Back to Journals » Veterinary Medicine: Research and Reports » Volume 10
Diagnosing bluetongue virus in domestic ruminants: current perspectives
Authors Rojas JM ![]() , Rodríguez-Martín D
, Rodríguez-Martín D ![]() , Martín V
, Martín V ![]() , Sevilla N
, Sevilla N
Received 15 October 2018
Accepted for publication 31 December 2018
Published 14 February 2019 Volume 2019:10 Pages 17—27
DOI https://doi.org/10.2147/VMRR.S163804
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Young Lyoo
José M Rojas, Daniel Rodríguez-Martín, Verónica Martín, Noemí Sevilla
Centro de Investigación en Sanidad Animal (CISA-INIA), Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria, Valdeolmos, Madrid, Spain
Abstract:This review provides an overview of current and potential new diagnostic techniques against bluetongue virus (BTV), an Orbivirus transmitted by arthropods that affects ruminants. Bluetongue is a disease currently notifiable to the World Organization for Animal Health (OIE), causing great economic losses due to decreased trade associated with bluetongue outbreaks and high mortality and morbidity. BTV cross-reacts with many antigenically related viruses including viruses that causes African Horse sickness and epizootic haemorrhagic disease of deer. Therefore, reliable diagnostic approaches to detect BTV among these other antigenically related viruses are used or being developed. The antigenic determinant for differentiation of virus species/serogroups among orbiviruses is the VP7 protein, meanwhile VP2 is serotype specific. Serologically, assays are established in many laboratories, based mainly on competitive ELISA or serum neutralization assay (virus neutralization assay [VNT]) although new techniques are being developed. Virus isolation from blood or semen is, additionally, another means of BTV diagnosis. Nevertheless, most of these techniques for viral isolation are time-consuming and expensive. Currently, reverse-transcription polymerase chain reaction (RT-PCR) panels or real-time RT-PCR are widely used methods although next-generation sequencing remains of interest for future virus diagnosis.
Keywords: bluetongue virus, diagnostic tests, virus isolation, serological tools
Introduction
Bluetongue (BT) is an infectious disease transmitted by Culicoides biting midges, affecting mainly domestic and wild ruminants. The causative agent is bluetongue virus (BTV), one of the 22 species or serogroups in the genus Orbivirus in the Reoviridae family.1
BTV causes severe morbidity and mortality in sheep, while the infection is subclinical in some domestic and wild ruminants. The BTV genomes are composed of ten fragments of double-stranded RNA encoding seven structural (VP1–7) and four nonstructural (NS1–4) proteins.2,3 It is classified into 27 serotypes based on the genetic and antigenic features of the neutralizing protein VP2.4 VP7 protein is a major determinant of serogroup specificity, and most of the serological assays to detect BTV are based on detecting anti-VP7 antibodies.5 The distribution of BTV has changed drastically in the last decades. It was primarily distributed in tropical regions of the world but since 1998 outbreaks through the Mediterranean to the Northern Europe have been reported.6
BTV is notifiable to the World Organization for Animal Health (OIE), mainly because new outbreaks implicate movement and trade restrictions, causing severe economic losses. Vaccination with live-attenuated vaccines currently contributes to control BTV outbreaks.7 Nevertheless, active surveillance to detect BTV infection by either virus isolation or other detection test or serology is implemented. In this review, we provide an overview of current and new diagnostic procedures for BTV (summarized in Table 1).
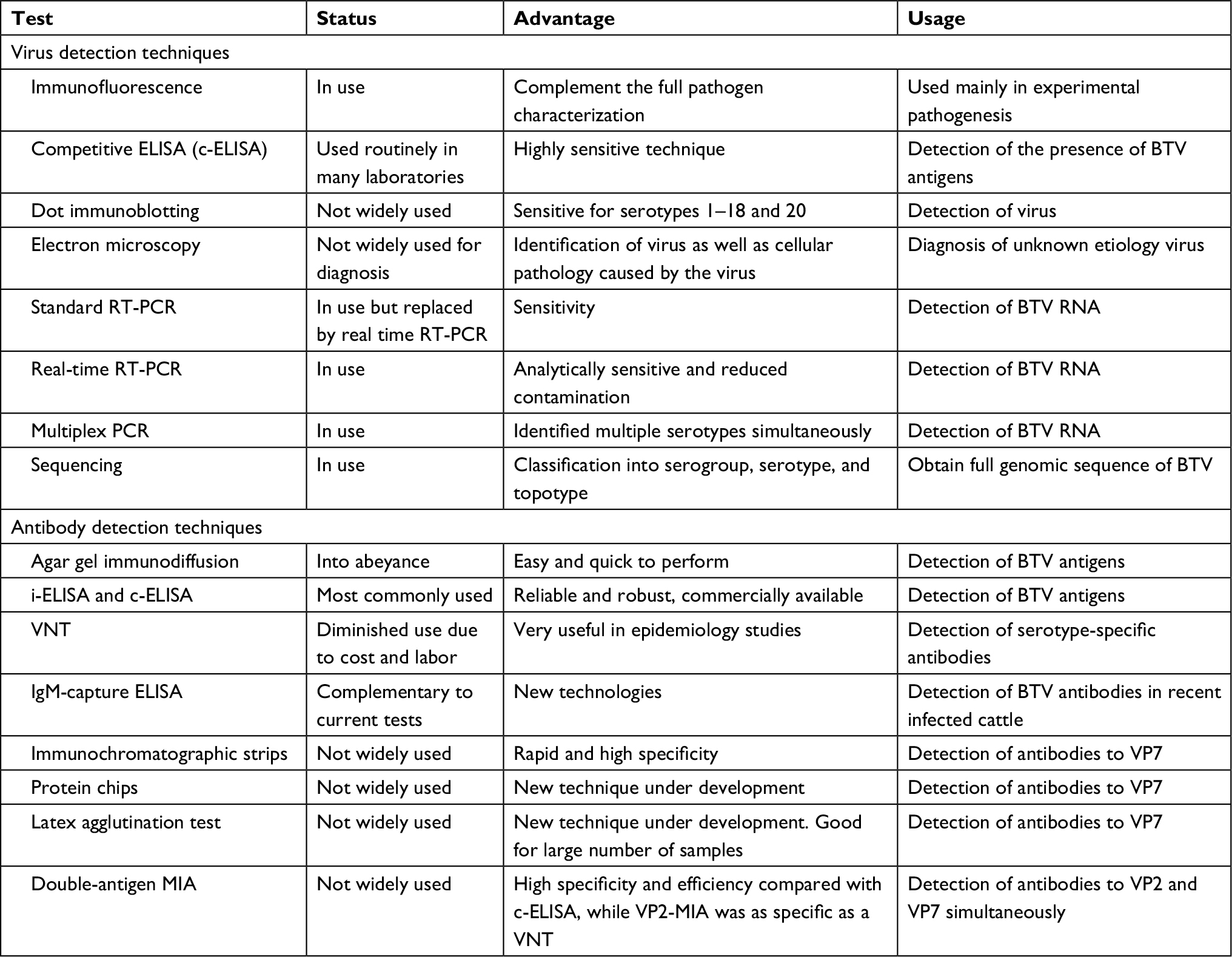 | Table 1 Virus detection techniques Abbreviations: BTV, bluetongue virus; i-ELISA, indirect ELISA; MIA, microsphere immunoassay; RT-PCR, reverse-transcription polymerase chain reaction; VNT, virus neutralization assay. |
BTV transmission and clinical signs
Transmission
BTV is an arbovirus, and until recently, its transmission was thought to be only mediated in cattle and ruminant through the bite of infected midges. This sole transmission route has been challenged recently with the emergence of reports of direct contact transmission with some serotypes and vertical transmission from mother to fetus.8
BTV is typically transmitted by the bite of Culicoides. Competent vectors were thought to be restricted to tropical and subtropical regions. However, outbreaks in North West Europe in 2006 have challenged this view, and it is now apparent that the virus can be transmitted by Culicoides species present in those latitudes.9 It is therefore critical to understand which Culicoides species are competent for BTV transmission. This is doubly important as this knowledge will allow for a better control of vector populations around livestock and for a better understanding of disease epidemiology.
Culicoides imicola in Africa and Asia, Culicoides brevitarsis in Australia, and Culicoides sonorensis in America are thought to be the main vectors in disease spread in those regions.10,11 C. imicola can now be found in the European Mediterranean basin and probably established itself from populations blown by winds from northern Africa.12 Culicoides obsoletus, Culicoides scoticus, and Culicoides pulicaris are nonetheless the most likely competent vectors that spread BTV in European northern latitudes.13–15 The distribution of these species in time and place matched BT outbreaks where C. imicola was absent.14,15 Moreover, BTV could be detected from wild-caught midge pools that did not contain C. imicola.15 Specific identification of the vector species responsible for outbreaks is however problematic as it often relies on morphological traits usually defined in male Culicoides, which limits the efficacy of trapping efforts as male do not contribute to BT epidemiology nor do they possess the same affinity for traps as female counterparts.16 Typically, midges are caught using light/suction traps, but different Culicoides species have preference for different light wavelengths.17,18 For instance, C. brevitarsis was found to prefer green light-emitting diode (LED) to incandescent light or ultraviolet (UV) LEDs,19 whereas C. obsoletus was caught in higher numbers with UV light traps than with incandescent light.20 It should be noted that odor baiting can greatly enhance Culicoides trapping.17 Carbon dioxide is a major attractant of biting midges to their blood host. Sampling around livestock and analysis of preferred host-feeding pattern are also important factors to consider when identifying the transmitting vector.
BTV diagnostic could be greatly helped by improving methodology for molecular identification of vectors in affected areas along with parallel detection of virus in captured Culicoides populations. This has helped better define the Culicoides competence in transmission of the recently discovered Schmallenberg virus (SBV). Culicoides pools for SBV testing were prepared from midge heads, while the rest of the carcasses were stored so that the number of infected individuals and species competence could be evaluated.21–23 The application of molecular strategies to define Culicoides vector competence in BTV similar to those applied to SBV could greatly help to identify the vectors competent for BTV spread in Europe and entomological surveillance. Therefore, this will have major implications for risk assessment of BTV incursion and spread.
BTV can be spread through the transport of infected livestock. To control this, restriction in animal movement and massive vaccination programs are usually set up when outbreaks occur. These measures are however not always effective as observed in Italy where infection has been spreading. Infection of wild ruminants can be damaging to these programs as they can act as a reservoir for the virus.10 The main risk of breaking contention restrictions however remains the dissemination of infected vectors. Culicoides are bad fliers, but their small size allows them to be carried by wind for long distances. Indeed, Culicoides movements due to wind have been reported over hundreds of kilometers.12,24 As a consequence, the regional spread of an outbreak is difficult to predict, which limits the efficacy of contention measures.
While in warmer African climates, BT outbreaks can occur throughout the year, in cooler climates like in the southern parts of South Africa the clinical disease usually becomes apparent in late summer and autumn and disappears with the first winter frosts.10 It has been suggested10 that this late emergence could be due to a BTV buildup in the vector during spring and early summer. Cattle rarely shows clinical signs but have prolonged viremia,25 which could contribute to this buildup effect. In Europe, the warm and wetter summers that facilitate vector propagation and the warmer winters that permit survival of some adults14,26 have allowed the disease to disappear overwinter and reemerge in spring. It is still unclear how this occurs, but a combination of these favorable climatological factors for the vector, the long BTV viremia detected in some animals, vertical transmission from mother to fetus, and infection of wild ruminants are likely to explain why BTV has become endemic in Southern European latitudes.
Although the bite of infected Culicoides remains the main source of BTV infection, it is critical to be aware of other possible routes of transmission. Vertical transmission from mother to fetus has been described.8,27,28 Transplacental infection is of great economic importance as it can result in abortions, still births, or nonviable offsprings. Oral transmission through ingestion of contaminated placenta is also possible.29 BTV can be detected in the semen of some viremic animals, which suggested that venereal transmission could be possible although this has not been documented.30 The use of shared needles for subcutaneous inoculations can also be a source of BTV transmission.31 Horizontal transmission has also been reported for some serotypes through close direct contact between animals in vector-free environments.27,32–34 The route of infection in those cases remains unclear although direct contact between animals appears essential. BTV is unlikely to spread horizontally over large distances; however, these findings should be considered in the context of closely housed farmed animals. Hygiene and veterinary practices in outbreaks should therefore be adapted accordingly.
BTV clinical signs
BTV can infect ruminants and camelids,35 but clinical disease is often more pronounced in sheep36–38 and in white tail deer.39,40 BTV seroconversion has been reported in numerous carnivores in Africa, although the epidemiologic significance of these findings remains unclear.41 Clinical signs however vary greatly depending on species and individual susceptibility. Cattle and goat are typically subclinically infected and are often considered as amplifying hosts in endemic regions,10 Although some serotypes like BTV-8 produce clinical disease in cattle, particularly in naïve region in Europe.36,37,42 In all livestock cases, BTV can nonetheless produce economic losses even in subclinical cases, as transplacental transmission can result in fetus malformation.8 Since subclinical infections are prominent in cattle and goats, it is important to set up sanitary control for BTV status in those animals to limit disease spread.
BT clinical signs are triggered by its preferred replication in endothelial cells.43 As a result of vascular damage, edema and hemorrhages take place. This damage in endothelial cells also produces a coagulopathy that participates in the hemorrhagic predisposition observed in BTV infections. Early signs of BT are however rather unspecific and resemble those of many other infectious diseases. Early clinical disease presents with pyrexia, lack of appetite, depression, or milk drop in lactating animals.44
In sheep, where clinical disease is more pronounced, disease usually manifest with fever, conjunctivitis, lacrimation, congestion of nasal and oral mucosa, and edema of the face and lips. In some cases, the clinical signs do not progress further, but in most cases these signs develop to more severe clinical features like salivation, mucopurulent nasal discharge, severe facial edema, and erosions, ulcerations, and hemorrhages in the nose, lips, and tongue.44 BT lesions are typically edematous, erosive, and hemorrhagic. These signs sometimes progress to cyanosis of the tongue, which gives its name to the disease. In most severe cases, pharyngeal and esophageal paresis and respiratory distress are observed. In most sheep, coronitis occurs 8–14 days after infection. This is characterized by lameness in more than one limb of variable intensity and diffused redness in the hoof coronary band. In most severe cases, serum exudate around the coronary band can be observed and animals appear hunched or kneeled and unwilling to stand or move.
In cattle, signs are usually milder than in sheep and only one or two signs manifest besides the pyrexia, lack of appetite, and depression.44 Cocirculation of several BTV serotypes in one region could possibly increase clinical signs in cattle in some cases.45
Considerations for BT differential diagnosis
Differential diagnosis for BT relies on the basic presentation of edema, ulcerations, and epithelial lesions. The type of lesions as well as their distribution in the infected animal can guide the veterinary practitioner toward a BT diagnosis. OIE notifiable diseases with similar presentation are one of the main instances in which a clinical differential diagnosis could help set up early control measures. Differential presentation traits will thus be broadly discussed for some of these diseases. Differential diagnosis should nonetheless also include endemic diseases.
BT initial signs are very similar to foot and mouth disease (FMD); however, FMD lesions are vesicular and erosive, whereas BT lesions are hemorrhagic, edematous, and erosive.44 Lesion distribution on the tongue also differs between FMD and BT. BT lesions are typically situated at the back and lateral borders of the tongue, whereas FMD lesions are situated at the tip and dorsum of the tongue. FMD lesions also rarely occur in eyes, whereas this is frequent in BTV infections.46 Disease transmission is also an indicator for differential diagnosis. BT presentation can be sporadic due to its transmission by biting midges, whereas FMD is highly contagious and as such morbidity will be high in the herd. Vesicular stomatitis is another disease that could be mistaken for BT. Lesion presentation is very similar to FMD, so the same criteria can be applied for differential diagnosis.
Peste des petits ruminants (PPRs) is another infectious disease with similar presentation as BT. In the case of PPRs, nasal discharge can also be mucopurulent, with lesions appearing in the oral and nasal tract, the nares, and the eyes. PPR lesions are hemorrhagic, necrotic, and erosive. Lesion distribution also differs from BT. They appear in the dorsum of the tongue, and respiratory infections are more characteristic of PPR. PPR also produces severe alimentary tract disease with diarrhea, but no coronitis. Moreover, like FMD, PPR is highly contagious, and thus, morbidity will be high in flocks.
Capripoxvirus-induced disease should also be included in the differential diagnosis. These viruses produce lumpy skin disease in cattle or sheep and goat pox. The lumpy skin disease nodular lesions47 and the sheep and goat pox pustular lesions that turn crusty48 are characteristically proliferative in these diseases. Their wide distribution in the skin of affected animal makes them distinguishable from BT lesions.
Ultimately, although these differential characteristics of BT presentation can guide a field diagnosis, only serological and molecular methods can provide a definitive BT diagnosis.
Overview of current tests for virus detection
Despite advances in molecular biology techniques, traditional diagnostic methods for the detection and identification of BTV remain in force. Different assays have been developed for the detection of BTV infection, based on either antibody detection or virus detection.49 An overview of current tests for antibody detection included in the serological techniques for diagnosis will be provided later in this review. This paragraph aims at summarizing the virologic techniques for BTV detection.
One of the biggest challenges in the diagnosis of BTV has been not only the detection but also differentiation of the different BTV serotypes. One important method for the identification of BTV is viral purification, adaptation, and amplification in embryonated chicken eggs, tissue culture, Culicoides-derived cells, or inoculation of susceptible ruminants. At the beginning of 1990s, some diagnostic tests were implemented based on reverse-transcription polymerase chain reaction (RT-PCR) that allowed the detection of a limited number of serotypes. Parts of the viral genome were specifically amplified in a faster and cheaper way than the virus isolation, providing precise information on serogroup, serotype, and topotype, but these assays required to run the samples on agarose gels, which made them difficult to implement in the field or in developing countries. These procedures have been improved along the years, and many RT-PCR protocols are now available, validated, and published. The next step has been the use of micorarrays50 and second- and third-generation sequencing51 for virus diagnostic, but these techniques are expensive and difficult to implement and maintain routinely for diagnosis and surveillance.
Virus isolation
BTV can be isolated from different types of samples:52
- Blood samples: BTV hosts present a transient viremia with a peak before the seroconversion, usually concomitant with fever. BTV has been found associated with red blood cells, and the isolation is possible from heparinized or sodium citrated whole blood.
- Semen samples: BTV transmission through semen in sheep and cattle has been suggested,53 and raw or extended semen samples can be used for virus isolation.54
- Tissue samples: Homogenized samples of different infected organs from the host collected in aseptic conditions and refrigerated also allows BTV isolation.55 Dead animals or aborted fetuses are optimal to get lung, spleen, lymph nodes, brain, bone marrow, and liver samples,55–57 while live animals can also be a source of BTV, from red bone marrow biopsy of adult sheep or liver and spleen from lambs. BTV can be obtained from swabs from the different lesions produced during the disease, through the digestive, vascular, and respiratory tracts.
The most sensitive method for BTV isolation is the intravenous inoculation of 10- to 11-day-old embryonated chicken eggs58 with sonicated blood or tissue suspension samples obtained from infected animals. In spite of the reduced isolation efficiency after virus inoculation of tissue culture cells with positively diagnosed samples, many cell lines are also used for this purpose such as the one derived from Culicoides variipennis or sonorensis midges (KC)59,60 or mouse L, baby hamster kidney (BHK-21), African green monkey kidney (Vero), or Aedes albopictus clone C6/36 (AA). However, as an alternative, that is, not routinely used, sheep can be inoculated intravenously or subcutaneously with washed blood cells or tissue homogenates from infected animals to amplify and isolate the virus.
Virus characterization
Once the virus is isolated, it can be serogrouped and serotyped.
Serogroup techniques are based on the reactivity of the antisera against conserved proteins, such as VP7, among Orbivirus isolates. Another serogroup, epizootic hemorrhagic disease virus, could be mistaken with BTV due to the cross-reactivity between them. For this reason, the use of monoclonal antibodies (MAbs) instead of polyclonal antibodies is recommended.
- Immunofluorescence (IFA): Different research groups have developed MAbs that bind specifically to different Orbivirus isolates, usually based on the VP7 protein, that is conserved within serogroup, allowing its recognition. These MAbs or anti-BTV sera are employed as primary antibodies in IFA procedures to recognize specifically BTV being detected with secondary antibodies coupled to different fluorophores.61–63 An alternative technique is the immunoperoxidase method used to detect BTV antigen in fixed cells or tissues,64 improving the sensitivity with the peroxidase–antiperoxidase detection, using BTV antiserum raised in sheep as primary antibody,65 or the avidin–biotin complex detection.66
- Competitive antigen capture ELISA (competitive ELISA [c-ELISA]): The virus is detected with a double antibody sandwich ELISA. A guinea pig and a murine hyperimmune serum are used for BTV capture, and a third conjugated goat anti-murine antibody is used to develop the assay.67 Despite being considered a highly sensitive technique, it has been questioned as a diagnostic tool for BTV detection from blood samples due to false negative registered as well as cross-reactivity between serotypes.68
- Dot immunoperoxidase assay: In this technique, the immune reaction takes place on nitrocellulose membranes where the samples are absorbed as dots and incubated with MAbs directed to the major group-specific antigen of BTV, detected with horseradish peroxidase-conjugated immunoglobulin showing a color reaction in the presence of H2O2 and a precipitating chromogen.58 Samples from BHK-21 cells infected with available BTV serotypes 1–18 and 20 but no other Orbivirus isolates are identified with this assay. However, as with the ELISA, BTV detection directly in infected sheep blood cells or tissue suspensions does not work.69
- Electron microscopy: BTV serogroup particles can be detected by transmission electron microscopy,70,71 but immunomicroscopy techniques are needed to identify BTV, for example, with a specific MAb to BTV and gold-labeled protein-A.72
- Viral genome detection: The Northern and Southern blot techniques or dot blots and in situ hybridizations are used to detect BTV genome through hybridization with cDNA or RNA probes from different segments of the BTV genome and serotypes. Although these assays are sensitive and specific, hybridization techniques are difficult to perform requiring special treatment of the samples, such as electrophoresis (most of them) and optimization of the test conditions. They do not present the optimal properties to be included as routine diagnostic tests. These techniques have been displaced by the PCR or RT-PCR development. Specific oligonucleotide pairs have been developed for serogrouping and serotyping73 BTV genome. The amplified product can be identified by hybridization,74–80 enzyme restriction, or sequencing techniques. A range of RT-PCR assays targeting different BTV genome segments have been developed since the early 1990s, but they were only able to detect some of the BTV serotypes.76,77,81,82 The rapid evolution of RT-PCR tests has resulted in many assays currently in use with different extraction and amplification methods (reverse transcriptase and polymerase enzymes, primers, or conditions), including the real-time PCR, the main diagnostic tool used in Europe for the detection of BTV in infected ruminants since 2006,83 which is able to differentiate simultaneously all BTV serotypes. For example, Hofmann et al developed a real-time PCR based on the nonstructural protein 3 (NS3) fragment of the BTV genome84 or the nested PCR85 targeting the NS1 gene, both able to differentiate 25 of 27 BTV serotypes. In the same way, duplex assays using two primer sets, one targeting the VP7 genome segment and the other the segment 1 (encoding the viral polymerase, VP1), were published.86 Many real-time RT-PCR kits have been developed and validated during the last years, which are able to detect the genome of the 27 BTV serotypes. Multiplex PCR in which different primer pairs, based on VP2-specific genes,87,88 were developed allowing the identification of multiple serotypes simultaneously. These techniques are the most commonly used because they provide the easiest, cheapest, and fastest as well as specific information on the presence of BTV nucleic acid in blood and tissues from suspected infected samples.82,87,89 It may be taken into account that RT-PCR detects virus-specific nucleic acid even if it is no longer viable or infectious.90
- Sequencing techniques: Classification into serogroup, serotype, and even topotype (geographical origin) could be attempted by BTV genome sequencing.73,91–93 As the full genomic sequence of BTV isolates becomes available, the design of new primers for further RT-PCR and possibly serotype-specific assays for BTV is greatly facilitated. A wide range of molecular tests based on these sequences are available, requiring however modification or scaling-up in response to the emergence of new variants or serotypes. In spite of advances in sequencing, the reduction of prices, and the increase in companies that routinely sequence, it is still an expensive technique to use as a routine diagnostic test.
Overview of current tests for antibody detection
Serological techniques for the detection of BTV serum antibodies are widely used in Animal Health Laboratories across the world for virus infection confirmation. BTV serological response develops 7–14 days after infection, generating both neutralizing and nonneutralizing anti-BTV antibodies, and they are usually considered long-lasting.94 Diagnostic tests for BTV infection can therefore be based around the detection of these anti-BTV antibodies.
Serological techniques for BTV detection can be divided into two main categories according to whether they aim at serogroup (detection of VP7 that is highly conserved within each serogroup) or serotype (neutralization techniques or detection of VP2 protein) specificity during detection. Agar gel immunodiffusion (AGID) and ELISA are the most used techniques for serogroup determination, although a wide range of different assays have been developed along the years for the same purpose. Serum neutralization is the most frequently used assay for BTV serotype identification.
Agar gel immunodiffusion
AGID95 is a variant of a classical serological reaction in which a soluble antigen in a clear medium is precipitated by the addition of the specific antibody. Three positive sera are placed in alternate wells with three test sera, while central well is filled with a crude preparation of BTV-infected BHK or Vero cells. Soluble antigens and antibodies diffuse passively through the gel toward each other, forming an insoluble immune complex seen as a precipitin line. The test is easy and quick to perform, and the antigen required is relatively easy to produce. AGID is however only considered by the OIE as an alternative test due to its disadvantages.96 Some of the disadvantages are its low sensitivity and specificity comparing with c-ELISA, the fact that is not a quantitative method and the result reading subjectivity factor.97,98 Since AGID can detect antibodies to other orbiviruses, especially those against the epizootic hemorrhagic disease, it is no longer considered a sufficiently accurate assay for international trade purposes, and AGID-positive sera results should be retested with a different serogroup-specific assay.96
Indirect ELISA (i-ELISA)
i-ELISAs are commercially available for the detection of BTV antibodies in bulk milk. VP7 is used as a substrate for sample BTV antibodies and detected by an anti-ruminant-IgG-conjugate and substrate. It has been shown99,100 to be a reliable and robust assay, with a 96.5% specificity and a 98.9% sensitivity, was useful for surveillance purposes.96 i-ELISA can also be used as a Differentiation of Infected from Vaccinated Animals (DIVA) system based on the detection of BTV NS3 antibodies.101 Recombinant NS3 was used as antigen, and unvaccinated cattle serum was compared with sera obtained after BTV infection or BTV vaccination with a bivalent inactivated BTV 2–4 vaccine. The greater levels of NS3 antibodies detected in infected animals than in vaccinated ones allow distinguishing between these two groups. This approach might be useful considering that competitive ELISA cannot differentiate infected animals from animals that received an inactivated vaccine.
Competitive ELISA
c-ELISA is a serogroup-specific assay able to measure BTV antibodies without the cross-reactivity problems of AGID.102,103 To do so, c-ELISA uses a known amount of one BT serogroup-reactive MAbs, which competes with the sample serum antibodies for the same conserved BTV VP7 antigen. This technique is a prescribed test for international trade by the OIE terrestrial manual96 and is extensively used in clinical laboratories for the detection of BTV antibodies. This test is a sensitive, specific, and reliable test compared with other aforementioned serological techniques like AGID or indirect ELISA.102,104 It is also quick and inexpensive and is considered an ideal technique to study BTV distribution, monitor vaccination campaigns, and eradicate campaign policies planning.68,96
Serum neutralization test (SNT)
This test is used for both identification and titration of BTV serotype-specific neutralizing antibodies. Due to the great variety of serotypes cocirculating around the world, being able to distinguish them with a serological test is key to surveillance. There are different SNT protocols that makes it difficult to interpret and compare results.105 Even though the OIE terrestrial manual96 published a standardized procedure after comparing different studies, conditional changes have been proposed to improve sensitivity while retaining specificity by some authors.105 SNT is based on serial dilutions of the test serum, which is incubated with a fixed amount of each BTV serotype. Cells (usually BHK or Vero) are then added and incubated for 3–5 days, after which cytopathic effect (CPE) is evaluated. Antibody titration is established by the maximum dilution capable of neutralizing >75% of CPE.96 The main advantage of SNT probably is its ability to not cross-react with other Orbivirus serogroups while distinguishing between BTV serotypes. SNT is considered a highly sensitive and specific assay106 that not only identifies the BTV serotype responsible for infection but can also be used to check the immune status of individual animals or populations after vaccination. SNT can also determine whether animals are free from infection before movement and provides essential information for eradication policy planning.96 SNT present some disadvantages too: it can be time-consuming, laborious, and requires specific reference viruses stocks for each serotype.106
Other serogroup-specific techniques
Numerous serogroup-specific assays have been developed along the years as alternative or, at least, complementary to the methods already in use.
- IgM-capture ELISA: This technique was developed for the detection of BTV antibodies in recently infected cattle.107 It is based on biotinylated capture of anti-bovine IgM antibodies bound to a streptavidin-coated ELISA plate, which can capture IgM antibodies present in test sera. These are detected by the addition of BTV VP7 antigen and an MAb specific for it.
- Immunochromatographic strips (ICS): This test uses a recombinant BTV VP7 protein immobilized on nitrocellulose membranes to test for the presence of serogroup-specific antibodies. Colloidal gold particles bound to streptococcal protein G are then used to develop the chromatographic strip.108 Its specificity and sensitivity correlated well with c-ELISA results. ICS thus appears as a rapid test that is suitable for the routine serological surveillance of BTV infection in the field, with a high specificity (97.6%) and sensitivity (100%) compared with c-ELISA.
- Protein chip detection method: In this technique,109 BTV VP7 is printed on a new solid supporting material known as polymer-coated initiator-integrated poly (dimethyl siloxane) (iPDMS). After test sera incubation, HRP-conjugated BTV-4H7 MAb is added to detect BTV anti-VP7 antibodies. Like other protein chip techniques, it allows performance of thousands of sample detections in parallel, plus a nearly zero nonspecific protein adsorption background due to iPDMS. Its sensitivity and specificity were reported as 98.6% and 94.8%, respectively.
- Latex agglutination test: This assay uses latex beads coupled with recombinant VP7 protein to detect anti-BTV antibodies in ruminant serum. Specificity and sensitivity correlated well with c-ELISA (99.0% and 93.0%, respectively). This could be ideal for the screening of large numbers of serum samples due to its quick and easy use.110
- Double-antigen microsphere immunoassay (MIA): This novel assay is both serogroup- and serotype-specific, as it was developed to detect BTV anti-VP2 and anti-VP7 serum antibodies simultaneously.111 MIA is based on color-coded florescent beads, each one coated with a different viral antigen. First, test serum and antigen-coated color beads are mixed. Then, biotinylated antibodies specific to the analyte are added to form an antibody–antigen complex which, finally, will be detected, thanks to the addition of a phycoerythrin-conjugated streptavidin and a set of lasers to detect color beads. Authors argue that VP7-MIA showed high specificity and efficiency compared with c-ELISA, while VP2-MIA was as specific as a VNT.
Despite advances in diagnostic techniques over the years, the tests used for the diagnosis that allows serogroup and serotyping are still ELISAs and VNT, respectively. These are only replaced, in the reference laboratories, by the techniques based on RT-PCRs for BTV identification as routine protocols. In France, the annual ring trials based on RT-PCR have been shown to be a good BTV surveillance method.83 The development of easy, rapid, and cheap BTV serotype identification methods is important for BTV surveillance, vaccination programs, and epidemiology studies.
Current perspectives
In spite of the efficacy of all the techniques developed for BTV diagnosis, there is still room for improvement. In fact, most current methods are either slow or present a high cost. The advantage of a quicker diagnosis to producers is that authorities will be able to rapidly introduce control measures if an outbreak emerges. Molecular techniques such as real-time RT-PCR, although fast enough to identify BTV, need isolated genetic materials and sophisticated instruments, which make these methods unsuitable for in-field analysis. Consequently, development and improvement of diagnosis assays for rapid BTV detection and identification with high sensitivity and selectivity are challenges in the field. Biosensors are attractive solutions for fast and efficient infectious disease diagnostics due to their utilization simplicity and their potential as real-time analysis tools.112 Biosensors recognize a target biomarker specific for the pathogen via an immobilized sensing element called bioreceptor that could be RNA, MAb, aptamers, or others. This bioreceptor is a crucial component that assures high sensitivity and selectivity, avoiding interferences from other pathogens. The interaction between the biomarker and the bioreceptor is converted into a measurable signal that is recorded, allowing qualitative and quantitative pathogen identification. Additional techniques to improve BTV diagnosis should also focus on the development of pen-side tests that allow for the rapid detection of target antigen or antibody, require little or no scientific equipment, and is sensitive and specific for the pathogen.
Acknowledgment
This work was funded by grantno. AGL2015-64290R from the Spanish Ministerio de Economía y Competitividad, and grant no. S2013/ABI-2906-PLATESA from the Comunidad de Madrid and the European Union (Fondo Europeo de Desarrollo Regional, FEDER funds).
Disclosure
The authors report no conflicts of interest in this work.
References
Mellor PS, Baylis M, Mertens PP. Bluetongue. London: Academic Press; 2009. | ||
Ratinier M, Caporale M, Golder M, et al. Identification and characterization of a novel non-structural protein of bluetongue virus. PLoS Pathog. 2011;7(12):e1002477. | ||
Roy P. Bluetongue virus proteins and particles and their role in virus entry, assembly, and release. Adv Virus Res. 2005;64:69–123. | ||
Jenckel M, Bréard E, Schulz C, et al. Complete coding genome sequence of putative novel bluetongue virus serotype 27. Genome Announc. 2015;3(2):e00016-15. | ||
Mertens P, Maan NS, Samuel AR, Attoui H. Orbiviruses. In: Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA, editors. Virus Taxonomy. Classification and Nomenclature of Viruses. Amsterdam, Netherlands: Elsevier Academic Press; 2005:466–483. | ||
Zientara S, Sánchez-Vizcaíno JM. Control of bluetongue in Europe. Vet Microbiol. 2013;165(1–2):33–37. | ||
Maclachlan NJ, Mayo CE. Potential strategies for control of bluetongue, a globally emerging, Culicoides-transmitted viral disease of ruminant livestock and wildlife. Antiviral Res. 2013;99(2):79–90. | ||
van der Sluijs MT, de Smit AJ, Moormann RJ. Vector independent transmission of the vector-borne bluetongue virus. Crit Rev Microbiol. 2016;42(1):57–64. | ||
Turner J, Bowers RG, Baylis M. Modelling bluetongue virus transmission between farms using animal and vector movements. Sci Rep. 2012;2(1):319. | ||
Coetzee P, Stokstad M, Venter EH, Myrmel M, van Vuuren M. Bluetongue: a historical and epidemiological perspective with the emphasis on South Africa. Virol J. 2012;9(1):198. | ||
Carpenter S, Veronesi E, Mullens B, Venter G. Vector competence of Culicoides for arboviruses: three major periods of research, their influence on current studies and future directions. Rev Sci Tech. 2015;34(1):97–112. | ||
Jacquet S, Huber K, Pagès N, et al. Range expansion of the bluetongue vector, Culicoides imicola, in continental France likely due to rare wind-transport events. Sci Rep. 2016;6(1):27247. | ||
Carpenter S, Mcarthur C, Selby R, et al. Experimental infection studies of UK Culicoides species midges with bluetongue virus serotypes 8 and 9. Vet Rec. 2008;163(20):589–592. | ||
Takken W, Van Rooij EMA, Verhlust NO. Bluetongue: an emerging vector-borne disease outbreak in north-western Europe. In: Takken W, Knols GJ, editors. Emerging pests and Vector-Borne Diseases in Europe. Wageningen: Wageningen Academic Publishers; 2007:113–121. | ||
Goffredo M, Catalani M, Federici V, et al. Vector species of Culicoides midges implicated in the 20122014 bluetongue epidemics in Italy. Vet Ital. 2015;51(2):131–138. | ||
Mordue AJ, Dallas JF, Nolan DV, Logan JG. Novel methods for the identification and control of Culicoides midges as vectors of emerging diseases. In: Takken W, Knols GJ, editors. Emerging Pests and Vector-Borne Diseases in Europe. Wageningen: Wageningen Academic Publishers; 2007:307–325. | ||
Qiu YT, Spitzen J, Smallegange RC, Knols BGJ. Monitoring systems for adult insect pests and disease vectors. In: Takken W, Knols GJ, editors. Emerging Pests and Vector-Borne Diseases in Europe. Wageningen: Wageningen Academic Publishers; 2007:329–353. | ||
González M, Alarcón-Elbal PM, Valle-Mora J, Goldarazena A. Comparison of different light sources for trapping Culicoides biting midges, mosquitoes and other dipterans. Vet Parasitol. 2016;226:44–49. | ||
Bishop AL, Bellis GA, Mckenzie HJ, et al. Light trapping of biting midges Culicoides spp. (Diptera: Ceratopogonidae) with green light-emitting diodes. Aust J Entomol. 2006;45(3):202–205. | ||
Mehlhorn H, Walldorf V, Klimpel S, et al. First occurrence of Culicoides obsoletus-transmitted bluetongue virus epidemic in central Europe. Parasitol Res. 2007;101(1):219–228. | ||
Elbers AR, Meiswinkel R, van Weezep E, Sloet van Oldruitenborgh-Oosterbaan MM, Kooi EA. Schmallenberg virus in Culicoides spp. biting midges, the Netherlands, 2011. Emerg Infect Dis. 2013;19(1):106–109. | ||
Perrin A, Cetre-Sossah C, Mathieu B, Baldet T, Delecolle JC, Albina E. Phylogenetic analysis of Culicoides species from France based on nuclear ITS1-rDNA sequences. Med Vet Entomol. 2006;20(2):219–228. | ||
Elbers AR, Meiswinkel R, van Weezep E, Kooi EA, van der Poel WH. Schmallenberg virus in Culicoides biting midges in the Netherlands in 2012. Transbound Emerg Dis. 2015;62(3):339–342. | ||
Ducheyne E, de Deken R, Bécu S, et al. Quantifying the wind dispersal of Culicoides species in Greece and Bulgaria. Geospat Health. 2007;1(2):177–189. | ||
Barratt-Boyes SM, Maclachlan NJ. Dynamics of viral spread in bluetongue virus infected calves. Vet Microbiol. 1994;40(3–4):361–371. | ||
Losson B, Mignon B, Paternostre J, et al. Biting midges overwintering in Belgium. Vet Rec. 2007;160(13):451–452. | ||
van der Sluijs MT, Schroer-Joosten DP, Fid-Fourkour A, et al. Transplacental transmission of bluetongue virus serotype 1 and serotype 8 in sheep: virological and pathological findings. PLoS One. 2013;8(12):e81429. | ||
Belbis G, Bréard E, Cordonnier N, et al. Evidence of transplacental transmission of bluetongue virus serotype 8 in goats. Vet Microbiol. 2013;166(3–4):394–404. | ||
Menzies FD, McCullough SJ, McKeown IM, et al. Evidence for transplacental and contact transmission of bluetongue virus in cattle. Vet Rec. 2008;163(7):203–209. | ||
Kirkland PD, Hawkes RA. A comparison of laboratory and “wild” strains of bluetongue virus--is there any difference and does it matter? Vet Ital. 2004;40(4):448–455. | ||
Darpel KE, Barber J, Hope A, et al. Using shared needles for subcutaneous inoculation can transmit bluetongue virus mechanically between ruminant hosts. Sci Rep. 2016;6(1):20627. | ||
Batten C, Darpel K, Henstock M, et al. Evidence for transmission of bluetongue virus serotype 26 through direct contact. PLoS One. 2014;9(5):e96049. | ||
Bréard E, Schulz C, Sailleau C, et al. Bluetongue virus serotype 27: experimental infection of goats, sheep and cattle with three BTV-27 variants reveal atypical characteristics and likely direct contact transmission BTV-27 between goats. Transbound Emerg Dis. 2018;65(2): e251–e263. | ||
van der Sluijs M, Timmermans M, Moulin V, et al. Transplacental transmission of bluetongue virus serotype 8 in ewes in early and mid gestation. Vet Microbiol. 2011;149(1–2):113–125. | ||
Coetzee P, van Vuuren M, Venter EH, Stokstad M. A review of experimental infections with bluetongue virus in the mammalian host. Virus Res. 2014;182:21–34. | ||
De Koeijer AA, Elbers ARW. Modelling of vector-borne disease and transmission of bluetongue virus in North-west Europe. In: Takken W, Knols GJ, editors. Emerging Pests and Vector-Borne Diseases in Europe. Wageningen: Wageningen Academic Publishers; 2007:99–112. | ||
Elbers AR, Backx A, Meroc E, et al. Field observations during the bluetongue serotype 8 epidemic in 2006. I. Detection of first outbreaks and clinical signs in sheep and cattle in Belgium, France and the Netherlands. Prev Vet Med. 2008;87(1–2):21–30. | ||
Katsoulos PD, Giadinis ND, Chaintoutis SC, et al. Epidemiological characteristics and clinicopathological features of bluetongue in sheep and cattle, during the 2014 BTV serotype 4 incursion in Greece. Trop Anim Health Prod. 2016;48(3):469–477. | ||
Howerth EW, Greene CE, Prestwood AK. Experimentally induced bluetongue virus infection in white-tailed deer: coagulation, clinical pathologic, and gross pathologic changes. Am J Vet Res. 1988;49(11):1906–1913. | ||
Drolet BS, Reister LM, Rigg TD, et al. Experimental infection of white-tailed deer (Odocoileus virginianus) with Northern European bluetongue virus serotype 8. Vet Microbiol. 2013;166(3–4):347–355. | ||
Alexander KA, Maclachlan NJ, Kat PW, et al. Evidence of natural bluetongue virus infection among African carnivores. Am J Trop Med Hyg. 1994;51(5):568–576. | ||
Zanella G, Martinelle L, Guyot H, Mauroy A, de Clercq K, Saegerman C. Clinical pattern characterization of cattle naturally infected by BTV-8. Transbound Emerg Dis. 2013;60(3):231–237. | ||
Howerth EW. Cytokine release and endothelial dysfunction: a perfect storm in orbivirus pathogenesis. Vet Ital. 2015;51(4):275–281. | ||
Williamson S, Woodger N, Darpel K. Differential diagnosis of bluetongue in cattle and sheep. In Practice. 2008;30(5):242–251. | ||
Brenner J, Batten C, Yadin H, et al. Clinical syndromes associated with the circulation of multiple serotypes of bluetongue virus in dairy cattle in Israel. Vet Rec. 2011;169(15):389. | ||
Watson P. Differential diagnosis of oral lesions and FMD in sheep. In Practice. 2004;26(4):182–191. | ||
Tuppurainen ESM, Oura CAL. Review: lumpy skin disease: an emerging threat to Europe, the Middle East and Asia. Transbound Emerg Dis. 2012;59(1):40–48. | ||
Rao TV, Bandyopadhyay SK. A comprehensive review of goat pox and sheep pox and their diagnosis. Anim Health Res Rev. 2000;1(2):127–136. | ||
Zientara S, Bréard E, Sailleau C. Bluetongue diagnosis by reverse transcriptase-polymerase chain reaction. Vet Ital. 2004;40(4):531–537. | ||
Palacios G, Quan P-L, Jabado OJ, et al. Panmicrobial oligonucleotide array for diagnosis of infectious diseases. Emerg Infect Dis. 2007;13(1):73–81. | ||
Wright CF, Morelli MJ, Thébaud G, et al. Beyond the consensus: dissecting within-host viral population diversity of foot-and-mouth disease virus by using next-generation genome sequencing. J Virol. 2011;85(5):2266–2275. | ||
Afshar A. Bluetongue: laboratory diagnosis. Comp Immunol Microbiol Infect Dis. 1994;17(3–4):221–242. | ||
Philpott M. The dangers of disease transmission by artificial insemination and embryo transfer. Br Vet J. 1993;149(4):339–369. | ||
Pearson JE, Ga G, Shafer AL, Alstad AD. Diagnosis of bluetongue and epizootic hemorrhagic disease. In: Thomas E, Walton and Bennie L Osburn, editors. Bluetongue, African Horse Sickness and Related Orbiviruses. Boca Raton (FL): CRC Press; 1991:533–546. | ||
Jochim MM. An overview of diagnostics for bluetongue. Prog Clin Biol Res. 1985;178:423–433. | ||
Luedke AJ, Erasmus BJ, Sawyer M, et al. WHO/FAO working team report: virology. Prog Clin Biol Res. 1985;178:665–688. | ||
Parsonson IM. Pathology and pathogenesis of bluetongue infections. Curr Top Microbiol Immunol. 1990;162:119–141. | ||
Clavijo A, Heckert RA, Dulac GC, Afshar A. Isolation and identification of bluetongue virus. J Virol Methods. 2000;87(1–2):13–23. | ||
Mcholland LE, Mecham JO. Characterization of cell lines developed from field populations of Culicoides sonorensis (Diptera: Ceratopogonidae). J Med Entomol. 2003;40(3):348–351. | ||
Wechsler SJ, McHolland LE, Tabachnick WJ. Cell lines from Culicoides variipennis (Diptera: Ceratopogonidae) support replication of bluetongue virus. J Invertebr Pathol. 1989;54(3):385–393. | ||
Blacksell SD, Lunt RA. Serotype identification of Australian bluetongue viruses using a rapid fluorescence inhibition test. J Virol Methods. 1993;44(2–3):241–250. | ||
Jochim MM, Barber TL, Bando BM. Identification of bluetongue and epizootic haemorrhagic disease viruses by the indirect fluorescent antibody procedure. Proc Ann Meeting Am Assoc Vet Lab Diagn. 1974;17:91–103. | ||
Pini A, Coackley W, Ohder H. Concentration of bluetongue virus in experimentally infected sheep and virus identification by immune fluorescence technique. Arch Gesamte Virusforsch. 1966;18(4):385–390. | ||
Wechsler SJ, Austin KJ, Wilson WC. Limits of detection of bluetongue virus with different assay systems. J Vet Diagn Invest. 1990;2(2):103–106. | ||
Cherrington JM, Hw G, Sawyer MM, Osburn BI. Detection of viral antigens in bluetongue virus-infected ovine tissues, using the peroxidase-antiperoxidase technique. Am J Vet Res. 1985;46(11):2356–2359. | ||
Anderson GA, Phillips DL, Waldvogel AS, Osburn BI. Detection of bluetongue virus in bovine fetuses using the avidin-biotin complex immunoperoxidase method. J Vet Diagn Invest. 1989;1(1):45–49. | ||
El Hussein A, Calisher CH, Holbrook FR, Schoepp RJ, Beaty BJ. Detection of bluetongue virus antigens in Culicoides variipennis by enzyme immunoassay. J Clin Microbiol. 1989;27(6):1320–1323. | ||
Hamblin C. Bluetongue virus antigen and antibody detection, and the application of laboratory diagnostic techniques. Vet Ital. 2004;40(4):538–545. | ||
Afshar A, Dubuc C, Dulac GC, Thomas FC, Nielsen K, Henning D. Dot immunoperoxidase assay using monoclonal antibody for detection of bluetongue virus antigens. J Virol Methods. 1991;31(1):105–112. | ||
Eaton BT, Hyatt AD, Brookes SM. The replication of bluetongue virus. Curr Top Microbiol Immunol. 1990;162:89–118. | ||
Gould AR, Hyatt AD, Eaton BT, et al. Current techniques in rapid bluetongue virus diagnosis. Aust Vet J. 1989;66(12):450–454. | ||
Nunamaker RA, Ellis JA, Wigington JG, Maclachlan NJ. The detection of intracellular bluetongue virus particles within ovine erythrocytes. Comp Biochem Physiol Comp Physiol. 1992;101(3):471–476. | ||
Mccoll KA, Gould AR. Detection and characterisation of bluetongue virus using the polymerase chain reaction. Virus Res. 1991;21(1):19–34. | ||
Akita GY, Chinsangaram J, Osburn BI, Ianconescu M, Kaufman R. Detection of bluetongue virus serogroup by polymerase chain reaction. J Vet Diagn Invest. 1992;4(4):400–405. | ||
Akita GY, Glenn J, Castro AE, Osburn BI. Detection of bluetongue virus in clinical samples by polymerase chain reaction. J Vet Diagn Invest. 1993;5(2):154–158. | ||
Jiménez-Clavero MA, Agüero M, San Miguel E, et al. High throughput detection of bluetongue virus by a new real-time fluorogenic reverse transcription-polymerase chain reaction: application on clinical samples from current Mediterranean outbreaks. J Vet Diagn Invest. 2006;18(1):7–17. | ||
Orrù G, Ferrando ML, Meloni M, Liciardi M, Savini G, de Santis P. Rapid detection and quantitation of bluetongue virus (BTV) using a molecular beacon fluorescent probe assay. J Virol Methods. 2006;137(1):34–42. | ||
Shaw AE, Monaghan P, Alpar HO, et al. Development and initial evaluation of a real-time RT-PCR assay to detect bluetongue virus genome segment 1. J Virol Methods. 2007;145(2):115–126. | ||
Toussaint JF, Sailleau C, Breard E, Zientara S, de Clercq K. Bluetongue virus detection by two real-time RT-qPCRs targeting two different genomic segments. J Virol Methods. 2007;140(1–2):115–123. | ||
Zientara S, Sailleau C, Bréard E, Viarouge C, Doceul V, Vitour D. Benefits of PCR and decentralization of diagnosis in regional laboratories in the management of bluetongue in France. Vet Ital. 2015;51(4):393–399. | ||
Billinis C, Koumbati M, Spyrou V, et al. Bluetongue virus diagnosis of clinical cases by a duplex reverse transcription-PCR: a comparison with conventional methods. J Virol Methods. 2001;98(1):77–89. | ||
Mertens PP, Maan NS, Prasad G, et al. Design of primers and use of RT-PCR assays for typing European bluetongue virus isolates: differentiation of field and vaccine strains. J Gen Virol. 2007;88(Pt 10):2811–2823. | ||
Sailleau C, Viarouge C, Breard E, Vitour D, Zientara S. Ring trial 2016 for bluetongue virus detection by real-time RT-PCR in France. Vet Med Sci. 2017;3(2):107–114. | ||
Hofmann MA, Renzullo S, Mader M, Chaignat V, Worwa G, Thuer B. Genetic characterization of toggenburg orbivirus, a new bluetongue virus, from goats, Switzerland. Emerg Infect Dis. 2008;14(12):1855–1861. | ||
Katz JB, Alstad AD, Gustafson GA, Moser KM. Sensitive identification of bluetongue virus serogroup by a colorimetric dual oligonucleotide sorbent assay of amplified viral nucleic acid. J Clin Microbiol. 1993;31(11):3028–3030. | ||
Shaw AE, Reid SM, Ebert K, Hutchings GH, Ferris NP, King DP. Implementation of a one-step real-time RT-PCR protocol for diagnosis of foot-and-mouth disease. J Virol Methods. 2007;143(1):81–85. | ||
Maan NS, Maan S, Belaganahalli MN, et al. Identification and differentiation of the twenty six bluetongue virus serotypes by RT-PCR amplification of the serotype-specific genome segment 2. PLoS One. 2012;7(2):e32601. | ||
Maan S, Maan NS, Belaganahalli MN, et al. Development and evaluation of real time RT-PCR assays for detection and typing of bluetongue virus. PLoS One. 2016;11(9):e0163014. | ||
Johnson DJ, Wilson WC, Paul PS. Validation of a reverse transcriptase multiplex PCR test for the serotype determination of U.S. isolates of bluetongue virus. Vet Microbiol. 2000;76(2):105–115. | ||
MacLachlan NJ. The pathogenesis and immunology of bluetongue virus infection of ruminants. Comp Immunol Microbiol Infect Dis. 1994;17(3–4):197–206. | ||
Gould AR. The complete nucleotide sequence of bluetongue virus serotype 1 RNA3 and a comparison with other geographic serotypes from Australia, South Africa and the United States of America, and with other orbivirus isolates. Virus Res. 1987;7(2):169–183. | ||
Pritchard LI, Gould AR, Wilson WC, Thompson L, Mertens PP, Wade-Evans AM. Complete nucleotide sequence of RNA segment 3 of bluetongue virus serotype 2 (Ona-A). phylogenetic analyses reveal the probable origin and relationship with other orbiviruses. Virus Res. 1995;35(3):247–261. | ||
Wilson WC, Ma HC, Venter EH, van Djik AA, Seal BS, Mecham JO. Phylogenetic relationships of bluetongue viruses based on gene S7. Virus Res. 2000;67(2):141–151. | ||
Stott JL, Barber TL, Osburn BI. Immunologic response of sheep to inactivated and virulent bluetongue virus. Am J Vet Res. 1985;46(5):1043–1049. | ||
Pearson JE, Jochim MM. Protocol for the immunodiffusion test for bluetongue. Proc Am Assoc Vet Lab Diagn. 1979;22:463–471. | ||
(OIE) OIdÉ. Chapter 2.1.3. Bluetongue (infection with bluetongue virus). In: Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Paris: OIE: World Organisation for Animal Health; 2018. | ||
Della-Porta AJ, Parsonson IM, McPhee DA. Problems in the interpretation of diagnostic tests due to cross-reactions between orbiviruses and broad serological responses in animals. Prog Clin Biol Res. 1985;178:445–453. | ||
Reddington JJ, Reddington GM, Maclachlan NJ. A competitive ELISA for detection of antibodies to the group antigen of bluetongue virus. J Vet Diagn Invest. 1991;3(2):144–147. | ||
Kramps JA, van Maanen K, Mars MH, Popma JK, van Rijn PA. Validation of a commercial ELISA for the detection of bluetongue virus (BTV)-specific antibodies in individual milk samples of Dutch dairy cows. Vet Microbiol. 2008;130(1–2):80–87. | ||
Mars MH, van Maanen C, Vellema P, Kramps JA, van Rijn PA. Evaluation of an indirect ELISA for detection of antibodies in bulk milk against bluetongue virus infections in the Netherlands. Vet Microbiol. 2010;146(3–4):209–214. | ||
Barros SC, Cruz B, Luís TM, et al. A Diva system based on the detection of antibodies to non-structural protein 3 (NS3) of bluetongue virus. Vet Microbiol. 2009;137(3–4):252–259. | ||
Afshar A, Thomas FC, Wright PF, Shapiro JL, Anderson J. Comparison of competitive ELISA, indirect ELISA and standard AGID tests for detecting blue-tongue virus antibodies in cattle and sheep. Vet Rec. 1989;124(6):136–141. | ||
Lunt RA, White JR, Blacksell SD. Evaluation of a monoclonal antibody blocking ELISA for the detection of group-specific antibodies to bluetongue virus in experimental and field sera. J Gen Virol. 1988;69(Pt 11):2729–2740. | ||
Afshar A, Thomas FC, Wright PF, Shapiro JL, Shettigara PT, Anderson J. Comparison of competitive and indirect enzyme-linked immunosorbent assays for detection of bluetongue virus antibodies in serum and whole blood. J Clin Microbiol. 1987;25(9):1705–1710. | ||
Worwa G, Chaignat V, Feldmann J, Thür B. Detection of neutralizing antibodies against bluetongue virus serotype 8 by an optimized plasma neutralization test. J Virol Methods. 2013;188(1–2):168–174. | ||
Chand K, Biswas SK, Pandey AB, Saxena A, Tewari N, Mondal B. A competitive ELISA for detection of group specific antibody to bluetongue virus using anti-core antibody. Biologicals. 2017;46:168–171. | ||
Zhou EM, Ridd D, Riva J, Fernando L, Clavijo A. Development and evaluation of an IgM-capture ELISA for detection of recent infection with bluetongue viruses in cattle. J Virol Methods. 2001;91(2):175–182. | ||
Yang J, Hua Q, Chen H, et al. Development and evaluation of an immunochromatographic strip for the detection of serum antibodies against bluetongue virus. J Virol Methods. 2010;163(1):68–73. | ||
Xu QY, Sun EC, Feng YF, et al. Development of a novel protein CHIP for the detection of bluetongue virus in China. J Virol Methods. 2016;234:28–33. | ||
Yang J, Hua Q, Chen H, Lv J, Chen B, Ruan Z. A rapid assay for detecting antibody against bluetongue virus with a latex agglutination test using recombinant VP7 antigen. J Vet Diagn Invest. 2010;22(2):242–244. | ||
Breard E, Garnier A, Despres P, et al. Development of a double-antigen microsphere immunoassay for simultaneous group and serotype detection of bluetongue virus antibodies. Transbound Emerg Dis. 2017;64(6):1837–1847. | ||
Yoo SM, Lee SY. Optical biosensors for the detection of pathogenic microorganisms. Trends Biotechnol. 2016;34(1):7–25. |
 © 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2019 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.