad Sci U S A. 2018;115(7):E1578–E1587. doi:10.1073/pnas.1714645115
16. Tang XK, Su YB, Ye HQ, et al. Glucose-potentiated amikacin killing of cefoperazone/sulbactam resistant Pseudomonas aeruginosa. Front Microbiol. 2021;12:800442. doi:10.3389/fmicb.2021.800442
17. Guan Y, Shen P, Lin M, Ye X. Exogenous alanine reverses the bacterial resistance to Zhongshengmycin with the promotion of the P cycle in Xanthomonas oryzae. Antibiotics. 2022;11(2):245. doi:10.3390/antibiotics11020245
18. Zhou Y, Yong Y, Zhu C, Yang H, Fang B. Exogenous D-ribose promotes gentamicin treatment of several drug-resistant Salmonella. Front Microbiol. 2022;13:1053330. doi:10.3389/fmicb.2022.1053330
19. Cheng ZX, Guo C, Chen ZG, et al. Glycine, serine and threonine metabolism confounds efficacy of complement-mediated killing. Nat Commun. 2019;10(1):3325. doi:10.1038/s41467-019-11129-5
20. Ye JZ, Lin XM, Cheng ZX, et al. Identification and efficacy of glycine, serine and threonine metabolism in potentiating kanamycin-mediated killing of Edwardsiella piscicida. J Proteomics. 2018;183:34–44. doi:10.1016/j.jprot.2018.05.006
21. Kou TS, Wu JH, Chen XW, Chen ZG, Zheng J, Peng B. Exogenous glycine promotes oxidation of glutathione and restores sensitivity of bacterial pathogens to serum-induced cell death. Redox Biol. 2022;58:102512. doi:10.1016/j.redox.2022.102512
22. Kim J, Hemachandran S, Cheng AG, Ricci AJ. Identifying targets to prevent aminoglycoside ototoxicity. Mol Cell Neurosci. 2022;120:103722. doi:10.1016/j.mcn.2022.103722
23. McDermott JH, Wolf J, Hoshitsuki K, et al. Clinical pharmacogenetics implementation consortium guideline for the use of aminoglycosides based on MT-RNR1 genotype. Clin Pharmacol Ther. 2022;111(2):366–372. doi:10.1002/cpt.2309
24. Zhang Y, Zhang N, Wang M, et al. The prevalence and distribution of aminoglycoside resistance genes. Biosafety Health. 2023;5(1):14–20. doi:10.1016/j.bsheal.2023.01.001
25. Wang N, Luo J, Deng F, Huang Y, Zhou H. Antibiotic combination therapy: a strategy to overcome bacterial resistance to aminoglycoside antibiotics. Front Pharmacol. 2022;13:839808. doi:10.3389/fphar.2022.839808
26. Lv B, Bian M, Huang X, et al. n-Butanol potentiates subinhibitory aminoglycosides against bacterial persisters and multidrug-resistant MRSA by rapidly enhancing antibiotic uptake. ACS Infect Dis. 2022;8(2):373–386. doi:10.1021/acsinfecdis.1c00559
27. Lu C, Zhang N, Kou S, et al. Sanguinarine synergistically potentiates aminoglycoside-mediated bacterial killing. Microb Biotechnol. 2022;15(7):2055–2070. doi:10.1111/1751-7915.14017
28. Fan L, Pan Z, Liao X, et al. Uracil restores susceptibility of methicillin-resistant Staphylococcus aureus to aminoglycosides through metabolic reprogramming. Front Pharmacol. 2023;14:1133685. doi:10.3389/fphar.2023.1133685
29. Vázquez NM, Fiorilli G, Guido PAC, et al. Carnosic acid acts synergistically with gentamicin in killing methicillin-resistant Staphylococcus aureus clinical isolates. Phytomedicine. 2016;23(12):1337–1343. doi:10.1016/j.phymed.2016.07.010
30. Sreepian A, Popruk S, Nutalai D, et al. Antibacterial activities and synergistic interaction of citrus essential oils and limonene with gentamicin against clinically isolated methicillin-resistant Staphylococcus aureus. Sci World J. 2022;2022:8418287.
31. Su YB, Kuang SF, Peng XX, Li H. The depressed P cycle contributes to the acquisition of ampicillin resistance in Edwardsiella piscicida. J Proteomics. 2020;212:103562. doi:10.1016/j.jprot.2019.103562
32. Yang XL, Wang P, Ye H, et al. Untargeted serum metabolomics reveals potential biomarkers and metabolic pathways associated with esophageal cancer. Front Oncol. 2022;12:938234. doi:10.3389/fonc.2022.938234
33. Chen X, Kuang S, He Y, et al. The differential metabolic response of oral squamous cell carcinoma cells and normal oral epithelial cells to cisplatin exposure. Metabolites. 2022;12(5):389. doi:10.3390/metabo12050389
34. Kok M, Maton L, van der Peet M, Hankemeier T, van Hasselt JGC. Unraveling antimicrobial resistance using metabolomics. Drug Discov Today. 2022;27(6):1774–1783. doi:10.1016/j.drudis.2022.03.015
35. Cheng ZX, Ma YM, Li H, Peng XX. N-acetylglucosamine enhances survival ability of tilapias infected by Streptococcus iniae. Fish Shellfish Immunol. 2014;40(2):524–530. doi:10.1016/j.fsi.2014.08.008
36. Smith CA, Want EJ, O’Maille G, Abagyan R, Siuzdak G. XCMS: processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal Chem. 2006;78(3):779–787. doi:10.1021/ac051437y
37. Chen YT, Yang KX, Dai ZY, et al. Repressed central carbon metabolism and its effect on related metabolic pathways in cefoperazone/sulbactam-resistant Pseudomonas aeruginosa. Front Microbiol. 2022;13:11. doi:10.3389/fmicb.2022.847634
38. Ma TC, Zhao HA, Liu CY, et al. Discrimination of natural mature acacia honey based on multi-physicochemical parameters combined with chemometric analysis. Molecules. 2019;24(14):2674. doi:10.3390/molecules24142674
39. Jiang M, Kuang SF, Lai SS, et al. Na(+)-NQR confers aminoglycoside resistance via the regulation of l-alanine metabolism. mBio. 2020;11(6):e02086–20. doi:10.1128/mBio.02086-20
40. Yong Y, Zhou Y, Liu K, Liu G, Wu L, Fang B. Exogenous citrulline and glutamine contribute to reverse the resistance of Salmonella to apramycin. Front Microbiol. 2021;12:759170. doi:10.3389/fmicb.2021.759170
41. Zou Z, Lin M, Shen P, Guan Y. Alanine-dependent TCA cycle promotion restores the Zhongshengmycin-susceptibility in Xanthomonas oryzae. Int J Mol Sci. 2023;24(3):3004. doi:10.3390/ijms24033004
42. Cui XD, Zhang JK, Sun YW, et al. Synergistic antibacterial activity of baicalin and EDTA in combination with colistin against colistin-resistant Salmonella. Poult Sci. 2023;102(2):102346. doi:10.1016/j.psj.2022.102346
43. Liu B, Yang Y, Wu H, et al. Zeolitic imidazolate framework-8 triggers the inhibition of arginine biosynthesis to combat methicillin-resistant Staphylococcus Aureus. Small. 2023:e2205682. doi:10.1002/smll.202205682
44. Nguyen TK, Selvanayagam R, et al. Co-delivery of nitric oxide and antibiotic using polymeric nanoparticles. Chem Sci. 2016;7(2):1016–1027. doi:10.1039/c5sc02769a
45. Howlin RP, Cathie K, Hall-Stoodley L, et al. Low-dose nitric oxide as targeted anti-biofilm adjunctive therapy to treat chronic Pseudomonas aeruginosa infection in cystic fibrosis. Mol Ther. 2017;25(9):2104–2116. doi:10.1016/j.ymthe.2017.06.021
46. Rouillard KR, Novak OP, Pistiolis AM, et al. Exogenous nitric oxide improves antibiotic susceptibility in resistant bacteria. ACS Infect Dis. 2021;7(1):23–33. doi:10.1021/acsinfecdis.0c00337
47. Zhang S, Wang J, Jiang M, et al. Reduced redox-dependent mechanism and glucose-mediated reversal in gentamicin-resistant Vibrio alginolyticus. Environ Microbiol. 2019;21(12):4724–4739. doi:10.1111/1462-2920.14811
48. Warraich AA, Mohammed AR, Perrie Y, Hussain M, Gibson H, Rahman A. Evaluation of anti-biofilm activity of acidic amino acids and synergy with ciprofloxacin on Staphylococcus aureus biofilms. Sci Rep. 2020;10(1):9021. doi:10.1038/s41598-020-66082-x
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
The Diagnostic Value of Bile Acids and Amino Acids in Differentiating Acute Coronary Syndromes
Yu Q, Zhao F, Wang S, Jia X, Shen S, Zhao X, Li Y, Song J, Sun M, Liu X, Liu Z
International Journal of General Medicine 2025, 18:179-189
Published Date: 15 January 2025
Integrating Therapeutic Drug Monitoring and Metabolomics to Explore Interindividual Variability in Lamotrigine Response in Epilepsy
Shen X, Jin R, Xu N, Miao Y, Zheng Z, Wang J, You M, Lin F, Hu L
Drug Design, Development and Therapy 2026, 20:593268
Published Date: 5 May 2026