")
Back to Journals » Journal of Inflammation Research » Volume 15
Dual Effect of the Extract from the Fungus Coriolus versicolor on Lipopolysaccharide-Induced Cytokine Production in RAW 264.7 Macrophages Depending on the Lipopolysaccharide Concentration
Authors Jędrzejewski T , Sobocińska J, Pawlikowska M, Dzialuk A , Wrotek S
Received 4 March 2022
Accepted for publication 26 May 2022
Published 20 June 2022 Volume 2022:15 Pages 3599—3611
DOI https://doi.org/10.2147/JIR.S364945
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Ning Quan
Tomasz Jędrzejewski,1 Justyna Sobocińska,1 Małgorzata Pawlikowska,1 Artur Dzialuk,2 Sylwia Wrotek1
1Department of Immunology, Faculty of Biological and Veterinary Sciences, Nicolaus Copernicus University, Torun, 87-100, Poland; 2Department of Genetics, Faculty of Biological Sciences, Kazimierz Wielki University, Bydgoszcz, 85-090, Poland
Correspondence: Tomasz Jędrzejewski, Department of Immunology, Faculty of Biological and Veterinary Sciences, Nicolaus Copernicus University, 1 Lwowska Str., Torun, 87-100, Poland, Tel +48 56 611 25 18, Email [email protected] Artur Dzialuk, Department of Genetics, Faculty of Biological Sciences, Kazimierz Wielki University, 10 Powstańców Wielkopolskich Ave., Bydgoszcz, 85-090, Poland, Tel +48 52 34 19 281, Email [email protected]
Purpose: Extract from the fungus Coriolus versicolor (CV) is classified as an immunological response modifier. Previously, we have shown that this extract induces interleukin 6 (IL-6)-related extension of lipopolysaccharide (LPS)-induced fever. This study investigated the effect of CV extract on the production of pro-inflammatory cytokines and the expression of components of signal transduction pathways leading to the secretion of cytokines from RAW 264.7 macrophages stimulated with different doses of LPS.
Methods: RAW 264.7 cells were stimulated with CV extract alone or co-treated with CV extract and LPS. The level of IL-6 and tumour necrosis factor α (TNF-α) in the culture media was measured using ELISA. Protein expression of Toll-like receptor (TLR) 4, phosphorylated IκB (p-IκB), CD14 glycoprotein and phospho-phosphatidylinositol 3-kinase (p-PI3K) was evaluated using Western blot. The effects of TLR4, nuclear factor κB (NF-κB) and p-PI3K on cytokine secretion were estimated using inhibitors: TAK-242, JSH-23 and LY294002.
Results: CV extract itself stimulates the secretion of IL-6 and TNF-α and increases the expression of TLR4, p-IκB and p-PI3K. The presence of CV extract during the treatment of cells with lower concentrations of LPS (10 and 100 ng/mL) increases the cytokine production. Co-stimulation of cells with CV extract and LPS at a higher dose (500 ng/mL) decreases the secretion of cytokines. This effect is related to the changes in the expression of TLR4, CD14 glycoprotein, p-IκB and p-PI3K.
Conclusion: This is the first report showing that the CV extract-induced production of cytokines is mediated by the PI3K signalling pathway. This extract acts antagonistically or additively with LPS on the production of IL-6 and TNF-α, depending on the LPS concentration. Our results are helpful for illustrating the mechanisms for the immunostimulatory effect of CV extract in inflammatory processes.
Keywords: Coriolus versicolor, pro-inflammatory cytokines, Toll-like receptor 4, phosphorylated IκB, phosphatidylinositol 3-kinase, CD14 glycoprotein
Introduction
Extract from the fungus Coriolus versicolor (CV extract) is a well-known agent with anti-tumour and immunomodulatory properties.1–3 Clinical studies have demonstrated that CV extract-related natural products have no toxic effect, and they improve the survival and quality of life of patients with cancers, hepatitis, hyperlipidaemia or chronic bronchitis,2,4,5 but the exact mechanism of their action is still poorly understood. It has been demonstrated that CV extract increases the activity of monocytes, the production of reactive oxygen species1 and the secretion of various pro-inflammatory cytokines, which is beneficial for patients with depressed immunity.2,4,6,7 In our previous research, we found that CV extract interferes with the crosstalk between breast cancer cells and macrophages by switching the macrophage phenotype from M2 to M1.8 We also observed inhibitory effects of CV extract on pro-inflammatory cytokine production by rat peripheral blood mononuclear cells (PBMCs)6 and human umbilical vein endothelial cells (HUVEC).9 However, our research on fever showed that CV extract extended the duration of LPS-induced fever in rats.10
Fever is a mechanism regulated, among others, by pro-inflammatory cytokines, and our previous research on mice with severe combined immunodeficiency (SCID) showed that in the process of fever induction, the involvement of the innate immune cells, such as macrophages, rather than lymphocytes, is needed.11 Based on these findings and the reports published by others showing different ways of macrophage activation depending on LPS concentration,12 we hypothesised that CV extract-induced effects differ depending on the severity of the inflammation.
Several studies have confirmed that macrophages are one of the main target cells of CV extract.8,13,14 These cells are both positive and negative regulators of inflammatory responses, playing a pivotal role in the development of inflammatory diseases.15 During initiation of inflammation, macrophages have three major function, namely antigen presentation, phagocytosis and immunomodulation through the production of various cytokines and growth factors. They also play a role in the maintenance and resolution of inflammation.16,17 Furthermore, Medzhitov described their role in the eradication of infectious microorganisms and in initiating the healing process.18
One of the most investigated models of inflammatory process is that induced by lipopolysaccharide (LPS). LPS is one of the best characterized pathogen-associated molecular patterns (PAMPs) derived from Gram-negative bacterial cell walls, which activates macrophages to initiate inflammation.19 The binding of LPS to the CD14/Toll-like receptor (TLR) 4 complex stimulates a cascade of effects, including degradation of phospho-IκB (p-IκB), and leads to the release of nuclear factor κB (NF-κB), which translocates into the nucleus. In consequence, the initiation of transcription of the pro-inflammatory cytokines, such as interleukin (IL) 6 and tumour necrosis factorα (TNF-α) is observed.20,21 In macrophages, it has been shown that LPS signalling can be regulated by the phosphatidyl-inositol-3 kinase (PI3K) pathway, which is also involved in a variety of different cell responses, such as cell survival, cell proliferation, and gene expression.22,23
In this study, we used two in vitro models that mimic mild and severe inflammation and analysed the signalling pathway leading to the expression of pro-inflammatory cytokines in macrophages treated with the CV extract. The aim of our research was to investigate whether CV extract is a modulator of the TLR4 signalling pathway and whether its effect differs in various (ie, mild and severe) inflammatory conditions.
Our study is the first report providing evidence that CV extract, depending on the LPS concentration, acts antagonistically or additively to LPS. This phenomenon is a consequence of its ability to affect the key factors involved in LPS-induced signalling [ie, TLR4, CD14 glycoprotein, p-IκB and phospho-phosphatidylinositol 3-kinase (p-PI3K)], leading to IL-6 and TNF-α expression.
Materials and Methods
Chemicals and Reagents
The CV extract was purchased from the MycoMedica Company (Police nad Metují, Czech Republic). Protein-bound polysaccharides, major soluble components of CV extract constituted 25% of the CV capsule and were isolated following described protocols.6,7,9 Briefly, the complete DMEM culture medium was used to dissolve the CV extract as a stock solution at a concentration of 4 mg/mL, which was extracted using continuous agitation (2500 rpm/min) for 48 h at room temperature. Insoluble material was removed by centrifugation at 2000 × g for 10 min. The soluble supernatant containing 1 mg/mL of the CV extract was sterilized using a 0.22-µm filter and stored at −80°C before use. The final concentration of protein-bound polysaccharides in the stock solution was calculated according to the manufacturer’s certificate of analysis and had been tested with the CP2010-UV method. The 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT), lipopolysaccharide derived from Escherichia coli 0111:B4 (LPS), protease inhibitor cocktail, phosphatase inhibitor cocktail 2 and 3 and all reagents used for cell culture were provided by Sigma-Aldrich (Darmstadt, Germany). The TAK-242 (TLR4 inhibitor) and JSH-23 (NF-κB inhibitor) were purchased from Cayman Chemical (Ann Arbor, MI, USA); LY294002 (PI3K inhibitor), rabbit anti-phosphorylated-IκB IgG, rabbit anti-mouse TLR4 IgG, rabbit phospho-PI3K p85 ((Tyr458)/p55 (Tyr199)) and rabbit-anti-mouse CD14 IgG were purchased from Cell Signalling Technology (Leiden, The Netherlands). Mouse anti-actin IgG was provided from MP Biomedicals (Santa Ana, CA, USA) and goat anti-mouse IgG or goat anti-rabbit IgG conjugated with horseradish peroxidase were obtained from Jackson ImmunoResearch (Cambridge, UK) and Merck Millipore (Burlington, MA, USA), respectively. Mouse IL-6 and TNF-α ELISA kits were purchased from Biorbyt Ltd (Cambridge, UK).
Cell Culture
Murine RAW 264.7 macrophage-like cells were purchased from the European Collection of Authenticated Cell Cultures (Salisbury, UK). The cells between 5 and 10 passage were cultured in a high-glucose (4.5 g/L) Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS), L-glutamine (1 mM), streptomycin (100 µg/mL) and penicillin (100 IU/mL) at 37°C, in a humidified atmosphere containing 5% CO2.
Cell Viability Assay
The effects of the CV extract and the inhibitors TLR4, PI3K, and NF-κB on cell viability were determined by the MTT assay. The cells (5 × 103/well) were pre-stimulated with 0.1% DMSO for 2 h followed by the treatment with CV extract (50, 100 and 200 µg/mL) for 24 h or pre-treated with the following inhibitors: TAK-242 (0.1 µM), JSH-23 (10 µM) and LY294002 (2 µM) for 2 h prior to stimulation with the CV extract for 24 h. All tested inhibitors were dissolved in DMSO and diluted with the culture medium; the final concentration of DMSO did not exceed 0.1% in the culture medium. The control cells were pre-treated with 0.1% DMSO or the inhibitors followed by incubation alone for 24 h. After treatment, the cells were incubated with a 0.5 mg/mL MTT solution for 3 h at 37°C, and the resulting purple formazan crystals were dissolved in DMSO. Optical density at 570 nm (with a reference wavelength of 630 nm) was measured using a Synergy HT Multi-Mode microplate reader (BioTek Instruments, Winooski, VT, USA). Cell viability was determined as a percentage of the untreated control cells, which served as 100%.
Cytokine Assay
Cells were seeded in 24-well tissue culture plates at a density of 0.1×106 cells/well and pre-incubated for 24 h. Subsequently, the macrophages were stimulated with the CV extract (50, 100 and 200 µg/mL) or co-treated with the CV extract and LPS (10, 100 and 500 ng/mL) for 24 h. The control cells were stimulated only with LPS or incubated alone in a complete DMEM. In separate experiments, to evaluate the roles of TLR4, PI3K and NF-κB in the cytokine production induced by the CV extract, the cells were pre-treated for 2 h with TLR4 inhibitor TAK-242 (0.1 μM), NF-κB inhibitor JSH-23 (10 µM) or PI3K inhibitor LY294002 (2 µM) prior to stimulation with the CV extract (200 µg/mL) for 24 h. In these experiments, the control cells were pre-treated with the inhibitors, followed by stimulation with LPS (100 ng/mL) for 24 h. After treatment, the total protein concentration of the viable cell pellets was determined using the Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. After stimulation, the culture plates were centrifuged at 800 × g for 5 min, and the culture media were collected and stored at −80°C. The concentrations of IL-6 and TNF-α were determined by standard ELISA kits according to the manufacturer’s instructions. Colorimetric changes in the assays were detected using the Synergy HT Multi-Mode Microplate Reader (BioTek Instruments, Winooski, VT, USA). Total amounts of the cytokines in the culture media were normalised to the total protein amount of the viable cell pellets.
Western Blot Analysis
Western blot analysis was performed to detect the protein levels of TLR4, CD14 glycoprotein, phosphorylated-IκB (p-IκB) and p-PI3K in the following RAW 264.7 macrophages: co-treated with the CV extract (200 µg/mL) and LPS (100 ng/mL or 500 ng/mL), stimulated only with CV extract or LPS and cultured in a DMEM for 1 h. After treatment, the cells were washed with ice-cold PBS and lysed in 100 µL of a buffer containing 4% SDS, 20% glycerol, 4% 2-mercaptoethanol, 0.004% bromophenol blue, 0.250 M Tris HCl (pH 6.8), 2 mM EDTA, protease inhibitor cocktail and phosphatase inhibitor cocktail. The protein concentration in the lysates was measured using a Pierce™ BCA Protein Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA), according to the manufacturer’s instruction. The lysates were supplemented with sample buffer and subjected to electrophoresis using a 12% precast polyacrylamide gel. After transfer onto nitrocellulose, the membranes were immunoblotted with the following antibodies: rabbit anti-mouse TLR4 IgG, rabbit anti-mouse CD14 IgG, rabbit anti-phosphorylated-IκB IgG, rabbit anti-phospho-PI3K p85 ((Tyr458)/p55 (Tyr199)) and mouse anti-actin IgG, followed, as appropriate, by anti-rabbit or anti-mouse IgG conjugated with horseradish peroxidase. Immunoreactive bands were visualised by chemiluminescence using SuperSignalWest Pico substrate (Thermo Fisher Scientific, Waltham, MA, USA) and analysed densitometrically using the ImageJ program (National Institute of Mental Health, Bethesda, MD, USA).
Statistical Analyses
The GraphPad Prism 7.0 software (GraphPad Software Inc., San Diego, CA, USA) was used for statistical analyses. All values are reported as mean ± standard error (SEM) and were analysed using analysis of variance (ANOVA) followed by Tukey’s multiple comparisons test, with the level of significance set at p < 0.05.
Results
Effect of CV Extract and Inhibitors of TLR-4 Signalling Pathway on RAW 264.7 Cell Viability
The cytotoxicity of the CV extract and the inhibitors used in the experiments was tested using the MTT assay (Figure 1). The RAW 264.7 cells were pre-treated with the inhibitors, such as TAK-242, LY294002 and JSH-23, followed by stimulation with various concentrations of the CV extract. The cell survival rates at all doses of the CV extract were above 86%. Importantly, the presence of all tested inhibitors during the stimulation of cells with the CV extract did not significantly change the viability of RAW 264.7 macrophages (cell viability was above 84%).
 |
Figure 1 Viability of RAW264.7 cells stimulated with different concentrations of CV extract in the presence or absence of the following inhibitors: TAK-242 (Toll-like receptor 4 inhibitor; (A)), JSH-23 (nuclear factor κB inhibitor; (B)) or LY294002 (phosphoinositide 3-kinase inhibitor (C)). Cell viability was assessed using the MTT assay. The results are presented as a percentages of the control cells cultured in DMEM (served as 100%; dash lines). The data are shown as the mean ± standard error of two independent experiments with six wells in each experiment. Asterisk denotes differences between the cells treated with the CV extract and/or the inhibitors compared with the control cells (*p < 0.05). |
Effect of TLR4, NF-κB and PI3K Inhibitors of TLR-4 Signalling Pathway on CV Extract-Induced Cytokine Production
Figure 2 shows the concentrations of IL-6 and TNF-α in the culture media collected after stimulation of RAW 264.7 cells with the CV extract only (grey columns). Compared with the unstimulated macrophages, the CV-treated cells produced significantly higher quantities of IL-6 and TNF-α in a dose-dependent manner. The IL-6 and TNF-α levels increased after CV extract treatment at doses of 50, 100 and 200 µg/mL, reaching maximum concentrations of 505.2 ± 26.7 pg/mL and 7668 ± 269 pg/mL for IL-6 and TNF-α, respectively. Upon stimulation with the CV extract at a dose of 200 µg/mL, the maximum increase in cytokine concentration relative to the unstimulated cells, approximately 30.8-fold for IL-6 and 14.2-fold for TNF-α, was observed.
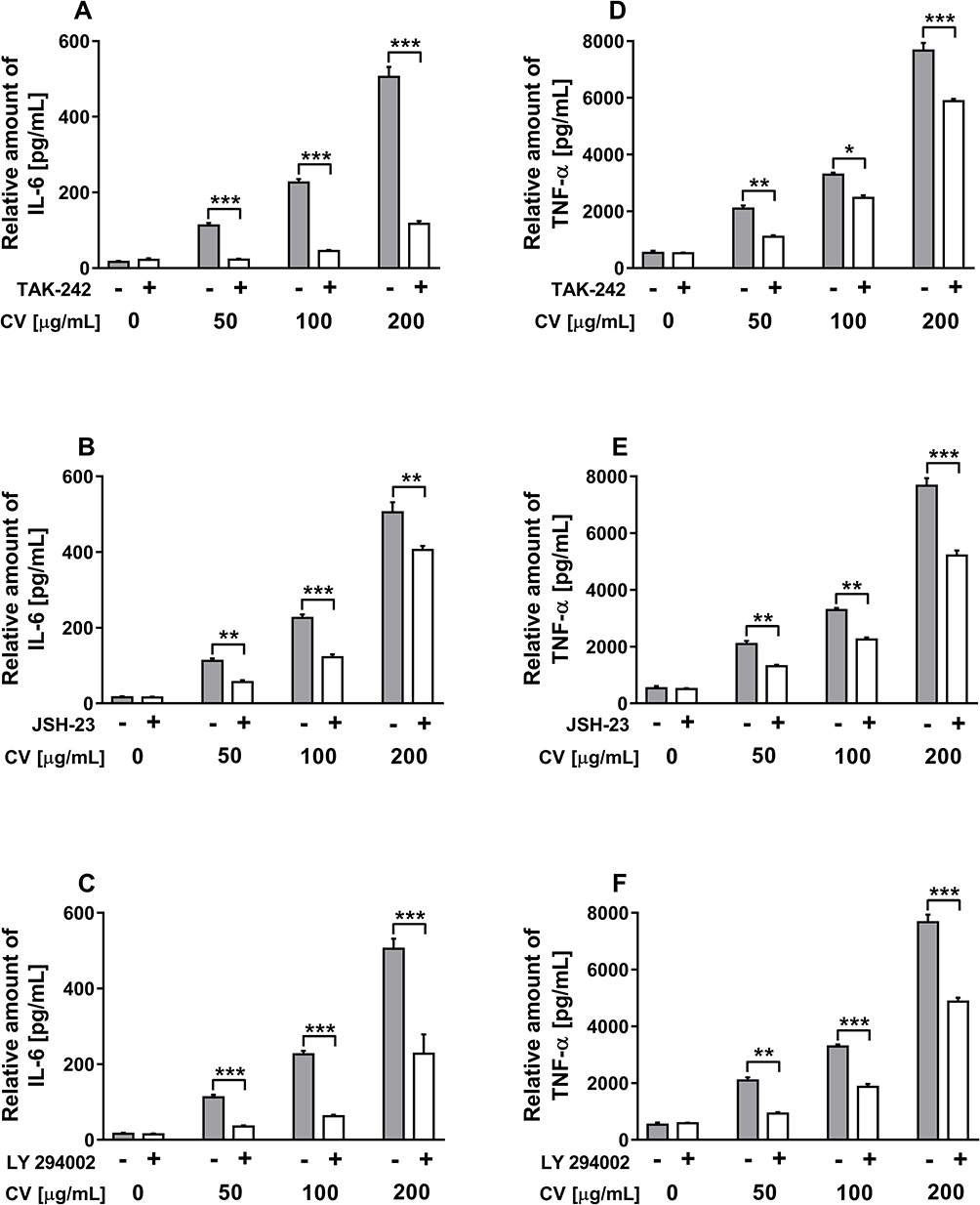 |
Figure 2 Effects of the selective inhibitors on the production of IL-6 (A–C) and TNF-α (D–F) in CV extract-stimulated RAW 264.7 cells. Cells were pre-treated with TLR4 inhibitor (TAK-242; 0.1 µM), NF-κB inhibitor (JSH-23; 10 µM), and PI3K inhibitor (LY294002; 2 µM) for 2 h prior to stimulation with CV extract (50, 100 and 200 µg/mL) for 24 h. Data are shown as the mean ± SEM of two independent experiments with five wells in each experiment. The concentration of the cytokines in the culture media was normalized to the total protein amount of the viable cell pellets and measured using ELISA. Asterisks indicate significant differences between the cells stimulated with the CV extract alone and the cells co-treated with the CV extract and the inhibitors (*p < 0.05; **p < 0.01; ***p < 0.001). |
Since it is well-established that the production of pro-inflammatory cytokines by macrophages is related with the TLR signalling pathway and NF-κB activation,24 in the present work, we investigated the effects of TLR4 and NF-κB on the CV extract-induced cytokine production using the following inhibitors: TAK-242 and JSH-23, respectively. It has also been shown that the PI3K signalling pathway is responsible for the expression of pro-inflammatory markers through IκB degradation and NF-κB activation.25 Therefore, we also studied the role of PI3K in cytokine production upon CV extract stimulation, using a PI3K inhibitor called “LY294002”. As shown in Figure 2, co-treatment of RAW 264.7 cells with the CV extract and all tested inhibitors significantly inhibited the CV extract-induced production of IL-6 (Figure 2A–C) and TNF-α (Figure 2D–F) compared with the cells stimulated with the CV extract only. This effect was observed for all tested doses of the CV extract. The control cells stimulated with the inhibitors alone did not release significantly different quantities of the cytokines compared with the untreated cells. The effectiveness of the tested doses of inhibitors on cytokine production was verified for the cells pre-treated with the inhibitors and then LPS in the same way as described for the treatment with the CV extract (Supplementary Figure 1).
Effect of CV Extract on the LPS-Induced Release of Pro-Inflammatory Cytokines from Cells
To assess the effect of CV extract on the LPS-induced production of IL-6 and TNF-α, the cells were co-stimulated with the CV extract (50, 100 and 200 µg/mL) and LPS (10, 100 and 500 ng/mL) for 24 h. The control macrophages were stimulated with LPS; as expected, these cells displayed notable increases in the levels of IL-6 and TNF-α in a dose-dependent manner (Figure 3). The production of pro-inflammatory cytokines was significantly increased by the CV extract in a dose-dependent manner when the cells were simultaneously stimulated with LPS at a dose of 10 and 100 ng/mL, except for the concentration of TNF-α produced by the cells co-treated with the CV extract and LPS at a dose of 10 ng/mL (Figure 3A, D, G and B, E, H, respectively). The opposite results were observed for the macrophages co-stimulated with various concentrations of the CV extract and LPS at a dose of 500 ng/mL. As shown, stimulation of RAW 264.7 cells with the CV extract and the highest tested concentration of LPS decreased the production of IL-6 (Figure 3C) and TNF-α (Figure 3F). This effect was observed was similar for all three tested doses of the CV extract (p < 0.001).
 |
Figure 3 Effect of CV extract on the production of IL-6 (A–C) and TNF-α (D–F) by RAW 264.7 cells stimulated with LPS. The cells were stimulated with various concentration of LPS alone (10, 100 and 500 ng/mL; grey bars) or with a combination of LPS and various doses of the CV extract (50, 100 and 200 µg/mL) for 24 h. The levels of pro-inflammatory cytokines released into the culture media were determined using ELISA assays. Data are shown as the mean ± SEM of two independent experiments with five wells in each experiment. The concentration of the cytokines was normalized to the total protein amount of the viable cell pellets. Asterisks indicate significant differences between the cells stimulated with LPS alone in comparison with the cells co-treated with the CV extract and LPS (*p < 0.05; **p < 0.01; ***p < 0.001). |
Effect of the CV Extract on LPS-Induced p-IκB, p-PI3K, TLR4 and CD14 Expression
To check whether the bilateral effect of cell co-stimulation with the CV extract and LPS is a result of different expressions of molecules involved in the TLR4 signalling pathways, Western blot analysis was applied. The expression of CD14 glycoprotein was also investigated since CD14 is a co-receptor especially required for LPS recognition by TLR4.26 Results of the Western blot analysis showed that the CV extract itself, similar to LPS, significantly increased the activation of TLR4, CD14 glycoprotein, p-IκB as well as p-PI3K in RAW 264.7 cells in comparison with unstimulated cells (Figure 4). The presence of the CV extract during the stimulation of cells with a lower concentration of LPS (100 ng/mL) had an additive effect on the protein expression of p-IκB and p-PI3K compared with the cells only treated with LPS (Figure 4B and C; p < 0.001 and p < 0.01, respectively). The opposite results were observed for the macrophages co-stimulated with the CV extract and a higher dose of LPS (500 ng/mL), where the CV extract decreased the protein expression of p-IκB and p-PI3K in the LPS-stimulated cells in comparison with the cells only stimulated with LPS at the same concentration (Figure 4B and C; p < 0.05 and p < 0.01, respectively). Moreover, the CV extract decreased the LPS-induced expression of TLR4 (Figure 4D) and CD14 (Figure 4E) both in the cells stimulated with LPS at a dose of 100 ng/mL (p < 0.05) as well as at a concentration of 500 ng/mL (p < 0.01 and p < 0.05, respectively).
 |
Figure 4 Representative Western blot images of p-IκB, p-PI3K, TLR4 and CD14 in RAW 264.7 cells co-stimulated with CV extract and LPS (A). The cells were stimulated only with CV extract (200 µg/mL) or LPS itself (100 or 500 ng/mL) or co-treated with CV extract and LPS for 1 h. Relative densitometric bar graphs showing the levels of p-IκB (B), p-PI3K (C), TLR4 (D) and CD14 (E) in the cell lysates were analysed by immunoblotting relative to the actin content. The blots were quantified by densitometry using the ImageJ software; results are expressed as a fold-change compared to unstimulated control cells. Data are shown as the mean ± SEM of three independent experiments with two repetition in each experiment. Asterisks indicate significant differences between stimulated and untreated cells (**p < 0.01; ***p < 0.001). Hash marks indicate a significant difference between the cells stimulated with the respective dose of LPS alone and the cells co-treated with LPS and CV extract (#p < 0.05; ##p < 0.05; ###p < 0.05). |
Discussion
The extract from CV fungus and its bioactive components have been widely used in the treatment of numerous diseases, including many types of cancers, hepatitis, hyperlipidaemia or chronic bronchitis.2,3,5,27 Although the effects of such therapy are encouraging, a signalling pathway induced by the CV extract is still poorly understood.2
Firstly, in the present paper, we examined how the CV extract itself affects the TLR4 signalling pathway and cytokine expression in immune cells. Our results showed that CV extract stimulated RAW 264.7 cells to secrete IL-6 and TNF-α in a dose-dependent manner. These findings are in line with studies previously published by us or other authors’ research on PBMCs,6 lymphocytes,7,28 endothelial cells,9 or dendritic cells.29
In this research, to confirm data about the involvement of TLR4 and NF-κB in CV extract-induced cytokine production,9,30 and to verify the role of the PI3K signalling pathway in this effect, we used the specific inhibitors TAK-242, JSH-23 and LY294002, respectively. These small-molecule inhibitors selectively bind to target proteins and reduce pro-inflammatory cytokine secretion from immune cells during their stimulation.31–33 They display desirable benefits over large-molecule inhibitors, such as monoclonal antibodies, including greater cell permeability and the ability to block intracellular target proteins and, through this, interfering with the downstream signalling pathways.34 Our results confirm for the first time, that the all tested inhibitors reduced CV extract-induced cytokine production. Additionally, we investigated whether TLR4 and factors such as p-PI3K and p-IκB are involved in a pathway leading to an enhanced cytokine production by measuring protein expression using the Western blot technique. We found that CV extract increased the levels of all above-mentioned proteins. Considerable levels of research have been devoted to understanding how CV extract interacts with immune cells; however, only few of them show that TLRs and NF-κB signalling pathways are involved in CV extract-induced immunomodulatory effects.29,35,36 Our results increase the knowledge about the immunomodulatory properties of CV extract, showing, for the first time, that its immunomodulatory properties are mediated by the PI3K signalling pathway, which was comprehensively confirmed on the level of protein expression and by using small-molecule inhibitors. Previously, it has been established in macrophages, that the PI3K pathway regulates the responses to different metabolic and inflammatory signals, modulates macrophage polarisation and controls cell survival, migration, and proliferation.15 Our results show that by using CV extract we can affect these processes.
For many years, CV extract has been one of the main subjects of research in our laboratory, among others, related to its influence on the processes of fever or inflammation. Previously, we have shown that this extract inhibited the LPS-induced synthesis of pro-inflammatory cytokines in PBMCs,6 MCF-7 human breast cancer cells and human umbilical vein endothelial cells.9 Taking into consideration that pro-inflammatory cytokines, such as IL-6 and TNFα, contribute to fever development, CV extract would be expected to suppress the fever. However, our study on LPS-induced fever in rats showed that CV extract did not reduce fever but rather extended its duration, which was accompanied by an increased plasma concentration of IL-6.10 In addition, there is more evidence that CV extract increases the sensitivity of immune cells to other stimuli and exerts a synergistic effect with other factors. For instance, peritoneal macrophages isolated from mice orally administered with CV extract showed increased TNF-α production in response to LPS stimulation.14 All these results indicate that CV extract induces different ways of immune cell activation in the inflammatory environment created by LPS. We hypothesized that the intensity of LPS-induced inflammation resulted from the LPS dose and may be a reason of these various effects. Therefore, in the present study, we investigated, in a model of mild and severe inflammation, whether CV extract modulates the TLR4 signalling pathways, leading to pro-inflammatory cytokine expression.
Our results show that the presence of CV extract during the stimulation of RAW 264.7 cells with LPS at a lower concentration (10 and 100 ng/mL) increased the production of IL-6 and TNF-α compared with the cells treated with LPS only. In contrast, CV extract decreased the levels of these cytokines when the cells were simultaneously stimulated with a high dose of LPS (500 ng/mL). So far, published data have shown that CV extract inhibited the pro-inflammatory cytokine production induced by LPS in rat PBMCs6 and murine peritoneal macrophages.37,38 However, in these works, the authors used primary cells stimulated with various doses of LPS. In a current paper, we provided evidence that CV extract modulates an inflammatory response of established monocyte/macrophage-like cells, depending on the LPS concentration.
Since there is evidence that the signalling pathway activated by the CV may be, at least partly, similar to the LPS-induced pathway (ie, TLR4, CD14 and NF-κB),9,30,35,36 in the present paper, we checked how the CV extract affects TLR4, CD14 and NF-κβ activation during co-treatment with different doses of LPS. Moreover, since a growing body of evidence reveals that the PI3K pathway is involved in TLR4 and NF-κB activation, leading to pro-inflammatory marker production in macrophages,25,39 we examined the level of PI3K in the macrophages co-stimulated with CV extract and different concentrations of LPS. To the best of our knowledge, this is the first study showing a role of PI3K in the cells treated with CV extract and LPS. Results of the Western blot analysis demonstrate that the presence of CV extract during the stimulation of cells with a lower concentration of LPS (100 ng/mL) had an additive effect on the protein expression of p-IκB and p-PI3K, whereas the co-treatment of cells with the CV extract and a higher dose of LPS (500 ng/mL) decreased their expression. These results correlate with the observed changes in the levels of pro-inflammatory cytokines and demonstrate the dual effect of CV extract on the macrophages stimulated with various doses of LPS. Since it is well-established that PI3K signalling enables NF-κB to enhance the activity of inflammatory mediator genes,40 and phosphorylation of IκB induces the nuclear translocation of NF-κB, leading to pro-inflammatory gene expression,41 our present study demonstrates that p-PI3K and NF-κB are involved in the dual effect of the CV extract on the different LPS-induced pro-inflammatory microenvironments.
The binding of LPS to the CD14/TLR4 complex activates NF-κB, which is a key transcription factor involved in the expression of pro-inflammatory cytokines.20 In contrast, only a few studies indicate that CV extract may activate immune cells via the TLR4 signalling pathway.30,35,36 Based on these reports, we investigated the effect of CV extract on the LPS-induced expression of TLR4 and CD14. Our results demonstrate that CV extract inhibited the expression of TLR4 and CD14 both in the RAW 264.7 cells stimulated with a lower (100 ng/mL) as well as a higher (500 ng/mL) concentration of LPS. These findings are in line with the results of Asai et al,37 who showed that CV extract displayed inhibitory activities against TLR4 in LPS-stimulated murine peritoneal macrophages through blocking the physical associations of LPS with LPS-binding protein (LBP) and CD14. These authors also demonstrated that this physical inhibition of TLR4 leads to the suppression of NF-κB activity. As we mentioned before, in our study, CV extract inhibited TLR4 and CD14 protein expression in macrophages stimulated with both concentrations of LPS (100 and 500 ng/mL). However, the decrease in the protein level of p-IκB was observed only when the cells were co-treated with CV extract and LPS at a dose of 500 ng/mL. The stimulation of cells with CV extract and LPS at a dose of 100 ng/mL had an additive effect on p-IκB expression. The dualistic effect of CV extract on the inflammatory environment can be associated with the involvement of the PI3K signalling pathway in the activation of CV extract-stimulated RAW 264.7 cells. As we have shown, the PI3K-related pathway modulates the production of pro-inflammatory cytokines upon stimulation with the CV extract itself as well as upon LPS and CV. Although the role of the PI3K pathway in the regulation of LPS signalling has been widely investigated,15,25,42 contradictory findings are still published, and it remains unclear how the PI3K pathway regulates the nuclear translocation of NF-κB and pro-inflammatory cytokine production.43 On the one hand, there is evidence that the activation of the PI3K pathway limits the LPS-induced activation of TLR and the NF-κB signalling pathways as well as the expression of inflammatory mediators in monocytes/macrophages.44–46 In contrast, other studies have shown that this pathway is required for LPS-induced NF-κB activation, which results in pro-inflammatory cytokine expression.47–49 Luyendyk et al indicate that these contradictory findings may be related to, in part, factors such as cell type, species or agonist used for stimulation.43 We presume that the LPS concentration and ability of the CV extract to activate the PI3K pathway may be the factors responsible for the dualistic effect of the CV extract in the inflammatory environment.
Conclusion
In summary, our results broaden the knowledge on the action of CV extract in an inflammatory environment. We indicate that, depending on the severity of inflammation, CV extract acts antagonistically or additively with LPS on the pro-inflammatory cytokine production in macrophages. To the best of our knowledge, this is the first report showing that the immunostimulatory properties of CV extract are mediated by the PI3K signalling pathway. However, additional, especially in vivo studies are required to clarify this effect. Our findings enable another step towards the more effective use of CV extract in clinical application since inflammation is a common phenomenon in patients suffering from various diseases (ie, cancers, hepatitis and chronic bronchitis).
Abbreviations
CV, Coriolus versicolor; DMSO, dimethyl sulfoxide; FBS, fetal bovine serum; JSH-23, nuclear factor κB inhibitor; IL, interleukin; LPS, lipopolysaccharide; LY294002, phosphatidylinositol 3-kinase inhibitor; PBMCs, peripheral blood mononuclear cells; p-IκB, phosphorylated IκB; p-PI3K; phosphatidylinositol 3-kinase; NF-κB, nuclear factor κB; TAK-242, Toll-like receptor 4 inhibitor; TLR4, Toll-like receptor 4; TNF-α, tumor necrosis factor α.
Data Sharing Statement
Supporting data used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Funding
This work was supported by the Polish Minister of Science and Higher Education under the program “Regional Initiative of Excellence” in 2019–2022 (Grant No. 008/RID/2018/19).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Sekhon BK, Sze DM, Chan WK, et al. PSP activates monocytes in resting human peripheral blood mononuclear cells: immunomodulatory implications for cancer treatment. Food Chem. 2013;138(4):2201–2209. doi:10.1016/j.foodchem.2012.11.009
2. Saleh MH, Rashedi I, Keating A. Immunomodulatory properties of Coriolus versicolor: the role of polysaccharopeptide. Front Immunol. 2017;(8):1087. doi:10.3389/fimmu.2017.01087
3. Habtemariam S. Trametes versicolor (Synn. Coriolus versicolor) polysaccharides in cancer therapy: targets and efficacy. Biomedicines. 2020;8(5):135. doi:10.3390/biomedicines8050135
4. Chang Y, Zhang M, Jiang Y, et al. Preclinical and clinical studies of Coriolus versicolor polysaccharopeptide as an immunotherapeutic in China. Discov Med. 2017;23(127):207–219.
5. Maehara Y, Tsujitani S, Saeki H, et al. Biological mechanism and clinical effect of protein-bound polysaccharide K (KRESTIN(®)): review of development and future perspectives. Surg Today. 2012;42(1):8–28. doi:10.1007/s00595-011-0075-7
6. Jędrzejewski T, Pawlikowska M, Piotrowski J, Kozak W. Protein-bound polysaccharides from Coriolus versicolor attenuate LPS-induced synthesis of pro-inflammatory cytokines and stimulate PBMCs proliferation. Immunol Lett. 2016;178:140–147. doi:10.1016/j.imlet.2016.08.013
7. Kowalczewska M, Piotrowski J, Jędrzejewski T, Kozak W. Polysaccharide peptides from Coriolus versicolor exert differential immunomodulatory effects on blood lymphocytes and breast cancer cell line MCF-7 in vitro. Immunol Lett. 2016;174:37–44. doi:10.1016/j.imlet.2016.04.010
8. Jędrzejewski T, Pawlikowska M, Sobocińska J, Wrotek S. Protein-bound polysaccharides from coriolus versicolor fungus disrupt the crosstalk between breast cancer cells and macrophages through inhibition of angiogenic cytokines production and shifting tumour-associated macrophages from the M2 to M1 subtype. Cell Physiol Biochem. 2020;54(4):615–628. doi:10.33594/000000244
9. Jędrzejewski T, Sobocińska J, Pawlikowska M, Dzialuk A, Wrotek S. Extract from the coriolus versicolor fungus as an anti-inflammatory agent with cytotoxic properties against endothelial cells and breast cancer cells. Int J Mol Sci. 2020;21(23):9063. doi:10.3390/ijms21239063
10. Jedrzejewski T, Piotrowski J, Kowalczewska M, Wrotek S, Kozak W. Polysaccharide peptide from Coriolus versicolor induces interleukin 6-related extension of endotoxin fever in rats. Int J Hyperthermia. 2015;31(6):626–634. doi:10.3109/02656736.2015.1046953
11. Kozak W, Wrotek S, Walentynowicz K, Waszak P. Fever and symptoms of sickness are present in mice lacking functional B and T lymphocytes. Acta Biol Crac Ser Zool. 2006;48:9–20.
12. Borzęcka K, Płóciennikowska A, Björkelund H, Sobota A, Kwiatkowska K. CD14 mediates binding of high doses of LPS but is dispensable for TNF-α production. Mediators Inflamm. 2013;2013(2013):824919. doi:10.1155/2013/824919
13. Chan SL, Yeung JH. Polysaccharide peptides from COV-1 strain of Coriolus versicolor induce hyperalgesia via inflammatory mediator release in the mouse. Life Sci. 2006;78(21):2463–2470. doi:10.1016/j.lfs.2005.10.011
14. Liu WK, Ng TB, Sze SF, Tsui KW. Activation of peritoneal macrophages by polysaccharopeptide from the mushroom, Coriolus versicolor. Immunopharmacology. 1993;26(2):139–146. doi:10.1016/0162-3109(93)90006-c
15. Vergadi E, Ieronymaki E, Lyroni K, Vaporidi K, Tsatsanis C. Akt signaling pathway in macrophage activation and M1/M2 polarization. J Immunol. 2017;198(3):1006–1014. doi:10.4049/jimmunol.1601515
16. Fujiwara N, Kobayashi K. Macrophages in inflammation. Curr Drug Targets Inflamm Allergy. 2005;4(3):281–286. doi:10.2174/1568010054022024
17. Watanabe S, Alexander M, Misharin AV, Budinger GRS. The role of macrophages in the resolution of inflammation. J Clin Investig. 2019;129(7):2619–2628. doi:10.1172/JCI124615
18. Medzhitov R. Inflammation 2010: new adventures of an old flame. Cell. 2010;140:771–776. doi:10.1016/j.cell.2010.03.006
19. Brightbill HD, Modlin RL. Toll-like receptors: molecular mechanisms of the mammalian immune response. Immunology. 2000;101(1):1–10. doi:10.1046/j.1365-2567.2000.00093.x
20. Ciesielska A, Matyjek M, Kwiatkowska K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol Life Sci. 2021;78(4):1233–1261. doi:10.1007/s00018-020-03656-y
21. Muniandy K, Gothai S, Badran KMH, Suresh Kumar S, Esa NM, Arulselvan P. Suppression of proinflammatory cytokines and mediators in LPS-induced RAW 264.7 macrophages by stem extract of Alternanthera sessilis via the inhibition of the NF-κB pathway. J Immunol Res. 2018;2018:3430684. doi:10.1155/2018/3430684
22. Cantley LC. The phosphoinositide 3-kinase pathway. Science. 2002;296(5573):1655–1657. doi:10.1126/science.296.5573.1655
23. Koyasu S. The role of PI3K in immune cells. Nat Immunol. 2003;4(4):313–319. doi:10.1038/ni0403-313
24. Dorrington MG, Fraser IDC. NF-κB signaling in macrophages: dynamics, crosstalk, and signal integration. Front Immunol. 2019;10(10):705. doi:10.3389/fimmu.2019.00705
25. Hawkins PT, Stephens LR. PI3K signalling in inflammation. Biochim Biophys Acta. 2015;1851(6):882–897. doi:10.1016/j.bbalip.2014.12.006
26. Ryu JK, Kim SJ, Rah SH, et al. Reconstruction of LPS transfer cascade reveals structural determinants within LBP, CD14, and TLR4-MD2 for efficient LPS recognition and transfer. Immunity. 2017;46(1):38–50. doi:10.1016/j.immuni.2016.11.007
27. Zong A, Cao H, Wang F. Anticancer polysaccharides from natural resources: a review of recent research. Carbohydr Polym. 2012;90(4):1395–1410. doi:10.1016/j.carbpol.2012.07.026
28. Ho CY, Lau CB, Kim CF, et al. Differential effect of Coriolus versicolor (Yunzhi) extract on cytokine production by murine lymphocytes in vitro. Int Immunopharmacol. 2004;4(12):1549–1557. doi:10.1016/j.intimp.2004.07.021
29. Engel AL, Sun GC, Gad E, et al. Protein-bound polysaccharide activates dendritic cells and enhances OVA-specific T cell response as vaccine adjuvant. Immunobiology. 2013;218(12):1468–1476. doi:10.1016/j.imbio.2013.05.001
30. Wang Z, Dong B, Feng Z, Yu S, Bao Y. A study on immunomodulatory mechanism of Polysaccharopeptide mediated by TLR4 signaling pathway. BMC Immunol. 2015;16:34. doi:10.1186/s12865-015-0100-5
31. Avni D, Glucksam Y, Zor T. The phosphatidylinositol 3-kinase (PI3K) inhibitor LY294002 modulates cytokine expression in macrophages via p50 nuclear factor κB inhibition, in a PI3K-independent mechanism. Biochem Pharmacol. 2012;83(1):106–114. doi:10.1016/j.bcp.2011.09.025
32. Matsunaga N, Tsuchimori N, Matsumoto T, Li M. TAK-242 (resatorvid), a small-molecule inhibitor of Toll-like receptor (TLR) 4 signaling, binds selectively to TLR4 and interferes with interactions between TLR4 and its adaptor molecules. Mol Pharmacol. 2011;79(1):34–41. doi:10.1124/mol.110.068064
33. Shin HM, Kim MH, Kim BH, et al. Inhibitory action of novel aromatic diamine compound on lipopolysaccharide-induced nuclear translocation of NF-kappaB without affecting IkappaB degradation. FEBS Lett. 2004;571(1–3):50–54. doi:10.1016/j.febslet.2004.06.056
34. Smith WM, Purvis IJ, Bomstad CN, et al. Therapeutic targeting of immune checkpoints with small molecule inhibitors. Am J Transl Res. 2019;11(2):529–541.
35. Yang SF, Zhuang TF, Si YM, Qi KY, Zhao J. Coriolus versicolor mushroom polysaccharides exert immunoregulatory effects on mouse B cells via membrane Ig and TLR-4 to activate the MAPK and NF-κB signaling pathways. Mol Immunol. 2015;64:144–151. doi:10.1016/j.molimm.2014.11.007
36. Price LA, Wenner CA, Sloper DT, Slaton JW, Novack JP. Role for toll-like receptor 4 in TNF-alpha secretion by murine macrophages in response to polysaccharide Krestin, a Trametes versicolor mushroom extract. Fitoterapia. 2010;81(7):914–919. doi:10.1016/j.fitote.2010.06.002
37. Asai Y, Takaori K, Yamamoto T, Ogawa T. Protein-bound polysaccharide isolated from basidiomycetes inhibits endotoxin-induced activation by blocking lipopolysaccharide-binding protein and CD14 functions. FEMS Immunol Med Microbiol. 2005;43(1):91–98. doi:10.1016/j.femsim.2004.07.008
38. Kuan YC, Wu YJ, Hung CL, Sheu F. Trametes versicolor protein YZP activates regulatory B lymphocytes - gene identification through de novo assembly and function analysis in a murine acute colitis model. PLoS One. 2013;8(9):e72422. doi:10.1371/journal.pone.0072422
39. Troutman TD, Bazan JF, Pasare C. Toll-like receptors, signaling adapters and regulation of the pro-inflammatory response by PI3K. Cell Cycle. 2012;11(19):3559–3567. doi:10.4161/cc.21572
40. Zhao Y, Qian Y, Sun Z, et al. Role of PI3K in the progression and regression of atherosclerosis. Front Pharmacol. 2021;12(12):632378. doi:10.3389/fphar.2021.632378
41. Oeckinghaus A, Ghosh S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb Perspect Biol. 2009;1(4):a000034. doi:10.1101/cshperspect.a000034
42. Stark AK, Sriskantharajah S, Hessel EM, Okkenhaug K. PI3K inhibitors in inflammation, autoimmunity and cancer. Curr Opin Pharmacol. 2015;23:82–91. doi:10.1016/j.coph.2015.05.017
43. Luyendyk JP, Schabbauer GA, Tencati M, Holscher T, Pawlinski R, Mackman N. Genetic analysis of the role of the PI3K-Akt pathway in lipopolysaccharide-induced cytokine and tissue factor gene expression in monocytes/macrophages. J Immunol. 2008;180(6):4218–4226. doi:10.4049/jimmunol.180.6.4218
44. Schabbauer G, Tencati M, Pedersen B, Pawlinski R, Mackman N. PI3K-Akt pathway suppresses coagulation and inflammation in endotoxemic mice. Arterioscler Thromb Vasc Biol. 2004;24(10):1963–1969. doi:10.1161/01.ATV.0000143096.15099.ce
45. Guha M, Mackman N. The phosphatidylinositol 3-kinase-Akt pathway limits lipopolysaccharide activation of signaling pathways and expression of inflammatory mediators in human monocytic cells. J Biol Chem. 2002;277(35):32124–32132. doi:10.1074/jbc.M203298200
46. Fukao T, Koyasu S. PI3K and negative regulation of TLR signaling. Trends Immunol. 2003;24(7):358–363. doi:10.1016/s1471-4906(03)00139-x
47. Ojaniemi M, Glumoff V, Harju K, Liljeroos M, Vuori K, Hallman M. Phosphatidylinositol 3-kinase is involved in Toll-like receptor 4-mediated cytokine expression in mouse macrophages. Eur J Immunol. 2003;33(3):597–605. doi:10.1002/eji.200323376
48. Park SJ, Lee SC, Hong SH, Kim HM. Degradation of IkappaBalpha in activated RAW264.7 cells is blocked by the phosphatidylinositol 3-kinase inhibitor LY294002. Cell Biol Toxicol. 2002;18(2):121–130. doi:10.1023/a:1015384201785
49. Saponaro C, Cianciulli A, Calvello R, Dragone T, Iacobazzi F, Panaro MA. The PI3K/Akt pathway is required for LPS activation of microglial cells. Immunopharmacol Immunotoxicol. 2012;34(5):858–865. doi:10.3109/08923973.2012.665461
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.