Back to Journals » International Journal of Nanomedicine » Volume 16
CpG-Based Nanovaccines for Cancer Immunotherapy
Authors Chen W, Jiang M, Yu W, Xu Z, Liu X, Jia Q, Guan X, Zhang W ![]()
Received 26 April 2021
Accepted for publication 14 July 2021
Published 5 August 2021 Volume 2021:16 Pages 5281—5299
DOI https://doi.org/10.2147/IJN.S317626
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Ebrahim Mostafavi
Wenqiang Chen,1 Mingxia Jiang,1 Wenjing Yu,1 Zhiwei Xu,1 Xinyue Liu,1 Qingmiao Jia,1 Xiuwen Guan,1– 3 Weifen Zhang1– 3
1College of Pharmacy, Weifang Medical University, Weifang, 261053, People’s Republic of China; 2Collaborative Innovation Center for Target Drug Delivery System, Weifang Medical University, Weifang, 261053, People’s Republic of China; 3Shandong Engineering Research Center for Smart Materials and Regenerative Medicine, Weifang Medical University, Weifang, 261053, People’s Republic of China
Correspondence: Xiuwen Guan; Weifen Zhang Tel +86-536-8462490
Email [email protected]; [email protected]
Abstract: Cancer has been a serious health hazard to the people all over the world with its high incidence and horrible mortality. In recent years, tumor vaccines in immunotherapy have become a hotspot in cancer therapy due to their many practical advantages and good therapeutic potentials. Among the various vaccines, nanovaccine utilized nanoparticles (NPs) as the carrier and/or adjuvant has presented significant therapeutic effect in cancer treatment. For tumor nanovaccines, unmethylated cytosine-phosphate-guanine oligodeoxynucleotide (CpG ODN) is a commonly used adjuvant. It has been reported that CpG ODN was the most effective immune stimulant among the currently known adjuvants. It could be recognized by toll-like receptor 9 (TLR9) to activate humoral and cellular immunity for preventing or treating cancer. In this review, the topic of CpG-based nanovaccines for cancer immunotherapy will be focused. The types and properties of different CpG will be introduced in detail first, and then some representative tumor nanovaccines will be reviewed according to the diverse loading modes of CpG, such as electrostatic adsorption, covalent bonding, hydrophilic and hydrophobic interaction, and DNA self-assembly, for summarizing the current progress of CpG-based tumor nanovaccines. Finally, the challenges and future perspectives will be discussed. It is hoped that this review will provide valuable references for the development of nanovaccines in cancer immunotherapy.
Keywords: cancer immunotherapy, nanovaccine, nanoparticle, adjuvant, CpG
Introduction
Malignant tumors have been commonly occurring diseases that seriously threatened human health and life with high morbidity and mortality.1 As cancer cells are originated from normal body cells, they are difficult to be monitored and completely eliminated at an early stage in the body, and these cells are easy to escape the systemic immune surveillance for rampantly developing into malignant tumors and further metastasizing to other normal tissues.2,3 Currently, the treatment of cancer is still mainly based on the traditional methods, such as surgical treatment, chemotherapy, and radiotherapy. However, according to the current situation of the high incidence and dreadful mortality of cancer in clinic, these traditional therapies are not sufficient to defeat the malignancies.4,5 In recent years, immunotherapy has become another effective anti-tumor tactic.6–10 Cancer immunotherapy can reinitiate and maintain the tumor-immune cycle, restore the body’s normal anti-tumor immune response to achieve the purpose of controlling or completely eliminating tumors.6,11–13
Currently, cancer immunotherapy mainly includes adoptive lymphocyte therapy, biological targeting antibody, tumor vaccine, immune checkpoint blockade, etc.14 Among them, tumor vaccine has become one of the research hotspots of immunotherapy, and some protein and peptide-based subunit antigens have been widely developed for tumor vaccine due to their high safety performance, easy manufacture and convenient storage.15–18 However, this type of vaccine usually has poor immunogenicity, unreliable stability, and impressionable degradation in the body.19 It can only induce a short-term immune response and cannot achieve an effective therapeutic effect.20 Therefore, new generations of tumor nanovaccines which utilized nanoparticles (NPs) as the carriers and/or adjuvants had been proposed.21 In recent years, the continuous and in-depth development of nanotechnology has brought new dawn for people who suffered from intractable diseases.22,23 Nanomaterials have been widely utilized as drug delivery carriers to effectively address the multifarious problems that occurred during the in vivo drug transportation, such as protect drugs from degradation, prolong systemic circulation, regulate drugs to distribute at the lesion site, promote drugs to cross the physiological barriers, facilitate drugs to penetrate the biological membranes, control drug release and other issues.24,25 Especially in the field of cancer therapy, nanomaterials have been extensively used to achieve effective and precise treatment.26,27 For cancer immunotherapy, in contrast to subunit vaccines, nanovaccines have many practical advantages. Nanovaccines can be designed with diverse features, such as effective penetration through the tissue barriers, high cellular uptake in antigen-presenting cells (APCs), reliable protection and efficient delivery for antigens, also tunable antigen release and effective cross-presentation, and so on. On the other hand, nanovaccines can co-deliver the antigens with the adjuvants to remarkably augment the immunogenicity, induce robust innate and adaptive immunity and potentiate antigen-specific immune responses. The antigens and adjuvants wrapped in the nanocarriers can be well protected to prevent the degradation and improve their stability.28 In addition, nanovaccines can also achieve targeting function to deliver the antigens and adjuvants to the specific cells.
Generally, nanovaccine is composed of antigen, adjuvant, and the nanocarrier. Among them, adjuvant, as an immune enhancer, plays a crucial role in nanovaccines, which can effectively stimulate and activate immune cells to boost the immune responses.28,29 And according to the research findings, unmethylated cytosine-phosphate-guanine (CpG) is a very potential immune adjuvant, which can stimulate a variety of immune cells to trigger strong innate and adaptive immune responses, and this powerful immune stimulation ability is closely related to the special structure of CpG.30 CpG oligodeoxynucleotide (ODN) is a short single-stranded synthetic DNA, its sequence consists of a cytosine triphosphate deoxynucleotide (“C”) and a guanine triphosphate deoxynucleotide (“G”). The “p” represents the phosphodiester link between the consecutive nucleotides. C is located at the 5ʹ end and G is located at the 3ʹ end. The CpG sequence is widely distributed in the DNA of bacteria and other prokaryotes, their DNA sequences contain a high frequency of CpG dinucleoside (about 1/16), which can stimulate the lymphocytes of mice and humans. However, CpG sequence is less distributed in mammals, and the frequency of CpG dinucleosides is only about 1/50, and most of them are methylated in an inhibited status and unable to stimulate lymphocytes.31 As unmethylated CpG motifs are ubiquitous in the genomes of bacterias and other prokaryotes, but not in vertebrate genomes. When this DNA sequences appear in mammals, CpG motifs will be recognized as pathogen-related molecular patterns (PAMPs). Water-soluble CpG is phagocytized by lymphocytes, like plasmacytoid dendritic cells (pDCs) and B cells, and then CpG enters the endosome, recognized by the pattern recognition receptor (PRR) Toll-like receptor 9 (TLR9), and further induces innate and acquired immune responses.32
CpG DNA can be derived from some lower organisms in nature, and also can be obtained by artificially synthesizing with specific sequences. CpG ODN is a synthetic short single-stranded DNA molecule which can be flexibly synthesized. Most strikingly, studies have shown that artificially synthesized oligonucleotide sequences containing unmethylated CpG motifs have more advantages. CpG ODN can replace bacterial CpG DNA to realize an extremely similar immunostimulatory effect for directly activating B cells and pDCs to induce innate and adaptive immune response as shown in Figure 1. Therefore, they can also be used as TLR9 agonists. In addition, CpG ODN has better stability and longer half-life than CpG of lower organisms such as bacteria. CpG ODN has a partial or complete phosphorothioate (PS) backbone, and the PS modification can protect CpG ODN from nuclease degradation in the body and enhance its uptake by cells.33 While bacterial genomic DNA is a natural phosphodiester backbone, which is unstable and easily degraded by nucleases in vivo. According to some reports, five types of CpG ODN have been identified (Type A, B, C, P, and S) based on the sequence, secondary structure and the effect on human peripheral blood mononuclear cells (PBMC), especially B cells and pDCs. Among them, Type A, Type B and Type C CpG ODN are particularly effective in stimulating human immune cells.34–36 In this review, the above three types of CpG ODN will be introduced, and we will also summarize some representative CpG-based nanovaccines for cancer immunotherapy according to the CpG loading pattern. We hope that this review will provide valuable references for the development of nanovaccines in cancer immunotherapy.
 |
Figure 1 The mechanism of CpG ODN inducing innate and adaptive immune response. |
The Types and Properties of CpG
Despite CpG ODNs of Type A, Type B and Type C are particularly effective in stimulating human immune cells, the patterns and degrees of the induced immune responses after they stimulating immune cells are significantly different due to their distinguishing structures.32,36 The comparison between Type A, Type B and Type C CpG ODNs was summarized in Table 1. The typical sequence of these three CpG types is different, and their unique structural features further contribute to the diverse immunomodulatory activity.
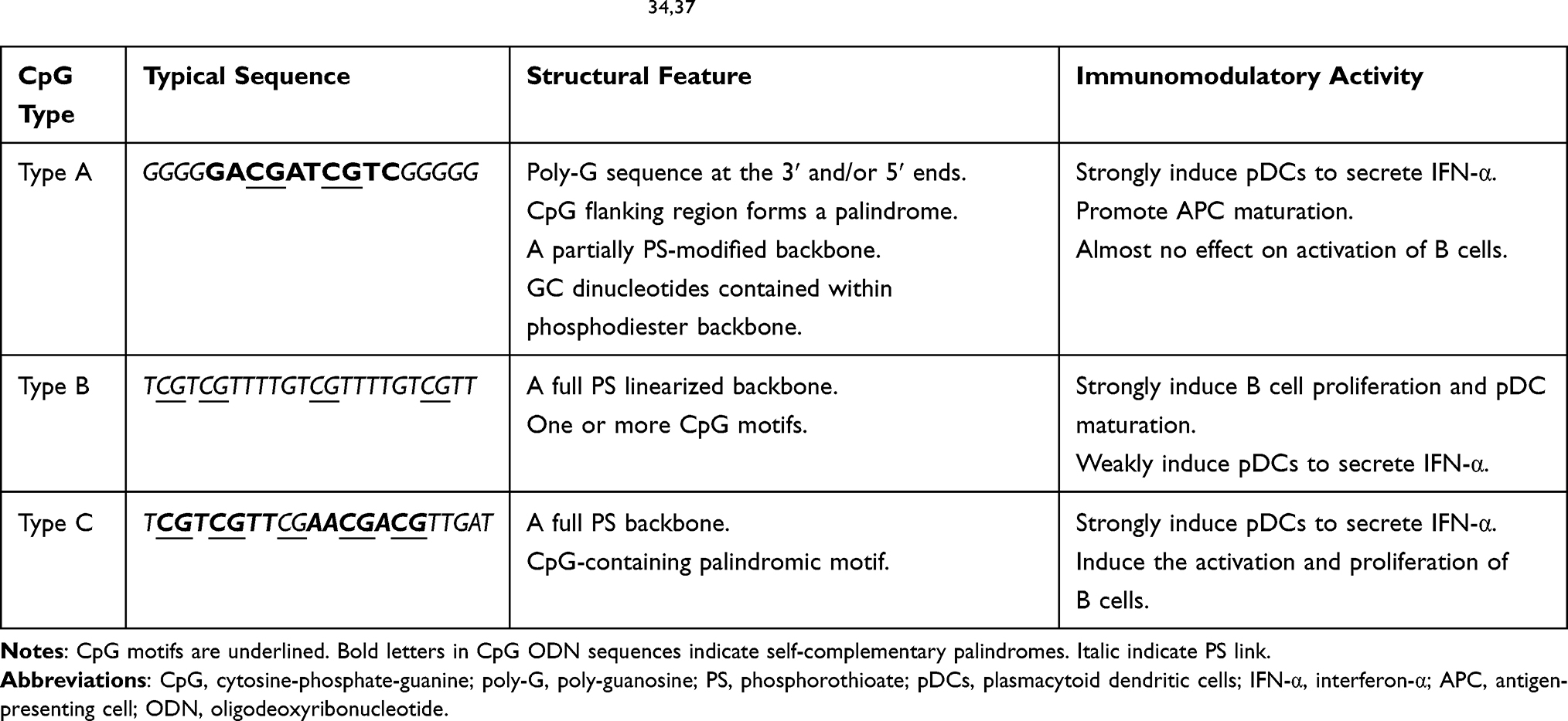 |
Type A CpG
Type A CpG ODN (also known as CpG-D), consists of partially PS modified phosphorothioate-phosphodiester-phosphorothioate chimeric backbone,38 contains a single hexameric purine-pyrimidine-CG-purine-pyrimidine CpG motif,37,39 which is located on the phosphodiester backbone.38,40 The flanking palindrome sequence of the CpG motif containing the GC dinucleotide is self-complementary base pairs and forms a stem-loop structure.37,39 The 3ʹ and 5ʹ flanking regions are located on the PS backbone,40 and there are three or more consecutive poly-guanosine (poly-G) sequences at the 3ʹ and/or 5ʹ ends. The PS chain and the special structure of poly-G can effectively protect Type A CpG ODNs from nuclease degradation and effectively prolong their half-life. Additionally, the poly-G tails on a Type A CpG ODN molecule can interact and form stable and complex G-tetrad structure.34,37,41 Type A CpG ODN can effectively activate natural killer cells (NKs),38 by directly inducing pDCs to secrete a large amount of tumor necrosis factor-α (TNF-α) and interferon-α (IFN-α).42–44 What’s more, they can also promote the maturation of APCs.37,39,42 But Type A CpG ODNs almost have no effect on activation of B cells or secretion of Th1-type cytokines.34,45–48
Since the discovery of Type A CpG, the practical application research of this kind of CpG has also been intensified. The production of “cytokine storm” can cause fatal endotoxin shock. Studies have found that Type A CpG played an important role in the treatment of endotoxin shock. Type A CpG can effectively induce the production of platelet-activating factor (PAF) hydrolase, which greatly improves the endotoxic shock induced by lipopolysaccharide and increases the survival rate to a certain extent.49 In contrast, Type B CpG does not have such a good therapeutic effect, and may even aggravate the situation.50 Additionally, Type A CpG also plays an active role in the treatment of experimental autoimmune encephalomyelitis (EAE) disease. Crooks et al found that Type A CpG could significantly reduce the severity of EAE and alleviate the acute phase of EAE disease, while also brought some other positive therapeutic effects.35 And studies have shown that the poly-G tail of Type A CpG had anti-tumor activity with its special structure. Kobayashi et al found that Type A CpG with the poly-G tail structure could induce the phosphorylation of lymphocyte-specific protein tyrosine kinases, enhance the secretion of interleukin-2 (IL-2) and the proliferation of CD8+ T cells, thereby inhibiting the development of tumor cells to achieve a good anti-tumor effect.51 However, the special poly-G structure of Type A CpG can be easily formed into multimer in solution, which also causes certain difficulties for the clinical application. In a recent study, Type A CpG was encapsulated in virus-like particles (VLP), which could effectively combat tumors through intratumoral delivery. Lemke-Miltner et al used Qβ phage capsid protein and Type A CpG to assemble CMP-001 (a new VLP) in a certain proportion, and found that CMP-001 had a good anti-tumor effect. Through intratumoral treatment, it effectively enhanced the infiltration of T cells and DCs into tumors, and induced prominent tumor regression.52 These studies had established theoretical and experimental basis for the potential applications of Type A CpG in the future.
Type B CpG
Type B CpG ODN (also known as CpG-K) is composed of a linearized oligonucleotide sequence containing one or more CpG motifs and located on the fully PS-modified backbone.34 Similarly, the backbone is phosphorylated by sulfuric acid, which enhances the ability to resist nuclease degradation and prolongs the half-life of systemic circulation.48,53 Type B CpG ODNs are weak inducers of IFN-α, but strong stimulators for B cells, which can induce B cells and monocytes to proliferate and secrete interleukin-6 (IL-6), thereby promoting the production of immunoglobulin M (IgM).32,34,54 In addition, they are also able to present good anti-tumor activity.55 Malignant tumors can create an immunosuppressive environment that maintains T cell tolerance and represses the anti-tumor immune responses.34 Type B CpG ODNs are strong inducers for Th1-type immune response, they can promote the production of Th1 cytokines (such as TNF-α) and induce the maturation of pDCs.39,46,48 Besides, Type B CpG ODN can up-regulate the expression of costimulatory molecules to relieve the immunosuppression state of the tumor microenvironment (TME) and convert the T cells from anergy status into awakening cytotoxic T lymphocytes (CTLs) to fight against tumors.34,40
Since the discovery of the powerful immunostimulating activity of Type B CpG ODN, it has been widely used in preclinical and clinical research in the field of disease treatment.56 For example, Type B CpG ODN has been developed as an immune adjuvant for hepatitis B virus (HBV) vaccine. Compared with the original HBV vaccine (hepatitis B surface antigen + alum), the HBV vaccine loaded with Type B CpG ODN has better immunostimulation ability to produce protective antibodies faster for achieving better preventive effect, and the vaccine is already in the advanced stage of clinical trials, which has good potential to replace the original HBV vaccine.57 Liang et al found that Type B CpG was helpful for the treatment of human chronic lymphocytic leukemia (CLL). It could induce the apoptosis of the CLL-related B cells by mediating the TLR9 signaling pathway. In mouse experiments, Type B CpG could reduce the number of leukemia cells in the body of mice.58 Furthermore, in recent years of research, Pul et al described that Type B CpG played an important role in combating the immunesuppression of sentinel lymph nodes (SLNs) induced by breast cancer. For inhibiting the JAK2/STAT3 pathway, Type B CpG could reactivate a subset of lymph node-resident (LNR) dendritic cells (DCs) and restore Th1-type anti-tumor response.59 These studies reflected that Type B CpG had occupied an important position in the field of cancer immunotherapy.
Type C CpG
Type C CpG ODN is a kind of amazing CpG sequence, which seems to simultaneously have the immunostimulatory ability of both Type A and Type B CpG ODNs.41,46 Therefore, Type C CpG ODN has been extensively used and developed due to its strong immunostimulatory activity. It has been reported that the application of Type C CpG ODN in cancer treatment has achieved good therapeutic effects.43 Type C CpG ODN has such a powerful stimulating ability mainly due to the special structure which combines the structural characteristics of Type A and Type B CpG ODNs: (1) It has a complete PS backbone similar to Type B CpG ODN, which prevents the degradation by nucleases and increases the activity of CpG.48 And it has a representative Type B CpG ODN motif (like “TCGTCG” or “GTCGTT”) at the 5ʹ end.38 (2) It has a stimulatory palindrome with CpG motif similar to the Type A CpG ODN at the 3ʹ end, which can form a stem-loop or dimer structure.34,43,48 Remarkably, Type C CpG ODN does not contain the poly-G motifs.38
Studies have shown that Type C CpG ODN seemed to be potent inducer of Th1-type cytokines, which could induce strong Th1-type immune response by activating TLR9.60,61 Specifically, Type C CpG ODN induces pDCs to secrete cytokine IL-12 to activate NKs. And similar to Type A CpG ODN, Type C CpG can also induce pDCs to significantly secrete cytokine IFN-α.62 Additionally, Type C CpG ODNs are able to effectively stimulate the activation and proliferation of B cells, thereby promoting the secretion of immunological active substances such as IL-6, IgM, and TNF-α.43,62 It was observed that Type C CpG ODN not only could effectively activate innate immunity, but also acquired immunity.48,61 In the past decades, many studies have been conducted on the Type C CpG ODN. For instance, mitochondrial DNA (mtDNA) belongs to the risk-related molecular model, and plays an important role in the development of post-traumatic inflammatory response and the transduction of antiviral signals. Studies have found that Type C CpG could quickly induce lymphocytes to release mtDNA, which was a resilient reticulum structure. The mtDNA network could act as a signal for rapid response and play a synergistic effect with cytokines and natural antibodies in the antiviral process.63 Mathur et al demonstrated that Type C CpG could reduce TLR4-induced acute inflammatory cardiac dysfunction by activating TLR9, and also be likely to have the ability to prevent cardiac dysfunction.64 On the basis that CpG ODN might have the ability to promote wound healing, Li et al observed the effect of a new Type C CpG ODN designed by their group on wound healing through experiments, which further proved that Type C CpG ODN had the ability to accelerate wound healing by regulating fibroblasts and activating the immune response.65 Furthermore, Sorski et al found that Type C CpG ODN might also have advantages in reducing the side effects of surgery.61 Since Type C CpG ODN has specific structure and prominent immunostimulating ability, it will have broad potential applications in the future.
CpG-Based Nanovaccines for Cancer Immunotherapy
It was reported that CpG ODN was considered to be new type of immune adjuvant with ideal immune stimulation and also the most effective immunostimulatory adjuvant known till now.37,66–72 Since CpG ODN has the ability to initiate immune activation and destroy immunosuppression in the TME, it has been applied to treat various cancers in preclinical and clinical studies.56 Nevertheless, compared with CpG ODN loaded in nanovaccines, the immune effect of free CpG ODN in clinical research did seem to be discouraging. It is well known that the cell membranes and CpG ODNs are generally negatively charged, which have a high probability of causing electrostatic repulsion to make it difficult for CpG ODN being taken up by the cells.73 In addition, there are also other problems: although PS-modified CpG is more resistant to nucleases than the natural CpG containing phosphodiester, the slightly prolonged half-life of the PS-modified CpG is still difficult to achieve an effective action time for the expected immunostimulating effect. Therefore, it is challenging to apply free CpG in vivo for the prevention and treatment of related diseases.73,74 Over the past few years, with the continuous development of nanomaterials, the above-mentioned problems could been solved by carrying CpG ODN in nanoparticles (NPs). These CpG-loaded nanovaccines were proven to have considerable promise and great advantages in cancer immunotherapy, such as prolonged half-life, enhanced delivery to APCs and lymph nodes (LNs), improved immune responses and so on,75 which has also brought a new dawn for cancer treatment.76 Free CpG ODN and CpG ODN-loaded nanovaccines have been tested in many clinical trials. The results shown that nanovaccine-based CpG ODN had higher immunostimulatory ability than free CpG ODN, because the bound CpG ODN was more stable in the body and had a longer half-life, and increased the chance to be phagocytized by the immune cells. The nanovaccines loaded with CpG ODN can efficiently play the role of tumor vaccine and maximally assist CpG ODN exerting the adjuvant function, thereby achieving effective cancer prevention and treatment. It thus would appear that there would be more and more researches on CpG-based nanovaccines for the treatment of various cancers.
It is worth noting that nanovaccines using Type B CpG ODN as adjuvants accounted for the majority of the studies for cancer immunotherapy.36 As a matter of fact, this is not surprising since Type B CpG ODNs have great anti-tumor activity. Secondly, Type C CpG ODN is also used as adjuvants for tumor nanovaccines. As described previously, Type C CpG has a special structure and immunostimulatory activity, it exhibits the activity of both Type A and Type B CpG ODNs, and has good prospect in the prevention and treatment of diseases.77 Nevertheless, since Type C CpG ODNs have been freshly studied in recent years, the specific process of immune response caused by Type C CpG needs to be gradually explored. As a consequence, in contrast with Type B CpG, there are relatively few studies using Type C CpG ODN as adjuvants for tumor nanovaccines. Unfortunately, the applied research on Type A CpG is minimal, due to the poly-G tail of Type A CpG is easy to form multimer in solution, in addition, it has almost no effect on the secretion of Th1-type cytokines. These above-mentioned factors have incurred many obstacles for the applications of Type A CpG ODNs.48,78
The CpG ODNs play an important role in tumor nanovaccines. Specifically, these CpG ODNs can be used to enhance the immune response, they can induce a strong immunostimulation and promote the secretion of Th1-type cytokines and the expression of co-stimulatory molecules on the surface of B cells and pDCs, and change the tolerance of T cells in the TME to achieve anti-tumor effects.40 In the following content, we will review some representative CpG-based nanovaccines for cancer immunotherapy according to the loading modes of CpG in nanovaccines, such as electrostatic adsorption, covalent bonding, hydrophilic and hydrophobic interaction, and DNA self-assembly.
CpG Loaded in Nanovaccines by Electrostatic Adsorption
It is well recognized that CpG ODN is negatively charged, so suitable positively charged NP carriers can be constructed to introduce CpG by electrostatic adsorption. Moreover, CpG loading by electrostatic adsorption usually involves convenient process and without excess chemical-modification, which can well preserve the immune activities of the adjuvants. For example, Liu et al had prepared poly(ethylene glycol)-b-poly(2-amino ethyl methacrylate)-b-poly(2-(hexamethylene imino) ethyl methacrylate-co-2-(dibutyl amino) ethyl methacrylate) (EAASc) amphiphilic cationic copolymer assembled NPs in their study. As the carrier was positively charged, the negatively charged CpG ODN and antigen peptide E75 (a T-cell epitope of HER2 protein from breast cancer cells) could be adsorbed in the nanocarrier through electrostatic interaction. The tumor nanovaccine was successfully constructed, and the nanovaccines significantly induced a strong immunostimulation response to promote the maturation of DCs and trigger an effective T cell activation, thereby achieving efficient anti-tumor immune effect.79 Guan et al developed a simple and effective antigen and adjuvant co-delivery nanovaccine that used positively charged polyethyleneimine (PEI) to attract negatively charged model antigen ovalbumin (OVA) and CpG through electrostatic interaction. The nanovaccine could be effectively phagocytized by DCs with enhanced uptake, and the PEI assisted the endosomal escape and promoted the cross-presentation of the antigen.16 Similarly, Kim et al designed a nanoplatform based on small lipid nanoparticle (SLNP), including a cationic molecule composed of mono arginine and cholesterol (MA-Chol). The MA-Chol could form complexes with negatively charged CpG through electrostatic adsorption, and finally the OVAPEP (a MHC I-restricted epitope of OVA) was incorporated into the nanoplatform by disulfide bond to successfully construct the nanovaccines. And according to the experimental data, the obtained nanovaccines had immensely effective anti-tumor activity in the prevention and treatment of E.G7 (mouse T lymphoma cell) tumor models. The combination of the small nanovaccines with PD-1 antibody further produced a better therapeutic effect, effectively inhibiting the recurrence of the tumors through a reasonable sequence of treatment.80 Shi et al constructed a peptide nanocomposite vaccine, which was formed by the electrostatic interaction between cell-penetrating peptide (CPP)-conjugated three melanoma-specific and MHC I-restricted epitopes and CpG. Their positively charged CPP R8 (RRRRRRRR)-conjugated derivatives could be combined with negatively charged CpG through electrostatic interaction, leading to spontaneous formation of nanocomposite vaccine. The nanovaccine complex could effectively migrate to the LNs to cause strong effector memory CD8+ T cells and Th1 immune responses (Figure 2).81 Ruiz-de-Angulo et al developed a micro-dose lipid-encapsulated magnetite NP vaccine and used radioactive 67Ga to mark the surface of the nanovaccine. By selecting the combination of different lipids to fill the micelles containing magnetite, when the cationic lipid DOTAP (1,2-dioleoyl-3-trimethylammonium-propane) was added to the mixture of PEGylated phospholipids (PEG PLs), the micelles filled with ionic polymer were positively charged. The negatively charged CpG ODNs were loaded in the micelles through electrostatic interaction. These nanovaccines could be accurately delivered into DCs in LNs, and achieved high-sensitivity tracking in vivo.82
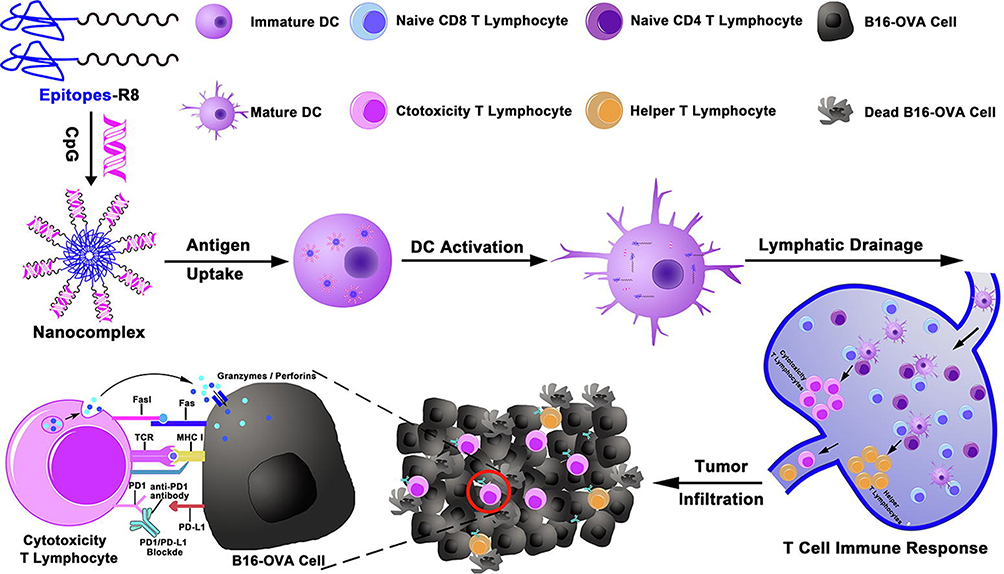 |
Figure 2 Schematic of the nanocomplex vaccine and specific anti-tumor T cell immune response. The nanovaccine was formed by co-assembly of the adjuvant CpG and the cationic Epitopes-R8. Reprinted from Chemical Engineering Journal, 399, Shi X, Song H, Wang C, et al, Co-assembled and self-delivered epitope/CpG nanocomplex vaccine augments peptide immunogenicity for cancer immunotherapy, 125854, Copyright 2020, with permission from Elsevier.81 |
Furthermore, it was found that as a commonly used nanocarrier, mesoporous silica nanoparticles (MSNs) had obvious advantages for being easily modified to be positively charged, so that CpG could be better loaded in MSNs through electrostatic interactions.83 Such as Liu et al firstly aminated the MSN surface, so that the acetylated peptide TY (a DC-targeting peptide) was attached to the MSN surface by covalent interaction. The aminated MSN-TY had a higher potential and contained some positive amino groups on the surface, which could attract CpG and OVA through electrostatic interaction, thereby forming a nanovaccine that precisely targeting DCs to effectively promote the maturation and activation of DCs, and facilitating the secretion of some cytokines by T cells to inhibit tumor growth for prolonging the survival period of the mice.83 Apart from the above studies which constructed the nanocarriers by external materials. Notably, some studies have reported that directly using OVA to construct nanocarriers through self-crosslinking for simultaneously act as the carrier and the antigen. For instance, Dong et al had built self-crosslinked positively charged OVA NPs (ONPs) which used OVA and PEI through the crosslinking of genipin. Subsequently, directly adsorbed the negatively charged CpG on the surface of ONPs through electrostatic interaction, thus successfully constructing multifunctional nanovaccines. The ONPs-CpG nanocavines could provide sufficient antigens, promote the maturation and activation of immune cells, and strongly induce specific anti-tumor immune responses.84
CpG Loaded in Nanovaccines by Covalent Bonding
Studies have shown that the covalent bonding was also a good way for loading CpG. The modified CpG can form covalent bonds with the nanocarrier to introduce them into the tumor nanovaccines. Wang et al designed a simple nanovaccine which was entirely composed of antigen OVA and a small amount of CpG adjuvant without the help of other external nanocarriers. Through the action of hydrophobic regulator sodium dodecyl sulfonate (SDS), the interaction between the sulfhydryl groups formed a self-assembled OVA NPs with disulfide bonds (-S-S-) network, and in the meantime, it could form disulfide bond with sulfhydryl-modified CpG to construct the nanovaccines with simple ingredients, and experiments had confirmed that the nanovaccines strongly induced CTLs to achieve a significant anti-tumor effect (Figure 3).85
 |
Figure 3 Design of “minimalist” OVA@CpG nanovaccine. (A) Traditional nanovaccine and “minimalist” nanovaccine. (B) The general process of constructing the “minimalist” nanovaccine. Adapted with permission from Wang K, Wen S, He L, et al. “Minimalist” nanovaccine constituted from near whole antigen for cancer immunotherapy. ACS Nano. 2018;12(7):6398–6409. Copyright 2018 American Chemical Society.85 |
Similarly, Liu et al used a hydrophobic biomimetic phospholipid material DOPE (1,2-dioleoyl-sn-glycero-3-phosphoethanolamine) as the carrier, First, under the action of 3-(2-pyridinyl dithiopropionic acid N-hydroxysuccinimide ester (SPDP), DOPE was pyridinedithiolated, and then pyridyldithiol-activated DOPE could be combined with the thiol-terminated hydrophilic adjuvant CpG ODN through forming the disulfide bonds, and finally loaded with OVA to construct the bionic nanovaccines with good prospects. The obtained nanovaccines could provide sustained and effective antigen stimulation, promote the release of some immune factors, further boost significantly anti-tumor immunity to inhibit tumor growth.86 Moreover, there is another way to introduce CpG into tumor nanovaccines through the formation of covalent bonds between S and other atoms. For example, Zhu et al developed albumin/albumin-binding vaccines (AlbiVax) nanocomplexes that achieved high efficiency co-delivery of antigen and adjuvant CpG. The mercaptated CpG could be covalently cross-linked with the maleimide functionalized Evans blue (EB) derivative to form a thioether covalent bond, thus realizing the stable loading of CpG in the nanovaccines. The albumin/AlbiVax nanocomplexes elicited a strong CTLs response to effectively inhibit or eliminate tumors, and the combination of nanovaccine with immune checkpoint inhibitors (anti-PD-1) could further enhance the efficiency of cancer immunotherapy.87 The use of S atoms to form strong polar covalent bonds with metal atoms has also been reported in previous studies, such as the covalent bonds with gold NPs.88
In addition, the way to introduce CpG into nanovaccines through covalent bond is not limited to the disulfide bonds or coordination bonds formed by the interaction with S, and other methods can also be used. For instance, Wang et al used aldehyde-terminated CpG instead of sulfhydryl-terminated CpG to covalently react with the amino group of the membrane protein on tumor-derived nanovesicles to form amide bonds, in order that CpG could successfully bind to the surface of the nanovesicles. The obtained tumor-derived vesicle-based nanovaccine was implemented in three different vaccine delivery modes, resulting in different degrees of anti-tumor effects.89 The introduction of CpG into tumor nanovaccines through this method has also been reported in the past.90 Compared with the electrostatic interaction, covalent bonding is a more stable way to introduce CpG into nanovaccines. However, CpG loading via covalent bond usually needs complicated chemical synthesis and modification under the participation of organic solvents, which may bring adverse effects on the immunological-activities of the antigens and adjuvants.
CpG Loaded in Nanovaccines by Hydrophilic and Hydrophobic Interaction
Despite CpG-loaded nanovaccine formed by covalent bonding is relatively stable, some chemical conjugating process is difficult to implement between CpG and nanocarriers.91 Hydrophilic and hydrophobic interaction is another method of introducing CpG into tumor nanovaccines. Owing to CpG is hydrophilic, it may be not easy to be delivered to the LNs. Using some suitable nanomaterials to encapsulate CpG through hydrophilic and hydrophobic interaction can achieve some advantages, such as prevented degradation, prolonged half-life, controlled release.92,93 Among the nanomaterials, liposomes are commonly used carriers, because of their earlier clinical application and more mature development. Zhao et al used the reversed-phase evaporation method to prepare nanovaccines encapsulating CpG by hydrophilic and hydrophobic interaction. Firstly, the compounds used to form the lipid film were dissolved. After the lipid film was formed, E7 peptide (Human papillomavirus type 16) and CpG ODN were added into the lipid film solution to form a water-in-oil emulsion, and finally got the liposome vaccine encapsulating CpG and E7 peptide through evaporation and other steps. The liposome nanovaccine was also modified with mannose for accurately targeting DCs, enhanced cellular immunity was realized to induce strong anti-tumor effects.94 Additionally, poly(lactic-co-glycolic acid) (PLGA) is also a commonly used degradable polymer with good biocompatibility. Nanovaccines made by PLGA have been widely used in cancer treatment. In the process of preparing PLGA NPs, most of them are prepared by the double emulsion solvent evaporation method. Sainz et al had successfully prepared PLGA nanovaccines co-encapsulated with CpG and other substances. The study had shown that the constructed nanovaccines worked synergistically with α-Galactosylceramide (GalCer), exhibited a high degree of anti-tumor effect in the established mouse tumor model of melanoma, and significantly inhibited the tumor growth.95 Kokate et al used solvent evaporation technology to prepare biodegradable PLGA NPs to co-encapsulate tumor antigen and CpG. Experimental results showed that the nanovaccine effectively promoted the maturation and activation of DCs derived from bone marrow, induced a strong CTLs immune response to effectively inhibited tumor growth.96 There are also studies using spray drying method to obtain PLGA microspheres.97 In addition to PLGA, there is also the recently discovered new polymer material poly(4,4ʹ-trimethylenedipiperdyl sulfide) (PNSN) that can be used to wrap CpG. Geary et al prepared PNSN microparticles by the aforementioned double emulsion solvent evaporation method, dissolving OVA and CpG as the internal water phase, and PNSN in dichloromethane as the oil phase. A secondary emulsion was obtained through a series of operations, and finally evaporated dichloromethane, and freeze-dried the PNSN particles co-encapsulating CpG and OVA.93
CpG Loaded in Nanovaccines Through DNA Self-Assembling
With the development of nucleic acid nanotechnology, it has been a potential strategy to accurately design nanostructures by using synthetic nucleic acids. On the basis of this technical theory, studies have proposed an innovative research idea, using CpG DNA not only as the immune adjuvant, but also as the carrier material of nanovaccines. Zhang et al had constructed the self-assembled DNA nanoflowers (NFs) with immunostimulatory capacity. Firstly, a linear template containing three CpG complementary sequences was connected to form a circular structure under the action of T4 deoxyribonucleic acid ligase. Subsequently, Φ29 DNA polymerase was used for rolling circle replication (RCR) through the action of a FITC-labeled primer, and finally the CpG NFs was obtained through liquid crystal self-assembly.98 Importantly, the self-assembly process of this NFs did not depend on the base pairing between the DNA chains.99 This is also the first time that the NFs assembly process has been proposed, and this method has been developed for the construction of CpG-based nanovaccines.98 CpG can be introduced into nanovaccines through DNA self-assembly. Briefly, DNA template containing CpG complementary sequence and primer chain was used to realize self-assembly of CpG NFs through RCR, and finally antigen was loaded to obtain the nanovaccines.67,100,101 For instance, Zhu et al described the DNA-inorganic hybrid nanovaccines containing not only the CpG sequence but also the inorganic magnesium pyrophosphate (Mg2PPi). The addition of Mg2PPi made the assembled nanovaccines more stable and resistant to nuclease degradation. Likewise, linearized DNA templates containing CpG sequences were used. After circularization under the action of T4 ligase, a large number of tandem CpG analogues were generated by RCR, and pyrophosphate ions were added into the buffer containing magnesium ions to form Mg2PPi. CpG analogues could be precipitated in Mg2PPi, and finally self-assembled to form nanovaccines. The experiments had shown that the nanovaccines were able to be effectively absorbed into APCs to produce effective immune stimulation, thereby achieving the effect of cancer immunotherapy.67 Subsequently, Zhu et al used this method to construct another self-assembled DNA-RNA nanovaccine. The tandem DNA and tandem small hairpin RNA (shRNA) nanostructures were constructed by co-crystallization of nucleic acids and inorganic supplements during RCR and rolling circle transcription (RCT) processes. The tandem DNA and shRNA were self-assembled into intertwining DNA-RNA microflowers (iDR-MFs). After constructing and forming iDR-MFs, they were shrunk to form intertwining DNA-RNA nanocapsules (iDR-NCs) under the action of PEG-grafted polypeptide (PPT-g-PEG), and finally the antigen was further loaded by hydrophobic interaction to obtain the self-assembled DNA-RNA nanovaccines (Figure 4).100 It was worth noting that the nanovaccines could induce immensely strong immune stimulus response, significantly inhibited the growth of colorectal tumors. In addition, similar studies have also appeared in the past year. Ni et al have designed and developed bi-adjuvant neoantigen nanovaccine (banNV) to achieve the co-delivery of the neoantigen and the two adjuvants (TLR7/8 agonist R848 and TLR9 agonist CpG). The main synthetic method of the nanometer vaccine was described below. In the first place, the amphiphilic poly(ethylene oxide)-block-poly(D,L-lactic acid) (PEG-b-PLA) micelles coated with DNA primers were used for RCR with the DNA encoding CpG template to obtain self-assembled microparticles composed of PEG-PLA micelle cores and polymerized CpG shells, and then the R848 adjuvant was physically loaded. Finally, with the help of PPT-g-PEG, the neoantigen was effectively introduced and the size of the self-assembled microparticles was also reduced to obtain NPs. The nanovaccines effectively realized the joint delivery of antigen and two adjuvants, which caused a strong immunostimulation, effectively promoted the maturation of DCs, and boosted the CTLs response. In addition, studies have shown that the nanovaccines combined with immune checkpoint inhibitors (especially PD-1 antibody) had a significant anti-tumor effect.101
 |
Figure 4 The iDR-NC/neoantigen nanovaccines constructed by DNA self-assembly loaded with CpG. (A) Tandem CpG and shRNAs were produced through RCR and RCT, and self-assembled into intertwining DNA-RNA MFs. (B) The above-mentioned MFs were contracted by PPT-g-PEG into iDR-NCs, which were further loaded with tumor neoantigens. (C) Specific anti-tumor response induced by iDR-NCs/neoantigen nanovaccines in mice. Adapted from Zhu G, Mei L, Vishwasrao HD, et al. Intertwining DNA-RNA nanocapsules loaded with tumor neoantigens as synergistic nanovaccines for cancer immunotherapy. Nat Commun. 2017;8(1):1482.100 |
Collectively, this CpG-loaded method has good flexibility. By designing multifarious DNA templates, different CpG ODNs could be loaded in the nanovaccines through DNA self-assembling, which could flexibly endow and regulate the function of the nanovaccines, and effectively improve the stability of CpG.102
CpG Loaded in Nanovaccines Through Other Methods
In addition to the common methods described above, there were also other strategies to package CpG into the nanovaccines. Based on the comprehensive consideration of the physicochemical properties of CpG and nanocarriers, suitable methods or innovative strategies were explored to introduce CpG into the nanovaccines. For instance, the method of inserting cholesterol-modified CpG into phospholipids is a commonly used method. For instance, Qian et al constructed α-AP-FNPs (ultra-small biocompatible nanoparticles) as carriers, loaded with three different antigenic peptide (AP) sequences, and inserted cholesterol-modified CpG into phospholipids on the surface of the NPs to successfully construct CpG-loaded nanovaccines. In this study, they observed that three different AP sequence did not affect the ability of targeting scavenger receptor class B1 (SR-B1), and choosing different AP sequences might have a targeted therapeutic effect on different types of cancers. It was worth noting that the nanovaccines without CpG also induced specific anti-tumor immune responses, while the nanovaccines loaded with CpG showed stronger anti-tumor efficacy.103 Kuai et al used a high-density lipoprotein (sHDL) nanodisk as a carrier, which was composed of phospholipids and mimetic peptides. The neoantigen-lipid conjugate and cholesterol-modified CpG could be inserted into the nanodisk through hydrophobic interaction, which improved the effective co-delivery of antigen and adjuvant. Comprehensive experimental data showed that the nanodisk vaccines induced an immensely strong immune response to motivate specific CTLs, and the anti-tumor effect was more prominent when combined with immune checkpoint inhibitors (Figure 5).104
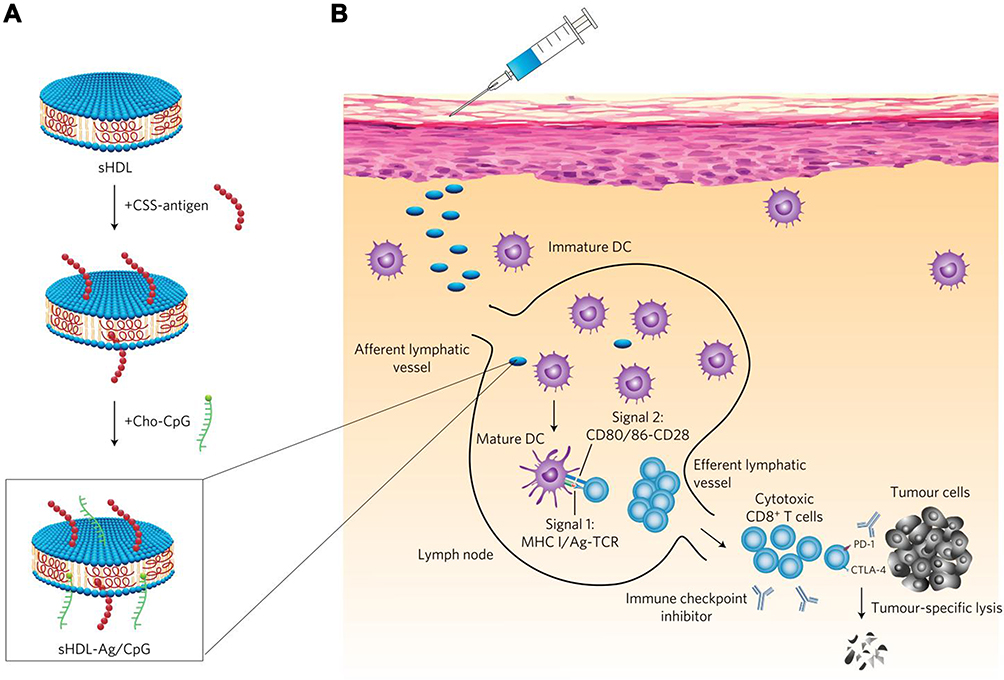 |
Figure 5 Construction of sHDL nanodisk platform for personalized vaccines for cancer immunotherapy. (A) The binding process of antigen and adjuvant CpG into sHDL nanodisk platform. (B) The designed personalized nanovaccines could induce a strong CTL response in vivo. Reprinted by permission from Springer Nature Customer Service Centre GmbH: Springer Nature, Nature Materials. Kuai R, Ochyl LJ, Bahjat KS, Schwendeman A, Moon JJ. Designer vaccine nanodiscs for personalized cancer immunotherapy. Nat Mater. 2017;16(4):489–496. doi:10.1038/nmat4822. Copyright 2017.104 |
Cai et al used tetrakis (4-carboxyphenyl) porphyrin (H2TCPP) and zirconium ions to construct a metal-organic framework (MOF), loaded with acriflavine (ACF) through the porous internal structure of the MOF, and then CpG was evenly distributed on the surface of the MOF under ultrasound treatment, finally directly coated with hyaluronic acid (HA), which also increased the loading rate of CpG. The results indicated that the system of nanovaccines could be used in combination with photodynamic therapy for cancer treatment.105
Liu et al used tannic acid (TA) as a nanocarrier material, and successfully constructed the tumor nanovaccines with a potent subunit antigen and the adjuvant CpG for Epstein-Barr virus (EBVs)-associated tumors. As well known, TA is a typical glucosyl compound, and its special structure also allows the particular activities. It can be combined with alkaloids, proteins, polysaccharides and other molecules through hydrogen bonding. As a carrier, TA can load CpG and tumor-associated protein antigens through hydrogen bonding. What’s more, the obtained nanovaccines could accurately target the LNs, induced strong immune stimulation, and reshaped the TME to break the tumor immune tolerance, which was promising to be used in the future for the effective treatment of clinical patients with EBVs positive tumors.106
The above-enumerated CpG-based nanovaccines for cancer immunotherapy were summarized in Table 2. These CpG-based nanovaccines were described according to the following information: loading technique, selected CpG type, used antigen, tumor type, therapeutic effect, and studied in vitro or in vivo.
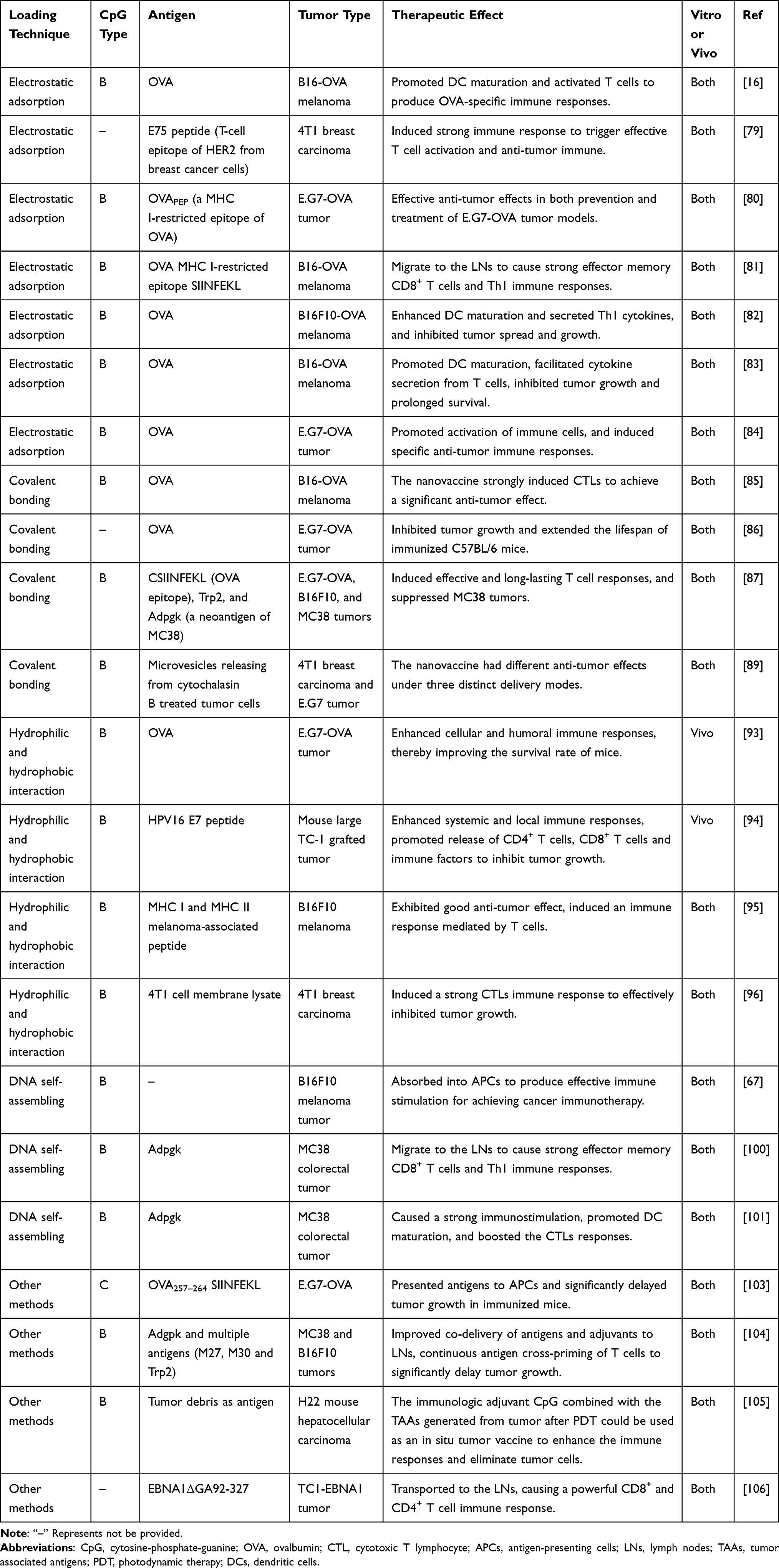 |
Table 2 Summary of the CpG-Based Nanovaccines for Cancer Immunotherapy |
Challenge and Future Perspectives
Although the studies to date have shown exciting and encouraging progress in CpG-loaded nanovaccines, there are still many intractable challenges which need to be addressed. First of all, it is crucial to comprehensively elucidate the action mechanisms of the CpG-based nanovaccines. The behaviors of nanovaccines in vivo are closely related to their physicochemical property parameters and the disease state of the patients, the complicated correlations between these issues must be fully understood. Most of these nanovaccines have not been evaluated in clinic, and some intractable clinical problems about using CpG-based nanovaccines have to be settled urgently, involving safety, practicability, and effectiveness. The materials used to construct these NPs may be synthetic or extracted from nature, which may lack effective evaluation in clinical practice and may produce a series of adverse reactions in vivo. In addition, most of the materials need to be obtained through complex and time-consuming preparation process, these high costs will hamper the development and application of these nanovaccines. There are also important issues about the effectiveness of nanovaccines. A large amount of basic research with CpG-based tumor nanovaccines had been conducted on the basis of animal experiments. Although animal models are able to provide some informative experimental data, they are incapable to fully simulate the complicated physiological pathological environments and the actual development of tumors in human body.107 Some nanovaccines showed effectual in research but lackluster in clinical translation. Hence, there are numerous issues for the nanovaccines that need to be considered from basic research to clinical translation, and Quality by Design (QbD) should be taken into consideration throughout the research and development of the nanovaccines for ensuring their quality. Extensive efforts in the future will be needed to further improve the safety and practicability of the CpG-based tumor nanovaccines, and the effectiveness should be optimized for insuring the cancer immunotherapy available and efficacious to the vast patients.
Conclusion
In this review, we have summarized recent progress on CpG-based nanovaccines for cancer immunotherapy. Three types of CpG ODNs (Type A, B and C) were introduced in detail along with their different properties. Some recently representative CpG-based nanovaccines for cancer immunotherapy were illustrated according to the diverse CpG loading patterns, including electrostatic adsorption, covalent bonding, hydrophilic and hydrophobic interaction, DNA self-assembly, and also other loading methods. In addition, the challenges including safety, practicability and effectiveness of CpG-based nanovaccines from basic research to clinical translation were also proposed. It is hoped that this review will provide valuable references for the development of nanovaccines in cancer immunotherapy.
Acknowledgments
The authors are thankful to the National Natural Science Foundation of China (81973671) and the Natural Science Foundation of Shandong Province (ZR2019BEM003) for financial support to this work.
Author Contributions
All authors contributed to data analysis, drafting or revising the article, have agreed on the journal to which the article will be submitted, gave final approval of the version to be published, and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Ma X, Yu H. Global burden of cancer. Yale J Biol Med. 2006;79(3–4):85–94.
2. Kim R, Emi M, Tanabe K. Cancer immunoediting from immune surveillance to immune escape. Immunology. 2007;121(1):1–14. doi:10.1111/j.1365-2567.2007.02587.x
3. Gonzalez H, Hagerling C, Werb Z. Roles of the immune system in cancer: from tumor initiation to metastatic progression. Genes Dev. 2018;32(19–20):1267–1284. doi:10.1101/gad.314617.118
4. Krishnamachari Y, Geary SM, Lemke CD, Salem AK. Nanoparticle delivery systems in cancer vaccines. Pharm Res. 2011;28(2):215–236. doi:10.1007/s11095-010-0241-4
5. Arruebo M, Vilaboa N, Sáez-Gutierrez B, et al. Assessment of the evolution of cancer treatment therapies. Cancers (Basel). 2011;3(3):3279–3330. doi:10.3390/cancers3033279
6. Mellman I, Coukos G, Dranoff G. Cancer immunotherapy comes of age. Nature. 2011;480(7378):480–489. doi:10.1038/nature10673
7. Rosenberg SA, Yang JC, Restifo NP. Cancer immunotherapy: moving beyond current vaccines. Nat Med. 2004;10(9):909–915. doi:10.1038/nm1100
8. Sharma P, Wagner K, Wolchok JD, Allison JP. Novel cancer immunotherapy agents with survival benefit: recent successes and next steps. Nat Rev Cancer. 2011;11(11):805–812. doi:10.1038/nrc3153
9. Zhang Y, Cui Z, Kong H, et al. One-shot immunomodulatory nanodiamond agents for cancer immunotherapy. Adv Mater. 2016;28(14):2699–2708. doi:10.1002/adma.201506232
10. Zhu G, Zhang F, Ni Q, Niu G, Chen X. Efficient nanovaccine delivery in cancer immunotherapy. ACS Nano. 2017;11(3):2387–2392. doi:10.1021/acsnano.7b00978
11. Banchereau J, Steinman RM. Dendritic cells and the control of immunity. Nature. 1998;392(6673):245–252. doi:10.1038/32588
12. Dubois B, Lamy PJ, Chemin K, Lachaux A, Kaiserlian D. Measles virus exploits dendritic cells to suppress CD4+ T-cell proliferation via expression of surface viral glycoproteins independently of T-cell trans-infection. Cell Immunol. 2001;214(2):173–183. doi:10.1006/cimm.2001.1898
13. Mailliard RB, Dallal RM, Son YI, Lotze MT. Dendritic cells promote T-cell survival or death depending upon their maturation state and presentation of antigen. Immunol Invest. 2000;29(2):177–185. doi:10.3109/08820130009062302
14. Farkona S, Diamandis EP, Blasutig IM. Cancer immunotherapy: the beginning of the end of cancer? BMC Med. 2016;14(1):73. doi:10.1186/s12916-016-0623-5
15. Foged C. Subunit vaccines of the future: the need for safe, customized and optimized particulate delivery systems. Ther Deliv. 2011;2(8):1057–1077. doi:10.4155/tde.11.68
16. Guan X, Chen J, Hu Y, et al. Highly enhanced cancer immunotherapy by combining nanovaccine with hyaluronidase. Biomaterials. 2018;171:198–206. doi:10.1016/j.biomaterials.2018.04.039
17. Moyle PM, Toth I. Modern subunit vaccines: development, components, and research opportunities. ChemMedChem. 2013;8(3):360–376. doi:10.1002/cmdc.201200487
18. Purcell AW, McCluskey J, Rossjohn J. More than one reason to rethink the use of peptides in vaccine design. Nat Rev Drug Discov. 2007;6(5):404–414. doi:10.1038/nrd2224
19. Gause KT, Wheatley AK, Cui J, Yan Y, Kent SJ, Caruso F. Immunological principles guiding the rational design of particles for vaccine delivery. ACS Nano. 2017;11(1):54–68. doi:10.1021/acsnano.6b07343
20. Maina TW, Grego EA, Boggiatto PM, Sacco RE, Narasimhan B, McGill JL. Applications of nanovaccines for disease prevention in cattle. Front Bioeng Biotechnol. 2020;8:608050. doi:10.3389/fbioe.2020.608050
21. Gheibi Hayat SM, Darroudi M. Nanovaccine: a novel approach in immunization. J Cell Physiol. 2019;234(8):12530–12536. doi:10.1002/jcp.28120
22. Siddique S, Chow JCL. Application of nanomaterials in biomedical imaging and cancer therapy. Nanomaterials (Basel). 2020;10(9):1700. doi:10.3390/nano10091700
23. Aghebati-Maleki A, Dolati S, Ahmadi M, et al. Nanoparticles and cancer therapy: perspectives for application of nanoparticles in the treatment of cancers. J Cell Physiol. 2020;235(3):1962–1972. doi:10.1002/jcp.29126
24. Siddique S, Chow JCL. Gold nanoparticles for drug delivery and cancer therapy. Appl Sci. 2020;10(11):3824. doi:10.3390/app10113824
25. Ansari MT, Ramlan TA, Jamaluddin NN, et al. Lipid-based nanocarriers for cancer and tumor treatment. Curr Pharm Des. 2020;26(34):4272–4276. doi:10.2174/1381612826666200720235752
26. Moore JA, Chow JCL. Recent progress and applications of gold nanotechnology in medical biophysics using artificial intelligence and mathematical modeling. Nano Express. 2021;2(2):022001. doi:10.1088/2632-959X/abddd3
27. Riley RS, June CH, Langer R, Mitchell MJ. Delivery technologies for cancer immunotherapy. Nat Rev Drug Discov. 2019;18(3):175–196. doi:10.1038/s41573-018-0006-z
28. Zhang Y, Lin S, Wang XY, Zhu G. Nanovaccines for cancer immunotherapy. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2019;11(5):e1559. doi:10.1002/wnan.1559
29. Vijayan V, Mohapatra A, Uthaman S, Park IK. Recent advances in nanovaccines using biomimetic immunomodulatory materials. Pharmaceutics. 2019;11(10):534. doi:10.3390/pharmaceutics11100534
30. Heeg K, Dalpke A, Peter M, Zimmermann S. Structural requirements for uptake and recognition of CpG oligonucleotides. Int J Med Microbiol. 2008;298(1–2):33–38. doi:10.1016/j.ijmm.2007.07.007
31. Krieg AM, Wu T, Weeratna R, et al. Sequence motifs in adenoviral DNA block immune activation by stimulatory CpG motifs. Proc Natl Acad Sci USA. 1998;95(21):12631–12636. doi:10.1073/pnas.95.21.12631
32. Krieg AM. CpG motifs in bacterial DNA and their immune effects. Annu Rev Immunol. 2002;20(1):709–760. doi:10.1146/annurev.immunol.20.100301.064842
33. Dalpke AH, Zimmermann S, Albrecht I, Heeg K. Phosphodiester CpG oligonucleotides as adjuvants: polyguanosine runs enhance cellular uptake and improve immunostimulative activity of phosphodiester CpG oligonucleotides in vitro and in vivo. Immunology. 2002;106(1):102–112. doi:10.1046/j.1365-2567.2002.01410.x
34. Krieg AM. Therapeutic potential of Toll-like receptor 9 activation. Nat Rev Drug Discov. 2006;5(6):471–484. doi:10.1038/nrd2059
35. Crooks J, Gargaro M, Vacca C, et al. CpG Type A induction of an early protective environment in experimental multiple sclerosis. Mediators Inflamm. 2017;2017:1380615. doi:10.1155/2017/1380615
36. Vollmer J, Krieg AM. Immunotherapeutic applications of CpG oligodeoxynucleotide TLR9 agonists. Adv Drug Deliv Rev. 2009;61(3):195–204. doi:10.1016/j.addr.2008.12.008
37. Klinman DM. Immunotherapeutic uses of CpG oligodeoxynucleotides. Nat Rev Immunol. 2004;4(4):249–258. doi:10.1038/nri1329
38. Ballas ZK. Modulation of NK cell activity by CpG oligodeoxynucleotides. Immunol Res. 2007;39(1–3):15–21. doi:10.1007/s12026-007-0066-3
39. Verthelyi D, Ishii KJ, Gursel M, Takeshita F, Klinman DM. Human peripheral blood cells differentially recognize and respond to two distinct CPG motifs. J Immunol. 2001;166(4):2372–2377. doi:10.4049/jimmunol.166.4.2372
40. Wooldridge JE, Weiner GJ. CpG DNA and cancer immunotherapy: orchestrating the antitumor immune response. Curr Opin Oncol. 2003;15(6):440–445. doi:10.1097/00001622-200311000-00006
41. Marshall JD, Fearon K, Abbate C, et al. Identification of a novel CpG DNA class and motif that optimally stimulate B cell and plasmacytoid dendritic cell functions. J Leukoc Biol. 2003;73(6):781–792. doi:10.1189/jlb.1202630
42. Krug A, Rothenfusser S, Hornung V, et al. Identification of CpG oligonucleotide sequences with high induction of IFN-alpha/beta in plasmacytoid dendritic cells. Eur J Immunol. 2001;31(7):2154–2163. doi:10.1002/1521-4141(200107)31:7<2154::AID-IMMU2154>3.0.CO;2-U
43. Li T, Wu J, Zhu S, et al. A novel C type CpG oligodeoxynucleotide exhibits immunostimulatory activity in vitro and enhances antitumor effect in vivo. Front Pharmacol. 2020;11:8. doi:10.3389/fphar.2020.00008
44. Wilson HL, Dar A, Napper SK, Marianela Lopez A, Babiuk LA, Mutwiri GK. Immune mechanisms and therapeutic potential of CpG oligodeoxynucleotides. Int Rev Immunol. 2006;25(3–4):183–213. doi:10.1080/08830180600785868
45. van Ojik HH, Bevaart L, Dahle CE, et al. CpG-A and B oligodeoxynucleotides enhance the efficacy of antibody therapy by activating different effector cell populations. Cancer Res. 2003;63(17):5595–5600.
46. Hartmann G, Battiany J, Poeck H, et al. Rational design of new CpG oligonucleotides that combine B cell activation with high IFN-alpha induction in plasmacytoid dendritic cells. Eur J Immunol. 2003;33(6):1633–1641. doi:10.1002/eji.200323813
47. Krug A, Towarowski A, Britsch S, et al. Toll-like receptor expression reveals CpG DNA as a unique microbial stimulus for plasmacytoid dendritic cells which synergizes with CD40 ligand to induce high amounts of IL-12. Eur J Immunol. 2001;31(10):3026–3037. doi:10.1002/1521-4141(2001010)31:10<3026::AID-IMMU3026>3.0.CO;2-H
48. Scheiermann J, Klinman DM. Clinical evaluation of CpG oligonucleotides as adjuvants for vaccines targeting infectious diseases and cancer. Vaccine. 2014;32(48):6377–6389. doi:10.1016/j.vaccine.2014.06.065
49. Yamamoto Y, Sugimura R, Watanabe T, et al. Class A CpG oligonucleotide priming rescues mice from septic shock via activation of platelet-activating factor acetylhydrolase. Front Immunol. 2017;8:1049. doi:10.3389/fimmu.2017.01049
50. Shirota H, Gursel I, Gursel M, Klinman DM. Suppressive oligodeoxynucleotides protect mice from lethal endotoxic shock. J Immunol. 2005;174(8):4579–4583. doi:10.4049/jimmunol.174.8.4579
51. Kobayashi N, Hong C, Klinman DM, Shirota H. Oligodeoxynucleotides expressing polyguanosine motifs promote antitumor activity through the upregulation of IL-2. J Immunol. 2013;190(4):1882–1889. doi:10.4049/jimmunol.1201063
52. Lemke-Miltner CD, Blackwell SE, Yin C, et al. Antibody opsonization of a TLR9 agonist-containing virus-like particle enhances in situ immunization. J Immunol. 2020;204(5):1386–1394. doi:10.4049/jimmunol.1900742
53. Mutwiri GK, Nichani AK, Babiuk S, Babiuk LA. Strategies for enhancing the immunostimulatory effects of CpG oligodeoxynucleotides. J Control Release. 2004;97(1):1–17. doi:10.1016/j.jconrel.2004.02.022
54. Gürsel M, Verthelyi D, Gürsel I, Ishii KJ, Klinman DM. Differential and competitive activation of human immune cells by distinct classes of CpG oligodeoxynucleotide. J Leukoc Biol. 2002;71(5):813–820.
55. Vollmer J, Weeratna R, Payette P, et al. Characterization of three CpG oligodeoxynucleotide classes with distinct immunostimulatory activities. Eur J Immunol. 2004;34(1):251–262. doi:10.1002/eji.200324032
56. Li T, Hua C, Yue W, et al. Discrepant antitumor efficacies of three CpG oligodeoxynucleotide classes in monotherapy and co-therapy with PD-1 blockade. Pharmacol Res. 2020;161:105293. doi:10.1016/j.phrs.2020.105293
57. Campbell JD. Development of the CpG adjuvant 1018: a case study. Methods Mol Biol. 2017;1494:15–27.
58. Liang X, Moseman EA, Farrar MA, et al. Toll-like receptor 9 signaling by CpG-B oligodeoxynucleotides induces an apoptotic pathway in human chronic lymphocytic leukemia B cells. Blood. 2010;115(24):5041–5052. doi:10.1182/blood-2009-03-213363
59. van Pul KM, Vuylsteke R, de Beijer MTA, et al. Breast cancer-induced immune suppression in the sentinel lymph node is effectively countered by CpG-B in conjunction with inhibition of the JAK2/STAT3 pathway. J Immunother Cancer. 2020;8(2):e000761. doi:10.1136/jitc-2020-000761
60. Jurk M, Schulte B, Kritzler A, et al. C-Class CpG ODN: sequence requirements and characterization of immunostimulatory activities on mRNA level. Immunobiology. 2004;209(1–2):141–154. doi:10.1016/j.imbio.2004.02.006
61. Sorski L, Melamed R, Levi B, et al. Prevention of liver metastases through perioperative acute CpG-C immune stimulation. Cancer Immunol Immunother. 2020;69(10):2021–2031. doi:10.1007/s00262-020-02596-7
62. Gray RC, Kuchtey J, Harding CV. CpG-B ODNs potently induce low levels of IFN-alphabeta and induce IFN-alphabeta-dependent MHC-I cross-presentation in DCs as effectively as CpG-A and CpG-C ODNs. J Leukoc Biol. 2007;81(4):1075–1085. doi:10.1189/jlb.1006606
63. Ingelsson B, Söderberg D, Strid T, et al. Lymphocytes eject interferogenic mitochondrial DNA webs in response to CpG and non-CpG oligodeoxynucleotides of class C. Proc Natl Acad Sci USA. 2018;115(3):E478–e487. doi:10.1073/pnas.1711950115
64. Mathur S, Walley KR, Boyd JH. The Toll-like receptor 9 ligand CPG-C attenuates acute inflammatory cardiac dysfunction. Shock. 2011;36(5):478–483. doi:10.1097/SHK.0b013e31822d6442
65. Li L, Xu Z, Zuo J, Ding J. A C-type CpG ODN accelerates wound healing via regulating fibroblasts and immune response. J Cell Biochem. 2019;120(5):7868–7875. doi:10.1002/jcb.28061
66. Hartmann G, Weiner GJ, Krieg AM. CpG DNA: a potent signal for growth, activation, and maturation of human dendritic cells. Proc Natl Acad Sci U S A. 1999;96(16):9305–9310. doi:10.1073/pnas.96.16.9305
67. Zhu G, Liu Y, Yang X, et al. DNA-inorganic hybrid nanovaccine for cancer immunotherapy. Nanoscale. 2016;8(12):6684–6692. doi:10.1039/C5NR08821F
68. Fourcade J, Kudela P, Andrade Filho PA, et al. Immunization with analog peptide in combination with CpG and montanide expands tumor antigen-specific CD8+ T cells in melanoma patients. J Immunother. 2008;31(8):781–791. doi:10.1097/CJI.0b013e318183af0b
69. Karbach J, Gnjatic S, Bender A, et al. Tumor-reactive CD8+ T-cell responses after vaccination with NY-ESO-1 peptide, CpG 7909 and Montanide ISA-51: association with survival. Int J Cancer. 2010;126(4):909–918.
70. Valmori D, Souleimanian NE, Tosello V, et al. Vaccination with NY-ESO-1 protein and CpG in Montanide induces integrated antibody/Th1 responses and CD8 T cells through cross-priming. Proc Natl Acad Sci USA. 2007;104(21):8947–8952. doi:10.1073/pnas.0703395104
71. Speiser DE, Liénard D, Rufer N, et al. Rapid and strong human CD8+ T cell responses to vaccination with peptide, IFA, and CpG oligodeoxynucleotide 7909. J Clin Invest. 2005;115(3):739–746. doi:10.1172/JCI23373
72. Katsuda M, Iwahashi M, Matsuda K, et al. Comparison of different classes of CpG-ODN in augmenting the generation of human epitope peptide-specific CTLs. Int J Oncol. 2011;39(5):1295–1302.
73. Ming J, Zhang J, Shi Y, et al. A trustworthy CpG nanoplatform for highly safe and efficient cancer photothermal combined immunotherapy. Nanoscale. 2020;12(6):3916–3930. doi:10.1039/C9NR09402D
74. Hanagata N. CpG oligodeoxynucleotide nanomedicines for the prophylaxis or treatment of cancers, infectious diseases, and allergies. Int J Nanomedicine. 2017;12:515–531. doi:10.2147/IJN.S114477
75. Zupančič E, Curato C, Kim JS, et al. Nanoparticulate vaccine inhibits tumor growth via improved T cell recruitment into melanoma and huHER2 breast cancer. Nanomedicine. 2018;14(3):835–847. doi:10.1016/j.nano.2017.12.011
76. Han S, Wang W, Wang S, et al. Multifunctional biomimetic nanoparticles loading baicalin for polarizing tumor-associated macrophages. Nanoscale. 2019;11(42):20206–20220. doi:10.1039/C9NR03353J
77. Adamus T, Kortylewski M. The revival of CpG oligonucleotide-based cancer immunotherapies. Contemp Oncol (Pozn). 2018;22(1a):56–60.
78. Shirota H, Klinman DM. Recent progress concerning CpG DNA and its use as a vaccine adjuvant. Expert Rev Vaccines. 2014;13(2):299–312. doi:10.1586/14760584.2014.863715
79. Liu X, Feng Z, Wang C, et al. Co-localized delivery of nanomedicine and nanovaccine augments the postoperative cancer immunotherapy by amplifying T-cell responses. Biomaterials. 2020;230:119649. doi:10.1016/j.biomaterials.2019.119649
80. Kim Y, Kang S, Shin H, et al. Sequential and timely combination of a cancer nanovaccine with immune checkpoint blockade effectively inhibits tumor growth and relapse. Angew Chem Int Ed Engl. 2020;59(34):14628–14638. doi:10.1002/anie.202006117
81. Shi X, Song H, Wang C, et al. Co-assembled and self-delivered epitope/CpG nanocomplex vaccine augments peptide immunogenicity for cancer immunotherapy. Chem Eng J. 2020;399:125854. doi:10.1016/j.cej.2020.125854
82. Ruiz-de-angulo A, Zabaleta A, Gómez-Vallejo V, Llop J, Mareque-Rivas JC. Microdosed lipid-coated (67) Ga-magnetiteenhances antigen-specific immunity by Image tracked delivery of antigen and CpG to lymph nodes. ACS Nano. 2016;10(1):1602–1618. doi:10.1021/acsnano.5b07253
83. Liu Y, Yao L, Cao W, et al. Dendritic cell targeting peptide-based nanovaccines for enhanced cancer immunotherapy. ACS Appl Bio Mater. 2019;2(3):1241–1254. doi:10.1021/acsabm.8b00811
84. Dong X, Liang J, Yang A, Qian Z, Kong D, Lv F. A visible codelivery nanovaccine of antigen and adjuvant with self-carrier for cancer immunotherapy. ACS Appl Mater Interfaces. 2019;11(5):4876–4888. doi:10.1021/acsami.8b20364
85. Wang K, Wen S, He L, et al. “Minimalist” nanovaccine constituted from near whole antigen for cancer immunotherapy. ACS Nano. 2018;12(7):6398–6409. doi:10.1021/acsnano.8b00558
86. Liu D, Liu J, Ma B, et al. A simple self-adjuvanting biomimetic nanovaccine self-assembled with the conjugate of phospholipids and nucleotides can induce a strong cancer immunotherapeutic effect. Biomater Sci. 2021;9(1):84–92. doi:10.1039/D0BM01333A
87. Zhu G, Lynn GM, Jacobson O, et al. Albumin/vaccine nanocomplexes that assemble in vivo for combination cancer immunotherapy. Nat Commun. 2017;8(1):1954. doi:10.1038/s41467-017-02191-y
88. Wang Y, Wang Y, Kang N, et al. Construction and immunological evaluation of CpG-Au@HBc virus-like nanoparticles as a potential vaccine. Nanoscale Res Lett. 2016;11(1):338. doi:10.1186/s11671-016-1554-y
89. Wang J, Wang S, Ye T, et al. Choice of nanovaccine delivery mode has profound impacts on the intralymph node spatiotemporal distribution and immunotherapy efficacy. Adv Sci (Weinh). 2020;7(19):2001108. doi:10.1002/advs.202001108
90. Kokate RA, Thamake SI, Chaudhary P, et al. Enhancement of anti-tumor effect of particulate vaccine delivery system by ‘bacteriomimetic’ CpG functionalization of poly-lactic-co-glycolic acid nanoparticles. Nanomedicine (Lond). 2015;10(6):915–929. doi:10.2217/nnm.14.144
91. Shi R, Hong L, Wu D, et al. Enhanced immune response to gastric cancer specific antigen peptide by coencapsulation with CpG oligodeoxynucleotides in nanoemulsion. Cancer Biol Ther. 2005;4(2):218–224. doi:10.4161/cbt.4.2.1472
92. Bavananthasivam J, Alkie TN, Astill J, et al. In ovo administration of Toll-like receptor ligands encapsulated in PLGA nanoparticles impede tumor development in chickens infected with Marek’s disease virus. Vaccine. 2018;36(28):4070–4076. doi:10.1016/j.vaccine.2018.05.091
93. Geary SM, Hu Q, Joshi VB, Bowden NB, Salem AK. Diaminosulfide based polymer microparticles as cancer vaccine delivery systems. J Control Release. 2015;220(Pt B):682–690. doi:10.1016/j.jconrel.2015.09.002
94. Zhao Y, Wang H, Yang Y, et al. Mannose-modified liposome co-delivery of human papillomavirus type 16 E7 peptide and CpG oligodeoxynucleotide adjuvant enhances antitumor activity against established large TC-1 grafted tumors in mice. Int J Nanomedicine. 2020;15:9571–9586. doi:10.2147/IJN.S275670
95. Sainz V, Moura LIF, Peres C, et al. α-Galactosylceramide and peptide-based nano-vaccine synergistically induced a strong tumor suppressive effect in melanoma. Acta Biomater. 2018;76:193–207. doi:10.1016/j.actbio.2018.06.029
96. Kokate RA, Chaudhary P, Sun X, et al. Rationalizing the use of functionalized poly-lactic-co-glycolic acid nanoparticles for dendritic cell-based targeted anticancer therapy. Nanomedicine (Lond). 2016;11(5):479–494. doi:10.2217/nnm.15.213
97. Mueller M, Reichardt W, Koerner J, Groettrup M. Coencapsulation of tumor lysate and CpG-ODN in PLGA-microspheres enables successful immunotherapy of prostate carcinoma in TRAMP mice. J Control Release. 2012;162(1):159–166. doi:10.1016/j.jconrel.2012.06.015
98. Zhang L, Zhu G, Mei L, et al. Self-assembled DNA immunonanoflowers as multivalent CpG nanoagents. ACS Appl Mater Interfaces. 2015;7(43):24069–24074. doi:10.1021/acsami.5b06987
99. Zhu G, Hu R, Zhao Z, Chen Z, Zhang X, Tan W. Noncanonical self-assembly of multifunctional DNA nanoflowers for biomedical applications. J Am Chem Soc. 2013;135(44):16438–16445. doi:10.1021/ja406115e
100. Zhu G, Mei L, Vishwasrao HD, et al. Intertwining DNA-RNA nanocapsules loaded with tumor neoantigens as synergistic nanovaccines for cancer immunotherapy. Nat Commun. 2017;8(1):1482. doi:10.1038/s41467-017-01386-7
101. Ni Q, Zhang F, Liu Y, et al. A bi-adjuvant nanovaccine that potentiates immunogenicity of neoantigen for combination immunotherapy of colorectal cancer. Sci Adv. 2020;6(12):eaaw6071. doi:10.1126/sciadv.aaw6071
102. Lv Y, Hu R, Zhu G, et al. Preparation and biomedical applications of programmable and multifunctional DNA nanoflowers. Nat Protoc. 2015;10(10):1508–1524. doi:10.1038/nprot.2015.078
103. Qian Y, Jin H, Qiao S, et al. Targeting dendritic cells in lymph node with an antigen peptide-based nanovaccine for cancer immunotherapy. Biomaterials. 2016;98:171–183. doi:10.1016/j.biomaterials.2016.05.008
104. Kuai R, Ochyl LJ, Bahjat KS, Schwendeman A, Moon JJ. Designer vaccine nanodiscs for personalized cancer immunotherapy. Nat Mater. 2017;16(4):489–496. doi:10.1038/nmat4822
105. Cai Z, Xin F, Wei Z, et al. Photodynamic therapy combined with antihypoxic signaling and CpG adjuvant as an in situ tumor vaccine based on metal-organic framework nanoparticles to boost cancer immunotherapy. Adv Healthc Mater. 2020;9(1):e1900996. doi:10.1002/adhm.201900996
106. Liu H, Chen H, Liu Z, et al. Therapeutic nanovaccines sensitize EBV-associated tumors to checkpoint blockade therapy. Biomaterials. 2020;255:120158. doi:10.1016/j.biomaterials.2020.120158
107. Zang X, Zhao X, Hu H, Qiao M, Deng Y, Chen D. Nanoparticles for tumor immunotherapy. Eur J Pharm Biopharm. 2017;115:243–256. doi:10.1016/j.ejpb.2017.03.013
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.