Back to Journals » Nature and Science of Sleep » Volume 17

Dexmedetomidine Alleviates Hippocampal Tissue Damage in Rapid Eye Movement Sleep-Deprived Rats by Activating BDNF/TrkB Signaling Pathway
Received 28 May 2025
Accepted for publication 16 September 2025
Published 1 October 2025 Volume 2025:17 Pages 2423—2435
DOI https://doi.org/10.2147/NSS.S539185
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Sarah L Appleton
Biqiong Zheng,1,2,* Yang Li,3,* Changyi Liu4,5
1Department of Anesthesiology, The First Affiliated Hospital of Fujian Medical University, Fuzhou, 350005, People’s Republic of China; 2Department of Anesthesiology, National Regional Medical Center, Binhai Campus of The First Affiliated Hospital, Fujian Medical University, Fuzhou, 350212, People’s Republic of China; 3Department of Anesthesiology, The First Affiliated Hospital of Xiamen University, School of Medicine, Xiamen University, Xiamen, Fujian Province, 361001, People’s Republic of China; 4Department of Urology, The First Affiliated Hospital of Fujian Medical University, Fuzhou, 350005, People’s Republic of China; 5Department of Urology, National Regional Medical Center, Binhai Campus of The First Affiliated Hospital, Fujian Medical University, Fuzhou, 350212, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Changyi Liu, Department of Urology, The First Affiliated Hospital of Fujian Medical University, Fuzhou, No. 20, Cha Zhong Road, Fuzhou, Fujian Province, People’s Republic of China, Email [email protected]
Introduction: Sleep deprivation often leads to marked neurobehavioral and cognitive deficits, yet few well-defined interventions exist to address these effects. Dexmedetomidine (DEX), a highly selective α2-adrenoceptor agonist, possesses sedative, hypnotic, analgesic, and sympathetic-blocking properties, closely mimic natural sleep state. In this study, we aim to investigate whether DEX protects hippocampal tissue against rapid eye movement sleep deprivation (RSD)-induced injury in rats and to explore the underlying molecular mechanisms.
Methods: In this study, a rapid eye movement sleep deprivation (RSD) rat model was created using a modified multi-platform method. The influence of dexmedetomidine (DEX) on hippocampal tissue morphology, the BDNF/TrkB signaling pathway, and cognitive function was then evaluated. Group comparisons were analyzed using one-way ANOVA followed by appropriate post hoc tests.
Results: In comparison with the control group, DEX significantly alleviated the impaired spatial learning and memory as reflected escape latency and increased the time spent in the garget quadrant. ANA-12 reversed these improvements, indicating DEX’s cognitive benefits. HE staining showed that DEX protected neurons from RSD-induced injury by preserving structural integrity and TUNEL assay demonstrated reduced neuron apoptosis in the DEX group. Co-treatment with ANA-12 abolished these protective effects, resulting in neuronal damage and apoptosis levels similar to those observed in RSD rats. Moreover, compared with the level of TNA alpha in RSD rats, IL 6, IL 1beta and MDA levels were lower in the hippocampus of DEX group, while SOD activity was enhanced. Western blot analysis revealed that DEX increased hippocampal BDNF (0.586 ± 0.036 vs 0.315 ± 0.034, ~1.86‑fold, P < 0.01), TrkB (0.774 ± 0.039 vs 0.518 ± 0.033, ~1.49‑fold, P < 0.01) and pro-TrkB expression. However, co-administration of ANA‑12 abolished these effects, returning expression levels close to those in the RSD group, implying that DEX’s neuroprotection is mediated via the BDNF/TrkB pathway.
Conclusion: These findings indicate that DEX exerts neuroprotective effects in RSD by activating the BDNF/TrkB pathway, offering valuable evidence for DEX-based therapeutic approaches to sleep deprivation-related brain injury.
Keywords: dexmedetomidine, rapid eye movement sleep deprivation, hippocampus, BDNF, TrkB, neuroprotection
Introduction
Sleep deprivation constitutes a prevalent health concern in modern society. Epidemiological studies estimate that approximately 35–40% of adults experience insufficient sleep, and nearly 20% suffer from significant sleep-related cognitive impairments.1 Among various forms of sleep loss, rapid eye movement sleep deprivation (RSD) particularly affects learning and memory, leading to hippocampal dysfunction and cognitive decline. RSD has been shown to impair synaptic plasticity, increase neuroinflammation, and downregulate brain-derived neurotrophic factor (BDNF) and TrkB expression, thereby contributing to hippocampal dysfunction and cognitive decline.2 The hippocampus, a critical region for learning and memory, is highly susceptible to the adverse effects of insufficient sleep.3
Dexmedetomidine (DEX), a selective α2-adrenergic agonist, exhibits sedative, analgesic, and neuroprotective properties.4 Emerging evidence indicates that DEX exerts hippocampal neuroprotection through multiple mechanisms. For example, DEX attenuates perioperative neurocognitive disorders by suppressing hippocampal neuroinflammation via the HMGB1/RAGE/NF-κB pathway, and reduces hypoxia-induced synaptic loss by inhibiting microglial NOX2 activation and oxidative stress.5,6 In addition, DEX alleviates LPS-induced cognitive dysfunction by modulating Nrf2 signaling, thereby reducing proinflammatory cytokines, enhancing antioxidant capacity, and promoting synaptic integrity.7 Notably, recent findings suggest that DEX may confer neuroprotection by modulating neurotrophic factor signaling pathways.8 Among various neuroprotective mechanisms, the brain-derived neurotrophic factor (BDNF) and it receptor tropomyosin receptor kinase B (TrkB) are particularly critical for neuronal survival, synaptic plasticity, and cognitive performance.9,10 The transcription factor cAMP response element-binding protein (CREB) serves as a key downstream effector of BDNF/TrkB signaling. Activation of TrkB stimulates CREB phosphorylation, which subsequently enhances BDNF transcription, forming a positive feedback loop essential for maintaining synaptic plasticity, learning, and memory.11 Activation of the BDNF/TrkB signaling cascade has been widely recognized as a promising neuroprotective mechanism.12,13 Recent studies further suggest that DEX can promote hippocampal neurogenesis and improve cognitive outcomes by activating the BDNF/TrkB/CREB pathway and mitigate hippocampal neuronal damage in epilepsy by downregulating MeCP2 to restore BDNF/TrkB activity.3,14 However, despite extensive evidence supporting DEX-mediated neuroprotection, whether DEX mitigates RSD-induced hippocampal damage and cognitive deficits via modulation of the BDNF/TrkB pathway remains unknown.
Therefore, the present study investigated the protective effects of DEX on RSD-induced hippocampal injury in rats and evaluated its potential involvement in the BDNF/TrkB pathway, aiming to provide a theoretical foundation for developing novel neuroprotective strategies.
Materials and Methods
Experimental Animals
A total of eighty healthy Specific Pathogen Free (SPF)-grade male Sprague-Dawley rats (Hangzhou Medical College Laboratory Animal Center) of body weights180-200 g were selected for this experiment [Licence NO. SCXK (Zhe) 2019–0002]. The number of animals per group was determined based on previous studies and to ensure sufficient statistical power for detecting intergroup differences. Additionally, considering that several investigators were involved in parallel experiments, a larger sample size was adopted to ensure data reliability. Rats were fed and watered ad libitum in polypropylene cages (4 rats/cage), with a 12h light-dark cycle, temperature of 22°C–25°C, and relative humidity of 45%–55%. All mice were allowed to acclimatize to the laboratory environment for 14 days before the start of the experiment under standard housing conditions. All behavioral experiments were conducted at a fixed time using a blinded method. All procedures were performed in accordance with the regulations of the China Animal Care Committee and approved by the Animal Ethics Committee of Fujian Medical University (IACUCFJMU 2023-Y-0768). The study was designed, conducted, and reported in compliance with the ARRIVE guidelines.
Grouping
Eighty SPF-grade healthy male SD rats were randomly assigned, using a numeric table method, into five groups: control, RSD, RSD+DEX (RSD treated with dexmedetomidine), DEX, and RSD+DEX+ANA-12 (an inhibitor of the BDNF receptor TrkB), with 16 rats in each group.
Modified Multi-Platform Method to Establish RSD Rat Models
The RSD, RSD+DEX, and RSD+DEX+ANA-12 groups underwent the modified multi-platform method (MMPM).15 Following established protocols, the rats were placed in 12 specialized RSD cages with free access to water and food.3 During REM sleep, low muscle tone causes the rats to fall into the water, prompting them to climb back onto the platform and remain awake. This procedure effectively deprived them of REM sleep for 96 hours. In contrast, the DEX and control groups were acclimated according to the large-platform water environment method,3 allowing the animals to feed, drink, move, and sleep at will on a larger platform. All rats were acclimated in a water tank for 2 hours per day over seven days, and the tank was cleaned and refilled daily.
DEX Intervention
In the RSD+DEX, DEX, and RSD+DEX+ANA-12 groups, rats received daily intraperitoneal injections of dexmedetomidine hydrochloride (25 μg/kg; H20090248, Jiangsu Hengrui, China) beginning 7 days before REM sleep deprivation and continuing until the conclusion of the Morris Water Maze experiment (Figure 1A).16 In contrast, the RSD and control groups were administered comparable volumes of saline using the same schedule.
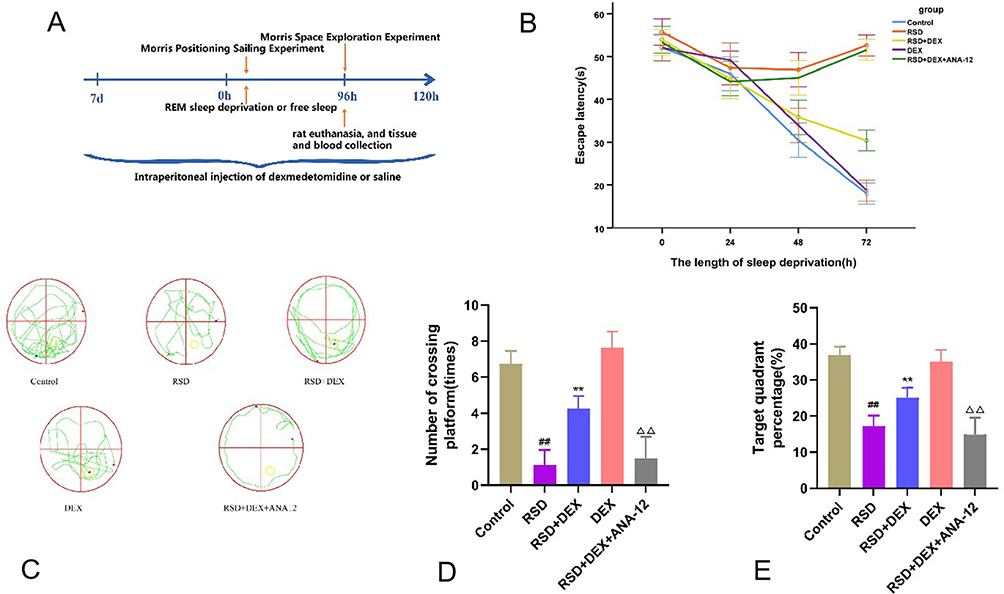 |
Figure 1 DEX improved the spatial learning ability of rats impaired by RSD, while ANA-12 inhibited the protective effect of dexmedetomidine. (A) Timeline for sleep deprivation, drug administration, behavioral testing, rat euthanasia, and tissue and blood collection. (B–E) Behavioral experiments included escape latency, exploration trajectory, number of crossing platform, and percentage time in target quadrant to assess the spatial learning memory of RSD rats. Results were expressed as mean ± standard deviation (n = 8). ##p<0.01 for the control group; **p <0.01 for the RSD group; ΔΔp<0.01 for the RSD+DEX group. |
Morris Water Maze Localization Navigation Experiment and Exploration Training
In the Morris Water Maze (LY-WMS, Xinruan Technology, China) localization and navigation task, each rat underwent four daily training sessions (twice in the morning and twice in the afternoon), with each session lasting 60 seconds. Animals were introduced into the pool from four distinct entry points, facing the pool wall, and allowed to remain on the platform for 10 seconds once located. The time elapsed from entry until the platform was discovered was recorded as the escape latency, reflecting spatial learning ability. At the end of sleep deprivation, the platform was removed, and a spatial search trial was conducted by placing the rats in the diagonal quadrant of the original platform quadrant. The cumulative percentage of time spent in the former platform quadrant and the number of platform crossings were documented, thereby assessing spatial memory.
Hematoxylin-Eosin Staining
Following the Morris Water Maze spatial exploration test, eight rats per group were selected using a random number table and subsequently anesthetized with 2% sodium pentobarbital (0.3 mL/100 g). The heart was exposed via thoracotomy, and 500 mL of sterile saline was quickly infused through the left ventricle. This was immediately followed by perfusion with 250 mL of ice-cold (4°C) 4% paraformaldehyde. Afterward, the rats were decapitated, and hippocampal tissue was harvested. Samples underwent a ethanol series, cleared in xylene, embedded in paraffin. Coronal sections of the hippocampus (5 μm thick) were cut and stained with hematoxylin (KGE1204-50, Jiangsu KGI, China) for 40 seconds, washed in running water, then stained with eosin for 1 minute. After ethanol dehydration and xylene washing, hematoxylin-eosin (HE)-stained sections were examined under a microscope (ECLIPSE E100, Nikon). The 20 μm and 50 μm scale bars were automatically generated by the microscope’s built-in calibration software based on the objective magnification.
TUNEL Apoptosis Assay
Coronal sections of the hippocampus were prepared and subjected to TUNEL and DAB staining according to the instructions of the TUNEL assay kit (KGA1400-100, Jiangsu KGI, China). After staining, neuronal cells in six randomly selected microscopic fields were photographed. The total number of neurons and the number of TUNEL-positive (apoptotic) neurons were quantified using ImageJ software. The apoptosis rate was then calculated as: Apoptosis rate (%) = (Number of apoptotic neurons / Total number of neurons) × 100%.
Enzyme Linked Immunosorbent Assay
Supernatants from fresh hippocampal tissues were collected and analyzed according to each ELISA kit’s protocol. Optical density (OD) was measured at 450 nm on a SpectraMax 190 microplate reader (Molecular Devices, USA). A polynomial quadratic regression equation was established based on the standard curve, and OD values were plugged back into this equation to determine tumor necrosis factor α (TNF-α), interleukin 6 (IL-6), interleukin 1β (IL-1β), malondialdehyde (MDA), and superoxide dismutase (SOD) activity levels. Kits for TNF-α, IL-6, and SOD were purchased from China Wuhan Cloud-Clone Corporation, whereas IL-1β and MDA kits were sourced from abcam (ab255730) and Jiangsu KGI (KGA7101-50), respectively.
Western Blot Assay
Eight additional rats in each group were anesthetized as before, and their hippocampi were carefully dissected to avoid contamination from adjacent brain regions. Each fresh hippocampus was immediately weighed using a precision balance (±0.1 mg), and then homogenized in a pre-chilled glass homogenizer on ice. For protein extraction, RIPA lysis buffer (Thermo Scientific, #89900, USA) supplemented with protease and phosphatase inhibitors was added proportionally to tissue weight, following the manufacturer’s recommendations. The homogenates were centrifuged at 5400 r/min for 10 minutes at 4 °C. The supernatant was collected, and protein concentration was measured with a BCA kit (P0010S, Beyotime, China). After adding 5× protein loading buffer (Laemmli buffer), samples were denatured in boiling water at 100°C for 10 minutes. SDS-PAGE was carried out using 10% resolving gels and a 5% stacking gel (P0012A, Beyotime, China). Samples were loaded, electrophoresed (DYCZ-26B, Beijing Liuyi Biotechnology, China), and then transferred onto PVDF membranes (IPVH00010, ISEQ00010, Millipore). Membranes were blocked with 5% skim milk for 1 hour, followed by overnight incubation at 4°C with primary antibodies against BDNF (1:1,500; GB11559, Servicebio), CREB (1:1,500; A11989, ABclonal), and TrkB (1:1,000; A2099, ABclonal). For TrkB detection, a rabbit polyclonal antibody against TrkB (ABclonal, Cat. No. A2099) was used. According to the manufacturer’s datasheet and our experimental results, the antibody primarily recognizes the full-length TrkB isoform (~140 kDa). After three 10-minute washes with TBST, membranes were incubated for 1 hour at room temperature with PBS-diluted secondary antibody (Goat Anti-Rabbit Antibody Conjugated to Horseradish Peroxidase, 1:1000, Bio-rad, Cat. No. 1662408EDU), then washed with TBST three times for 10 minutes each wash. Protein bands were visualized by enhanced chemiluminescence (ECL), using a GelDoc XR+ imaging system (Bio-Rad, USA) for image acquisition, and the band gray values were quantified with ImageJ software (NIH, USA).
Statistical Analysis
Statistical analyses were performed using SPSS (version 25.0). First, normality was assessed using the Shapiro–Wilk test, and variance homogeneity was evaluated using Levene’s test. Continuous variables with normal distributions and equal variances were expressed as mean ± standard deviation (SD) and analyzed by one-way analysis of variance (ANOVA), followed by the Least Significant Difference (LSD) post hoc test for multiple pairwise comparisons. When the assumption of homogeneity of variances was violated, Dunnett’s T3 test was employed instead. For non-normally distributed data, the Kruskal–Wallis test followed by Dunn’s post hoc test was applied. Effect sizes (η²) were calculated to evaluate the magnitude of differences where appropriate. Statistical significance was set at P < 0.05.
Results
DEX Alleviates Impairment of Spatial Learning Memory Capacity in REM Sleep-Deprived Rats and ANA-12 Inhibits the Protective Effect of DEX
In the Morris Water Maze localization navigation test, REM sleep-deprived (RSD) rats showed notably prolonged escape latencies compared to the control group, signifying impaired spatial learning (P<0.01) (Table 1). However, dexmedetomidine (DEX) administration in RSD rats shortened escape latencies, reflecting a protective effect against RSD-induced deficits (P<0.01). When ANA-12 (a TrkB inhibitor) was added, the DEX-mediated improvement disappeared, and escape latencies again increased (P<0.01). There was no difference between the DEX-alone group and control group (P>0.05). Additionally, there was no significant difference between the RSD group and RSD+DEX+ANA-12 group (P>0.05), suggesting that DEX’s benefits were dependent on TrkB signaling (Figure 1B).
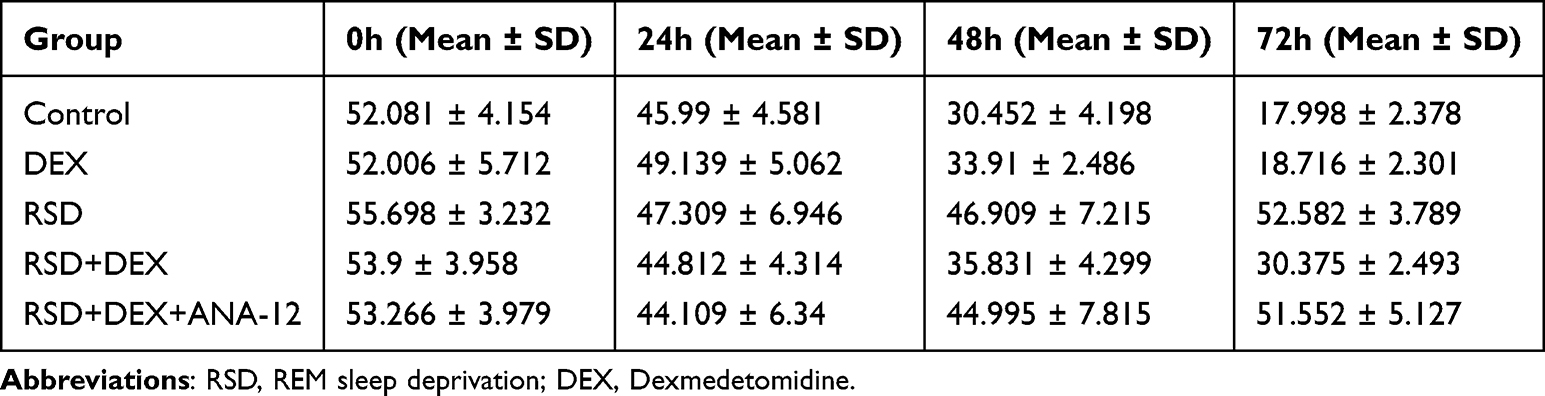 |
Table 1 Escape Latency Summary |
In the Morris Water Maze, RSD rats displayed fewer platform crossings and spent less time in the target quadrant compared to controls (P<0.01) (Table 2). DEX treatment significantly reversed these deficits (P<0.01). However, co-administration of ANA-12, a TrkB inhibitor, nullified DEX’ s protective effects (P<0.01). No significant differences were observed between the RSD group and RSD+DEX+ANA-12 group, or between the DEX group and control group (P>0.05), indicating that the DEX benefit depends on TrkB signaling (Figure 1C–E).
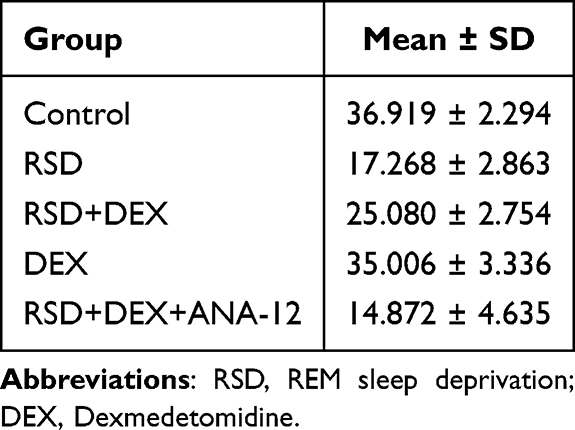 |
Table 2 Target Quadrant Percentage (%) Summary |
DEX Alleviates Cell Damage in CA1 Area of Hippocampal Tissue in RSD Rats and ANA-12 Inhibits the Protective Effect of DEX
HE staining (Figure 2) revealed that RSD caused substantial neuronal damage in the CA1 region, as shown by disorganized pyramidal cell arrangement, nuclear condensation, and widened cellular gaps compared with controls. DEX treatment in RSD rats markedly improved neuronal morphology, indicating reduced tissue damage. However, adding ANA-12 largely reversed DEX’s protective effect, once again displaying pronounced CA1 neuronal damage, comparable to that seen in the RSD group.
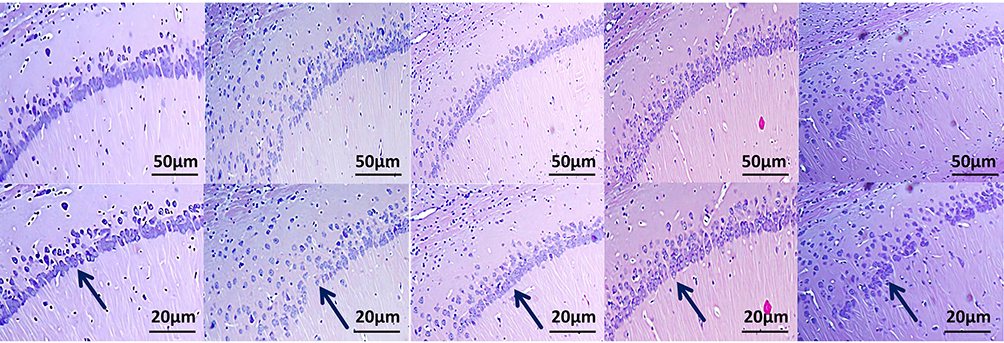 |
Figure 2 The effect of DEX on the histopathological changes in rat hippocampal neurons induced by RSD. HE-stained cells in the CA1 region of the hippocampus, scale bars: 50 μm and 20 μm. Blue arrows indicate the number and arrangement of neuronal cells in the CA1 region of the hippocampus. Results are expressed as mean ± standard deviation (n = 3). |
DEX Alleviates the Rate of Apoptosis in Hippocampal RSD Rats and ANA-12 Inhibits the Protective Effect of DEX
TUNEL staining (Figure 3) showed a marked increase in TUNEL-positive nuclei in the CA1 pyramidal neurons of the RSD group compared with controls (P<0.01), indicating higher levels of apoptosis (Table 3). These apoptotic cells displayed shrunken bodies and condensed or peripherally located nuclei. In contrast, the RSD+DEX group exhibited a significant reduction in TUNEL-positive cells compared with the RSD group (P<0.01), suggesting a protective effect of DEX. However, co-administration of ANA-12 reversed this protection, with TUNEL-positive cells in the RSD+DEX+ANA-12 group increasing to levels comparable to the RSD group (P>0.05).
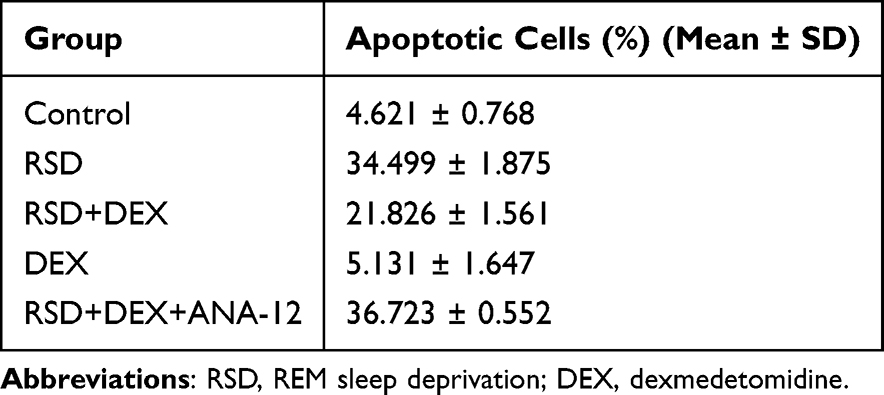 |
Table 3 Apoptotic Proportion |
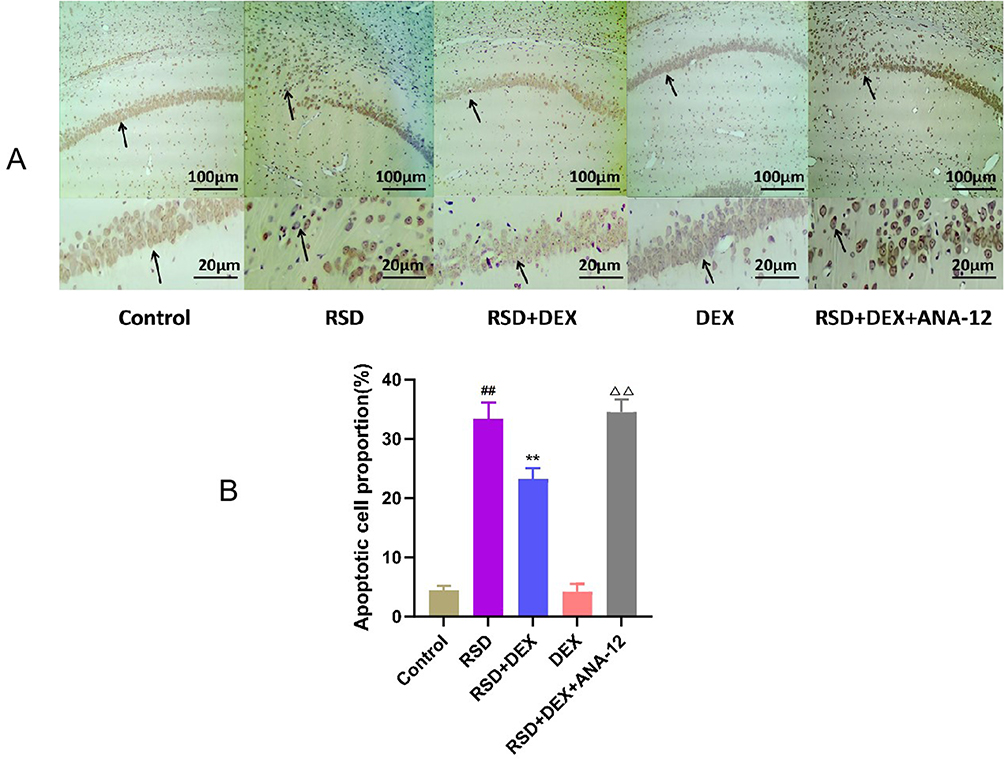 |
Figure 3 The protective effect of DEX on RSD-induced hippocampal neurons in rats. (A) Representative images of TUNEL staining in the hippocampal CA1 region of each group. Black arrows indicate TUNEL-positive apoptotic cells, characterized by shrunken cell bodies and condensed or peripherally located nuclei, scale bars: 50 μm and 20 μm; (B) Proportion of TUNEL-positive apoptotic cells in the CA1 region of the hippocampus. Results are expressed as mean ± standard deviation (n = 3). ##p<0.01 for the control group; **p <0.01 for the RSD group; ΔΔp<0.01 for the RSD+DEX group. |
DEX Increases SOD Activity and Decreases Expression of TNF-α, IL-6, IL-1β, and MDA Content in Hippocampal Tissues of RSD Rats, While ANA-12 Inhibits the Effects of DEX
ELISA (Figure 4) revealed that, compared to controls, RSD significantly decreased hippocampal SOD activity (P<0.01), while levels of TNF-α, IL-6, IL-1β, and MDA were significantly elevated (P<0.01) (Table 4). In the RSD+DEX group, SOD activity was restored (P<0.01) and TNF-α, IL-6, IL-1β, and MDA levels were all markedly reduced (P<0.01), indicating that DEX mitigates oxidative stress and inflammation induced by RSD. Compared to RSD+DEX, the RSD+DEX+ANA-12 group showed significantly lower hippocampal SOD activity and significantly higher TNF-α, IL-6, IL-1β, and MDA levels (P<0.01). Meanwhile, there were no significant differences in these parameters between the RSD and RSD+DEX+ANA-12 groups (P>0.05), suggesting that ANA-12 negates the protective effect of DEX.
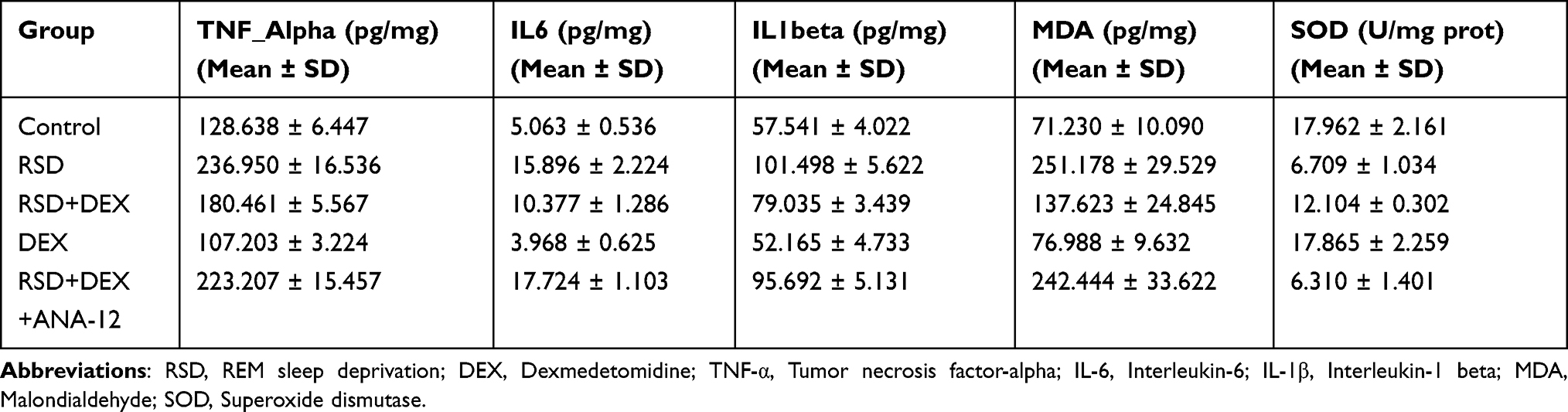 |
Table 4 Levels of Inflammatory Cytokines and Oxidative Stress Markers Measured by ELISA |
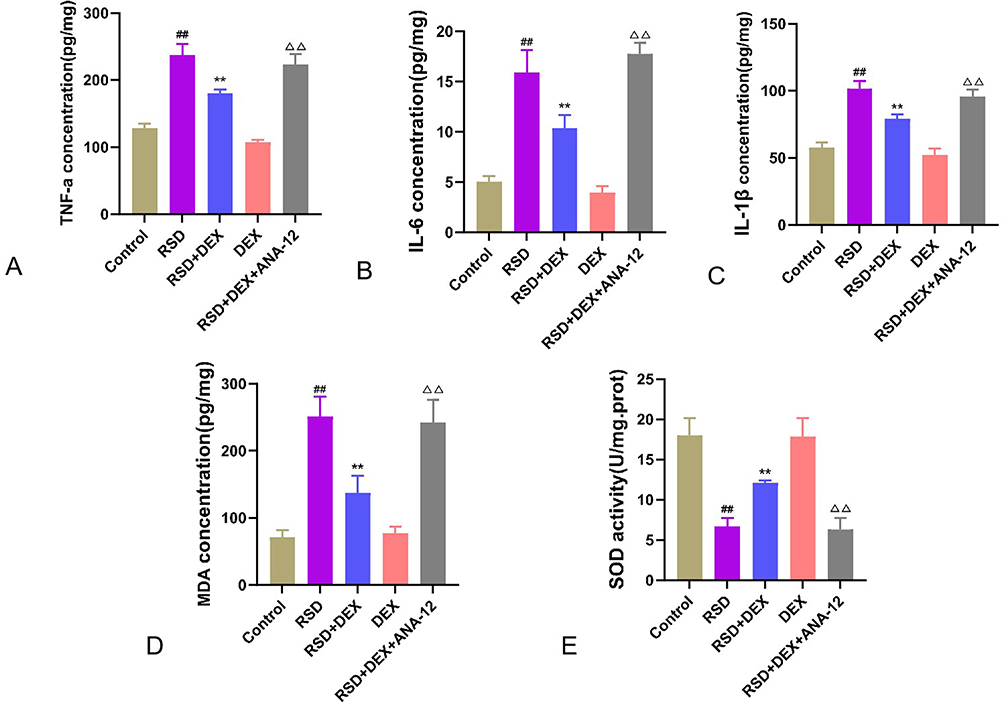 |
Figure 4 The protective effect of DEX on RSD-induced hippocampal neurons in rats. (A–D) ELISA detection of TNF-α (A), IL-6 (B), IL-1β (C), MDA (D) expression levels, and SOD (E) activity in hippocampal tissue. Results are expressed as mean ± standard deviation (n = 3). ##p<0.01 for the control group; **p <0.01 for the RSD group; ΔΔp<0.01 for the RSD+DEX group. |
DEX Increases BDNF, TrkB, and CREB Protein Expression in Hippocampal Tissues of RSD Rats While ANA-12 Inhibits Its Effects
Western blot analysis (Figure 5) revealed that RSD significantly downregulated the protein expression of BDNF, TrkB, and CREB in the hippocampus compared to control rats (P<0.01) (Table 5). Dexmedetomidine (DEX) treatment countered this reduction and significantly increased these protein levels (P<0.01), suggesting activation of the BDNF/TrkB pathway. However, co-administration of the TrkB inhibitor ANA-12 suppressed these increases (P<0.01), indicating that DEX’s neuroprotective effect relies on TrkB signaling.
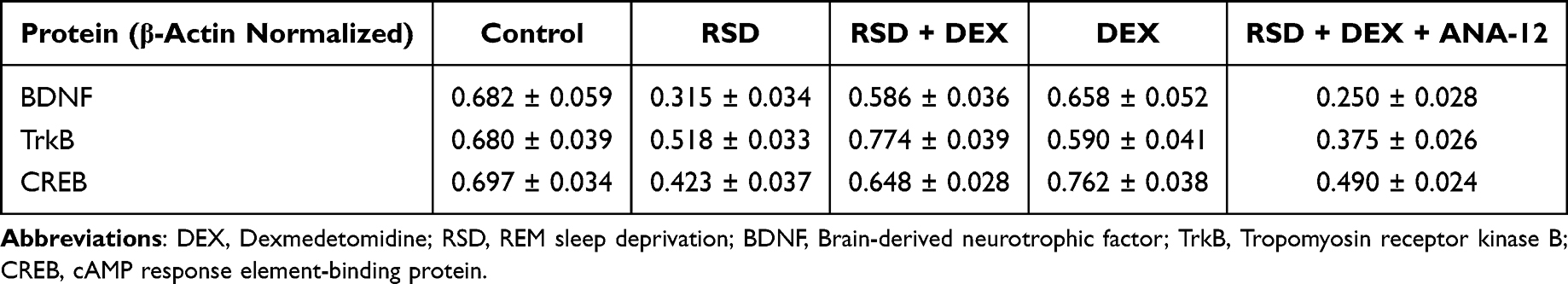 |
Table 5 Quantification of BDNF, TrkB, and CREB Protein Expression Normalized to β-Actin (Mean ± SD) |
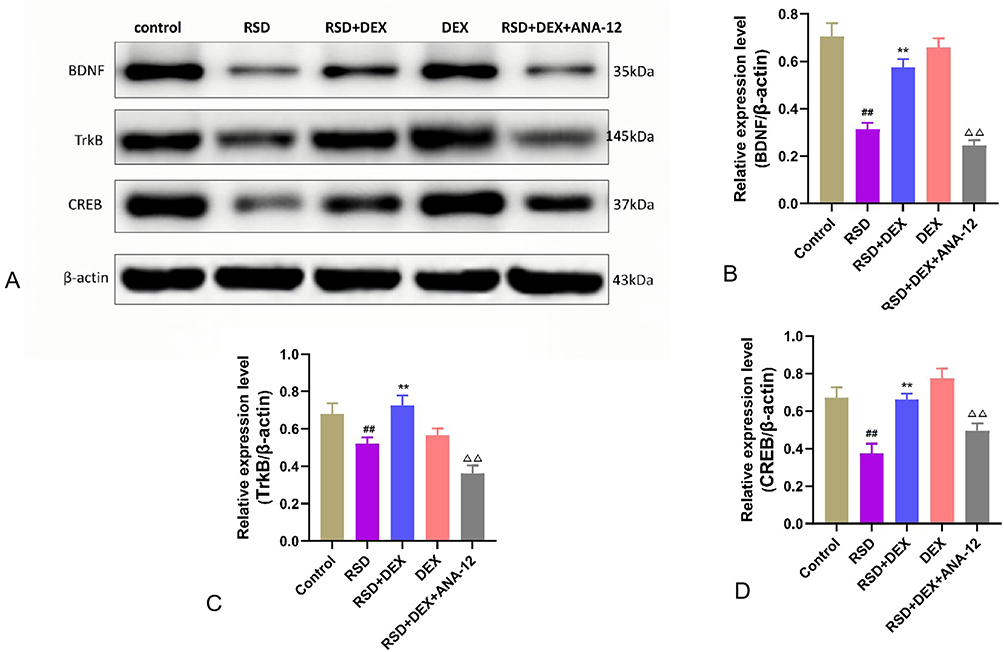 |
Figure 5 The effect of DEX on the BDNF/TrkB/CREB signaling pathway in the hippocampus of RSD-induced rats. (A) Western blot analysis of BDNF, TrkB, and CREB expression in the hippocampus; (B–D) Semi-quantitative analysis of BDNF, TrkB and CREB protein expression. Data are expressed as mean ± standard deviation (n = 3). Compared to the control group, ##p <0.01; compared to the RSD group, **p <0.01; compared to the RSD+DEX group, ΔΔp <0.01. |
Discussion
Sleep is divided into REM sleep and non-REM (NREM) sleep, alternating between the two types, where REM sleep plays a crucial role in learning memory function.17,18 The modified multiplatform aquatic environment method of sleep deprivation involves rats falling into water due to a decrease in generalized muscle tone when the rat enters the REM sleep stage.19 In this study, the spatial learning memory ability of RSD rats was impaired. HE staining showed disorganized cell arrangement, nuclear consolidation, and widening of cell gaps, and the rate of TUNEL-positive cells was increased, indicating that the rat RSD model established in this experiment was successful.
We specifically focused on the hippocampal CA1 region because this subfield is well recognized to be highly vulnerable to stress-related injury, including REM sleep deprivation. CA1 pyramidal neurons have higher metabolic demand, denser glutamatergic synaptic input, and greater NMDA receptor expression compared with CA3 or the dentate gyrus, making them more susceptible to excitotoxic and oxidative stress-induced damage. In contrast, CA3 neurons and the dentate gyrus are relatively resistant due to stronger inhibitory GABAergic networks and greater neurogenic capacity, resulting in less pronounced neuronal loss in these regions under similar conditions.20
DEX promotes NREM sleep and induces restorative sleep, boosts sleep efficiency, and benefits overall sleep patterns.21,22 It also helps mitigate depression-like behaviors resulting from sleep deprivation.23 In the present study, DEX intervention in RSD rats improved learning and memory function and reduced hippocampal cell damage and apoptosis. However, administering ANA-12, a selective TrkB inhibitor, negated these benefits, indicating that DEX’s protective effects in RSD likely depend on BDNF/TrkB signaling. α2-adrenergic agonists such as dexmedetomidine (DEX) have been widely reported to exert neuroprotective effects through multiple mechanisms. By activating presynaptic α2-adrenoceptors, DEX inhibits norepinephrine release and suppresses excessive sympathetic outflow, thereby reducing excitotoxicity and energy demand in vulnerable neuronal populations.24 DEX also decreases neuroinflammation by inhibiting microglial activation and lowering the production of proinflammatory cytokines such as TNF-α, IL-6, and IL-1β.25 Furthermore, DEX has been shown to attenuate oxidative stress by enhancing antioxidant enzyme activity and reducing lipid peroxidation.26 Consistent with these reports, our results demonstrated that DEX reduced hippocampal neuronal apoptosis, suppressed inflammatory cytokine expression, and restored antioxidant enzyme levels in REM sleep-deprived rats. Similar neuroprotective effects of DEX have been observed in ischemic brain injury, traumatic brain injury, and hypoxia-induced cognitive dysfunction, supporting the translational relevance of our findings.27–29
Following brain injury, BDNF dampens the inflammatory response and fosters an environment conducive to neural regeneration, predominantly through its modulation of immune cell activity and inflammatory factor release.30–32 However, an elevated release of inflammatory factors that are neurotoxic can coincide with downregulation of BDNF.33 In this study, RSD rats displayed a pronounced neuroinflammatory response, evidenced by increased levels of IL-6, IL-1α, and TNF-α. DEX reduced these inflammatory markers in the hippocampus of RSD rats, whereas co-administration of ANA-12 blocked DEX’s anti-inflammatory effect, indicating that BDNF/TrkB signaling plays a pivotal role in limiting neuroinflammation under RSD conditions. Previous studies indicate that oxidative stress-induced neuronal injury is primarily related to apoptotic pathway activation.34 The BDNF/TrkB pathway provides a protective effect by elevating SOD activity and lowering MDA levels, thus mitigating neuronal damage and apoptosis.35 DEX, an esterophilic compound capable of penetrating the blood-brain barrier, safeguards neuronal cells from inflammation, oxidative stress, apoptosis, and diverse neurodegenerative processes.36–39 Aligned with these findings, the current study demonstrates that DEX diminishes hippocampal neuron apoptosis in RSD rats by reducing MDA levels and boosting SOD activity. Notably, DEX is considered safe under normal conditions, with minimal potential to induce apoptosis.40 Thus, in RSD rats, DEX confers protection against oxidative stress and apoptosis, potentially ameliorating sleep deprivation-induced cognitive impairment. However, the explicit mechanism through which DEX confers this neuroprotection remains elusive. Moreover, the inhibitory effect of ANA-12 on DEX-mediated protection underscores the importance of the BDNF/TrkB pathway in mitigating oxidative stress and neuronal apoptosis associated with RSD.RSD can inhibit the BDNF/TrkB pathway, leading to hippocampal neuronal damage and cognitive dysfunction.41 As an α2-adrenergic receptor agonist, DEX may protect against such damage by modulating neurotransmitter release, enhancing BDNF expression, and activating TrkB (Saral S. et al, 2024). BDNF itself is vital for neural development and plasticity, promoting neuronal survival, growth, and differentiation through TrkB dimerization and downstream CREB activation.42,43 CREB plays an essential role in neuronal development, long-term memory formation, and survival.44 In this study, sleep-deprived rats treated with dexmedetomidine (DEX) showed elevated BDNF, TrkB, and CREB protein levels, suggesting that DEX can mitigate RSD-induced hippocampal damage and cognitive deficits. Conversely, blocking TrkB with ANA-12 reduced these protein levels and inhibited DEX’s neuroprotective effect, indicating that DEX’s mechanism largely depends on activating the BDNF/TrkB/CREB pathway.
Several limitations of this study should be acknowledged. First, we used a rat model of REM sleep deprivation (RSD), which may not fully recapitulate the complex mechanisms of sleep disturbances and cognitive impairment in humans. Second, although ANA-12 was used to block TrkB activation, we cannot completely rule out the involvement of other signaling pathways downstream of BDNF or independent neuroprotective mechanisms of dexmedetomidine. Third, our study primarily focused on hippocampal outcomes, while other brain regions involved in cognition, such as the prefrontal cortex and amygdala, were not examined and may contribute to the observed effects. Additionally, while our results strongly suggest that the BDNF/TrkB pathway mediates dexmedetomidine’s protective effects, it is possible that alternative mechanisms—such as modulation of GABAergic activity, direct antioxidant properties, or systemic anti-inflammatory effects—also play a role. Future studies employing genetic knockdown models, pathway-specific inhibitors, or multi-regional brain analyses are warranted to confirm causality and extend the translational relevance of these findings. In this study, a modified multi-platform method was used to create the RSD model, and the findings demonstrated both behaviorally and histologically that DEX protected RSD rats from spatial learning and memory deficits, as well as from neuronal damage and apoptosis in the hippocampal CA1 region.
Conclusion
In summary, our study demonstrates that dexmedetomidine (DEX) effectively mitigates hippocampal neuronal damage and cognitive impairment in REM sleep-deprived (RSD) rats. Mechanistically, DEX upregulated BDNF expression, enhanced TrkB activation, and subsequently reduced neuronal apoptosis and neuroinflammation, suggesting that its neuroprotective effects are mediated, at least in part, through the BDNF/TrkB signaling pathway. These findings not only provide mechanistic insights into the protective role of DEX against sleep deprivation-induced hippocampal dysfunction but also support its potential as a therapeutic strategy for preventing or treating sleep deprivation-related neurological disorders.
Abbreviations
DEX, Dexmedetomidine; RSD, Rapid eye movement sleep deprivation; BDNF, Brain-derived neurotrophic factor; SPF, Specific Pathogen Free; OD, Optical density; TNFα, Tumor necrosis factor α; MDA, Malondialdehyde; IL-1β, Interleukin 1β; SOD, Superoxide dismutase; ECL, Enhanced chemiluminescence; MMPM, Modified multi-platform method.
Data Sharing Statement
The raw data supporting the findings of this study are available from the corresponding upon reasonable request.
Ethics Statement
This animal experiment was approved by the Animal Ethics Committee of Fujian Medical University. The study was conducted in accordance with the local legislation and institutional requirements.
Acknowledgments
We are grateful to the members of the laboratory for their valuable discussions. We also appreciate Bohan Biotechnology Co. (Fuzhou, China) for providing technical guidance and acknowledge Anesthesiology Research Institute, The First Affiliated Hospital, Fujian Medical University for providing instrumentation facilities. The authors declare that no Generative AI was used in the creation of this paper.
Author Contributions
B. Z. and Y. L. are the co-first authors. BZ: Data curation, Funding acquisition, Writing–original draft, Writing–review & editing. YL: Data curation, Methodology, Writing–original draft. CL: Data curation, Methodology, Writing–original draft. All authors agreed on the journal to which the article will be submitted; agreed on the final version accepted for publication; and agree to take responsibility and be accountable for the contents of the article.
Funding
The authors declare that the research, writing and/or publication of this article was supported by Joint Funds for the Innovation of Science and Technology, Fujian Province (No. 2021Y9127); Natural Science Foundation of Fujian Province (No. 2023J01593); and Startup Fund for Scientific Research of Fujian Medical University (No. 2021QH1069).
Disclosure
The authors declare that this study was conducted without any business or financial relationship that could be considered a potential conflict of interest.
References
1. Lyons LC, Vanrobaeys Y, Abel T. Sleep and memory: the impact of sleep deprivation on transcription, translational control, and protein synthesis in the brain. J Neurochem. 2023;166(1):24–46. doi:10.1111/jnc.15787
2. Gao J, Jiang Z, Tian L, Chen H. Effects of phototherapy on memory and BDNF/TrkB signaling pathway in sleep-deprived mice. In: AOPC 2021: Biomedical Optics. Vol. 12067. SPIE; 2021:92–96.
3. Dai D, Zheng B, Yu Z, Lin S, Tang Y, Chen M. Right stellate ganglion block improves learning and memory dysfunction and hippocampal injury in rats with sleep deprivation. BMC Anesthesiol. 2021;21(1):272. doi:10.1186/s12871-021-01486-4
4. Chen Z, Zuo Z, Song X, Zuo Y, Zhang L, Ye Y. Mapping Theme Trends and Research Frontiers in Dexmedetomidine Over Past Decade: a Bibliometric Analysis. Drug Des Devel Ther. 2024;18:3043–3061. doi:10.2147/DDDT.S459431
5. Wang J, Xin Y, Chu T, Liu C, Xu A. Dexmedetomidine attenuates perioperative neurocognitive disorders by suppressing hippocampal neuroinflammation and HMGB1/RAGE/NF-κB signaling pathway. Biomed Pharmacother. 2022;150:113006. doi:10.1016/j.biopha.2022.113006
6. Chen X, Chen D, Li Q, et al. Dexmedetomidine alleviates hypoxia‐induced synaptic loss and cognitive impairment via inhibition of microglial NOX2 activation in the hippocampus of neonatal rats. Oxid Med Cell Longev. 2021;2021(1):6643171. doi:10.1155/2021/6643171
7. Chen L, Yue Z, Liu Z, et al. The impact of Nrf2 knockout on the neuroprotective effects of dexmedetomidine in a mice model of cognitive impairment. Behav Brain Res. 2024;469:115006. doi:10.1016/j.bbr.2024.115006
8. Jeon S, Baik J, Kim J, Lee J, Do W, Kim E. Intrathecal dexmedetomidine attenuates mechanical allodynia through the downregulation of brain-derived neurotrophic factor in a mild traumatic brain injury rat model. Korean J Anesthesiol. 2023;76(1):56–66. doi:10.4097/kja.22209
9. Azman KF, Zakaria R. Recent Advances on the Role of Brain-Derived Neurotrophic Factor (BDNF) in Neurodegenerative Diseases. Int J Mol Sci. 2022;23(12):6827. doi:10.3390/ijms23126827
10. Numakawa T, Kajihara R. The Role of Brain-Derived Neurotrophic Factor as an Essential Mediator in Neuronal Functions and the Therapeutic Potential of Its Mimetics for Neuroprotection in Neurologic and Psychiatric Disorders. Molecules. 2025;30(4):848. doi:10.3390/molecules30040848
11. Gupta AK, Gupta S, Mehan S, Khan Z, Das Gupta G, Narula AS. Exploring the Connection Between BDNF/TrkB and AC/cAMP/PKA/CREB Signaling Pathways: potential for Neuroprotection and Therapeutic Targets for Neurological Disorders. Molecular Neurobiol. 2025;1–33. doi:10.1007/s12035-024-04220-6
12. Ali NH, Al-Kuraishy HM, Al-Gareeb AI, Alexiou A, Papadakis M, AlAseeri AA. BDNF/TrkB activators in Parkinson’s disease: a new therapeutic strategy. J Cell Mol Med. 2024;28(10):e18368. doi:10.1111/jcmm.18368
13. Numakawa T, Kajihara R. An Interaction between Brain-Derived Neurotrophic Factor and Stress-Related Glucocorticoids in the Pathophysiology of Alzheimer’s Disease. Int J Mol Sci. 2024;25(3):1596. doi:10.3390/ijms25031596
14. Chen X, Chen A, Wei J, et al. Dexmedetomidine alleviates cognitive impairment by promoting hippocampal neurogenesis via BDNF/TrkB/CREB signaling pathway in hypoxic–ischemic neonatal rats. CNS Neurosci Ther. 2024;30(1):e14486. doi:10.1111/cns.14486
15. Kang JY, Lee JS, Wang JH, Son CG. Sleep deprivation in adolescent mice impairs long-term memory till early adulthood via suppression of hippocampal astrocytes. Sleep. 2024;47(10):zsae143. doi:10.1093/sleep/zsae143
16. Jimenez-Tellez N, Iqbal F, Pehar M, Casas-Ortiz A, Rice T, Syed NI. Dexmedetomidine does not compromise neuronal viability, synaptic connectivity, learning and memory in a rodent model. Sci Rep. 2021;11(1):16153. doi:10.1038/s41598-021-95635-x
17. Beaudin AE, Younes M, Gerardy B, Raneri JK, Hirsch allen AJM, Gomes T. Association between sleep microarchitecture and cognition in obstructive sleep apnea. Sleep. 2024;47(12):zsae141. doi:10.1093/sleep/zsae141
18. Figorilli M, Meloni M, Lanza G, Casaglia E, Lecca R, Saibene FL. Considering REM Sleep Behavior Disorder in the Management of Parkinson’s Disease. Nat Sci Sleep. 2023;15:333–352. doi:10.2147/NSS.S266071
19. Fang S, Dai J, Guo W, Ma T. Effect of sleep deprivation on general anesthesia in rats. Int J Burns Trauma. 2020;10(3):47–54. PMID: 32714627; PMCID: PMC7364414.
20. Jones EAA, Rao A, Zilberter M, et al. Dentate gyrus and CA3 GABAergic interneurons bidirectionally modulate signatures of internal and external drive to CA1. Cell Rep. 2021;37(13):1.
21. Zhang Z, Ferretti V, İ G, Moro A, Steinberg EA, Ye Z. Neuronal ensembles sufficient for recovery sleep and the sedative actions of α2 adrenergic agonists. Nat Neurosci. 2023;18(4):553–561. doi:10.1038/nn.3957
22. Brito RA, Viana SMDNR, Beltro BA, Magalhes CBDA, Bruin VMSD, Bruin PFCD. Pharmacological and non-pharmacological interventions to promote sleep in intensive care units: a critical review. Sleep Breath. 2019;24(1):25–35. doi:10.1007/s11325-019-01902-7
23. Moon EJ, Ko IG, Kim SE, Jin JJ, Hwang L, Kim CJ. Dexmedetomidine Ameliorates Sleep Deprivation-Induced Depressive Behaviors in Mice. Int Neurourol J. 2018;22(Suppl 3):S139–146. doi:10.5213/inj.1836228.114
24. Bo JH, Wang JX, Wang XL, et al. Dexmedetomidine attenuates lipopolysaccharide-induced sympathetic activation and sepsis via suppressing superoxide signaling in paraventricular nucleus. Antioxidants. 2022;11(12):2395. doi:10.3390/antiox11122395
25. Cheng W, Song Y, Liu Y, Sun X, Ren W. Impact of dexamethasone preconditioning on prevention of development of cognitive impairment following acute inflammation. Contrast Media Mol Imag. 2022;2022(1):6064007. doi:10.1155/2022/6064007
26. Zeng L, Zhao C, Cui C, et al. Dexmedetomidine improves DM-induced oxidative stress injury to protect liver function through Nrf2 pathway. Am J Transl Res. 2023;15(3):1715.
27. Hu Y, Zhou H, Zhang H, et al. The neuroprotective effect of dexmedetomidine and its mechanism. Front Pharmacol. 2022;13:965661. doi:10.3389/fphar.2022.965661
28. Tao Z, Li P, Zhao X. Progress on the mechanisms and neuroprotective benefits of Dexmedetomidine in brain diseases. Brain and Behavior. 2024;14(11):e70116. doi:10.1002/brb3.70116
29. Unchiti K, Leurcharusmee P, Samerchua A, Pipanmekaporn T, Chattipakorn N, Chattipakorn SC. The potential role of dexmedetomidine on neuroprotection and its possible mechanisms: evidence from in vitro and in vivo studies. Eur J Neurosci. 2021;54(9):7006–7047. doi:10.1111/ejn.15474
30. He Z, Zang H, Zhu L, Huang K, Yi T, Zhang S. An anti-inflammatory peptide and brain-derived neurotrophic factor-modified hyaluronan-methylcellulose hydrogel promotes nerve regeneration in rats with spinal cord injury. Int J Nanomed. 2019;14:721–732. doi:10.2147/IJN.S187854
31. Xiong HY, Hendrix J, Schabrun S, Wyns A, Campenhout JV, Nijs J. The Role of the Brain-Derived Neurotrophic Factor in Chronic Pain: links to Central Sensitization and Neuroinflammation. Biomolecules. 2024;14(1):71. doi:10.3390/biom14010071
32. Nociti V, Romozzi M. The Role of BDNF in Multiple Sclerosis Neuroinflammation. Int J Mol Sci. 2023;24(9):8447. doi:10.3390/ijms24098447
33. Porter GA, O’Connor JC. Brain-derived neurotrophic factor and inflammation in depression: pathogenic partners in crime? World J Psychiatry. 2022;12(1):77–97. doi:10.5498/wjp.v12.i1.77
34. Chen L, Cao J, Cao D, Wang M, Cong H. Protective effect of dexmedetomidine against diabetic hyperglycemia-exacerbated cerebral ischemia/reperfusion injury: an in vivo and in vitro study. Life Sci. 2022;235:116553. doi:10.1016/j.lfs.2019.116553
35. Dong J, Wei R, Zong F, Wang Z, Ma S, Zhao W. Phosphodiesterase 7 inhibitor reduces stress-induced behavioral and cytoarchitectural changes in C57BL/6J mice by activating the BDNF/TrkB pathway. Front Pharmacol. 2024;15:1411652. doi:10.3389/fphar.2024.1411652
36. Cho I, Koo BN, Kim SY, Park S, Kim EJ, Kam EH. Neuroprotective Effect of Dexmedetomidine against Postoperative Cognitive Decline via NLRP3 Inflammasome Signaling Pathway. Int J Mol Sci. 2022;23(15):8806. doi:10.3390/ijms23158806
37. Puls R, von Haefen C, Bührer C, Endesfelder S. Dexmedetomidine Protects Cerebellar Neurons against Hyperoxia-Induced Oxidative Stress and Apoptosis in the Juvenile Rat. Int J Mol Sci. 2023;24(9):7804. doi:10.3390/ijms24097804
38. Chen Y, Zhang X, Zhang B, He G, Zhou L, Xie Y. Dexmedetomidine reduces the neuronal apoptosis related to cardiopulmonary bypass by inhibiting activation of the JAK2-STAT3 pathway. Drug Des Devel Ther. 2017;11:2787–2799. doi:10.2147/DDDT.S140644
39. Tao QY, Liu D, Wang XJ, Wang X, Ouyang RN, Niu JY. Effects of Esketamine Combined with Dexmedetomidine on Early Postoperative Cognitive Function in Elderly Patients Undergoing Lumbar Spinal Surgery: a Double-Blind Randomized Controlled Clinical Trial. Drug Des Devel Ther. 2024;18:5461–5472. doi:10.2147/DDDT.S481173
40. Inoue T, Suzuki-Karasaki Y. Mitochondrial superoxide mediates mitochondrial and endoplasmic reticulum dysfunctions in TRAIL-induced apoptosis in Jurkat cells. Free Radic Biol Med. 2013;61:273–284. doi:10.1016/j.freeradbiomed.2013.04.020
41. Mahboubi S, Nasehi M, Imani A, Sadat-Shirazi MS, Zarrindast MR, Vousooghi N. Benefit effect of REM-sleep deprivation on memory impairment induced by intensive exercise in male wistar rats: with respect to hippocampal BDNF and TrkB. Nat Sci Sleep. 2019;24:179–188. doi:10.2147/NSS.S207339
42. Saral S, Mercantepe T, Topçu A, Kaya AK, Öztürk A. Dexmedetomidine Improves Learning Functions in Male Rats Modeling Cognitive Impairment by Modulating the BDNF/TrkB/CREB Signaling Pathway. Life. 2024;14(12):1672. doi:10.3390/life14121672. PMID: 39768379; PMCID: PMC11728090.
43. Cao J, Gorwood P, Ramoz N, Viltart O. The Role of Central and Peripheral Brain-Derived Neurotrophic Factor (BDNF) as a Biomarker of Anorexia Nervosa Reconceptualized as a Metabo-Psychiatric Disorder. Nutrients. 2024;16(16):2617. doi:10.3390/nu16162617
44. Koga Y, Tsurumaki H, Aoki-Saito H, Sato M, Yatomi M, Takehara K. Roles of Cyclic AMP Response Element Binding Activation in the ERK1/2 and p38 MAPK Signalling Pathway in Central Nervous System, Cardiovascular System, Osteoclast Differentiation and Mucin and Cytokine Production. Int J Mol Sci. 2019;20(6):1346. doi:10.3390/ijms20061346
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Empagliflozin Improves Learning and Memory Deficits in Streptozotocin-Induced Hyperglycemic Male Wistar Rats
Isawi IH, Al-Sawalha NA, Mahmoud F, Al-Khawaldeh I, Alzoubi KH
Journal of Experimental Pharmacology 2026, 18:561692
Published Date: 11 February 2026
Pharmacological Inhibition of Tropomyosin Receptor Kinase B Reverses Remifentanil-Induced Postoperative Hyperalgesia and Synaptic Remodeling
Yang L, Huang Y, Long B
Journal of Pain Research 2026, 19:598801
Published Date: 29 June 2026