Back to Journals » Cancer Management and Research » Volume 18

Cytotoxic Effect of Halimeda macroloba Crude Extract on Cervical Cancer Spheroid Cells via p-mTOR/mTOR, Apoptosis, and Ferroptosis Pathways
Authors Nittayaboon K ![]() , Leetanaporn K
, Leetanaporn K ![]() , Kerdkumthong K, Mayakun J, Navakanitworakul R
, Kerdkumthong K, Mayakun J, Navakanitworakul R
Received 25 March 2026
Accepted for publication 3 July 2026
Published 10 July 2026 Volume 2026:18 611650
DOI https://doi.org/10.2147/CMAR.S611650
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Kesara Nittayaboon,1 Kittinun Leetanaporn,1,2 Kankamol Kerdkumthong,1 Jaruwan Mayakun,3 Raphatphorn Navakanitworakul1,2
1Department of Biomedical Sciences and Biomedical Engineering, Faculty of Medicine, Prince of Songkla University, Hat Yai, Songkhla, 90110, Thailand; 2Translational Medicine Research Center, Faculty of Medicine, Prince of Songkla University, Hat Yai, Thailand; 3Division of Biological Science, Faculty of Science, Prince of Songkla University, Hat Yai, Songkhla, 90110, Thailand
Correspondence: Jaruwan Mayakun, Department of Biomedical Sciences and Biomedical Engineering, Faculty of Medicine, Prince of Songkla University, Hat Yai, Songkhla, 90110, Thailand, Email [email protected] Raphatphorn Navakanitworakul, Division of Biological Science, Faculty of Science, Prince of Songkla University, Hat Yai, Songkhla, 90110, Thailand, Email [email protected]
Background: Because of the limited efficacy of existing therapeutic strategies and the emergence of treatment resistance, cervical cancer remains a major global health burden. Recently, marine-derived natural compounds have attracted attention as potential sources of novel anticancer agents. We aimed to investigate the cytotoxic effects of crude Halimeda macroloba extract on cervical cancer spheroid cells and to explore its underlying molecular mechanisms.
Methods: Three-dimensional spheroid cultures of SiHa cervical cancer cells were treated with varying concentrations of H. macroloba extract, followed by assessment of migration, cell viability, caspase activity, and protein expression profiles using liquid chromatography–tandem mass spectrometry, with selected findings validated by Western blotting.
Results: Treatment with H. macroloba extract significantly reduced spheroid migration and decreased cell viability in a dose-dependent manner, accompanied by increased caspase activity. Proteomic analysis demonstrated altered expression of proteins associated with mitochondrial function, apoptosis-related pathways, and oxidative stress-related processes. Mechanistic studies further revealed downregulation of phosphorylated mTOR (p-mTOR) and total mTOR, together with increased expression of pro-apoptotic markers.
Conclusion: The H. macroloba crude extract exerts anticancer effects in cervical cancer spheroids, primarily through apoptosis-associated mechanisms and modulation of the mTOR signaling pathway, supporting its potential for further investigation as a marine-derived bioactive compound.
Keywords: cervical cancer, natural extracts, proteomic analysis, anticancer agents
Introduction
Natural products exhibit a wide range of biological properties beneficial for medical applications. These compounds are being increasingly recognized as crucial contributors to drug discovery and development. Among them, the marine calcified green seaweed, Halimeda (J. V. Lamouroux), belonging to the Halimedaceae family, Bryopsidales order, and Bryopidopyceae class, has emerged as a promising candidate for biomedical research. Halimeda is broadly distributed across tropical and subtropical regions and is abundant in numerous reef systems such as the Florida Keys, Southwest Atlantic Ocean, and Indo-Pacific Ocean.1–6 In Thailand, Halimeda macroloba is particularly abundant in the shallow subtidal and intertidal zones, and forms dense populations in the intertidal zone of Lidee Island, Satun Province.7 H. macroloba is an important calcium carbonate producer that contributes to the carbonate budget.8,9 Halimeda produces hydrocolloids, which are utilized in both the food and pharmaceutical industries, thereby making it economically relevant. Furthermore, Halimeda is a rich source of bioactive compounds including carotenoids, xenicane lactones, sym-triazine derivatives, bis-indole alkaloids, bis-nor diterpene derivatives, phenolics, flavonoids, terpenoids, and polysaccharides. This species also exhibits high phenolic and flavonoid content, contributing to its strong antioxidant potential. These secondary metabolites display diverse pharmacological activities including antioxidant, anticancer, anti-inflammatory, antimicrobial, and antiviral properties.2,10
Cervical cancer is the fourth most prevalent cancer in women worldwide. Breast cancer is the second most common type of cancer in Thailand. The introduction of screening programs and human papilloma virus (HPV) vaccination has contributed to a reduction in incidence rates.11 Standard treatment options for cervical cancer include surgery, radiotherapy, chemotherapy, and combined chemoradiotherapy.11,12 However, the mortality rate associated with cervical cancer in Thailand has increased, thereby indicating a critical gap in research and intervention. Natural compounds represent a major source of clinically approved anticancer agents and continue to serve as a critical reservoir for oncologic drug discovery.13 The potential therapeutic activity of Halimeda has been reported in various cancer types, including cervical cancer.10 However, the chemical composition of Halimeda varies depending on environmental conditions and nutrient availability across different geographical regions. Despite the reported bioactive properties of Halimeda species, the anticancer effects and underlying molecular mechanisms of H. macroloba in cervical cancer remain poorly understood, particularly in physiologically relevant 3D tumor models. Three-dimensional spheroid cultures more closely mimic the tumor microenvironment, including cell–cell interactions, nutrient gradients, hypoxic regions, and drug penetration characteristics observed in solid tumors, thereby providing a more predictive platform for evaluating anticancer activity than conventional 2D monolayer cultures.14 In addition, growing evidence suggests that marine-derived natural compounds may modulate oxidative stress and lipid peroxidation-associated pathways, including ferroptosis-related mechanisms, which have emerged as potential therapeutic targets in cancer treatment. However, the possible involvement of ferroptosis-associated pathways in the anticancer effects of H. macroloba has not yet been investigated.
In this study, we aimed to investigate the cytotoxic effects of H. macroloba extract on cervical cancer spheroids and explored the associated molecular mechanisms, including apoptosis- and ferroptosis-related pathways. Figure 1 shows the research workflow.
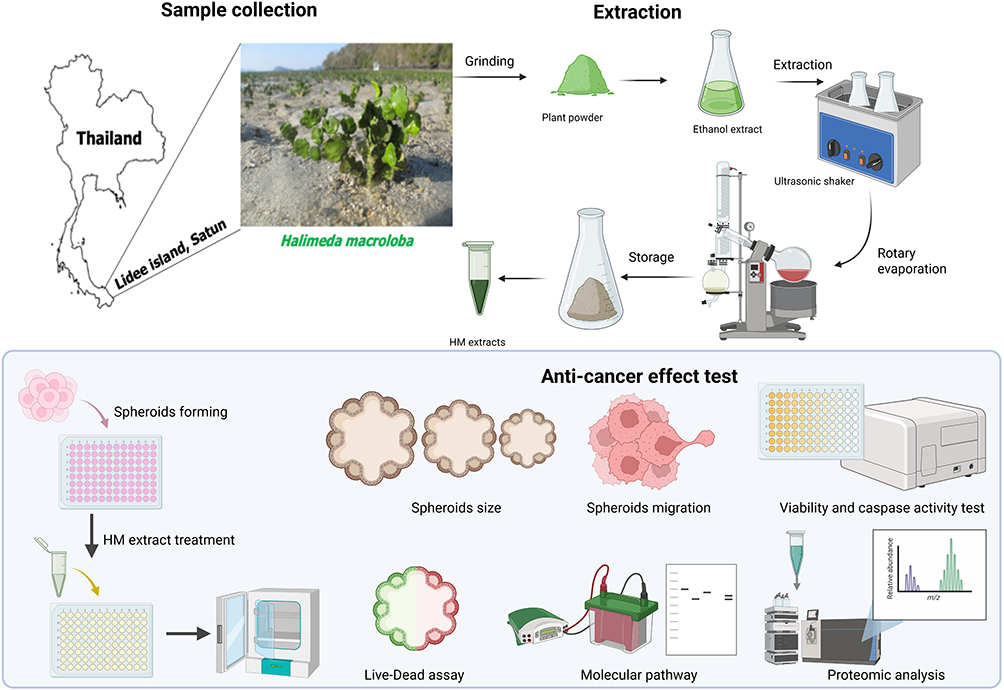 |
Figure 1 Research workflow. The upper and lower panels illustrate the crude extract preparation process, including sample collection and extraction step, and anticancer testing procedure, respectively. |
Materials and Methods
Cervical Cancer Cell Culture and Spheroid Culture
A squamous cell carcinoma of cervical cancer cells, SiHa (ATCC HTB-35), was purchased from the American Type Culture Collection (ATCC) (Manassas, VA). The cells were cultured in Dulbecco’s Modified Eagle’s Medium (DMEM, Gibco, Thermo Fisher Scientific, USA) supplemented with 10% fetal bovine serum (FBS, Gibco, Thermo Fisher Scientific, USA), L-glutamine, streptomycin (100 µg/mL, Gibco, Thermo Fisher Scientific, USA), and penicillin (100 U/mL, Gibco, Thermo Fisher Scientific, USA), and incubated in humidified air with 5% CO2 incubator at 37 °C.
Spheroid cells were grown on poly-(2-hydroxyethyl methacrylate)-or poly HEMA-coated plates (Sigma-Aldrich, USA). The low-attachment plates were prepared by transferring an aliquot of 20 μL of Poly-HEMA working solution (5 mg/mL) into a single well of a 96-well U-bottom plate (Thermo Fisher Scientific, USA). Subsequently, the plates were dried in an incubator for 3 days. SiHa cells were seeded into the plates at a density of 5000 cells/well in 10% FBS-DMEM complete medium to generate spheroid cells.14
H. macroloba Extract Preparation
H. macroloba extracts were prepared and subjected to phytochemical analysis. Briefly, H. macroloba was dried, ground, extracted with ethanol, and concentrated under reduced pressure. Phytochemical screening and thin-layer chromatography were performed as previously described.7 The major identified bioactive constituents included alkaloids, flavonoids, phenolic compounds, terpenoids, steroids, and saponins. For cytotoxicity assays, the extracts were dissolved in dimethyl sulfoxide (DMSO).
Spheroid Size and Migration Evaluation
Spheroid size was evaluated using three independent biological replicates, with each replicate containing three spheroids per condition. Spheroids were imaged on day 3 following treatment with H. macroloba extract, and spheroid diameters were measured using ImageJ software (version 1.54). The average spheroid diameter was calculated and compared among treatment groups. For the spheroid migration assay, spheroids were collected on day 3 post-treatment and transferred to flat-bottom plates containing fresh complete medium. The spheroids were further incubated for 72 h to allow cell migration. Migration areas were subsequently imaged and quantified using ImageJ software. Both spheroid diameter and migration area were analyzed quantitatively.
Cytotoxicity Test
Cell viability and cytotoxicity were evaluated following a 3-day treatment with H. macroloba extract using a LIVE/DEAD Cell Imaging Kit (Thermo Fisher Scientific, USA). Experiments were performed using three independent biological replicates, each consisting of three spheroids per treatment condition. Calcein AM was used to identify viable cells, whereas BOBO-3 iodide was used to detect dead cells. Prior to staining, the culture medium volume was adjusted to 30 µL. Following incubation with the staining solution at 37 °C for 30 min in the dark, fluorescence images were acquired using a Lionheart FX live-cell imaging system.
The mode of cell death was further characterized following a 3-day treatment using the ApoLive-Glo™ Multiplex Assay (Promega, WI, USA). The assay was performed using three independent biological replicates, each containing three spheroids per condition. This assay simultaneously measures protease activity in viable cells through fluorescence-based detection and caspase-3/7 activity in apoptotic cells via luciferase-based luminescence.
Proteomic Analysis Using Liquid Chromatography Tandem Mass Spectroscopy (LC–MS/MS)
Proteomic analysis was conducted to evaluate protein expression following H. macroloba extract treatment. Spheroids with and without treatment were collected in three independent passages, and proteins were extracted using a filter-aided sample preparation (FASP) protocol with minor modifications.15 In brief, 50 μg of protein was diluted in 50 mM Tris-HCl buffer (pH 8.0) containing 4% sodium dodecyl sulfate (SDS) and heated at 95 °C for 5 min. Thereafter, the samples were washed thrice with 8 M urea in 5 mM Tris-HCl and incubated with 5 mM DTT at 37 °C for 60 min, followed by 15 mM IAA at 25 °C for 30 min, and washed thrice with NH4CO3. Protein digestion was conducted using trypsin at a 50:1 (protein: enzyme, w/w) ratio in 50 mM NH4CO3 at 37 °C overnight. The reaction was quenched with 1% formic acid (FA), and peptides were desalted using ZipTip (Sigma Aldrich) prior to LC-MS/MS analysis.
For LC–MS/MS analysis, protein samples were analyzed using a Vanquish Neo UHPLC system (Thermo Fisher Scientific, USA) coupled with an Orbitrap Exploris 480 mass spectrometer (Thermo Fisher Scientific). Tryptic digest (1 µg) was loaded onto an Acclaim PepMap C18 trap column (5 μm, 300 μm × 5 mm), followed by peptide separation on a PepMap C18 analytical column (2 μm, 75 μm × 500 mm) at a flow rate of 0.3 μL/min with an electrospray voltage of 2 kV. Chromatographic separation was achieved using a linear gradient from 2% to 45% solvent B (solvent A: 2% acetonitrile with 0.1% formic acid; solvent B: 80% acetonitrile with 0.1% formic acid) for 70 min, and was followed by a 5-min wash with 95% solvent B. The column was maintained at 60 °C throughout the experiments. MS1 scans were acquired within an m/z range of 400–1000 at a resolution of 120,000 with an automatic gain control (AGC) target of 300%. MS2 spectra were generated using data-independent acquisition (DIA) with optimized isolation windows. DIA settings included 20 m/z isolation windows, a resolution of 30,000, an AGC target of 1000%, and a normalized collision energy of 28% using high-energy collisional dissociation.
For data processing, raw spectral files were analyzed using the DIA-NN version 1.9.2.16 The data were searched against the UniProt Homo sapiens proteome (UP000005640) with the following parameters: allowance for one missed cleavage, N-terminal methionine excision, carbamidomethylation of cysteines as a fixed modification, generic scoring, MS1 and MS2 mass accuracies set to 20 ppm and 10 ppm, respectively, retention time (RT)-time-dependent normalization, high-precision quantification strategy, and match-between-runs functionality. Precursor and protein identification were filtered using a 1% false discovery rate.
For data analysis, proteins detected in fewer than 50% of samples were excluded from further analyses. The remaining missing values were imputed using the Minimal Detection Value method in the imputeLCMD package of R. Additionally, protein expression patterns were visualized using heat maps (MetaboAnalyst). The number of proteins expressed in the treatment and control groups was visualized using Venn diagrams (ref WebBase). Differential expression was determined using the limma package. Consequently, proteins were classified as differentially expressed when meeting the thresholds of p < 0.05 and fold change > |1.5|. Finally, upregulated and downregulated proteins were classified using PANTHER (Protein ANalysis THrough Evolutionary Relationships), and gene enrichment was performed with the clusterProfiler package in R for molecular function (MF) and biological process (BP) categories using Fisher’s exact test with Benjamini–Hochberg correction (p < 0.2).
Molecular Mechanism of H. macroloba Extract Active Compounds
Western blot analysis of spheroids was performed as previously described.14 Briefly, spheroid pellets collected from three independent passages, with each passage containing nine spheroids, were lysed in radioimmunoprecipitation assay (RIPA) buffer (Pierce Biotechnology, Waltham, MA, USA). The protein concentration was measured using the Bradford assay (Bio-Rad, USA). A total of 30 μg of protein extract was subjected to SDS-polyacrylamide gel electrophoresis (SDS-PAGE), followed by transfer to polyvinylidene difluoride membranes (Amersham Pharmacia Biotech, USA). The membranes were blocked in 5% nonfat milk in Tris-buffered saline containing 0.1% (v/v) Tween-20 (TBS-T) for 1 h at room temperature. The cells were then washed twice with TBS-T for 10 min. Each membrane was incubated overnight at 4 °C, while shaking continuously with primary antibodies (1:1000 diluted with 1% non-fat milk in TBS-T) specific to Akt (cat. no. 4691; Cell Signaling Technology, Inc)., phospho-AKT (Ser473; cat. no. 4060; Cell Signaling Technology, Inc)., phosphorylated mTOR (Ser2448; cat. no. 5536; Cell Signaling Technology, Inc)., USA), mTOR (cat. no. 2983; Cell Signaling Technology, Inc., Danvers, MA, USA), cRaf (cat. no. 9421; Cell Signaling Technology, Inc)., Bax (cat. no. 2772; Cell Signaling Technology, Inc)., cleavage-caspase 3 (cat. no. 9664; Cell Signaling Technology, Inc)., cytochrome C (cat. no. 4272; Cell Signaling Technology, Inc)., and β-actin (cat. no. 4967; Cell Signaling Technology) were used as internal control. After incubation, the membranes were washed thrice (10 min/wash) with TBS-T and incubated with a secondary antibody (Anti-rabbit IgG horseradish peroxidase; cat. no. 7074; Cell Signaling Technology, Inc)., at 1:1000 diluted in 1% nonfat milk in TBS-T for 2 h. Thereafter, the membranes were again washed three times with TBS-T, and the last washing was performed using TBS for 10 min. Protein expression was visualized using an enhanced chemiluminescence reagent (Pierce ECL Western Blotting Substrate, Thermo Fisher Scientific, USA). Densitometry was performed using a Chemiluminescence & Epi Fluorescence Alliance Q9 Advanced Imager (UVITEC, UK).
Statistical Analysis
Statistical analyses were performed using the GraphPad Prism Software (version 9.00; GraphPad, San Diego, CA, USA). Student’s t-test (two-tailed) was used to compare two independent group means, whereas one- or two-way ANOVA (followed by Tukey‘s or Sidak’s multiple comparison tests) was used to compare more than two independent group means. A p-value < 0.05 were considered statistically significant.
Results
H. macroloba Extracts Inhibited Cervical Cancer Spheroid Migration but Not Spheroid Size
Prior to investigating the anticancer activity of H. macroloba extract, its safety profile was evaluated in non-cancerous cells using a 2D monolayer culture model. The extract exhibited minimal cytotoxicity toward L929 fibroblast cells, with an IC50 value exceeding 80 μg/mL. Similar findings were observed in HEK-293 cells in our previous study, where the IC50 value was also greater than 80 μg/mL.7 These results indicate a favorable preliminary safety profile and supported the subsequent evaluation of the extract in cervical cancer cell models. The corresponding data are provided in Supplementary Table S1.
Following confirmation of its preliminary safety profile, the effects of H. macroloba extract on spheroid morphology and migration were investigated. The morphology of the spheroids was captured after treatment with H. macroloba extract. The extracts showed anti-migration activity in spheroids, while spheroid size remained unaffected. Figure 2A shows comparable spheroid growth across treatment conditions. These results indicate that H. macroloba extracts did not affect spheroid size. Figure 2B shows a significant decrease in spheroid migration. This result indicates that H. macroloba extracts may inhibit the migration of SiHa spheroid cells.
 |
Figure 2 Effect of Halimeda macroloba extracts on cervical cancer spheroid. Cell morphology of SiHa spheroid after treatment for 72 h (upper panel). The migration of spheroids was observed after 72 h post-treatment (lower panel) (A). H. macroloba extracts showed no difference on spheroid size (B). Spheroid migration was significantly decreased (C). Scale bar = 400 µm. Statistical analysis was performed using the Student’s t-test. Significance: **** p < 0.0001. |
H. macroloba Extracts Induced Cervical Cancer Spheroid Death
After spheroid treatment, cell viability was determined by staining the spheroids using a LIVE/DEAD cell imaging kit. Figure 3 shows the cytotoxic effects of H. macroloba extracts on spheroid cells. The results demonstrated a decrease in green fluorescence from calcein AM-stained viable cells, and an increase in red fluorescence from nucleic acid staining-BOBO-3 for dead cells, thereby indicating enhanced cell death in the H. macroloba extract-treated group compared to the control. Our results revealed that dead cells—indicated by the red fluorescent signal—were located in the center of the SiHa spheroid, while the edge of spheroids showed a green fluorescent signal—indicating living cells.
 |
Figure 3 Cytotoxic effects of Halimeda macroloba extracts on cervical cancer spheroids. After treatment with H. macroloba extracts at various concentration for 72 h, the LIVE/DEAD staining (A, live = green, dead = red) of the spheroids was visualized with a LionheartFX live cell imager (10x magnification). Scale bar = 500 μm. The ApoLive-Glo™ assay was used to determine cell viability (B) and caspase-3/7 activity (C). Statistical analysis was performed using the Student’s t-test. Significance:*** p < 0.001, **** p < 0.0001. |
H. macroloba Extracts Induced Spheroid Death Through Caspase Activation
The mode of cell death after spheroid treatment was investigated using the ApoLive-Glo™ assay kit. Cell viability was evaluated using live cell protease activity to produce fluorescent AFC. Caspase activity was evaluated based on the cleavage of caspase, which is a substrate for luciferase (aminoluciferin). Protease activity was calculated as a percentage of viable cells. Figures 3B and C show the cell viability and caspase activity of the H. macroloba extracts, respectively. The results revealed a decrease in cell viability, which correlated with the LIVE/DEAD assay. These results confirmed that H. macroloba extracts exhibited dose-dependent cytotoxicity against SiHa spheroids. Moreover, caspase activity was measured to confirm apoptosis. We observed an increase in caspase activity in the H. macroloba extract-treated groups. However, caspase activity decreased slightly at 25 µg/mL and increased at 50 µg/mL. We hypothesized that this is attributed to negative feedback in the 25 µg/mL treatment group.
Proteomic Analysis Revealed Cell Death Induction Through Apoptosis and Ferroptosis Pathway
The protein expression profiles of SiHa spheroids incubated with or without H. macroloba extract are shown in Figure 4. The Venn diagram showed that 120 proteins were uniquely expressed in the control group (HM-0), whereas 47 proteins were uniquely expressed in the treatment group (HM-200). Gene Ontology analysis of these unique proteins revealed distinct expression patterns between the groups. In terms of MF, both groups exhibited proteins associated with structural molecular activity, MF regulation, and, most prominently, catalytic activity, with a higher percentage in the HM-200 treatment group. Additionally, transcriptional regulator and transporter activities were higher in the control group than in the treatment group. Analysis of BPs indicated that proteins involved in the response to stimuli and biological regulation were classified in the control group. Meanwhile, proteins associated with homeostatic, metabolic, and cellular processes were classified in the treatment group. Classification by protein class further demonstrated that metabolite interconversion enzymes and membrane trafficking proteins were highly expressed in the HM-200 treatment group.
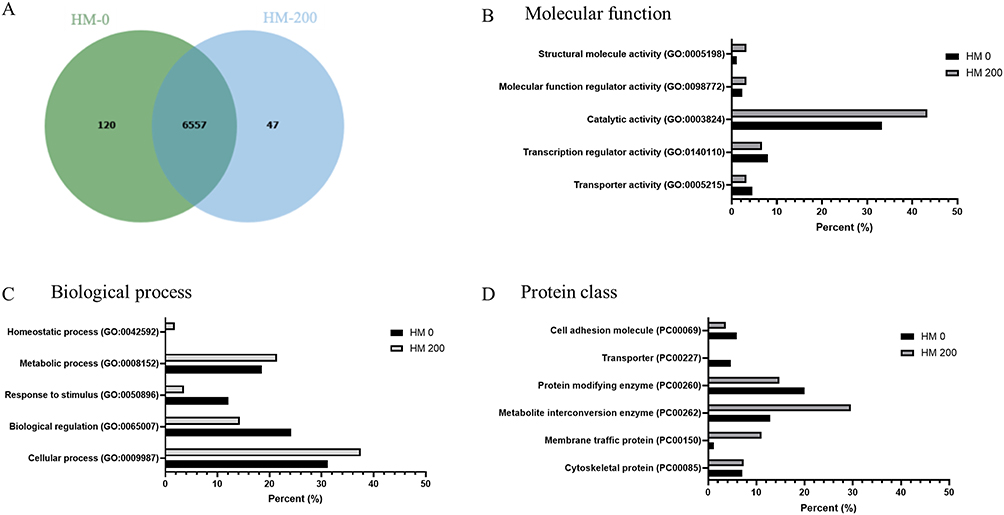 |
Figure 4 Proteomic data revealed protein expression changes associated with the bioactive effects of Halimeda macroloba extract in cervical cancer spheroids. Venn diagram of control (HM-0) and H. macroloba extracts (HM-200) on cervical cancer spheroid (A). A unique protein list in each group was subjected to the Panther database (PANTHER 19.0) to identify the molecular function using GO terms annotated (B), biological process (C), and protein class (D). |
Figure 5A presents a heatmap of the top 100 differentially expressed proteins, and Figures 5B and C illustrate the enrichment analyses. The heat map demonstrates a different pattern of proteins in the treatment and control groups. The dot plots in Figures 5B and C represent the enrichment of MF and BP, respectively. In the treatment group, MF enrichment included ubiquitin-related proteins, such as ubiquitin ligase, ubiquitin transferase, and ubiquitin-like modifiers, which are associated with the induction of cell death. In contrast, proteins related to microtubule and tubulin binding, as well as structural constituents, were enriched in the MF category of the control group.
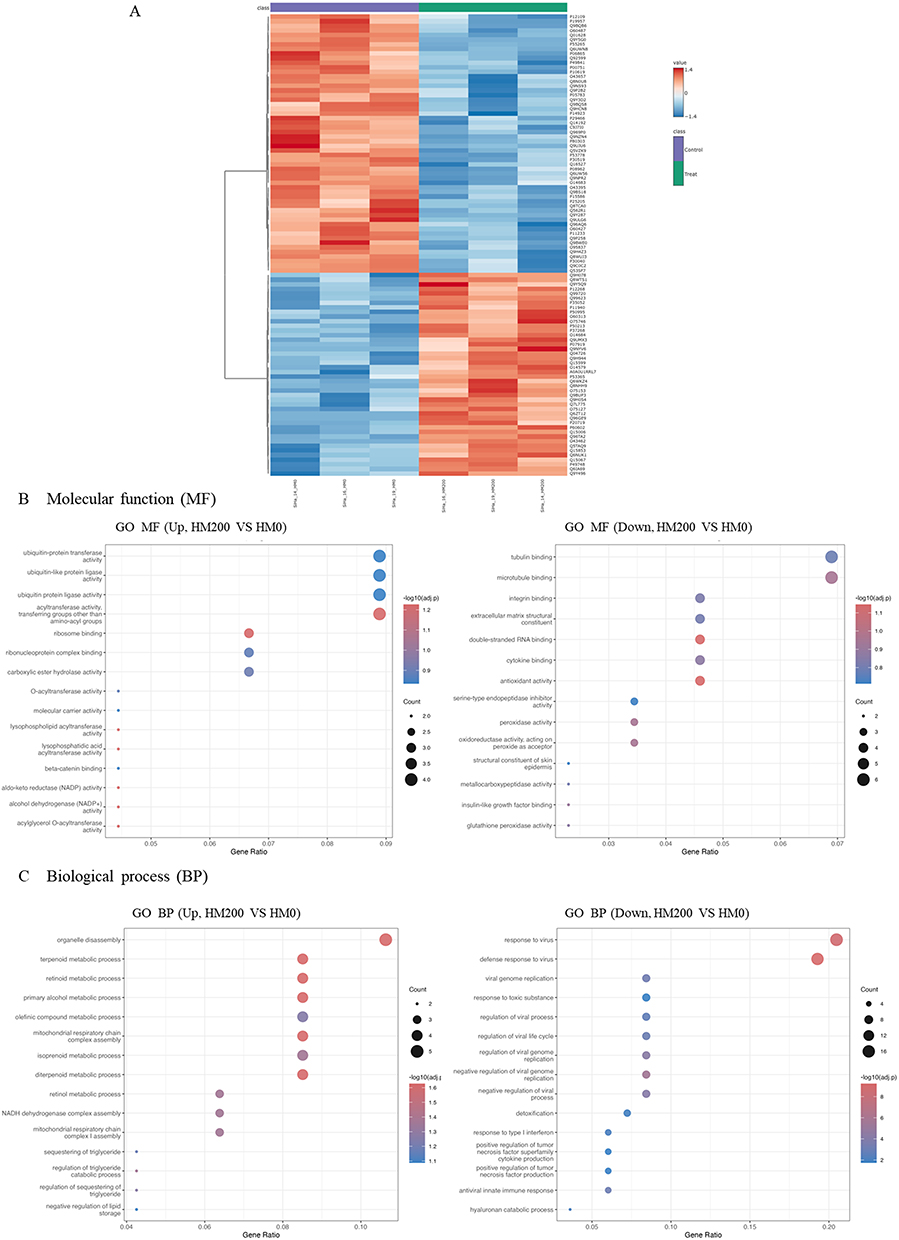 |
Figure 5 Heat map providing intuitive visualization of a proteomic data of control (HM-0) and Halimeda macroloba extracts (HM-200) treatment on cervical cancer spheroid. The top 100 of upregulated and downregulated expression in each group (A). Control group (HM-0) is shown in purple, while treatment group (HM-200) in shown in green. Blue represents low expression (−1.4) and red represents high expression (1.4). Enrichment analysis was performed using the clusterProfiler package. Gene Ontology (GO) enrichment was conducted for molecular function (MF), (B) and biological process (BP), (C) categories. |
Notably, proteins involved in various metabolic pathways, including retinoid, diterpenoid, terpenoid, isoprenoid, and retinol metabolism, were enriched in the BP of the treatment group. Additionally, proteins in the mitochondrial respiratory chain were enriched. This is consistent with mitochondrial disintegration leading to apoptosis and ferroptosis. Conversely, BP enrichment in the control group revealed the presence of virus-related proteins, including response to viruses, defense response to viruses, negative regulation of genome replication, and regulation of viral life cycle processes—notably those associated with human papillomavirus type 16 genome infection in SiHa cells. Table 1 lists 100 differentially expressed proteins in the control and treatment groups.
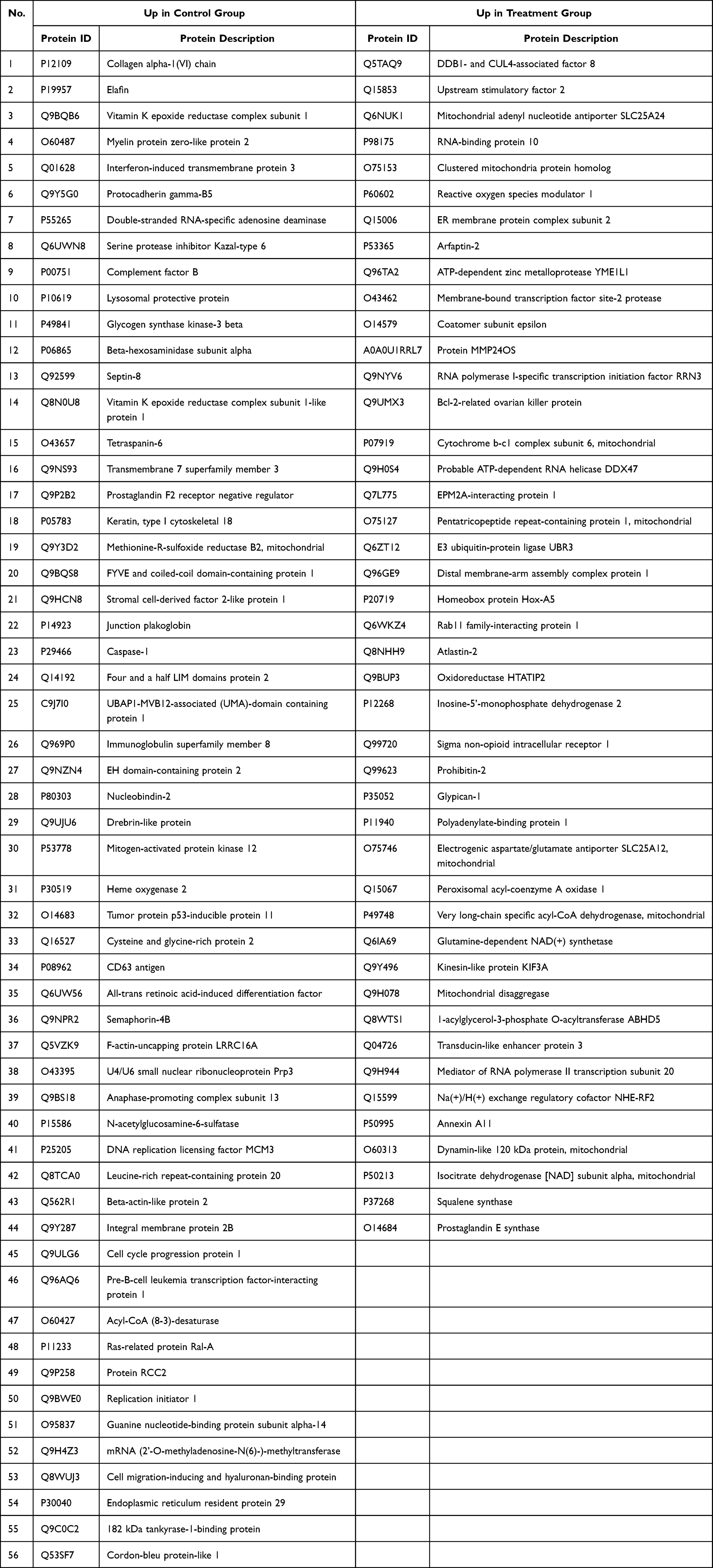 |
Table 1 List of Top 100 Proteins in Heatmap Analysis |
Molecular Mechanism of H. macroloba Extracts in Cervical Cancer Spheroids
The molecular mechanism of H. macroloba (HM) extract has previously been associated with its flavonoid constituents, which exhibit anticancer activities through modulation of multiple signaling pathways involved in cell proliferation, survival, and apoptosis. To investigate the potential molecular effects of HM treatment, the expression of proteins associated with the Akt/mTOR signaling network and apoptotic pathways was evaluated by Western blot analysis (Figure 6 and Supplementary Figure 1). HM treatment resulted in a dose-dependent increase in p-Akt/Akt expression, whereas p-mTOR/mTOR expression was significantly reduced. These findings indicate that HM modulates components of the Akt/mTOR signaling network; however, the observed expression pattern does not support a simple inhibition of the entire pathway and may reflect the involvement of additional regulatory mechanisms. In contrast, the expression levels of apoptosis-related proteins, including Bax, cleaved caspase-3, and cytochrome c, increased in a dose-dependent manner, suggesting activation of the mitochondrial apoptotic pathway. Collectively, these results support the pro-apoptotic activity of HM extract in cervical cancer cells, while further studies are required to elucidate the precise relationship between Akt activation and mTOR suppression.
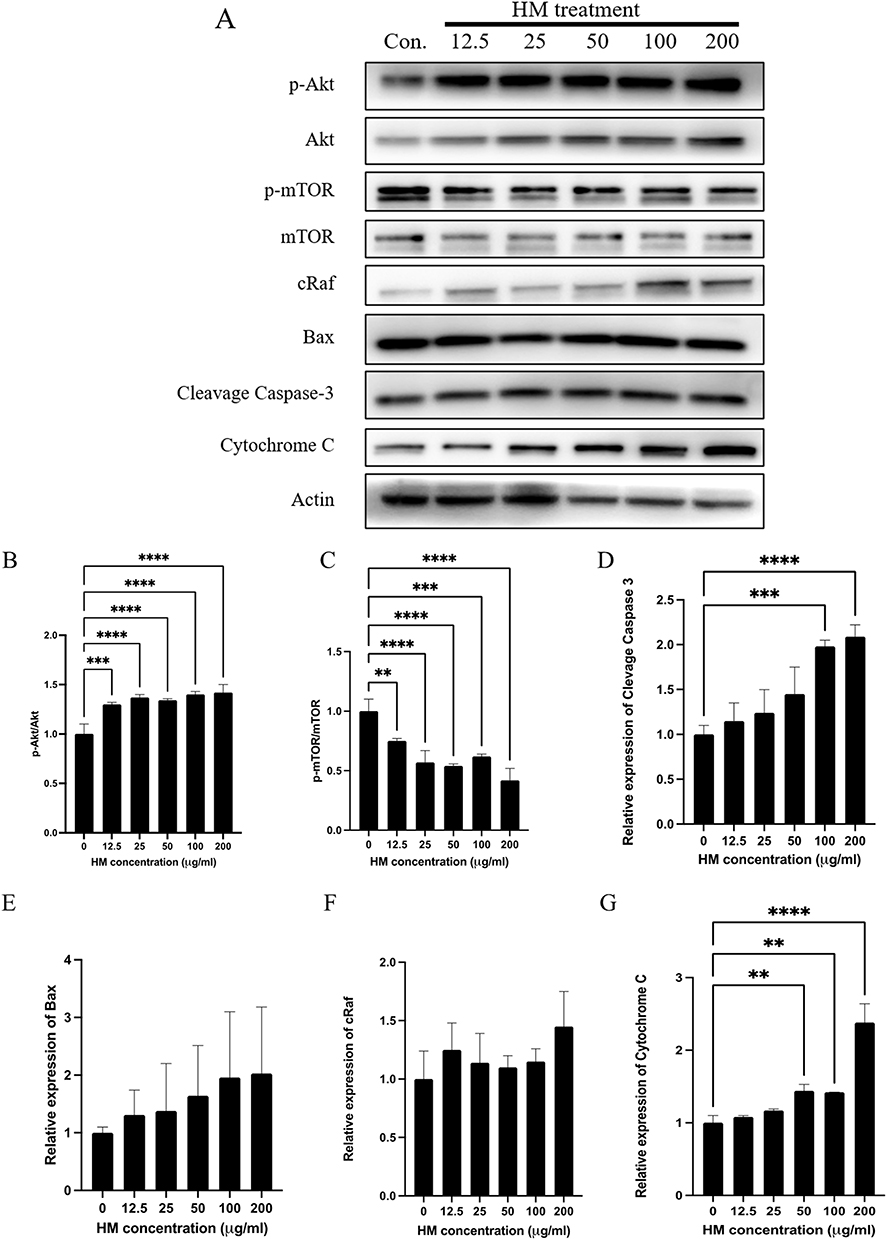 |
Figure 6 Effect of Halimeda macroloba extracts on the Akt, mTOR, and apoptosis pathway. The spheroids were treated with H. macroloba extracts at various concentration for 72 h. After incubation, the spheroids were collected and lysed. (A) The proteins were loaded on SDS-PAGE, and those of interest were examined by Western blot analysis. Actin was used as internal control. The quantitative proteins of interest, including (B) p-Akt/Akt, (C) p-mTOR/mTOR, (D) cleavage-Caspase 3, (E) Bax, (F) cRaf, and (G) Cytochrome C, were normalized to actin band intensity. Data are shown as the mean ± SD of triplicate experiments. Statistical analysis was performed using Student’s t-test. Significance: ** p < 0.01 compared with the control. *** p < 0.001 compared with the control. **** p < 0.0001 compared with the control. |
Discussion
Various phytochemical constituents, including terpenoids, alkaloids, steroids, saponins, phenolic compounds, and flavonoids, have been identified in the crude extracts of H. macroloba. Flavonoids and steroids are considered potential bioactive compounds associated with anticancer mechanisms. The anticancer activity of these metabolites involves the generation of reactive oxygen species (ROS), which leads to mitochondrial damage, caspase activation, and cytochrome c release, all of which ultimately result in apoptosis. Our findings demonstrated that H. macroloba extracts exhibited cytotoxic potential against SiHa cervical cancer spheroid cells. Initially, Cytotoxicity was assessed by measuring spheroid size; however, no significant differences were observed among the treatment groups. This result may be explained by the loose aggregation morphology of the SiHa spheroids. Notably, LIVE/DEAD assays revealed that cell death predominantly occurred in the spheroid core, whereas cells in the outer layer remained viable. This observation is consistent with the structural characteristics of SiHa spheroids, which typically exhibit a flat morphology with a densely packed center and a less compact periphery.14 Based on these findings, we propose that the H. macroloba extract exerts stronger effects under hypoxic conditions than under normoxic conditions. Similar anticancer properties have been reported in related Halimeda species. Methanolic extracts of Halimeda tuna demonstrated anticancer activity against HeLa cervical cancer cells and A549 lung cancer cells.17,18 Additionally, studies on breast, colorectal, and liver cancers have shown the cytotoxic activity of methanolic H. macroloba extracts, supporting their potential antiproliferative effects.19 Furthermore, Halimeda gracilis extract induced cytotoxicity in MDA-MB-231 breast cancer cells through ROS overproduction, mitochondrial dysfunction, and apoptosis.2 Previous studies have also reported that H. macroloba extracts exhibit diverse therapeutic activities, including antioxidant, antiplasmodial, α-glucosidase inhibitory, antimicrobial, antiproliferative, and wound-healing effects.5 Investigations of its antioxidant activity identified flavonoids and steroids as major active constituents.6,10 However, the molecular mechanisms underlying these anticancer effects remain poorly understood. In this study, we investigated the molecular effects of H. macroloba extract on SiHa spheroids while focusing on the Akt/mTOR and apoptotic pathways, which are established targets of its bioactive compounds. Proteomic analysis further revealed enrichment of proteins associated with mitochondrial function and organelle disassembly in the treatment group, suggesting the activation of apoptosis-related pathways and the possible involvement of ferroptosis-associated mechanisms. Proteins uniquely expressed in the H. macroloba extract-treated group were enriched for higher catalytic activity, metabolic processes, and metabolite interconversion enzymes than in the control group. The proteins in the heatmap were highly expressed in the treatment group, including the ROS modulator 1 (P60602 and ROMO1), which plays a role in inducing oxidative DNA damage and oxidative stress.20 This leads to the expression of apoptosis-related proteins, such as the Bcl-2-related ovarian killer protein (Q9UMX), cytochrome b-c1 complex subunit 6, mitochondria (P07919), and E3 ubiquitin-protein ligase UBR3. The key anticancer activity of ROMO1 is its role as a ROS-regulating protein that can increase the activation of c-Jun N-terminal kinase, thereby leading to apoptotic cell death. Enrichment analysis of molecular functions demonstrated a high gene ratio for acyltransferase activity and various ubiquitin ligase and transferase activities in the H. macroloba extract-treated group. Additionally, enrichment analysis of BPs revealed various metabolic processes in the H. macroloba extract treatment group, including retinoids, diterpenoids, terpenoids, and olefinic compounds, which may play a key role in the anticancer activity of cervical cancer spheroids.
Notably, our results revealed a significant downregulation of mTOR accompanied by a dose-dependent increase in apoptotic proteins, supporting the role of both apoptotic and ROS-mediated pathways in the cytotoxic effects of H. macroloba extract.
Conclusion
H. macroloba extract demonstrated significant cytotoxic potential against cervical cancer spheroids, with evidence suggesting the involvement of apoptosis-related pathways and possible ferroptosis-associated mechanisms. The enhanced effects observed in spheroid cultures may be associated with microenvironmental characteristics of the 3D spheroid model, including limited oxygen diffusion, together with proteomic enrichment of mitochondrial dysfunction- and organelle disassembly-related pathways. Consistent with previous reports on related Halimeda species and other cancer models, these findings support the potential of H. macroloba as a source of bioactive compounds with anticancer activity. However, additional studies incorporating specific molecular markers and functional validation are required to confirm the involvement of ferroptosis-related pathways, clarify the underlying cell death mechanisms, and determine the role of hypoxia-related effects Furthermore, isolation and characterization of the active compounds, together with evaluation of their bioavailability and pharmacokinetic properties, as well as validation in preclinical in vivo models, are necessary before clinical translation can be considered. This study is also limited by the evaluation of only short-term treatment responses in the 3D spheroid model; therefore, further investigation of long-term effects is warranted. In addition, while L929 and HEK-293 cells were used as representative non-cancerous controls to evaluate general cytotoxicity and safety, future studies should include immortalized normal cervical epithelial cells and corresponding 3D spheroid models to further assess tissue-specific selectivity and compare the effects of H. macroloba extract on normal and malignant cervical tissues within a more physiologically relevant microenvironment. Collectively, this study provides preliminary evidence supporting the anticancer potential of H. macroloba extracts against cervical cancer.
Acknowledgments
The authors gratefully acknowledge the Seaweed and Seagrass Research Unit team, Prince of Songkla University, who assisted us with fieldwork on H. macroloba extract preparation from Parinuch Chumkaew, Faculty of Science and Industrial Technology, Prince of Songkla University, Surat Thani Campus, and Surat Thani. The authors acknowledge the use of an AI platform (ChatGPT) for grammar checks and English language editing.
Funding
This research was funded by the National Science and Research and Innovation Fund (NSRF), Prince of Songkla University (SCI6801020S) and the Postdoctoral Fellowship Program from the Faculty of Medicine, Prince of Songkla University.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Hawas UW, Husain S, El-Kassem LTA. Biochemical composition and biological properties of some red and green algae from the Saudi Red Sea coast. Thalassas. 2025;41(2):104. doi:10.1007/s41208-025-00856-w
2. Ramalingam V, Narendra Kumar N, Harshavardhan M, et al. Chemical profiling of marine seaweed Halimeda gracilis using UPLC-ESI-Q-TOF-MSE and evaluation of anticancer activity targeting PI3K/AKT and intrinsic apoptosis signaling pathway. Food Res Int. 2022;157:111394. doi:10.1016/j.foodres.2022.111394
3. Cremen MCM, Huisman JM, Marcelino VR, Verbruggen H. Taxonomic revision of Halimeda (Bryopsidales, Chlorophyta) in south-western Australia. Aust Syst Bot. 2016;29(1):41–14. doi:10.1071/SB15043
4. Kooistra WHCF, Coppejans EGG, Payri C. Molecular systematics, historical ecology, and phylogeography of Halimeda (Bryopsidales). Mol Phylogenet Evol. 2002;24(1):121–138. doi:10.1016/S1055-7903(02)00221-X
5. Saber AA, Rashedy SH, Rushdi MI, et al. Insights into the phytochemical and pharmacological natural products of the green macroalga Halimeda (Chlorophyta). Nat Prod Commun. 2025;20(3). doi:10.1177/1934578X251331137
6. Widowati R, Handayani S, Suprihatin ILR. Phytochemicals and antioxidant of methanol extract of Gracilaria salicornia, Halimeda gracilis, Halimeda macroloba, and Hypnea asperi from Tidung Island coastal region. Eur J Mol Clin Med. 2021;8:1–12.
7. Navakanitworakul R, Nittayaboon K, Chumkaew P, Phetcharat S, Mayakun J. Phytochemical screening and cytotoxic activities of Enhalus acoroides (L.f.) Royle and Halimeda macroloba Decaisne on cervical cancer cell lines. Trop J Pharm Res. 2025;23(12):2053–2057. doi:10.4314/tjpr.v23i12.10
8. Mayakun J, Prathep A. Calcium carbonate productivity by Halimeda macroloba in the tropical intertidal ecosystem: the significant contributor to global carbonate budgets. Phycol Res. 2019;67(2):94–101. doi:10.1111/pre.12361
9. Mayakun J, Liao CP, Liu SL. The standing stock and CaCO 3 contribution of Halimeda macroloba in the tropical seagrass-dominated ecosystem in Dongsha Island, the main island of Dongsha Atoll, South China Sea. J Mar Biol Assoc UK. 2020;100(8):1219–1227. doi:10.1017/S0025315420001113
10. Nazarudin MF, Yasin ISM, Mazli NAIN, et al. Preliminary screening of antioxidant and cytotoxic potential of green seaweed, Halimeda opuntia (Linnaeus) Lamouroux. Saudi J Biol Sci. 2022;29(4):2698–2705. doi:10.1016/j.sjbs.2021.12.066
11. Wongpratate M, Bumrungthai S. Cervical cancer in Thailand: 2023 update. Obstet Gynecol Sci. 2024;67(3):261–269. doi:10.5468/ogs.23277
12. Tewari KS. Cervical cancer. N Engl J Med. 2025;392(1):56–71. doi:10.1056/NEJMra2404457
13. Rocha DHA, AML Seca, Pinto DCGA. Seaweed secondary metabolites in vitro and in vivo anticancer activity. Mar Drugs. 2018;16:410. doi:10.3390/md16110410
14. Molika P, Leetanaporn K, Chiangjong W, Choochuen P, Navakanitworakul R. Proteomic analysis reveals cadherin, actin, and focal adhesion molecule-mediated formation of cervical cancer spheroids. Cells. 2024;13(23):2004. doi:10.3390/cells13232004
15. Potriquet J, Laohaviroj M, Bethony JM, Mulvenna J. A modified FASP protocol for high-throughput preparation of protein samples for mass spectrometry. PLoS One. 2017;12(7):e0175967. doi:10.1371/journal.pone.0175967
16. Demichev V, Messner CB, Vernardis SI, Lilley KS, Ralser M. DIA-NN: neural networks and interference correction enable deep proteome coverage in high throughput. Nat Meth. 2020;17(1):41–44. doi:10.1038/s41592-019-0638-x
17. Gazali M, Husni A, Sukmadewi AP, et al. Anticancer activity of marine macroalgae Halimeda tuna from Aceh waters against cervical cancer cells. J Fish Environ. 2024;48:120–131.
18. Husni A, Gazali M, Nurjanah N, Syafitri R, Matin A, Zuriat Z. Cytotoxic activity of green seaweed Halimeda tuna methanolic extract against lung cancer cells. J Multidiscip Appl Nat Sci. 2024;4(1):16–29. doi:10.47352/jmans.2774-3047.172
19. Nazarudin MF, Isha A, Mastuki SN, et al. Chemical composition and evaluation of the α-glucosidase inhibitory and cytotoxic properties of marine algae Ulva intestinalis, Halimeda macroloba, and Sargassum ilicifolium. Evid Based Complement Alternat Med 2020;2020(1):2753945. doi:10.1155/2020/2753945
20. Amini MA, Talebi SS, Karimi J. Reactive oxygen species modulator 1 (ROMO1), a new potential target for cancer diagnosis and treatment. Chonnam Med J. 2019;55(3):136–143. doi:10.4068/cmj.2019.55.3.136
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.