Back to Journals » Journal of Inflammation Research » Volume 19
Dihydromyricetin Modulates Neuropathic Pain and Depression: A Correlation with Hippocampal Vascular Endothelial Growth Factor in Mice
Authors Li T ![]() , Ye LQ, Liu HF, Chen XH, Wang Y
, Ye LQ, Liu HF, Chen XH, Wang Y ![]() , Chen ZM, Zheng Y
, Chen ZM, Zheng Y ![]() , Zeng WX, Li SE, Wan L
, Zeng WX, Li SE, Wan L ![]()
Received 30 May 2025
Accepted for publication 22 October 2025
Published 4 February 2026 Volume 2026:19 538350
DOI https://doi.org/10.2147/JIR.S538350
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ning Quan
Tian Li,1,2 Liu-Qing Ye,1,2 Hua-Feng Liu,1,2 Xiao-Hui Chen,1,2 Yu Wang,1,2 Zuo-Ming Chen,1,2 Yu Zheng,1,2 Wen-Xin Zeng,1,2 Si-En Li,1,2 Li Wan1
1Department of Pain Medicine, The State Key Clinical Specialty in Pain Medicine, The Second Affiliated Hospital, Guangzhou Medical University, Guangzhou, 510260, People’s Republic of China; 2Central Laboratory, The Second Affiliated Hospital, Guangzhou Medical University, Guangzhou, Guangdong, 510260, People’s Republic of China
Correspondence: Li Wan, Email [email protected]
Introduction: Neuropathic pain (NP) is a multidisciplinary problem of medicine, psychology, sociology and economics. The emotional disorders caused by neuropathic pain are seriously underestimated. The dynamic interaction between the NP and emotional disorders greatly increasing the difficulty in clinical treatment, highlighting the importance of finding an effective treatment for neuropathic pain combined with depression. Dihydromyricetin (DHM) has been shown to be effective in improving neurological diseases due to its potent anti-inflammatory and neuroprotective effects.
Methods: In this study, through network pharmacology and bioinformatics, the potential targets and signaling pathways of DHM in the treatment of neuropathic pain (NP) and depression (DP) comorbidities were predicted. Further animal experiments utilized the spared nerve injury (SNI) model in mice. Mice received daily intraperitoneal injections of DHM (20 mg kg− 1) or vehicle for 14 days. Mechanical sensitivity was assessed with the paw withdrawal mechanical threshold (PWMT), and depressive-like behaviors were assessed by tail suspension test immobility time, open field test total distance moved and central time. Hippocampal levels of vascular endothelial growth factor (VEGFA), vascular endothelial growth factor receptor 2 (VEGFR2), IL-1β, and TNF-α were quantified by Western blot and immunofluorescence staining.
Results: VEGFA was found to have the high “degree” value after ranking by target correlation based on the analysis from network pharmacology. SNI model evoked persistent the pain and depressive-like behavior that were paralleled by marked hippocampal down-regulation of VEGFA/VEGFR2 and a concomitant surge in IL-1β and TNF-α. However, the pain and depressive-like behavior can be relieved by DHM treatment. Both VEGFA and VEGFR2 were up-regulated in the hippocampus, and the changes of IL-1β and TNF-α were improved after DHM treatment.
Discussion: This study elucidates that DHM alleviates neuropathic pain and depressive-like behavior, potentially be related to the upregulation of the VEGFA/VEGFR2 signaling pathway and subsequent attenuation of neuroinflammation.
Keywords: neuropathic pain, depression, vascular endothelial growth factor
Introduction
Neuropathic pain (NP) affects about 10% of the population in the global.1 It often has a worse prognosis and poor treatment effect in clinical practice compared with other types of pain. Pain processing in the brain is complex, which can activate circuits involved in a variety of psychiatric disorders, such as fear, withdrawal, depression and anxiety.2 More than half of patients with neuropathic pain are suffer from depression and anxiety.3 Medications used to treat depression (TCAs and SNRI antidepressants) are known to be beneficial for neuropathic pain.4 It can be inferred that neuropathic pain and depression serve as mutual risk factors for each other. However, the exact physiological mechanism has not been revealed, so that significant challenges to the clinical management of chronic pain comorbid with depression. Consequently, the development of a therapeutic approach that simultaneously alleviates pain and depression is of utmost necessity.
Vascular endothelial growth factor (VEGFA) is an important neurotrophic factor and has been shown to have prominent effects on different brain regions of the central nervous system, especially the hippocampus.5,6 VEGFA affects long-term potentiation of neural signaling by regulating neurotransmitter transmission and synaptic plasticity.7 In animal models of sciatic nerve ligation, chronic constriction injury, or diabetic neuropathy, VEGFA blockade significantly attenuates nociceptive responses.8,9 The overexpression of VEGFA drives angiogenesis, which is a pivotal mechanism underlying peripheral sensitization and chronic inflammation.10 However, the role of VEGFA in pain signaling is controversial. Several studies have demonstrated that neuropathic pain is often accompanied by impaired nerve blood flow.11 Especially, there is a significant damage to the peripheral vasculature in diabetic neuropathic pain.11 Pharmacologic interventions in cancer patients, such as the administration of bevacizumab in combination with oxaliplatin to inhibit VEGFA binding to its endothelial receptor, have been shown to exacerbate neuropathy.12 Therefore, it is reasonable to conclude that VEGFA is deeply involved in the occurrence of neuropathic pain. In addition, a recent cross-species transcriptional analysis demonstrated that hippocampal vascular endothelial growth factor (VEGF), along with other growth factors such as fibroblast growth factor and insulin-like growth factor-1, is dysregulated in both patients with major depression and in a rat model of depression.13 In vitro experiments showed that VEGFA promoted the proliferation of mouse cortical neuronal progenitor cells, increased the diameter of neuronal cells, and increased the number of developing axons.14 At the cellular and behavioral levels, VEGFA was found to be essential for antidepressant response and was significantly decreased in serum of depressed patients and increased in serum of patients taking antidepressants for a long time.15 Therefore, the role of VEGFA in neuropathic pain and depression may be mediated by regulating its signaling pathways. So, we hypothesized that VEGFA might be a target for the comorbidity of neuropathic pain and depression (DP). However, it has not been reported that comorbid NP and DP affect VEGFA expression in the hippocampus.
Dihydromyricetin (DHM), a flavonoid derived from grapevine vine, boasts a diverse array of significant pharmacological properties, including anti-alcoholism, anti-inflammation, antibacterial, anti-oxidation, anti-tumor, lipid and glucose regulation, and neuroprotection.16 Given that DHM is capable of crossing the blood-brain barrier and diffusing into brain tissue, it shows great potential for application in the treatment of human neurodegenerative diseases.17 Recently, a growing body of research has elucidated the effects of DHM on the central nervous system, demonstrating its ability to mitigate oxidative stress and neuroinflammation, provide neuroprotection, and regulate cell proliferation.18–21 A consensus conclusion shown that DHM exerts analgesic and antidepressant effects by inhibiting the AGE-RAGE and PI3K/AKT signaling pathways, suppressing NLRP3 inflammasome activation and the expression of pro-inflammatory cytokines (IL-1β, IL-6, and TNF-α), and upregulating the neurotrophic factor BDNF.20–25 Based on the spread of bioinformatics and the improvement of various databases in recent years, network pharmacology has become an efficient method for the development of effective drugs.26 In this study, network pharmacology analysis showed that VEGFA is one of the potential targets of DHM. On this basis, we observed the changes of VEGFA in the hippocampus of mice with comorbid NP and DP which were induced by spared nerve injury (SNI), and explored whether the analgesic and antidepressant effects of DHM are related to the regulation of VEGFA expression in the hippocampus.
Materials and Methods
Animals
Male C57BL/6J mice aged 8–10-weeks and weighing 18–25 g were provided from Guangdong Laboratory Animal Center and kept in the Animal Center of the Second Affiliated Hospital of Guangzhou Medical University in the following conditions: temperature 20–22°C, light 8:00–20:00 bright/20:00–8:00 dark room, free drinking water and diet, and humidity 50%–60%. Any experimental design and operation should follow the international code of ethics for animal pain research and be approved by the Animal Ethics Committee of Guangzhou Medical University (No. GY-A2021-028).
Chemicals
Dihydromyricetin (MCE Biotechnology Corporation, USA) was formulated in 0.9% saline containing 0.2% dimethyl sulfoxide (DMSO) (PYG0040, Boster Biological Technology, China). In this experiment, dihydromyricetin was administered intraperitoneally (i.p)., and the injection dose was 20mg/kg according to body weight.19
Experimental Animal Grouping
All the animals were assigned random ear notch codes by an independent experimenter and were randomly divided into four groups: sham operation group (SHAM, n = 6), spared nerve injury group (SNI, n = 6), sham operation + dihydromyricetin group (SHAM+DHM, n = 6), and spared nerve injury + dihydromyricetin group (SNI+DHM, n = 6). Starting from 2nd week, in SHAM group and SNI group, the mice were given 0.1% DMSO dissolved in 0.9% saline (ip) daily for 14 days (days 15~28), after sham surgery or receiving spare nerve injury (SNI) to sciatica, respectively. In SNI + DHM group and SHAM+DHM group mice, the mice were accepted daily DHM (20 mg/kg) (ip) for 14 consecutive days (days 15~28), after SNI or after sham surgery, respectively. For behavioral experiments, the mechanical pain withdrawal thresholds (PMWT) were tested once a week after surgery, and the tail suspension test (TST) and open field test (OFT) were tested from the second week after surgery.
SNI Model
Firstly, mice were put into an anesthesia machine and anesthetized with isoflurane (RWD Life Science, China), then placed on a constant temperature heating plate for inhalation anesthesia apparatus. The left lower limb fur of mice was removed, and the skin was sterilized with 75% alcohol. The incision was about 1cm marked with the skin of the middle and lower 1/3 of the femur, the biceps femoris is bluntly separated with scissors, exposing the left entire sciatic nerve and identifying the branch of the sciatic nerve: the smallest and earliest branch on the right is the sural nerve, and the two thicker parallel ones on the left are the peroneal nerve and the tibial nerve. Then, a tweezer with a curved tip was used to pick out the peroneal nerve and the tibial nerve, the ends were tied with two threads of 3–0 about 1cm apart. The peroneal nerve and the tibial nerve were cut between the two ligation wires, and then the transected nerves were put back in place. Pay attention to refrain from damaging or touching the intact sural nerve throughout all steps. Lastly, the incision was closed with 3–0 wires. For the SHAM group, only the skin was cut and the biceps femoris was blunt-separated until the sciatic, sural, peroneal, and tibial nerves were clearly visible, and the wound was closed without touching any of these nerves.27
Behavioral Tests
Paw Withdrawal Mechanical Threshold
Paw withdrawal mechanical threshold (PWMT) was detected by von frey filaments (Ugo Basile SRL, Italy). Put the mice in an acrylic cylinder with a brown-black plastic film on a wire grid plate and acclimated for 30 min until the mice calmed down. Choose different specifications (0.04, 0.07, 0.16, 0.4, 0.6, 1, 1.4 and 2 g) von frey filament to stab left hind paws (between the third and fourth metatarsal) in mice, the filament should be bent at an angle with each touch, maintained for 3s and each time interval for 5 min. Immediate paw reduction or licking was marked as a positive response. If three or more positive responses occurred in five trials, they were recorded as the mechanical threshold. Each mouse underwent more than three repeated rounds of experiments, with at least a 10-min interval between each round.21 The final PWMT was represented by the mean of three repeated rounds of experiments.
Open Field Test
Mice were moved to the test environment for 1h before testing. Then the mice were placed in a box measuring 40 * 40 * 30 cm3. Each mouse was gently placed in the center of the open-field arena, and its behavior was video-recorded for 5 min. Recording videos were analyzed and processed using ToxTrac to determine the time in seconds that the mouse remained continuously in a positively central range that accounted for half the area of the entire box and the total distance moved in millimeters of the mice traveled in the whole box. The device was cleaned with 75% alcohol disinfectant before the next animal was put into the box.28
Tail Suspension Test
The mice were put into the test environment to adapt for 1 hour, attach the cellophane tape to the third end of the mouse’s tail to make it hang. The head of the mouse was 20 cm from the bottom of the device. The whole experiment was observed for a total of 6 minutes from the beginning of the suspension. The mice were allowed to adapt for the first 2 minutes without any recording, and the immobility time of the mice was recorded for the last 4 minutes. The immobility time was automatically quantified over the last 4 min using VisuTrack. The device was cleaned with 75% alcohol disinfectant before the next animal was put into. When the ethanol was completely evaporated, the experiment of the next mouse was started.29
A different experimenter, who was blinded to the group assignment, performed all behavioral tests. The mice were identified by ear notch codes only.
Western Blot
The animals were sacrificed under isoflurane anesthesia on 28th day after completing all behavioral tests. The hippocampal tissues were separated on ice, and the blood was washed with 4°C double distilled H2O. The tissues were blotted dry with absorbent paper, loaded into 1.5 mL EP tubes, and placed in jars containing liquid nitrogen to freeze for 30 ~ 60 min. RIPA buffer (P0013B, Beyotime, China), protease inhibitors and phosphatase inhibitors (P8340, Sigma-Aldrich, USA) were added for lysis. The tissue was ground fully at low temperature to avoid degradation and centrifuged at 12000 rpm for 30 min at 4°C. After centrifugation, separating into three distinct layers was visible; the upper supernatant was retained as the protein of the tissue, and the lower two layers were discarded. Protein concentrations were determined according to the instructions of the BCA kit (23225, Thermo Fisher Scientific, China). The samples were prepared at 2 ug/mL using SDS and double-distilled water and then placed at −20°C until use. Using Bio-Rad electrophoresis apparatus, 10 mL (20 ug) of each group of samples was aspirated into 10% SDS-polyacrylamide gel wells, and the proteins were separated by a constant pressure of 110 V and transferred to polyvinylidene difluoride (PVDF) membrane (Millipore, Billerica, USA) by constant current of 220 mA. 5% skim milk powder in 1* TBST configuration incubated PVDF membrane for 1 h at 23 ± 1°C. After further washing with 1* TBST, the PVDF membranes were incubated with rabbit antibodies (TNF-α (1:800, Proteintech, China), IL-1β (1:800, Proteintech, China), VEGFA (1:1000, Abcam, USA), VEGFR2 (1:1000, Cell Signaling Technology, USA), mouse antibodies GAPDH (1: 25000, Proteintech, China), TUBULIN (1: 25000, Proteintech, China) overnight at 4°C. The membranes were washed with 1* TBST every 5 min for four times. The corresponding HRP secondary antibodies were diluted in 1* TBST and incubated for 1 h at 23 ± 1°C. After washing with 1* TBST for four times every 5 min, then the bands signals were visualized by chemiluminescence on a Bio-Rad system. Bands were quantified using Image-J software, and target proteins were normalized against the reference gene GAPDH or TUBULIN.30
Immunofluorescence Staining
Anesthesia of the mice with isoflurane was required to ensure that the circulatory system was still present, and then cardiac perfused with PBS and 4% paraformaldehyde (PFA). The hippocampus tissues were obtained on ice and immersed in 4% PFA solution for fixation. The next day, the samples were immersed in 10%, 20%, and 30% sucrose solutions for gradient dehydration, and when the sample sank to the bottom, the next gradient of sucrose solution was replaced. The hippocampal tissue was cut into 10μm sections using a cryomicrotome (Leica).31 The sections were blocked in 10% BSA solution for 1 h at 23 ± 1°C, and incubated overnight in a 4°C refrigerator with primary antibody (rabbit anti-VEGFA, 1:200, Abcam, USA; Mouse anti-VEGFR2, 1:200, Abcam, USA; Mouse anti-NEUN, 1:200, Ptoteintech, China; rabbit anti-NEUN, 1:200, Ptoteintech, China). With 1* TBST washed, the second day the slide was incubated with secondary antibody (Alexa 594 goat anti-mouse 1:1000, Abcam, USA; Alexa 488 goat anti-rabbit 1:500, Abcam, USA) for 1 h at 23 ± 1°C. Sections were washed with PBS, then sealed with an anti-fluorescence quench containing DIPA (Abcam, USA) for 10 min and finally sealed with a cover slip and glycerin to seal the edge. Observation was performed using a Leica microscope (Olympus, Japan).
All harvested hippocampus tissues were labeled with ear notch codes by a third individual. Analysts performing Western blotting, immunofluorescence, and image quantification remained blind to group identity and codes were revealed only after statistical analyses were complete.
Network Pharmacological Analysis
Prediction of the Intersection Targets of DHM with Depression and Neuropathic Pain
Using the PubChem database to obtain the DHM specification SMILES string construction, the string input Swiss target prediction database and TargetNet database, p value >0 as a filter condition, to obtain drug targets. The above targets were input into Uniprot KB and “Homo sapiens” was selected to obtain the standardized format of drug target gene name. Importing the keywords “depression” and “neuropathic pain” enter Genecards database (correlation score >1 is selected), TTD database and OMIM database, to filtrate genes that intersect depression and neuropathic pain in the species “Homo sapiens”.24
Construction of Drug-Disease Target PPI Network
Using online tools Venny 2.1 to determine and visualize DHM with common targets between depression and neuropathic pain. The common genes obtained from the Venny graph were imported into the STRING database to obtain PPI relationships. Downloading the network diagram, and saving the results as TSV format files. Then imported the files into Cytoscape 3.9.1 software with degree centrality (DC) as the sorting standard for visual analysis.
Biological Function and Pathway Enrichment Analysis
Forty-five intersection targets import the Metascape database and set p-value <0.05 as screening threshold to execute Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis and annotate the Gene Ontology (GO). Select the biological processes, cellular component, molecular function of the top 10 GO terms and top 25 KEGG pathways imported into the Weishengxin platform to draw charts.
Statistical Analysis
All data were presented as mean ± standard error of the mean (SEM). Graphs were generated from GraphPad Prism 8 and Adobe Illustrator. Statistical analyses were performed by ImageJ and GraphPad Prism 8. In cases of multiple comparisons, a One-way analysis of variance (ANOVA) was used for statistical analysis, for longitudinal behavioral data over time, a Two-way repeated measures ANOVA was used for statistical analysis, followed by Fisher’s post hoc test, p < 0.05 was considered significant.
Result
DHM Increased the Paw Withdrawal Mechanical Threshold (PWMT) and Alleviated Depressive-Like Behaviors in Mice
As shown in Figure 1, we observed changes of PWMT and depressive-like behaviors in mice after modeling and after two weeks of DHM treatment. The PWMT of mice in the SNI group were significantly lower than those in the SHAM group, from the first day after modeling, the PWMT of the SNI group began to decrease, especially on the 3rd day, the difference was very significant between the SNI group and the SHAM group (p < 0.0001), and the pain sensitivity lasted until the 28th day (p < 0.0001). After 2 weeks of DHM treatment, the PWMT in SNI + DHM group was significantly increased compared to the SNI group (p = 0.0034). In the TST, the immobility time of the SNI group was much longer than that of the SHAM group at the second week after successful modeling (p = 0.0047). At the 3rd and 4th weeks of the SNI group, the immobility time exhibited an increasing trend compared to that observed during the 1st and 2nd weeks. It was also observed that the total moving distances of the OFT in the SNI group was shorter than in the SHAM group at the 2nd week (p = 0.0012), but there was no significant difference between the SNI group and the SHAM group at the 3rd or 4th week. The center time of OFT was lower in the SNI group than that in the SHAM group until the 3rd week (p = 0.0030) and 4th week (p = 0.0252). In addition, following two weeks of DHM treatment, the treatment group exhibited a significantly shorter immobility time in the TST (p = 0.0017) and a significantly longer center time in the OFT (p = 0.0095) compared with the SNI group.
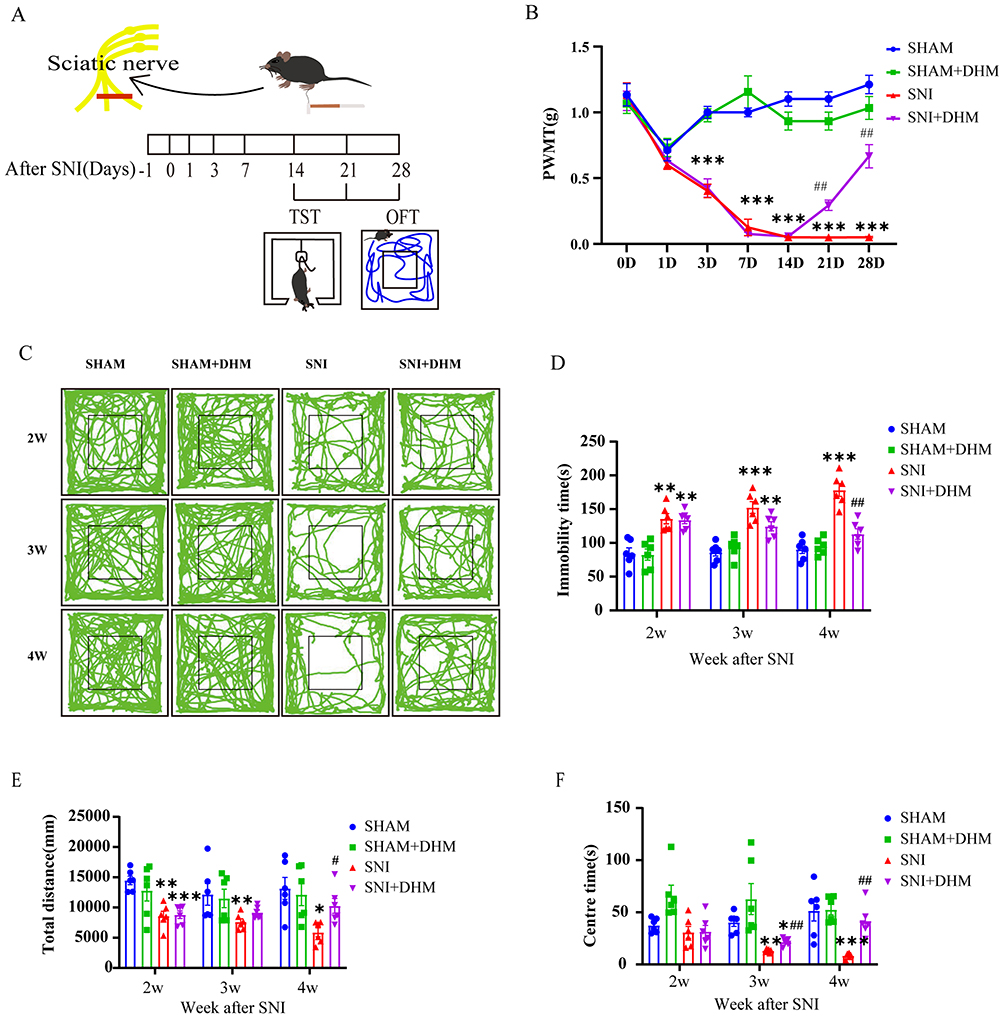 |
Figure 1 (A) Schematic of the SNI and behavior tests at the time points after the SNI model. (B) The PWMT of the SNI model and the effects of dihydromyricetin treatment in mice. (C) The total distance moved traveled in OFT of each group recorded. (D) The immobility time of the four groups in the TST. (E) The total distance moved of four groups in the OFT. (F) The center time of four groups in the OFT. n = 6, Two-way ANOVA, data are displayed as the means ± standard error of the mean, *p < 0.05, **p < 0.01 and ***p < 0.001 vs SHAM group; #p < 0.05 and ##p < 0.01 vs SNI group. |
The above data showed that the total distance moved by mice in SNI group in the OFT were lower than those in SHAM group at the second week after modeling, while the total distance moved of mice in the SNI group and SHAM group had no significant difference in OFT at the 3rd week after modeling. It is possible that SNI surgery affect the normal activities of mice within two weeks. After two weeks, the effect of surgery on mouse behaviors can be excluded. The immobility time of TST and the center time of OFT could represent the depressive behaviors of mice. So, our results showed that mice developed depressive symptoms 4 weeks after modeling, and DHM treatment for 2 weeks can significantly alleviate depression-related behaviors.
Summary of Disease Targets for Neuropathic Pain and Depression and DHM Drug Targets
To explore the potential targets and signaling pathways of DHM in the treatment of neuropathic pain with depression, we employed a network pharmacology approach. Specifically, as shown in Figure 2, we identified 1700 and 3751 genes associated with neuropathic pain and depression, respectively, from the GeneCards database. Additionally, we retrieved genes from the OMIM database (16 genes for neuropathic pain and 3 genes for depression), the TTD database (53 genes for neuropathic pain and 33 genes for depression), and the PharmGKB database (148 genes for neuropathic pain and 602 genes for depression). Furthermore, we obtained 70 and 114 potential targets of DHM from the Swiss Target Prediction platform and TargetNet database, respectively. Upon integrating and de-duplicating all the collected genes, we ultimately identified 1849 target genes for neuropathic pain, 4163 target genes for depression, and 148 target genes for DHM. Notably, through analysis using the Venny database, we pinpointed 45 genes that serve as potential targets for DHM in the treatment of both neuropathic pain and depression. We uploaded the 45 common targets to the STRING 12.0 database for analysis to construct the disordered protein–protein interaction (PPI) network map. Upon examination, we found that the gene galanin receptor 3 (Galr3) did not interact with any of the remaining 44 genes. As a result, the PPI network map displays only the 44 nodes and their interactions. The visualization of the disordered PPI network map was further refined according to the “degree” value using Cytoscape 3.9.1. In this network, the nodes represent the 44 common targets shared between the drug and the disease, while the line segments connecting the nodes signify the interaction relationships between the target proteins. Based on the “degree” value, we selected the top 25 targets for further analysis.
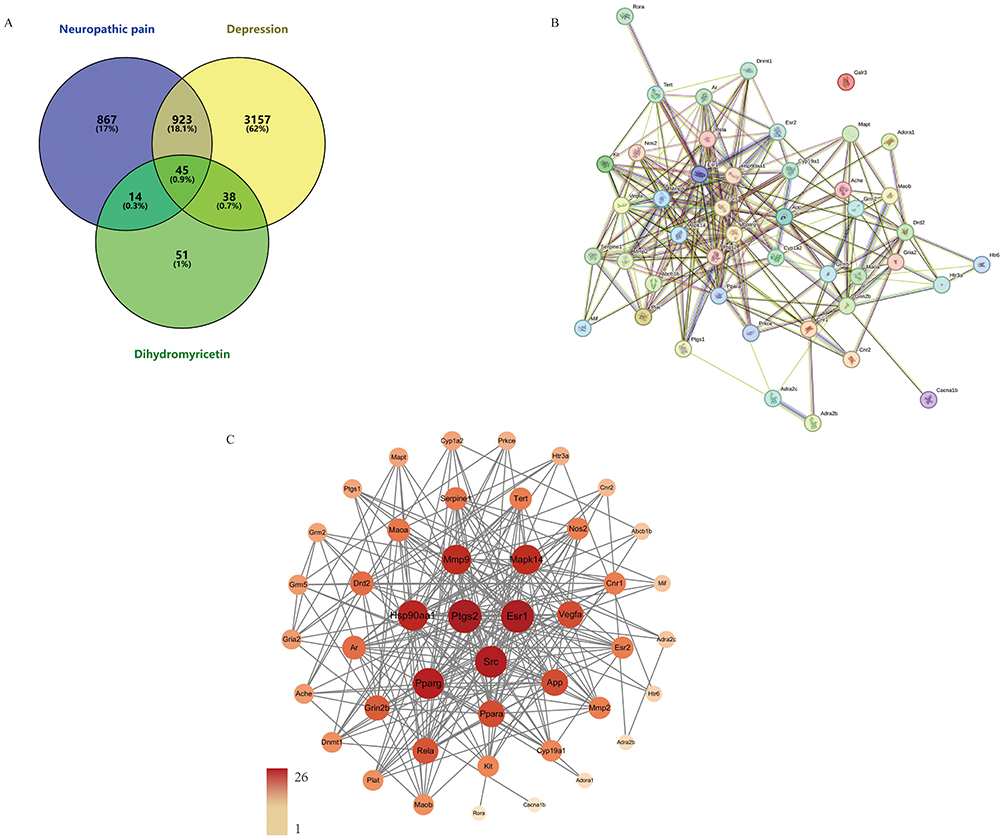 |
Figure 2 (A) Venn diagram of dihydromyricetin treatment for neuropathic pain and depression targets. (B) 45 common targets showed by disordered protein–protein interaction (PPI) network map. (C) 44 common targets showed by protein–protein interaction (PPI) network analysis. Both the size and color depth of the node are calculated from the “degree” value, the larger and darker nodes have a higher degree. |
Gene Ontology (GO) Enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Enrichment Analyses
The 45 common targets of DHM, neuropathic pain, and depression were subjected to Gene Ontology (GO) enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analyses by the Metascape database. As shown in Figure 3, this comprehensive analysis yielded a total of 85 KEGG signaling pathways and 1096 GO items, which were further categorized into 926 biological process (BP) items, 67 cellular component (CC) items, and 103 molecular function (MF) items. Under the significance threshold of (p < 0.01), the top 10 terms for each category (BP, CC, and MF) were selected based on their p-values for detailed analysis. The results indicate that these potential targets are primarily associated with the activity of neurotransmitter receptors, transmitter-gated ion channels, and nuclear (steroid) receptors, which play crucial roles in mediating neuropathic pain and depression.32–34 Similarly, the top 25 KEGG signaling pathways were selected and visualized in a map. These pathways include ligand-receptor interaction, calcium signaling pathways, IL-17 signaling pathways, retrograde endocannabinoid signaling, Rap1 signaling pathway, cAMP signaling pathway, AGE-RAGE signaling, estrogen signaling, prolactin signaling, and VEGF signaling pathways. These pathways are particularly relevant to the comorbidity of neuropathic pain and depression.32,35–39
 |
Figure 3 (A) Bubble plot showing the top 25 significantly enriched pathways from KEGG enrichment analysis. Pathways with more enriched genes are represented by larger bubbles. The color of the bubbles indicates the significance level based on the p value. (B) Bar graph showing the top 10 significantly enriched pathways obtained from GO enrichment analysis. Pathways with more enriched genes are represented by higher bars. The different color represents category. |
DHM Increase the Expression Levels of VEGFA, VEGFR2, IL-1β and TNF-α in the Hippocampus
The intensity of Western blot bands represents VEGFA, VEGFR2, IL-1β and TNF-α protein expression. According to the data, as shown in Figure 4, the expression levels of VEGFA (p = 0.0018) and VEGFR2 (p = 0.0033) in the hippocampus of the SNI group were lower than those of the SHAM group. After 2 weeks of DHM treatment, compared with the SNI group, the signals of VEGFA (p = 0.0058) and VEGFR2 (p = 0.0489) bands were enhanced in the SNI + DHM group. However, the expression of IL-1β (p < 0.0001) and TNF-α (p = 0.0006) protein was enhanced in the SNI group. After two weeks of DHM treatment, the expression of IL-1β (p = 0.0005) and TNF-α (p = 0.0077) protein was reduced.
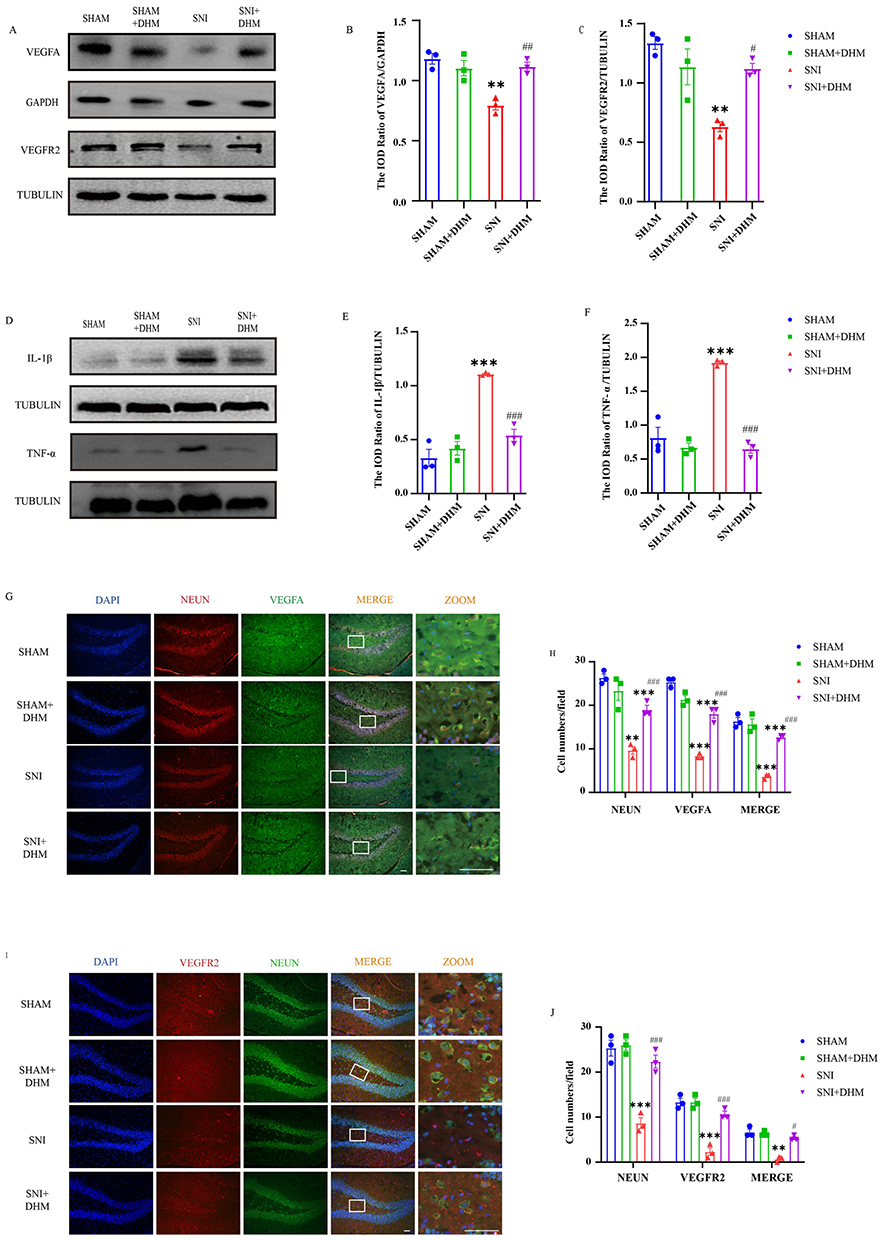 |
Figure 4 (A) Protein bands of VEGFA and VEGFR2 in the hippocampus. (B and C) Statistical analysis of VEGFA and VEGFR2 by normalizing to GAPDH or TUBULIN. (D) Protein bands of IL-1β and TNF-α in the hippocampus. (E and F) Statistical analysis of IL-1β and TNF-α by normalizing to TUBULIN. (G) Immunofluorescence staining of mice hippocampus: DAPI (blue), VEGFA (green), NEUN (red). Scale bar: 50μm. (H) Statistical analysis of VEGFA-positive, NEUN-positive expression and the double-positive expression of NEUN-VEGFA in the hippocampus in mice. (I) Immunofluorescence staining of mice hippocampus: DAPI (blue), VEGFR2 (red), NEUN (green). Scale bar: 50μm. (J) Statistical analysis of VEGFR2-positive, NEUN-positive expression and the double-positive expression of NEUN-VEGFR2 in hippocampus in mice. n = 3, One-way ANOVA, data are displayed as the means ± standard error of the mean, **p < 0.01, ***p < 0.001 vs SHAM group; #p < 0.05, ##p< 0.01 and ###p< 0.001, vs SNI group. |
Immunofluorescence double labeling was used to detect the immune reactivity in the hippocampus. NEUN represents mature neurons. The positive rate per visual field of NEUN, VEGFA, VEGFR2 (p < 0.0001) and the co-expression of VEGFA-NEUN (p < 0.0001) and VEGFR2-NEUN (p = 0.0005) in the hippocampus were lower in the SNI group than those in the SHAM group. After two weeks of DHM treatment, the positive rate per visual field of NEUN, VEGFA, VEGFR2 (p < 0.0001) and the co-expression of VEGFA-NEUN (p = 0.0006) and VEGFR2-NEUN (p = 0.0195) were increased in the SNI + DHM group.
Discussion
Clinically, patients with chronic pain are usually complicated with mental diseases such as anxiety, depression, and cognitive impairment. Depression is one of the most common comorbidities, and in turn, patients with depression are also prone to complicating of chronic pain.21 Traditional analgesic drugs or antidepressant drugs are far from meeting the needs of these patients, and most of them show poor prognosis and low therapeutic effect, which greatly affects the quality of life of patients and causes a terrible financial burden.40 This prompts us to pay attention to the mechanism of the comorbidity of chronic pain and depression and seek new therapeutic drugs according to the mechanism. Dihydromyricetin exhibits multiple pharmacological properties, including anti-inflammatory, antioxidant and neuroprotective effects.17 DHM can cross the blood-brain barrier, exerting positive effects on the brain without the risk of toxicity.41,42 In this study, chronic neuropathic pain was modeled by ligating and transection of the proximal tibial and common peroneal nerves, and pain status was assessed via PWMT. After SNI was established, the open field test and tail suspension test were used to evaluate the depression level of mice and to evaluate whether SNI models could induce neuropathic pain and depressive-like behavior, and whether dihydromyletin could improve pain behavior and depression behavior. With the help of network pharmacology analysis, we predicted that the alleviation of neuropathic pain with depression by DHM might involve complex signaling pathways and multiple targets. Cytoscape3.9.1 was used to rank the correlation of all intersecting targets, and the target VEGFA was determined to be the most strongly associated with neuropathic pain comorbid with depression. Then the protein expression levels of VEGFA/VEGFR2 and related inflammatory factors were detected by Western blot. The expression and co-localization of VEGFA/VEGFR2 and NEUN were detected by immunofluorescence double labeling. Our results strongly support that DHM treatment can significantly alleviate the persistent mechanical hyperalgesia and depression-like behavior at the 14th day to 28th day after SNI surgery. And the possible mechanism of DHM alleviate neuropathic pain is related to reducing the inflammation level of the central nervous system and normalizing the VEGFA/VEGFR2 signaling.
The changes of PWMT can accurately reflect pain behavior. The results of PWMT was significantly decreased after the model was established and was increased after DHM treatment. Depressive-like behaviors can be decomposed into anhedonia and manifestations of despair.43 In this experiment, depression-like behaviors were evaluated mainly from the locomotor activity and exploratory behavior of the mice. The time spent in the central zone in the open field test can reflect the exploration ability of mice. Meanwhile, mice in the SNI group spent significantly less time in the central area compared to those in the SHAM group, indicating a marked reduction in exploratory behavior. Conversely, mice in the DHM group exhibited an increased central zone exploration time. These findings suggest that the SNI group displayed depressive-like behavior characterized by diminished exploratory activity, whereas the treatment of DHM effectively alleviated these depressive-like symptoms. Due to the variety of depression-like behaviors, it is difficult to comprehensively respond to the situation by a single behavioral exploration. Therefore, the TST is another traditional method employed to assess depression-like behaviors in animals, which is by putting mice in an inverted suspension position to observe their desire to struggle.29 In our experiment, the SNI group showed an increase in immobility time at 2 weeks after modeling. However, the DHM treatment group showed a significant decrease in immobility time compared with the SNI group. Above results confirmed that SNI model can successfully simulate neuropathic pain and depression comorbidity in mice, and DHM has the potential to alleviate pain and improve depression-like behavior.
Network pharmacology is a systems biology approach that has been widely used to explore potential targets between drugs and diseases.44 In recent years, with the rapid improvement of the internet and the establishment and improvement of various databases, the improved accuracy and effectiveness have made it widely used to identifying effective therapeutic compounds for relevant diseases from traditional Chinese medicine. Based on the analysis of previous research finding and network pharmacology methods, DHM has multiple targets related to relieving chronic pain and antidepressant function. Among these core targets, most of them are related to inflammatory response, immune regulation and neurotransmitter transmission. Ten targets were identified with a high degree of relevance, including PTGS, SRC, ESR1, PPARG, HSP90AA1, MAPK14, MMP9, APP, VEGFA and PPARA. In addition, enrichment of these related targets by KEGG signaling pathway, 85 signaling pathways were screened. Based on the ranking by p-value, the top 25 signaling pathways were identified. Drawing on our group’s previous research findings, we determined that VEGFA acts on VEGFR2 and contributes to SNI-induced neuropathic pain via the p-Akt/TRPV1 pathway.30 Given this evidence, the VEGF signaling pathway was selected for further in vivo validation.
VEGFA is a member of growth factors, which is indispensable in the pathophysiology of angiogenesis, tumor progression and inflammatory factor recruitment.45 It has been proposed that VEGFA is involved in the signals communication between the vascular system and nervous system. In addition to its role in regulating vascular processes, VEGFA directly regulates neuronal function by activating VEGFR2.46 Therefore, more and more scholars have proposed that VEGFA may mediate the formation of chronic pain. Following partial ligation of the sciatic nerve (PSL), the levels of VEGFA in the spinal cord were observed to increase at both the protein and nucleic acid levels.10 Additionally, the administration of inhibitors targeting VEGFA and its receptor, VEGFR, has been shown to result in elevates mechanical and thermal pain thresholds.10 Given that VEGFA has neurobiological activities in the peripheral and central nervous systems, including neurotrophic and neuroprotective function, it directly modulates neuronal and glial cell functions.47 Several studies indicate that VEGFA, functioning as a neurotrophic factor, influences neurogenesis, thus suggesting it is a promising target for the development of antidepressants.48 Several studies considered that the baseline of VEGFA is down-regulated in the central system of depression model mice and clinically depressed patients.49,50 Meanwhile, both animal models and clinical applications of antidepressants have been shown to exert neuroprotective and nerve regeneration effects through VEGF signaling.50 We evaluated the alterations in VEGFA/VEGFR2 in the hippocampus of SNI-induced neuropathic pain in depressed mice. Western blot showed that the protein expression of VEGFA and VEGFR2 in the hippocampus of the SNI group was decreased. However, the decrease of VEGFA and VEGFR2 can be reversed after DHM treatment. The immunoreactivity of VEGFA and VEGFR2 in the hippocampus also showed decreased expression in the SNI group. Surprisingly, we observed that VEGFA-NEUN and VEGFR2-NEUN were strongly co-expressed in the dentate gyrus of the hippocampus. But the co-expression of VEGFA-NEUN and VEGFR2-NEUN in the hippocampus of the SNI group was decreased. These changes can also be reversed by DHM treatment. Our results indicate that the expression of VEGFA/VEGFR2 in the hippocampus of neuropathic pain combined with depression is decreased, which is different from that of chronic pain. This divergence may reflect region-specific or time-dependent responses: spinal VEGFA rise could support acute nociceptive transmission, while hippocampal VEGFA decline may underlie chronic affective-cognitive sequelae. Otherwise, the sample size for Western blot and immunofluorescence analyses in this study was set at n = 3 per group, consistent with sample sizes used in previous preliminary studies of similar pain-depression comorbidity models. We acknowledge that this limited sample size may affect the statistical robustness and reliability of the results. Further direct comparative studies employing larger sample sizes should be conducted to investigate VEGFA dynamics in both the spinal cord and hippocampus within the same model. DHM markedly reduces both pain and depressive-like behavior, an effect tightly linked to concurrent up-regulation of VEGFA/VEGFR2 expression. VEGFA-NEUN and VEGFR2-NEUN were highly co-expressed in the dentate gyrus of the hippocampus. We propose that a low VEGFA/VEGFR2 signaling mechanism is present in the hippocampus during the comorbidity of neuropathic pain and depression, while DHM demonstrates therapeutic potential in the comorbidity of neuropathic pain and depression, possibly be related to its activation of the VEGFA/VEGFR2 signaling pathway and its neuroprotective effects on neurons within the hippocampal dentate gyrus.
Studies have supported that both chronic pain and depressive-like behaviors are strongly related to neuroinflammation.51 At the same time, many treatments for neuropathic pain and depression (including neuromodulation, radiofrequency ablation, spinal cord stimulation, etc.) involve anti-inflammatory efficacy.52–54 Our results confirmed that the expression of IL-1β and TNF-α in the hippocampus was significantly higher in SNI group compared to the SHAM group. DHM treatment reduced the levels of these inflammatory factors. We hypothesized that the co-occurrence of pain and depression may be attributable to reduced expression of VEGFA/VEGFR2 in the hippocampus, which could be associated with elevated levels of inflammatory factors. Therefore, the alleviation of neuropathic pain and depression by dihydromyricetin is likely associated with the upregulation of VEGFA/VEGFR2 signaling and the concomitant reduction of neuroinflammation. The current experiments have not evaluated whether there is a causal relationship between these observed effects. To fully elucidate the underlying mechanisms, further investigation is warranted to explore whether the VEGFA signaling pathway is a necessary or sufficient condition for the analgesic and antidepressant effects of DHM and other and other mechanisms that may be implicated.
Conclusion
Our study demonstrates that DHM can effectively alleviate neuropathic pain and depression symptoms by SNI. This effect is associated with the upregulation of VEGFA and VEGFR2 expression in the hippocampus and the reduction of inflammatory factors IL-1β and TNF-α. These findings suggest that DHM may serve as a promising therapeutic agent for the treatment of comorbid neuropathic pain and depression.
Ethical Statement
All animal experiments were performed in accordance with the guidelines of the National Institutes of Health and the International Association for the Study of Pain and were approved by the Institutional Animal Care and Use Committee at Guangzhou Medical University (No. GY-A2021-028). All human data are exempt from ethical review and are in accordance with Article 32 of the “Notice on Issuing the Measures for Ethical Review of Life Sciences and Medical Research Involving Humans” (revised in 2023) of the People’s Republic of China, which states that “for life sciences and medical research involving humans that uses human information data or biological samples in the following situations, ethical review may be waived if it does not cause harm to human subjects, does not involve sensitive personal information or commercial interests, in order to reduce unnecessary burdens on researchers and promote the conduct of life sciences and medical research involving humans”. This study falls under the first item: “research using publicly available data obtained legally, or data generated through observation without interfering with public behavior”.
Funding
The research was supported by Guangzhou Science and Technolgy Bureau(2024A03J0198) and by the National Natural Science Foundation of China (No. 81771182) to Li Wan.
Disclosure
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
References
1. Vieira WF, Coelho DRA, Litwiler ST. et al. Neuropathic pain, mood, and stress-related disorders: a literature review of comorbidity and co-pathogenesis. Neurosci Biobehav Rev. 2024;161:105673. doi:10.1016/j.neubiorev.2024.105673
2. Navratilova E, Porreca F. Reward and motivation in pain and pain relief[J/OL]. Nat Neurosci. 2014;17(10):1304–14. doi:10.1038/nn.3811
3. Hooten WM. Chronic pain and mental health disorders: shared neural mechanisms, epidemiology, and treatment[J/OL]. Mayo Clin Proc. 2016;91(7):955–970. doi:10.1016/j.mayocp.2016.04.029
4. Finnerup NB, Attal N, Haroutounian S, et al. Pharmacotherapy for neuropathic pain in adults: systematic review, meta-analysis and updated NeuPSIG recommendations[J/OL]. Lancet Neurol. 2015;14(2):162. doi:10.1016/S1474-4422(14)70251-0
5. Cao L, Jiao X, Zuzga DS, et al. VEGF links hippocampal activity with neurogenesis, learning and memory[J/OL]. Nature Genet. 2004;36(8):827–835. doi:10.1038/ng1395
6. Denninger JK, Miller LN, Walters A, et al. Neural stem and progenitor cells support and protect adult hippocampal function via vascular endothelial growth factor secretion[J/OL]. bioRxiv. 2023;4(24):537801. doi:10.1101/2023.04.24.537801
7. De Rossi P, Harde E, Dupuis JP, et al. A critical role for VEGF and VEGFR2 in NMDA receptor synaptic function and fear-related behavior[J/OL]. Mol Psychiatry. 2016;21(12):1768–1780. doi:10.1038/mp.2015.195
8. Micheli L, Parisio C, Lucarini E, et al. VEGF-a/VEGFR-1 signalling and chemotherapy-induced neuropathic pain: therapeutic potential of a novel anti-VEGFR-1 monoclonal antibody[J/OL]. J Experiment Clin Cancer Res. 2021;40:320. doi:10.1186/s13046-021-02127-x
9. Xie X, Ma L, Xi K, et al. MicroRNA-183 suppresses neuropathic pain and expression of AMPA receptors by targeting mTOR/VEGF signaling pathway[J/OL]. Cell. Physiol Biochem. 2017;41(1):181–192. doi:10.1159/000455987
10. Kiguchi N, Kobayashi Y, Kadowaki Y, et al. Vascular endothelial growth factor signaling in injured nerves underlies peripheral sensitization in neuropathic pain[J/OL]. J Neurochem. 2014;129(1):169–178. doi:10.1111/jnc.12614
11. Schratzberger P, Walter DH, Rittig K, et al. Reversal of experimental diabetic neuropathy by VEGF gene transfer[J]. J Clin Investig. 2001;107(9):1083–1092.
12. Alejandro LM, Behrendt CE, Chen K, et al. Predicting acute and persistent neuropathy associated with oxaliplatin[J/OL]. Am J Clin Oncol. 2013;36(4):331–337. doi:10.1097/COC.0b013e318246b50d
13. Carboni L, Marchetti L, Lauria M, et al. Cross-species evidence from human and rat brain transcriptome for growth factor signaling pathway dysregulation in major depression[J/OL]. Neuropsychopharmacology. 2018;43(10):2134–2145. doi:10.1038/s41386-018-0117-6
14. Harms KM, Li L, Cunningham LA. Murine neural stem/progenitor cells protect neurons against ischemia by HIF-1α–regulated VEGF signaling[J/OL]. PLoS One. 2010;5(3):e9767. doi:10.1371/journal.pone.0009767
15. Ryan KM, McLoughlin DM. Vascular endothelial growth factor plasma levels in depression and following electroconvulsive therapy[J/OL]. Eur Archiv Psychiatry Clin Neurosci. 2018;268(8):839–848. doi:10.1007/s00406-018-0919-x
16. Wang R, Pi J, Su X, et al. Dihydromyricetin suppresses inflammatory responses in vitro and in vivo through inhibition of IKKβ activity in macrophages[J/OL]. Scanning. 2016;38(6):901–912. doi:10.1002/sca.21339
17. Chen J, Li M, Gao Q, et al. Dihydromyricetin, a flavonoid from vine tea (ampelopsis grossedentata) provides hepatoprotection by modulating gut microbiota-mediated bile acid homeostasis[J/OL]. J Agric Food Res. 2024;18:101376. doi:10.1016/j.jafr.2024.101376
18. Sun P, Yin JB, Liu LH, et al. Protective role of Dihydromyricetin in Alzheimer’s disease rat model associated with activating AMPK/SIRT1 signaling pathway[J/OL]. Biosci Rep. 2019;39(1):BSR20180902. doi:10.1042/BSR20180902
19. Guo CH, Cao T, Zheng LT, et al. Development and characterization of an inducible Dicer conditional knockout mouse model of Parkinson’s disease: validation of the antiparkinsonian effects of a sigma-1 receptor agonist and dihydromyricetin[J/OL]. Acta Pharmacol Sin. 2020;41(4):499–507. doi:10.1038/s41401-020-0379-5
20. Wei Y, Hu Y, Qi K, et al. Dihydromyricetin improves LPS-induced sickness and depressive-like behaviors in mice by inhibiting the TLR4/Akt/HIF1a/NLRP3 pathway[J/OL]. Behav Brain Res. 2022;423:113775. doi:10.1016/j.bbr.2022.113775
21. Ge H, Guan S, Shen Y, et al. Dihydromyricetin affects BDNF levels in the nervous system in rats with comorbid diabetic neuropathic pain and depression[J/OL]. Sci Rep. 2019;9:14619. doi:10.1038/s41598-019-51124-w
22. Guan S, Shen Y, Ge H, et al. Dihydromyricetin Alleviates Diabetic Neuropathic Pain and Depression Comorbidity Symptoms by Inhibiting P2X7 Receptor[J/OL]. Fronti Psychiat. 2019;10:770. doi:10.3389/fpsyt.2019.00770
23. Ge H, Sun M, Wei X, et al. Protective effects of dihydromyricetin on primary hippocampal astrocytes from cytotoxicity induced by comorbid diabetic neuropathic pain and depression[J/OL]. Purinergic Signalling. 2020;16(4):585–599. doi:10.1007/s11302-020-09752-9
24. Huang J, Chen B, Wang H, et al. Dihydromyricetin Attenuates Depressive-like Behaviors in Mice by Inhibiting the AGE-RAGE Signaling Pathway[J/OL]. Cells. 2022;11(23):3730. doi:10.3390/cells11233730
25. Zhang W, Yang L, Li L, et al. Dihydromyricetin attenuates neuropathic pain via enhancing the transition from M1 to M2 phenotype polarization by potentially elevating ALDH2 activity in vitro and vivo[J/OL]. Ann Transl Med. 2020;8(18):1151. doi:10.21037/atm-20-5838
26. Li T, Zhang W, Hu E, et al. Integrated metabolomics and network pharmacology to reveal the mechanisms of hydroxysafflor yellow a against acute traumatic brain injury[J/OL]. Comput Struct Biotechnol J. 2021;19:1002–1013. doi:10.1016/j.csbj.2021.01.033
27. Decosterd I, Woolf CJ. Spared nerve injury: an animal model of persistent peripheral neuropathic pain[J/OL]. Pain. 2000;87(2):149–158. doi:10.1016/S0304-3959(00)00276-1
28. Antiorio AT, Alemán-Laporte J, Zanatto DA, et al. Mouse behavior in the open-field test after meloxicam administration[J/OL]. JAALAS. 2022;61(3):270. doi:10.30802/AALAS-JAALAS-21-000046
29. Can A, Dao DT, Terrillion CE, et al. The tail suspension test[J/OL]. JoVE. 2012;2012(59):3769. doi:10.3791/3769
30. Peng Z, Yang F, Huang S, et al. Targeting vascular endothelial growth factor a with soluble vascular endothelial growth factor receptor 1 ameliorates nerve injury-induced neuropathic pain[J/OL]. Molecular Pain. 2022;18:17448069221094528. doi:10.1177/17448069221094528
31. Gorris MAJ, Halilovic A, Rabold K, et al. Eight-Color Multiplex Immunohistochemistry for Simultaneous Detection of Multiple Immune Checkpoint Molecules within the Tumor Microenvironment[J/OL. J Iimmunol. 2018;200(1):347–354. doi:10.4049/jimmunol.1701262
32. Gomez K, Santiago U, Nelson TS, et al. A peptidomimetic modulator of the CaV2.2 N-type calcium channel for chronic pain[J/OL]. Proc Natl Acad Sci USA. 2023;120(47):e2305215120. doi:10.1073/pnas.2305215120
33. Johnston JN, Henter ID, Zarate Jr CA. The antidepressant actions of ketamine and its enantiomers[J/OL]. Pharmacol Ther. 2023;246:108431. doi:10.1016/j.pharmthera.2023.108431
34. Witkin JM, Lippa A, Smith JL, et al. The imidazodiazepine, KRM-II-81: an example of a newly emerging generation of GABAkines for neurological and psychiatric disorders[J/OL]. Pharmacol Biochem Behav. 2022;213:173321. doi:10.1016/j.pbb.2021.173321
35. Austin PJ, Berglund AM, Siu S, et al. Evidence for a distinct neuro-immune signature in rats that develop behavioural disability after nerve injury[J/OL]. J Neuroinflammation. 2015;12:96. doi:10.1186/s12974-015-0318-4
36. Bolay H, Berman NE, Akcali D. Sex-related differences in animal models of migraine headache[J/OL]. Headache. 2011;51(6):891–904. doi:10.1111/j.1526-4610.2011.01903.x
37. Chen SC, Yh C, Song Y, et al. Upregulation of phosphodiesterase 7A contributes to concurrent pain and depression via inhibition of cAMP-PKA-CREB-BDNF signaling and neuroinflammation in the hippocampus of mice[J/OL]. Int J Neuropsychopharmacol. 2024;27(10):yae040. doi:10.1093/ijnp/pyae040
38. Mecca CM, Chao D, Yu G, et al. Dynamic change of endocannabinoid signaling in the medial prefrontal cortex controls the development of depression after neuropathic pain[J/OL]. J Neurosci. 2021;41(35):7492. doi:10.1523/JNEUROSCI.3135-20.2021
39. Medeiros GC, Greenstein D, Kadriu B, et al. Treatment of depression with ketamine does not change plasma levels of brain-derived neurotrophic factor or vascular endothelial growth factor[J/OL]. J Affective Disorders. 2020;280(Pt A):136. doi:10.1016/j.jad.2020.11.011
40. Bair MJ, Brizendine EJ, Ackermann RT, et al. Prevalence of pain and association with quality of life, depression and glycaemic control in patients with diabetes[J/OL]. Diabetic Med. 2010;27(5):578–584. doi:10.1111/j.1464-5491.2010.02971.x
41. Zhang J, Chen Y, Luo H, et al. Recent update on the pharmacological effects and mechanisms of dihydromyricetin[J/OL]. Front Pharmacol. 2018;9:1204. doi:10.3389/fphar.2018.01204
42. Martínez-Coria H, Mendoza-Rojas MX, Arrieta-Cruz I, et al. Preclinical research of dihydromyricetin for brain aging and neurodegenerative diseases[J/OL]. Front Pharmacol. 2019;10:1334. doi:10.3389/fphar.2019.01334
43. Hales CA, Stuart SA, Anderson MH, et al. Modelling cognitive affective biases in major depressive disorder using rodents[J/OL]. Br J Pharmacol. 2014;171(20):4524–4538. doi:10.1111/bph.12603
44. Hopkins AL. Network pharmacology: the next paradigm in drug discovery[J/OL]. Nat Chem Biol. 2008;4(11):682–690. doi:10.1038/nchembio.118
45. Claesson-Welsh L, Welsh M.
46. Stratton HJ, Boinon L, Gomez K, et al. Targeting the VEGF-a/neuropilin 1 axis for relief of neuropathic pain[J/OL]. Pain. 2022;164(7):1473. doi:10.1097/j.pain.0000000000002850
47. Carmeliet P, Ruiz de Almodovar C. VEGF ligands and receptors: implications in neurodevelopment and neurodegeneration[J/OL]. Cell Mol Life Sci Cmls. 2013;70(10):1763–1778. doi:10.1007/s00018-013-1283-7
48. Nowacka MM, Obuchowicz E. Vascular endothelial growth factor (VEGF) and its role in the central nervous system: a new element in the neurotrophic hypothesis of antidepressant drug action[J/OL]. Neuropeptides. 2012;46(1):1–10. doi:10.1016/j.npep.2011.05.005
49. Cui J, Gong C, Cao B, et al. MicroRNA-27a participates in the pathological process of depression in rats by regulating VEGFA[J/OL]. Exp Ther Med. 2018;15(5):4349. doi:10.3892/etm.2018.5942
50. Deyama S, Duman RS. Neurotrophic mechanisms underlying the rapid and sustained antidepressant actions of ketamine[J/OL]. Pharmacol Biochem Behav. 2019;188:172837. doi:10.1016/j.pbb.2019.172837
51. Campos ACP, Antunes GF, Matsumoto M, et al. Neuroinflammation, pain and depression: an overview of the main findings[J/OL]. Front Psychol. 2020;11:1825. doi:10.3389/fpsyg.2020.01825
52. Lurie DI. An integrative approach to neuroinflammation in psychiatric disorders and neuropathic pain[J/OL]. J Experiment Neurosci. 2018;12:1179069518793639. doi:10.1177/1179069518793639
53. Shen Y, Guan S, Ge H, et al. Effects of palmatine on rats with comorbidity of diabetic neuropathic pain and depression[J/OL]. Brain Res Bull. 2018;139:56–66. doi:10.1016/j.brainresbull.2018.02.005
54. Xu X, Chen R, Yu Y, et al. Pulsed radiofrequency on DRG inhibits hippocampal neuroinflammation by regulating spinal GRK2/p38 expression and enhances spinal autophagy to reduce pain and depression in male rats with spared nerve injury[J/OL]. Int Immunopharmacol. 2024;127:111419. doi:10.1016/j.intimp.2023.111419
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Research Progress on the Mechanism of Ketamine in Neuropathic Pain Comorbid Depression
Li Q, Liu S, Qin B, Liu X
Neuropsychiatric Disease and Treatment 2025, 21:1623-1636
Published Date: 9 August 2025
Depression and Insomnia Mediate the Link Between Problematic Internet Use and Neuropathic Low Back Pain: Evidence from a Cross-Sectional Survey
Radvanyi I, Tibold A, Herczeg V, Paphazi V, Beke S, Mák K, Hargitai-Kiss K, Feher G
Journal of Multidisciplinary Healthcare 2025, 18:7895-7905
Published Date: 7 December 2025