Back to Journals » Infection and Drug Resistance » Volume 13
Comparison and Correlation of Genetic Variability of the HBV Pre-S Region in HIV/HBV Co-Infected Patients: Quasispecies Perspective
Authors Nie Y ![]() , Deng X, Lan Y, Li L, Li F, Hu F
, Deng X, Lan Y, Li L, Li F, Hu F
Received 27 August 2020
Accepted for publication 17 November 2020
Published 1 December 2020 Volume 2020:13 Pages 4327—4334
DOI https://doi.org/10.2147/IDR.S278415
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Yuan Nie,* Xizi Deng,* Yun Lan, Linghua Li, Feng Li, Fengyu Hu
Research Institute, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, Guangzhou, Guangdong, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Fengyu Hu; Feng Li
Research Institute, Guangzhou Eighth People’s Hospital, Guangzhou Medical University, No. 627, Dongfeng Road, Yuexiu District, Guangzhou, Guangdong 510030, People’s Republic of China
Email [email protected]; [email protected]
Background: Human immunodeficiency virus (HIV)/hepatitis B virus (HBV) co-infection can accelerate HBV-induced liver disease. A previous study showed that variation in the HBV pre-S region and quasispecies heterogeneity (Sn, mean genetic distance, dS, dN, and dS/dN) are both related to HBV-induced terminal liver disease in HBV mono-infection. Currently, data are lacking on quasispecies variation of the HBV pre-S region in HIV/HBV co-infection. Investigating the quasispecies variation of the HBV pre-S region and its related factors in HIV/HBV co-infection will help to better explore the pathogenic mechanism of HIV/HBV co-infection.
Methods: According to the HIV antibody results obtained before treatment, chronic HBV-infected patients were divided into HIV/HBV co-infected and HBV mono-infected groups. The clinical characteristics of all patients were collected, and DNA was extracted from the serum. The HBV pre-S region was amplified by nested PCR and was further TA cloned. BioEdit software 7.0 was used for sequence alignment with reference to the standard sequence of the matched HBV genotype. We used 1:1 propensity score matching (PSM) to control for baseline confounding factors between the two groups.
Results: After 1:1 PSM, we identified 100 patients with similar propensities: 50 HIV/HBV co-infected patients and 50 HBV mono-infected patients. HBV quasispecies indices were lower in the HIV/HBV co-infected group than those in the HBV mono-infected group. A significant correlation was observed between all quasispecies indices and soluble cluster of differentiation 163 (sCD163) and interleukin-18 (IL-18) in the HIV/HBV co-infected group; however, this phenomenon was not found in the HBV mono-infected group.
Conclusion: Combined HIV infection reduces quasispecies heterogeneity in the HBV pre-S region, and the quasispecies heterogeneity is related to the sCD163 and IL-18 levels.
Keywords: HIV, HBV, quasispecies, pre-S region, inflammation
Introduction
Hepatitis B virus (HBV)/human immunodeficiency virus (HIV) are common viruses because both HIV and HBV can be transmitted through blood, from mother to child, and via injection of drugs and sexual contact. Previous studies have indicated that 10% of HIV-infected patients have HIV/HBV co-infection. In some countries, such as China, the HIV/HBV co-infection rate can be as high as 20%.1,2 HIV/HBV co-infection can accelerate the development of liver disease and delay immune reconstitution during highly active antiretroviral therapy (HAART). HIV/HBV co-infection is a global public health problem, and terminal liver disease is the leading cause of death in HIV-infected patients. However, the pathogenic mechanism of HIV/HBV co-infection is not yet clear and requires further study.2,3
The HBV genome consists of incomplete circular double-stranded DNA replicated by an error-prone polymerase through an RNA intermediate. As documented in many studies, numerous viral variants that differ in genetic sequence occur during HBV replication in vivo, which are called quasispecies.4,5 Since viral mutants are constantly produced by dynamic conditions under host immune pressure, quasispecies variability can provide insight into viral-host interactions, especially the interactions between viral evolution and host immunity.6 The emergence of HBV quasispecies is an important cause of continuous infection with
HBV is a reflection of infection evolution and is related to the emergence of drug resistance, the effects of antiviral treatment, and the development of liver disease.6–8 The intracellular persistence of HBV leads to liver disease through the host immune response.8 The intensity of the immune response can be critical to disease progression and to the complexity and diversity of virus strains. The HBV genome consists of four open reading frames (ORFs), namely, the pre-core/core, Polymerase, X and pre-S/S ORFs. The HBV pre-S region was reported to contain abundant immunogenic epitopes and functional regional sites and is related to the host’s immune response.9–11 Previous studies have indicated that, before treatment, the HBV pre-S deletion A1762T/G1764A is more common in HIV/HBV co-infected patients than in HBV mono-infected patients; however, these studies were only based on direct sequencing, and quasispecies studies have been very limited.12,13 When the expression product of the HBV pre-S region is deleted, affinity with the corresponding cell receptors is reduced, so that HBV escapes immune surveillance. Investigation of the quasispecies variation of the HBV pre-S region in HIV/HBV co-infected patients and the factors related to quasispecies evolution might help to better understand the pathogenic mechanisms of HIV/HBV co-infection.
Materials and Methods
Study Subjects
Patients hospitalized with chronic HBV infection (CHB) from April 2018 to December 2019 were included. CHB patients were confirmed to be positive for HBsAg for more than 6 months. HIV-infected patients were confirmed by ELISA and Western blot assays. The exclusion criteria were as follows: (1) patients with anti-HBV treatment and anti-HIV treatment (HAART); (2) patients with terminal liver disease or cancer; (3) patients with other types of hepatitis virus infection (such as HAV, HCV, HDV); (4) patients with other apparent opportunistic infections; (5) patients who were <18 years old, >65 years old, pregnant or lactating women; (6) patients with cardiovascular disease or renal failure. CHB patients were separated into HIV/HBV co-infection and HBV mono-infection groups. The cross-sectional study protocol conformed to the Declaration of Helsinki and was approved by the Institutional Ethics Committee of Guangzhou Eighth People’s Hospital (No. 20180307). Written informed consent was obtained from patients.
Serological Examination
An ELISA was used to detect HBsAg HBeAg/anti-HBe (Zhong Shan Da An Company, Guangdong, China). A COBAS TaqMan HBV Test was used to detect the HBV DNA load. An AU-2700 automatic biochemical detector was used to detect alanine aminotransferase (ALT) and aspartate aminotransferase (AST) levels. BD Multitest CD3/CD8/CD45/CD4 reagents were used with a BD FACSCanto II flow cytometer to determine the absolute CD4 cell count and the CD8 cell count (Invitrogen, USA). Soluble cluster of differentiation 14 (sCD14), soluble cluster of differentiation 163 (sCD163), interleukin-18 (IL-18), interferon-inducible protein-10 (IP-10), soluble cluster of differentiation 27 (sCD27), and monocyte chemoattractant protein 1 (MCP-1) levels were determined by ELISAs (Invitrogen, USA), according to the manufacturer’s guidelines. Routine blood indices, including platelet counts, were measured by an XE-5000 automatic blood analyzer.
Cloning and Sequencing
Total DNA was extracted from a 200 µL serum sample collected from each patient using a QIAamp DNA Mini Kit. The primers for the first round of nested PCR were 5ʹ-GCCTCATTTTGYGGGTCACCATATTC-3ʹ and 5ʹ-GGGTCACCATATTCTTGGGAACAAGA-3ʹ. The primers for the second round of nested PCR were 5ʹ-GGGTCACCATATTCTTGGGAACAAGA-3ʹ and 5ʹ-AATTCGTTGACANACTTTCCAATCAAT-3ʹ. The first round of PCR amplification was performed in a 25 μL reaction system using 4 μL of DNA template (DNA extracted from serum). The amplification conditions (20 cycles) of first round were as follows: denaturation at 98°C for 10 s, annealing at 56°C for 30 s, and extension at 72°C for 30 s. The second round of PCR amplification was performed in a 100 μL reaction system using 25 μL of DNA template. The amplification conditions (35 cycles) of the second round were as follows: denaturation at 98°C for 10 s, annealing at 56°C for 30 s, and extension at 72°C for 30 s. The primer-amplified region is shown in Scheme 1. The purified product was ligated into a pMD-19T/A vector and transformed into JM109 cells. Nearly 50 clones were selected and sequenced.
Sequence Analysis
Multiple alignments were performed using by BioEdit 7.0 software, and viral quasispecies heterogeneity was evaluated using two parameters: complexity and diversity. Quasispecies complexity is the index of distribution of different variants in the virus population, which is expressed by the standardized Shannon entropy (Sn). The Sn was calculated by the following formula: Sn=-∑I (pi ln pi)/ln N, where pi is the frequency of each quasispecies, and N is the total number of quasispecies. MEGA 6.0 software was used to evaluate the quasispecies diversity, mean genetic distance, number of synonymous substitutions per synonymous site (dS), and number of nonsynonymous substitutions per nonsynonymous site (dN). The mean genetic distances at the nucleotide and amino acid levels were calculated under the Kimura 2 parameter model and Jones-Taylor-Thornton model, respectively. The Jukes-Cantor mode was used to calculate the dS and dN.14,15
Statistical Analysis
We used 2 groups of 1:1 nearest neighbor propensity score matching (PSM) to control baseline confounding factors (including sex, age, HBeAg, HBV genotype, ALT, AST, platelets, AST to platelet ratio (APRI), HBV DNA, HBsAg).16 Statistical analyses were performed using SPSS software version 22.0 (SPSS Inc, Chicago, IL, USA), and graphs were produced using GraphPad Prism 8.0 software. Continuous variables are described as the median (interquartile range [IQR]) because of an abnormal distribution according to the results of normality testing (Kolmogorov–Smirnov test). Categorical variables are described as the frequency (percentage [%]). Continuous variables were compared using the Mann–Whitney U-test, and categorical variables were compared using the Chi-squared test or Fisher’s exact test. Correlation coefficient analyses were performed by Spearman rank correlation. All of the statistical tests were two-sided, and a value of P<0.050 was considered statistically significant.
Results
Demographic Data
In the study, 170 CHB patients successfully underwent PCR product extraction, cloning, and sequencing, including 92 patients in the HIV/HBV co-infected group and 78 patients in the HBV mono-infected group. A total of 100 patients were included in the final analysis after 1:1 PSM, including 50 HIV/HBV co-infected patients and 50 HBV mono-infected patients. The characteristics of the two groups of patients are listed in Table 1. In the HIV/HBV co-infected group, the HIV genotype mainly consisted of HIV-1, and the HIV-infected patients were in the AIDs phase (CD4<200 cell/mL); 35/15 had B/C genotypes, and 30 patients were HBeAg positive. The CD4 cell count and CD4/CD8 ratio in the HIV/HBV co-infected group were 75.5 (0.75–139) cells/mL and 0.110 (0.058–0.198) before HAART. Among HBV mono-infected patients, 36/14 had B/C genotypes, and 32 patients were HBeAg positive.
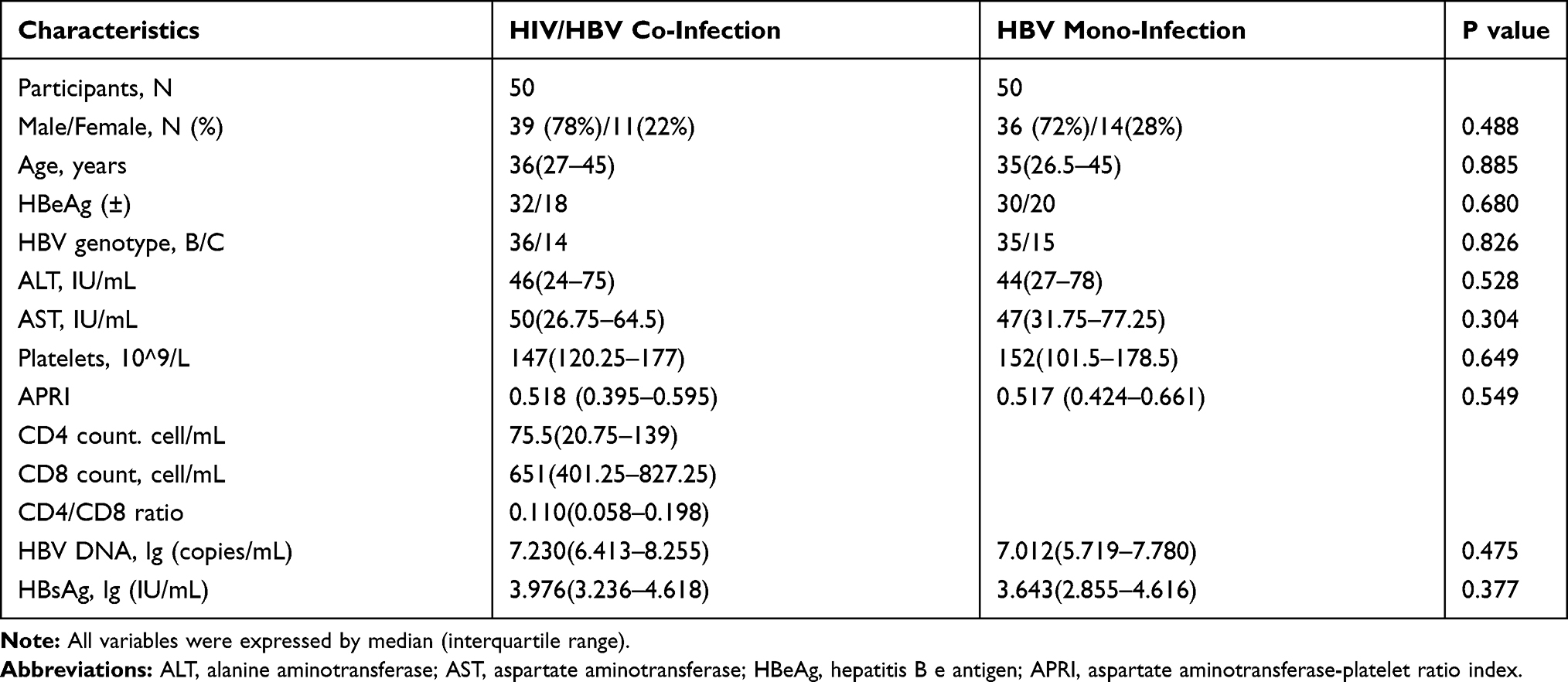 |
Table 1 Demographic and Clinical Features of Patients |
HBV Quasispecies Complexity and Diversity for Different Infectious Statuses
HBV quasispecies complexity and diversity were assessed according to the sequences obtained from TA clones. The HBV quasispecies complexity at the nucleotide and amino acid levels in the HIV/HBV co-infected group (0.671 (0.467–0.779); 0.479 (0.406–0.577)) was significantly lower than that in the HBV mono-infected group (0.760 (0.665–0.838); 0.610 (0.466–0.779)) (P<0.05). In terms of quasispecies diversity, the dN and dS of HBV quasispecies in the HIV/HBV co-infected group (6.564 (3.991–8.476); 6.361 (3.904–8.213)) were also significantly lower than those in the HBV mono-infected group (8.997 (8.248–10.820); 8.072 (5.840–9.085)) (P<0.05). The dN/dS in the HIV/HBV co-infected group (0.957 (0.600–1.259)) was significantly higher than that in the HBV mono-infected group (0.788 (0.597–1.007)) (P<0.05). No significant difference in genetic distance was found at the nucleotide level between two groups; however, the data showed a smaller genetic distance in the HIV/HBV co-infected group. The genetic distance at the amino acid level was significantly smaller in the HIV/HBV co-infected group (6.305 (4.232–8.264)) than in the HBV mono-infected group (8.070 (6.518–9.596)) (P<0.05).
Correlation of Quasispecies Indices and Immune Cell Count, Liver Enzymes, and Inflammatory Cytokines
No correlations were found between any quasispecies indices and CD4 cell counts in the HIV/HBV co-infected group (all P>0.05). Additionally, there was no correlation between quasispecies indices and CD8 cell count or CD4/CD8 ratio (all P>0.05). No significant correlations were found between quasispecies indices and ALT or AST in both the HIV/HBV co-infected group and the HBV mono-infected group (all P>0.05).
In the HIV/HBV co-infected group, significant correlations were observed between all 7 quasispecies indices and sCD163 (r=0.400, P=0.007; r=0.387, P=0.009; r=0.465, P=0.001; r=0.309, P=0.039; r=0.369, P=0.013; r=0.443, P=0.002; r=0.488, P=0.001). Significant correlations were found between quasispecies indices and IL-18 (r=0.383, P=0.042; r=0. 0.397, P=0.028; r=0.310, P=0.045; r=0.367, P=0.040; r=0.276, P=0.049; r=0.327, P=0.041; r=0.355, P=0.040). No correlations were found between quasispecies indices and sCD14 or MCP-1 (P>0.05). Significant correlations were found between sCD27 and complexity at the nucleotide level, genetic distance at the nucleotide level, genetic distance at the amino acid level, dN, and dN/dS. The correlations between sCD27 and complexity at the amino acid level and dS were not statistically significant. The correlation analysis results of the HIV/HBV co-infected group are listed in Table 2. In the HBV mono-infected group, no correlations were found between any of the quasispecies indices and inflammatory cytokines (P>0.05). The correlation analysis results of the HBV mono-infected group are listed in Supplementary Table 1. A heat map of correlation coefficients between quasispecies indices and the clinical parameters and inflammatory cytokines is shown in Figure 1.
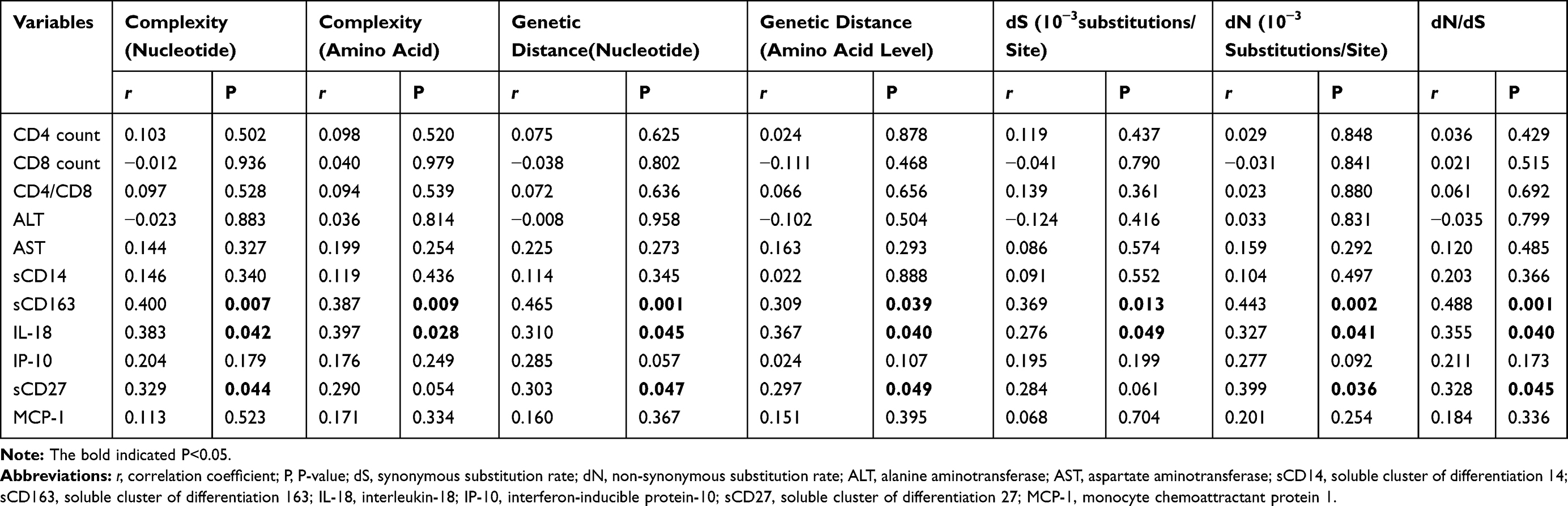 |
Table 2 Correlation of Quasispecies Indexes and Immune Cells Count, Liver Enzymes, Inflammatory Cytokines of HIV/HBV Co-Infection |
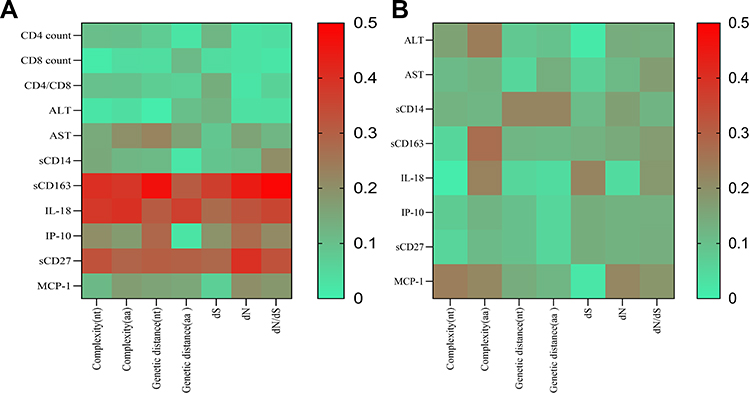 |
Figure 1 Heat map of correlation coefficient between quasispecies indexes and clinical parameters, inflammatory cytokines. (A) Heat map of correlation coefficient for HIV/HBV co-infection; (B) Heat map of correlation coefficient for HBV mono-infection. |
Discussion
The high replication capacity of HBV and the lack of proofreading activity of HBV polymerase lead to the development of quasispecies complexity and diversity during replication. One study showed a strong negative correlation between survival time of HIV-infected patients and HBV co-infection, but the effect of HIV co-infection on HBV, and especially HBV evolution, is unknown.17 A previous study used directed sequence analysis to demonstrate that HIV/HBV co-infection may affect HBV evolution and that HBV sequences in HIV/HBV co-infected patients have a greater number of variants than those in HBV mono-infected patients.12,13 Another previous study used direct sequencing analysis to show that pre-C A1762T/G1764A and pre-S deletions are more common in HIV/HBV co-infected patients than in HBV mono-infected patients in China.12 However, because only a limited number of studies of minor variants have been conducted, they might be overlooked, which could exclude useful information about quasispecies. Cassino L et al examined viral quasispecies in 8 patients infected with HBV and 9 patients infected with HIV/HBV before treatment and at 3 years.18 A previous study analyzing the pre-C/C and S genome regions suggested that low quasispecies complexity and diversity and few evolutionary characteristics existed in HIV/HBV co-infected patients. Mondal R’s study also compared the diversity of the pre-S region in 32 patients with HBV genotype D from India and suggested that the diversity in HIV/HBV co-infected patients is lower than that in HBeAg-positive and HBeAg-negative patients.19 However, their study was limited to Western populations, and patients in the two groups were limited in number and not matched regarding basic information. In this study, analysis of 2004 and 1992 clone sequences from HIV/HBV co-infected patients and HBV mono-infected patients, respectively, showed that the quasispecies complexity and diversity of the pre-S region are lower in HIV/HBV co-infected patients than in HBV mono-infected patients, which is an important finding. Although our study is inconsistent with the study of Cassino L in the gene region and genotype, the results were consistent in that the quasispecies complexity and diversity were lower in HIV/HBV co-infected patients than in HBV mono-infected patients.
The evolutionary characteristics of quasispecies are related to the emergence of drug resistance, effect of antiviral treatment, and development of liver disease.7,20 One study indicated that some miRNAs of HIV-infected patients were significantly correlated with viral load and CD4 cell count, and miRNAs seem to influence the clinical progression of HIV-1 infection in naive patients.21,22 The results could suggest that HIV infection has a significant impact on the state of the body. We performed further analyses to explore the correlation between quasispecies indices and acquired immunity (including the CD4 cell count and CD4/CD8 ratio) and innate immunity. Our results showed a significant correlation between all quasispecies indices and sCD163 and IL-18 in the HIV/HBV co-infected group after analyzing the correlation between quasispecies indices and clinical parameters. The results suggested that HIV co-infection might impact the molecular evolution of HBV, which in turn is associated with inflammatory activation. sCD163, also called the hemoglobin (Hb) scavenger receptor, is expressed only in monocyte-macrophage cell lines, and the upregulated expression of this receptor is one of the major changes in the macrophage switch to alternative activated phenotypes in inflammation.23 The sCD163 level is related to HIV replication activity, monocyte-macrophage immune activation, and the degree of liver inflammation.24 Mononuclear macrophages have been indicated to be an important reservoir of HIV.25 Therefore, it is speculated that a correlation exists between quasispecies indices and sCD163 in HIV/HBV co-infected patients, which could be caused by cross-reacting with immune activation. IL-18 is secreted primarily by innate immune cells and is related to the degree of liver inflammation and the effect of antiviral therapy. A previous study showed a significant correlation between IL-18 promoter polymorphisms and the progression of HBV-related liver disease.26 As liver inflammation decreases, IL-18 also decreases and can be used as an immune agent to promote specific cellular immune functions.27,28 In HIV/HBV co-infected patients, HBeAg conversion after anti-HBV antiviral regimens is closely related to IL-18 levels before HAART.29 The correlation analysis results indicate that innate immune activation could play an important role in HBV evolution in HIV/HBV co-infection.
This study also had some limitations. First, this was a cross-sectional study, and the dynamic changes in quasispecies heterogeneity, especially after HAART, could not be evaluated. Second, the main route of HBV in China is mother-to-child transmission, and the predominant HBV genotypes are B and C, which are different from those in Western countries. Finally, no liver biopsy or liver stiffness results were obtained before HAART; therefore, the APRI and platelets were used as indicators in this study.30,31
In conclusion, this study investigated the differences in quasispecies complexity and diversity of the HBV pre-S region between HBV mono-infected and HIV/HBV co-infected patients as well as the correlation of quasispecies indices with immune cell counts, liver enzymes, and inflammatory cytokines. Low quasispecies complexity and diversity were found in HIV/HBV co-infected patients, and significant correlations were observed between all quasispecies indices and sCD163 and IL-18. These results suggest that combined HIV infection reduces quasispecies heterogeneity in the HBV pre-S region, and quasispecies heterogeneity is related to the sCD163 and IL-18 levels.
Ethical Approval
This study was performed in accordance with the institutional committee protocols of Guangzhou Eighth People’s Hospital (No. 20180307), and informed consent was obtained from all patients.
Funding
We would like to thank the Guangzhou Science and Technology Plan Project (No. 202002030028); National Grand Program on Key Infectious Disease Control (2017ZX10202101; 2017ZX10202203-004-002; 2018ZX10301404-003-002); National Natural Science Foundation of China (No. 81670536; 81770593); and Major Special Project for Innovation of Health Care in Guangzhou (No. 201704020219).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Kourtis AP, Bulterys M, Hu DJ, et al. HIV–HBV Coinfection — A Global Challenge. N Engl J Med. 2012;366(19):1749–1752. doi:10.1056/NEJMp1201796
2. Zhang F, Zhu H, Wu Y, et al. HIV, hepatitis B virus, and hepatitis C virus co-infection in patients in the China National Free Antiretroviral Treatment Program, 2010–12: a retrospective observational cohort study. Lancet Infect Dis. 2014;14(11):1065–1072. doi:10.1016/S1473-3099(14)70946-6
3. Tanday S. HBV and HCV co-infection increases cancer risk in HIV patients[J]. Lancet Oncol. 2016;17(11):e484. doi:10.1016/S1470-2045(16)30512-5
4. RODRIGUEZ-FRIAS F, et al. Quasispecies structure, cornerstone of hepatitis B virus infection: mass sequencing approach[J]. World J Gastroenterol. 2013;19(41):6995–7023. doi:10.3748/wjg.v19.i41.6995
5. Jazayeri SM, Alavian SM, Carman WF. Hepatitis B virus: origin and evolution[J]. J Viral Hepat. 2010;17(4):229–235. doi:10.1111/j.1365-2893.2009.01193.x
6. Domingo E, Gomez J. Quasispecies and its impact on viral hepatitis[J]. Virus Res. 2007;127(2):131–150. doi:10.1016/j.virusres.2007.02.001
7. Liu F, Chen L, Yu Dm, et al. Evolutionary patterns of hepatitis B virus quasispecies under different selective pressures: correlation with antiviral efficacy[J]. Gut. 2011;60(9):1269–1277. doi:10.1136/gut.2010.226225
8. Yang Z-T, Huang S-Y, Chen L, et al. Characterization of Full-Length Genomes of Hepatitis B Virus Quasispecies in Sera of Patients at Different Phases of Infection[J]. J Clin Microbiol. 2015;53(7):2203–2214. doi:10.1128/JCM.00068-15
9. Desmond CP, Bartholomeusz A, Gaudieri S, et al. A systematic review of T-cell epitopes in hepatitis B virus: identification, genotypic variation and relevance to antiviral therapeutics[J]. Antivir Ther. 2008;13(2):161–175.
10. Li X, Qin Y, Liu Y, et al. PreS deletion profiles of hepatitis B virus (HBV) are associated with clinical presentations of chronic HBV infection[J]. J Clin Virol. 2016;82:27–32. doi:10.1016/j.jcv.2016.06.018
11. Chen BF, Liu CJ, Jow GM, et al. High prevalence and mapping of pre-S deletion in hepatitis B virus carriers with progressive liver diseases[J]. Gastroenterology. 2006;130(4):1153–1168. doi:10.1053/j.gastro.2006.01.011
12. Li KW, Kramvis A, Liang S, et al. Higher prevalence of cancer related mutations 1762T/1764A and PreS deletions in hepatitis B virus (HBV) isolated from HBV/HIV co-infected compared to HBV-mono-infected Chinese adults[J]. Virus Res. 2017;227:88–95. doi:10.1016/j.virusres.2016.10.002
13. Audsley J, Littlejohn M, Yuen L, et al. HBV mutations in untreated HIV-HBV co-infection using genomic length sequencing[J]. Virology. 2010;405(2):539–547. doi:10.1016/j.virol.2010.06.038
14. Tamura K, Stecher G, Peterson D, et al. MEGA6: molecular Evolutionary Genetics Analysis version 6.0[J]. Mol Biol Evol. 2013;30(12):2725–2729. doi:10.1093/molbev/mst197
15. Tamura K, Peterson D, Peterson N, et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods[J]. Mol Biol Evol. 2011;28(10):2731–2739. doi:10.1093/molbev/msr121
16. Ali MS, Groenwold RHH, Belitser SV, et al. Reporting of covariate selection and balance assessment in propensity score analysis is suboptimal: a systematic review[J]. J Clin Epidemiol. 2015;68(2):112–121. doi:10.1016/j.jclinepi.2014.08.011
17. Maracy MR, Mostafaei S, Moghoofei M, et al. Impact of HIV risk factors on survival in Iranian HIV-infected patients: A Bayesian approach to retrospective cohort[J]. HIV AIDS Rev. 2017;2:100–106. doi:10.5114/hivar.2017.68117
18. Cassino L, Laufer N, Salomon H, et al. Hepatitis B precore/core promoter mutations in isolates from HBV-monoinfected and HBV–HIV coinfected patients: A 3-yr prospective study. J Clin Virol. 2009;46(4):354–359. doi:10.1016/j.jcv.2009.09.015
19. Mondal RK, Khatun M, Ghosh S, et al. Immune-driven adaptation of hepatitis B virus genotype D involves preferential alteration in B-cell epitopes and replicative attenuation—an insight from human immunodeficiency virus/hepatitis B virus coinfection. Clin Microbiol Infect. 2015;21(7):710–711. doi:10.1016/j.cmi.2015.03.004
20. Shen T. Hepatitis B virus genetic mutations and evolution in liver diseases[J]. World J Gastroenterol. 2014;20(18):5435–5441. doi:10.3748/wjg.v20.i18.5435
21. Moghoofei M, Bokharaei-Salim F, Esghaei M, et al. microRNAs 29, 150, 155, 223 level and their relation to viral and immunological markers in HIV-1 infected naive patients[J]. Future Virol. 2018;13(9):637–645. doi:10.2217/fvl-2018-0055
22. Yousefpouran S, Mostafaei S, Manesh PV. The assessment of selected MiRNAs profile in HIV, HBV, HCV, HIV/HCV, HIV/HBV Co-infection and elite controllers for determination of biomarker. Microb Pathog. 2020;147:104355. doi:10.1016/j.micpath.2020.104355
23. Etzerodt A, Moestrup Sk. CD163 and inflammation: biological, diagnostic, and therapeutic aspects[J]. Antioxid Redox Signal. 2013;18(17):2352–2363. doi:10.1089/ars.2012.4834
24. Hiraoka A, Horiike N, Akbar SMF, et al. Soluble CD163 in patients with liver diseases: very high levels of soluble CD163 in patients with fulminant hepatic failure[J]. J Gastroenterol. 2005;40(1):52–56. doi:10.1007/s00535-004-1493-8
25. Campbell JH, Hearps AC, Martin GE, et al. The importance of monocytes and macrophages in HIV pathogenesis, treatment, and cure[J]. AIDS. 2014;28(15):2175–2187. doi:10.1097/QAD.0000000000000408
26. Migita K, Sawakami-Kobayashi K, Maeda Y, et al. Interleukin-18 promoter polymorphisms and the disease progression of Hepatitis B virus-related liver disease[J]. Transl Res. 2009;153(2):91–96. doi:10.1016/j.trsl.2008.11.008
27. Kimura K, Kakimi K, Wieland S, et al. Interleukin-18 inhibits hepatitis B virus replication in the livers of transgenic mice[J]. J Virol. 2002;76(21):10702–10707. doi:10.1128/JVI.76.21.10702-10707.2002
28. Lee M-O, Choi Y-H, Shin E-C, et al. Hepatitis B virus X protein induced expression of interleukin 18 (IL-18): a potential mechanism for liver injury caused by hepatitis B virus (HBV) infection[J]. J Hepatol. 2002;37(3):380–386. doi:10.1016/S0168-8278(02)00181-2
29. Li Y, Xie J, Wang H, et al. Elevated pre-treatment IL-18 level is associated with HBeAg seroconversion in HIV–HBV coinfection. Antivir Ther. 2017;22(6):523–527. doi:10.3851/IMP3136
30. Liu R, Guo J, Lu Y, et al. Changes in APRI and FIB-4 in HBeAg-negative treatment-naive chronic hepatitis B patients with significant liver histological lesions receiving 5-year entecavir therapy[J]. Clin Exp Med. 2019;19(3):309–320. doi:10.1007/s10238-019-00560-z
31. Laffi G, Marra F, Gresele P, et al. Evidence for a storage pool defect in platelets from cirrhotic patients with defective aggregation[J]. Gastroenterology. 1992;103(2):641–646.
 © 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2020 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.