Back to Journals » Infection and Drug Resistance » Volume 16
Comparative Genomics Identified PenR E151V Substitution Associated with Carbapenem-Resistance Burkholderia cepacia Complex and a Novel Burkholderia cepacia Complex Specific OXA-1043 Subgroup
Authors Liao YC ![]() , Huang YT, Tseng CH, Liu CW
, Huang YT, Tseng CH, Liu CW ![]() , Liu PY
, Liu PY
Received 30 April 2023
Accepted for publication 10 August 2023
Published 28 August 2023 Volume 2023:16 Pages 5627—5635
DOI https://doi.org/10.2147/IDR.S418969
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Professor Suresh Antony
Ya-Chun Liao,1 Yao-Ting Huang,2,* Chien-Hao Tseng,1 Chia-Wei Liu,1 Po-Yu Liu1,3,4,*
1Division of Infectious Diseases, Department of Internal Medicine, Taichung Veterans General Hospital, Taichung, Taiwan; 2Department of Computer Science and Information Engineering, National Chung Cheng University, Chiayi, Taiwan; 3Department of Post-Baccalaureate Medicine, National Chung Hsing University, Taichung, Taiwan; 4Genome Center for Infectious Diseases, Taichung Veterans General Hospital, Taichung, Taiwan
*These authors contributed equally to this work
Correspondence: Po-Yu Liu, Division of Infectious Diseases, Department of Internal Medicine, Taichung Veterans General Hospital, No. 1650, Sec. 4, Taiwan Blvd., Xitun District, Taichung, Taiwan, Tel +886-4-2359-2525, Fax +886-4-2359-5046, Email [email protected]
Purpose: Burkholderia cepacia complex (Bcc) is a known significant opportunistic pathogen causing morbidity and mortality, particularly in those with cystic fibrosis, chronic granulomatous disease, or immunocompromising host. Mortality of Bcc bloodstream infections among non-cystic fibrosis patients remained high. The antibiotic treatment for Bcc infection is quite challenging due to its intrinsic resistance to most antibiotics, and the resistance to carbapenems was the biggest concern among them. We aimed to realize the mechanism of carbapenem resistance in Bcc.
Patients and Methods: Ten strains of Bcc were identified by the MALDI-TOF MS, and the drug susceptibility test was using VITEK 2 system. The Burkholderia cepacia complex genomes were sequenced via Nanopore GridIon. We also downloaded another ninety-five strains of Bcc from the National Center for Biotechnology Information database to evaluate the divergence between carbapenem-resistance and carbapenem-sensitive strains.
Results: The genetic organization between carbapenem-sensitive and carbapenem-resistant strains of Bcc showed no difference. However, in the carbapenem-sensitive strain, E151V substitution in PenR was detected. In addition, a novel specific OXA family subgroup, blaOXA-1043 in Burkholderia cenocepacia was discovered.
Conclusion: The E151V substitution in PenR may be associated with carbapenem-sensitive in Bcc. Moreover, the V151E mutation in PenR may be related to the activation of PenB, leading to Bcc resistance to carbapenems. Besides, a novel OXA family subgroup, blaOXA-1043, was found in Burkholderia cenocepacia, which differs from the previous OXA family.
Keywords: Burkholderia cepacia complex, Burkholderia cenocepacia, carbapenem-resistant, whole-genome sequencing
Introduction
Burkholderia cepacia complex (Bcc), formerly known as Pseudomonas cepacia, is a group of aerobic, non-fermenting gram-negative bacilli, composed of more than twenty species. It was first discovered by Walter Burkholder in 1949, causing onion skin rot. In the 1950s, Bcc was described as a human pathogen and later reported as a cystic fibrosis respiratory pathogen, associated with accelerated decline in pulmonary functions and reduced survival, especially among those with more advanced lung disease or who had undergone lung transplantation.1 It is also an important pathogen that causes morbidity and mortality in hospitalized patients, especially those with cystic fibrosis, chronic granulomatous disease, or immunocompromising host.2,3 Mortality of Bcc bloodstream infections among non-cystic fibrosis patients in a 17-year nationwide study was 16%, 25%, and 36% at 14, 30, and 90 days, respectively.3
Treatment options for Bcc remain limited due to its intrinsic resistance to most antibiotics. Recommendation regimens for Bcc infections include carbapenem, fluoroquinolone, minocycline, and trimethoprim-sulfamethoxazole.4 However, a study of Bcc isolates from cystic fibrosis patients revealed 23% of strains were susceptible to ceftazidime, 38% of strains were susceptible to minocycline, 5% of strains were susceptible to trimethoprim-sulfamethoxazole, and 26% of strains susceptible to meropenem.5 Due to limiting antibiotic choice for Bcc infection and the high mortality rate in Bcc bloodstream infection, understanding the mechanism of carbapenem resistance in Bcc helps physicians select a proper antibiotic treatment and may contribute to developing new treatment options in the future. However, the mechanism of carbapenem resistance remained unclear.
As far as we know, carbapenemase production was vital in carbapenem-resistance gram-negative bacteria. The induction of PenB, also a class A β-lactamases family, led to the carbapenem-resistant in Burkholderia species.6 Another research suggested PenB conferred β-lactam resistance in Bcc and established that carbapenem resistance in Burkholderia ubonensis is due to an inducible class A PenB β-lactamase.7 However, another previous study in 2018, using an agar diffusion method to analyze the antibiotic resistance (across sixteen antibiotics and combinations) in five clinical and one environmental Burkholderia cenocepacia strains, the members of Bcc, showed a similar number of antibiotic resistances. Interestingly, the MC0-3 strain showed carbapenem sensitive, while most other strains exhibited carbapenem-resistant.8 Other mechanisms may regulate the expression of PenB, and impact the phenotype of carbapenem resistance in Bcc. Hence, we would like to figure out if the potential amino acid change in these two strains is associated to the phenotype between carbapenem-sensitive and carbapenem-resistant Bcc, despite both strains having the same genetic organization.
This study analyzed ten strains of Bcc, comparing the arrangement of amino acids between carbapenem-sensitive and carbapenem-resistant strains, and also retrieved another ninety-five strains of Bcc from the National Center for Biotechnology Information database to evaluate the divergence between the carbapenem-sensitive and carbapenem-resistant Bcc to figure out the change of amino acids alignment may associate to carbapenem resistance in Bcc.
Materials and Methods
Isolates
Ten strains of carbapenem-resistant Bcc isolated from blood culture in the past 5 years in Taichung Veterans General Hospital were included in the study. Preliminary identification was performed by MALDI-TOF MS (matrix-assisted laser desorption-ionization time-of-flight, mass spectrometry), and all protocols were performed according to the manufacturer’s instructions. The rest of ninety-five strains of Bcc were retrieved from the National Center for Biotechnology Information (NCBI) database.
Antimicrobial Susceptibility Testing
Antimicrobial susceptibility test (AST) was using VITEK 2 system. Once a reliable direct identification by MALDI-TOF MS was achieved, the pellet of the third aliquot was diluted in a saline solution 0.45% and its density adjusted to 0.5 McFarland turbidity standard. Then, it was used for a direct AST by the VITEK-2 Compact System. All protocols were performed according to the manufacturer’s instructions.
Genomic DNA Extraction, Sequencing, and Annotation
The Bcc genomes were sequenced using Nanopore GridIon. The DNA library was prepared using the rapid barcoding kit (SQK-RBK004) following the manufacturer’s instructions, and the sequencing was performed in an R9.4.1 flow cell for 48 h. The sequenced reads were basecalled using Guppy 4.4.2. Adaptor sequences left in the reads were trimmed using Porechop (v0.2.4). The clean reads were assembled into contigs via Flye (v2.7),9 and the remaining sequencing errors were sequentially polished by Racon, Medaka, and Homopolish.10,11 The assembled contigs were classified into chromosomes and plasmids via both NCBI BLAST and PlasmidFinder. Protein-coding genes and coding and non-coding RNAs in chromosomes and plasmids were annotated via the NCBI Prokaryotic Genome Annotation Pipeline (PGAP).12 In addition, we used the Protein Variation Effect Analyzer (PROVEAN) to find the possible functional mutations between carbapenem-sensitive and carbapenem-resistant Bcc.
Resistance Genes Analysis
Antibiotic-resistant genes (ARGs) were predicted by aligning protein-coding genes against the Comprehensive Antibiotic Resistance Database (CARD).13 Only ARGs with alignment coverage greater than 90% were retained. Efflux pumps were excluded from ARG analysis. Beta lactamases were separately predicted by the curated hidden Markov models in NCBI AMRFinderPlus.14 To assess whether the E151V was a random point mutation or a divergent evolution between resistant and sensitive Bcc strains, we also downloaded PenR sequences of ninety-five strains of Bcc in NCBI. The phylogeny of PenR and OXA was carried out by Mega and visualized by interactive Tree Of Life (iTOL).
Results
Genomic Organization of Bcc Strains Revealed No Difference
A total of one hundred and five strains of Bcc were included in our study. Nine strains of carbapenem-resistant and one strain of carbapenem-sensitive Bcc, identified by MALDI-TOF MS, were included (Table 1), and the rest of the ninety-five strains were retrieved from the NCBI database. The drug susceptibility test of these ten strains of Bcc is shown in Table 1. Both the genetic organization of carbapenem-sensitive and carbapenem-resistant strains of Bcc were identical (Figure 1), with PenB followed by PenR, and the latter is a LysR-type regulator that governs the expression of PenB.
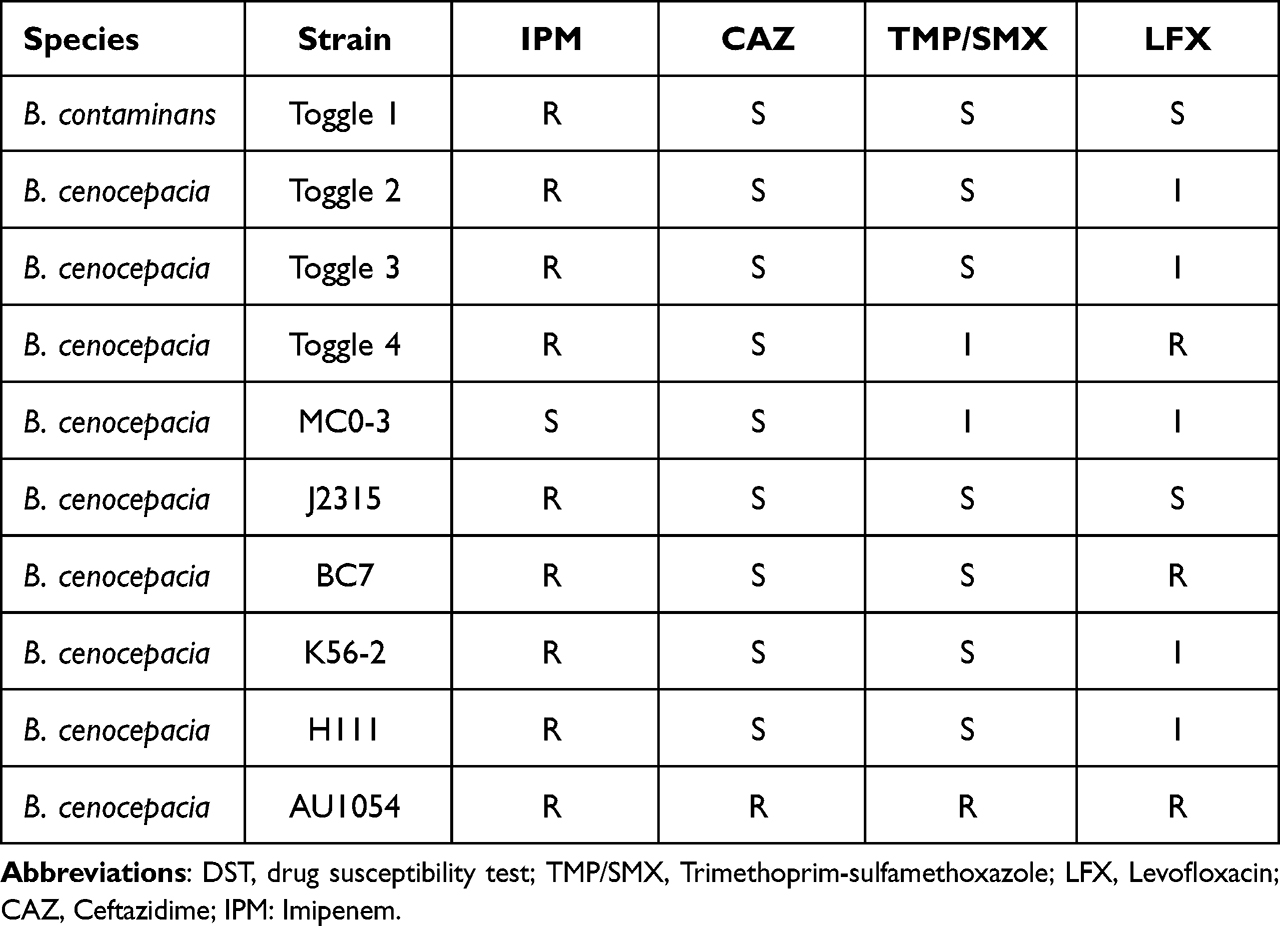 |
Table 1 DST of Different Antibiotics in Different Strains of B. cenocepacia |
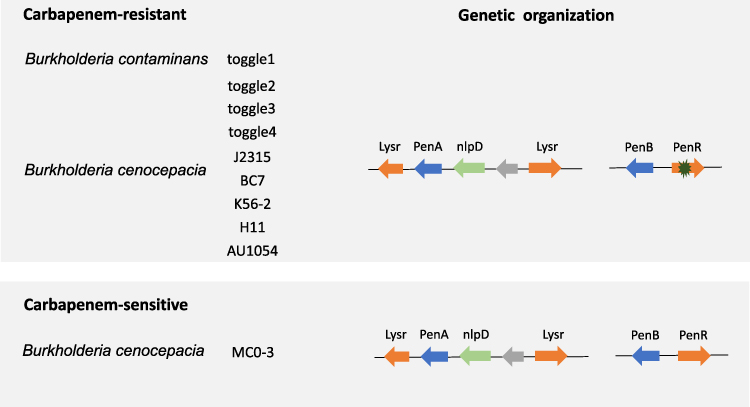 |
Figure 1 Genetic organization of carbapenem-resistant and carbapenem-sensitive in different strains of Burkholderia cepacia complex. Carbapenem-resistant Burkholderia cepacia complex include Burkholderia cenocepacia strain toggle 2, toggle 3, toggle 4, J2315, BC7, K56-2, H111 and AU1054, and Burkholderia contaminans strain toggle 1. The only carbapenem-sensitive strain is Burkholderia cenocepacia strain MC0-3. However, the genetic organization shows no difference between carbapenem-resistant and carbapenem-sensitive strain. |
E151V Mutation in Bcc Led to Sensitive of Carbapenem
Multiple sequence alignments of PenR and PenB revealed carbapenem resistance-associated mutation (Figure 2a and b). The functional importance of these mutations was analyzed by PROVEAN, which revealed only one deleterious substitution at E151V in PenR (Table 2). No other deleterious mutations were found in PenB system (Table S1). Therefore, the initial analysis suggested that E151V mutation may be associated with sensitivity to carbapenem in Bcc.
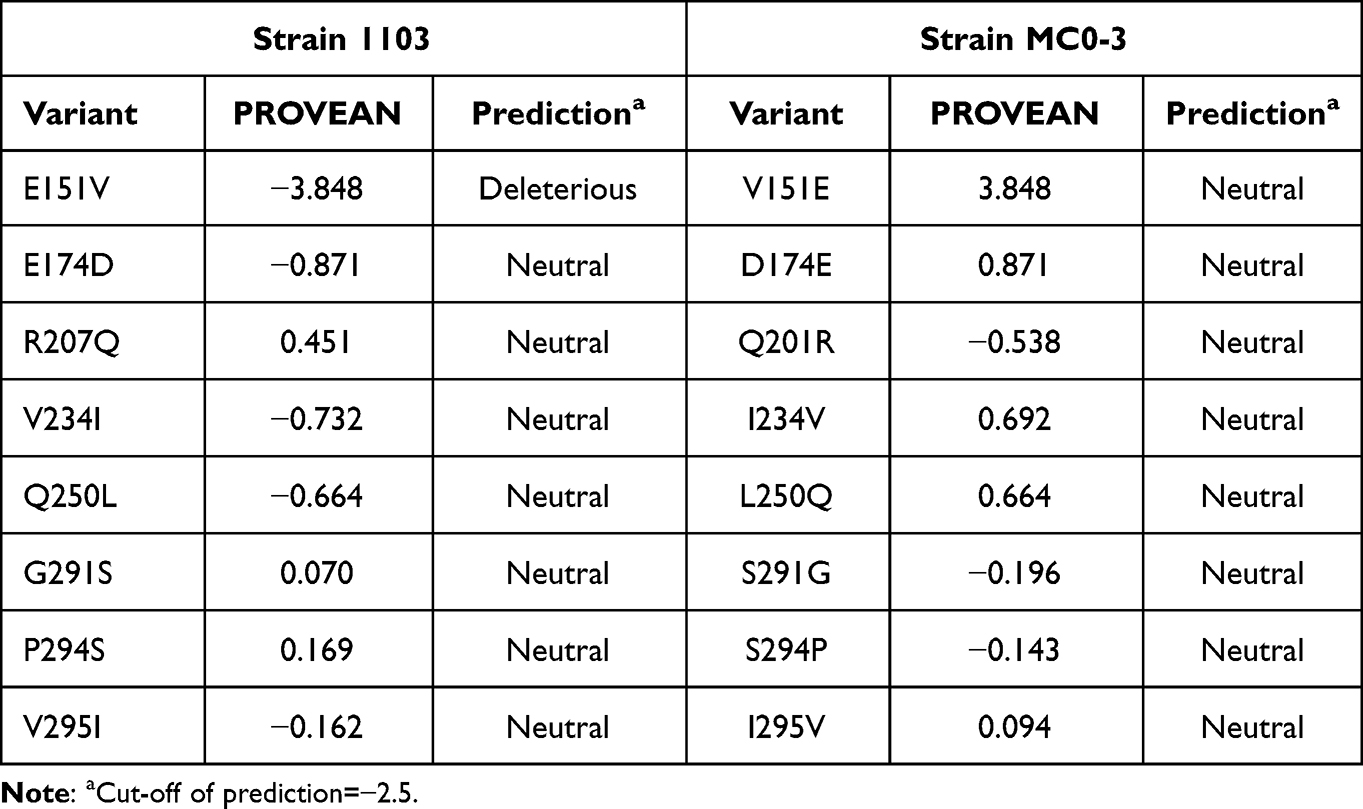 |
Table 2 PROVEAN of B. cenocepacia Strain 1103 and Strain MC0-3 in PenR |
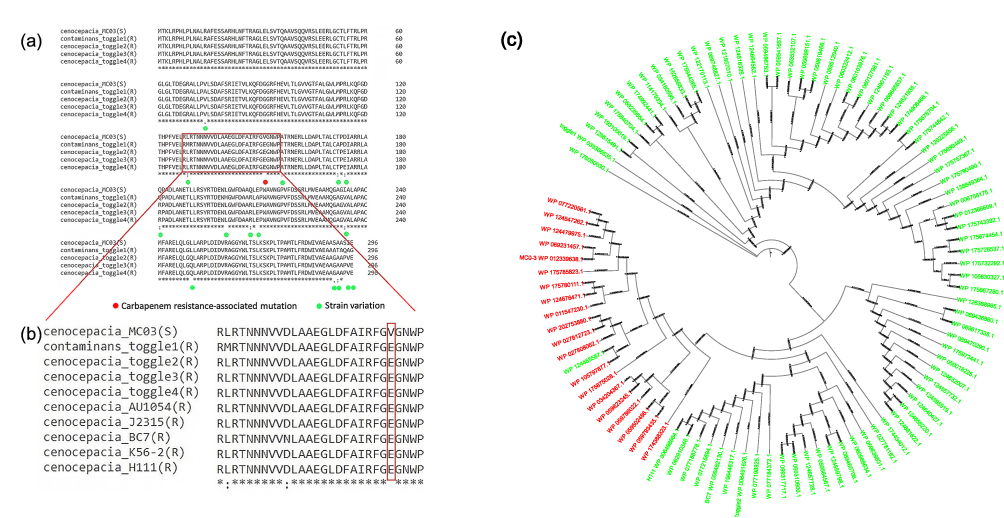 |
Figure 2 (a and b) Multiple sequence alignment of PenR in different strains of Burkholderia cepacia complex. The red box in (a) focuses on the possible deleterious mutations in multiple sequence alignments of PenR, and the red box in (b) indicated the deleterious substitution at E151V in PenR. Red dots represent carbapenem resistance associated mutations. Green dots indicate strain variation. E151V mutation in Burkholderia cepacia complex remained sensitive to carbapenem. The asterisk (*) indicated the position without mutation or variation of the multiple sequence alignments in PenR. (c) Phylogenetic tree of Burkholderia spp. Ninety-five strains of Burkholderia spp. are analyzed and the phylogenetic trees E151V mutation consisted in other carbapenem-sensitive strains of Burkholderia spp. (red color). |
Divergent Evolution of Resistant and Sensitive Bcc Strains
To assess whether the E151V mutation is a random effect or significantly associated with carbapenem-resistance in Bcc, we downloaded ninety-five PenR sequences in NCBI and reconstructed their phylogeny (see Methods). Two lineages, one corresponding to the E151V (sensitive) mutations and the other belonging to the V151E (resistant) strains, were clustered in two clades (Figure 2c). While the majority of PenR carried the resistant PenR alleles, a minority contained the sensitive one. Hence, our results suggested that the allele status mutation at E151V of PenR may not activate PenB sequences, causing the specific Bcc strain (MC0-3) to remain sensitive to carbapenem.
Novel OXA Gene Subgroup
A novel OXA gene is found in Burkholderia cenocepacia stains toggle 2, toggle 3, and toggle 4, which is named as blaOXA-1043 (OXA-1043 family class D β-lactamase). This new class D β-lactamase, designated OXA-1043, has a total of 7414 genes and 7082 coding sequences with protein. It was first identified on the chromosome 2 of the Burkholderia cenocepacia. We used the Mega to carry out the phylogeny of OXA family, which was visualized by iTOL. Phylogenetic tree and taxonomic ranks also reveal that blaOXA-1043 is different from previous OXA family, suggesting a new subgroup of oxacillinase (Figure 3).
 |
Figure 3 Phylogenetic tree of OXA β-lactamases family in Burkholderia spp. OXA family groups are listed in the phylogenetic tree, and the different OXA β-lactamases families are identified in different colors. A novel specific OXA family subgroup, blaOXA-1043 was found in Burkholderia cenocepacia, marked in blue. |
Discussion
In our study, the Bcc strain remaining sensitive to carbapenem might associate with E151V mutation in PenR, and the V151E mutation in PenR might associate with the activation of PenB, causing carbapenem resistance in Bcc. The divergence between carbapenem-sensitive and carbapenem-resistant Bcc showed two lineages rather than a random mutation. In addition, a novel specific OXA family subgroup, blaOXA-1043, was discovered in Burkholderia cenocepacia.
As far as we know, extended-spectrum β-lactamase (ESBL)-producing and carbapenem-resistant bacteria have become a global issue. Antibiotic options are limited for these pathogens. Carbapenems are the more effective antibiotic options for ESBL than colistin.15 However, when it comes to carbapenem-resistant bacteria, such as carbapenem-resistant Acinetobacter baumannii (CRAB), a long course of colistin therapy resulted in a lower 30-day mortality rate in critically ill patients.16 The primary mechanism of carbapenem resistance in carbapenem-resistant bacteria is the production of carbapenemases. Carbapenemase is divided into classes A, B, and D by using the Ambler classification method.17 Burkholderia species produces at least three active β-lactamase, including two class A enzymes (PenA and PenB) and one class C enzyme (AmpC).7 A newly identified β-lactamase, PenB, demonstrated its hydrolysis activity toward ß-lactams and the distribution of its gene among Bcc isolates in 2009.6 Focus on the carbapenem resistance in Bcc had been reported in Burkholderia ubonensis, a member of the Bcc, which is also due to an inducible class A PenB β-lactamase by mutational analyses. In the mutation analysis, a previous study compared the PenB protein expression between carbapenem-resistant strain Bu278 and the carbapenem-susceptible B. ubonensis strain MSMB2152, and found that PenB protein activity was much higher in the carbapenem-susceptible strain.9 Similar to many Gram-negative bacteria, a β-lactamase is induced in response to peptidoglycan perturbation, resulting in transpeptidase inhibition by β-lactam antibiotics. The system in Burkholderia cenocepacia is PenR/PenB.18,19 PenR is a LysR-type protein encoded by a gene that is transcribed divergently from penB.9 Moreover, the mutations in the key cell wall-recycling enzyme AmpD (N-acetyl-anhydromuramyl-L-alanine amidase) led to the induction of two β-lactamases, AmpC and PenB. However, in Burkholderia cenocepacia and many Bcc species, ampC and its regulator, ampR, named penR in Burkholderia genomes are not associated, sharing a divergent promoter, as in other previously studied species. In Bcc species, penB is instead associated with the ampR homolog.18
Other class A β-lactamases genes could also be identified in other Enterobacteriaceae. A plasmid-mediated carbapenemase gene, designating Klebsiella pneumoniae carbapenemase (KPC), was first reported in the Klebsiella pneumoniae strain in 2001.20 Since then, blaKPC has spread widely in the United States and South America, causing outbreaks of KPC-producing Enterobacteriaceae to be reported in most European regions.21,22 As for the class B β-lactamases, blaIMP has spread throughout Japan since the IMP-1 was first discovered among 105 strains of Serratia marcescens in Okazaki in 1993.23 Besides, blaNDM-associated carbapenem-resistant KPN was first reported in India and mainly spread in Asia.24,25 In recent years, NDM has become the second most common carbapenemase found among carbapenem-resistant Enterobacteriaceae in China, and blaNDM is more prevalent in Escherichia coli.26,27
Our study also found that both carbapenem-resistant and carbapenem-sensitive Bcc strains had the same genomic organization, and penB was associated with the penR homolog, which matched the previous studies. Moreover, the V151E mutation on PenR was found in carbapenem-resistant strains. The PenR sequences of other ninety-five strains of Bcc from the NCBI database also clustered in two lineages, one corresponding to the E151V mutations (sensitive strains) and the other belonging to V151E mutations (resistant strains). Therefore, the V151E mutation on PenR may be associated with the activation of PenB, causing carbapenem-resistant in Bcc.
The Class D β-lactamases were another important mechanism of beta-lactam resistance, which composed of 14 families and the majority of the member enzymes are included in the OXA family.28 This group of enzymes presents a distinguished ability to hydrolyse cloxacillin/oxacillin as well as carbenicillin. In 1985, the first OXA-encoding gene was found in an Acinetobacter baumannii isolate from the United Kingdom and designated blaOXA-23.29 Immediately after, the number of OXA family members gradually demonstrated in the Enterobacteriaceae, including OXA-23-like, OXA-48-like, OXA-40-like, OXA-51- like, and OXA-58-like.30 Mostly, the subfamilies of a cluster are intrinsic to the same bacterial genus.28 The OXA-22-like enzyme is innate in Ralstonia spp.31 The OXA-42-like enzymes are intrinsic to Burkholderia pseudomallei.32 The members of the OXA-114-like, OXA-258-like, and OXA-243-like subfamilies are innate in Achromobacter spp. The OXA-12-like-associated subfamilies are inherent in Aeromonas spp.33,34 The OXA-55-like and revised OXA-48-like subfamilies are innate in Shewanella spp.35–37 In our study, a new subfamily of OXA family, blaOXA-1043, is inherent in Burkholderia cenocepacia, which has not been reported before. The discovery of the blaOXA-1043 gene in Burkholderia species might imply the possibility of resistance to carbapenem, which may affect antibiotic treatment choice in clinical medicine.
Finally, some limitations of our study existed. First, only a small number of strains of Bcc were used in this study, although we tried to obtain more Bcc strains from the NCBI database, which may only represent some species of Bcc. Second, we only used PROVEAN to discover the possible point mutation between carbapenem-resistant and carbapenem-sensitive strains. The genotype mutation may not entirely correspond to the phenotype in carbapenem-resistant strains of Bcc. Retrospectively, the electrophoretic mobility shift assay (EMSA) is used to detect protein complexes with nucleic acids, and it is the core technology underlying a wide range of qualitative and quantitative analyses for the characterization of interacting systems. The assay is highly sensitive using radioisotope-labeled nucleic acids, allowing the assays to be performed with small protein and nucleic acid concentrations (0.1 nM or less) and small (20 mL or less) sample volumes.38 In previous studies, EMSA has been widely applied in detecting antimicrobial resistance among multiple microorganisms, such as demonstrating that the AtrR conferred clinically significant azole resistance in Aspergillus fumigatus.39 Further functional assay, such as EMSA, to demonstrate the impact of the V151E in PenR on PenB activation may be required to investigate whether this mutation responds to the resistance phenotype of carbapenem in Bcc. In addition, further experiments to test the activity of the novel OXA gene subgroups in other bacterial strains were also needed to strengthen our findings.
Conclusion
Our study found that the E151V mutation in PenR might be associated with carbapenem-sensitive in Bcc, which may help us understand the mechanism of carbapenem-resistant Bcc more. And the V151E mutation in PenR may relate to the activation of PenB, causing carbapenem resistance in Bcc. However, our results need further functional assays to prove that the carbapenem-resistant Bcc could detect the E151V substitution. Besides, blaOXA-1043, a novel specific OXA family, was first found in Burkholderia cenocepacia, which also shows a trend of specificity in different bacterial species like the previous OXA family.
Acknowledgments
The authors would like to thank all colleagues and students who contributed to this study. Our thanks go to Dr. Liu for his guidance, and Professor Huang for his expertise. The abstract of this paper was presented at the IDWeek as an abstract presentation with interim findings. The poster’s abstract was published in “Poster Abstracts” in Open Forum Infectious Disease: https://doi.org/10.1093/ofid/ofac492.221.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by the Ministry of Science and Technology under Grant 109-2221-E-194 -038 -MY3 and NSTC 112-2314-B-075A-006; Taichung Veterans General Hospital under CVGH-1123901C, CVGH-1123901D, TCVGH-PU1128105, TCVGH-1117201D, TCVGH-1127201D.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Isles A, Maclusky I, Corey M, et al. Pseudomonas cepacia infection in cystic fibrosis: an emerging problem. J Pediatr. 1984;104(2):206–210. doi:10.1016/s0022-3476(84)80993-2
2. Baul SN, De R, Mandal PK, Roy S, Dolai TK, Chakrabarti P. Outbreak of Burkholderia cepacia infection: a systematic study in a hematolooncology unit of a tertiary care hospital from Eastern India. Mediterr J Hematol Infect Dis. 2018;10(1):e2018051. doi:10.4084/MJHID.2018.051
3. El Chakhtoura NG, Saade E, Wilson BM, Perez F, Papp-Wallace KM, Bonomo RA. A 17-year nationwide study of Burkholderia cepacia complex bloodstream infections among patients in the United States Veterans Health Administration. Clin Infect Dis. 2017;65(8):1253–1259.
4. Gilbert DN, Chambers HF, Saag MS, Pavia A, Boucher T, Helen W. The Sanford Guide to Antimicrobial Therapy.
5. Zhou J, Chen Y, Tabibi S, Alba L, Garber E, Saiman L. Antimicrobial susceptibility and synergy studies of Burkholderia cepacia complex isolated from patients with cystic fibrosis. Antimicrob Agents Chemother. 2007;51(3):1085–1088. doi:10.1128/AAC.00954-06
6. Poirel L, Rodriguez-Martinez JM, Plesiat P, Nordmann P. Naturally occurring Class A ss-lactamases from the Burkholderia cepacia complex. Antimicrob Agents Chemother. 2009;53(3):876–882. doi:10.1128/AAC.00946-08
7. Somprasong N, Hall CM, Webb JR, et al. Burkholderia ubonensis meropenem resistance: insights into distinct properties of class A beta-Lactamases in Burkholderia cepacia complex and Burkholderia pseudomallei complex bacteria. mBio. 2020;11(2):10–128.
8. Bodilis J, Denet E, Brothier E, Graindorge A, Favre-Bonte S, Nazaret S. Comparative genomics of environmental and clinical Burkholderia cenocepacia strains closely related to the highly transmissible epidemic ET12 lineage. Front Microbiol. 2018;9:383. doi:10.3389/fmicb.2018.00383
9. Kolmogorov M, Yuan J, Lin Y, Pevzner PA. Assembly of long, error-prone reads using repeat graphs. Nat Biotechnol. 2019;37(5):540–546. doi:10.1038/s41587-019-0072-8
10. Vaser R, Sovic I, Nagarajan N, Sikic M. Fast and accurate de novo genome assembly from long uncorrected reads. Genome Res. 2017;27(5):737–746. doi:10.1101/gr.214270.116
11. Huang Y-T, Liu P-Y, Shih P-W. Homopolish: a method for the removal of systematic errors in nanopore sequencing by homologous polishing. Genome Biol. 2021;22(1):95. doi:10.1186/s13059-021-02282-6
12. Tatusova T, DiCuccio M, Badretdin A, et al. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016;44(14):6614–6624. doi:10.1093/nar/gkw569
13. Alcock BP, Raphenya AR, Lau TTY, et al. CARD 2020: antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020;48(D1):D517–D525. doi:10.1093/nar/gkz935
14. Feldgarden M, Brover V, Gonzalez-Escalona N, et al. AMRFinderPlus and the Reference Gene Catalog facilitate examination of the genomic links among antimicrobial resistance, stress response, and virulence. Sci Rep. 2021;11(1):12728. doi:10.1038/s41598-021-91456-0
15. Katip W, Yoodee J, Uitrakul S, Oberdorfer P. Efficacy of loading dose colistin versus carbapenems for treatment of extended spectrum beta lactamase producing Enterobacteriaceae. Sci Rep. 2021;11(1):18. doi:10.1038/s41598-020-78098-4
16. Katip W, Rayanakorn A, Oberdorfer P, Taruangsri P, Nampuan T. Short versus long course of colistin treatment for carbapenem-resistant A. baumannii in critically ill patients: a propensity score matching study. J Infect Public Health. 2023;16(8):1249–1255. doi:10.1016/j.jiph.2023.05.024
17. Ambler RP. The structure of beta-lactamases. Philos Trans R Soc Lond B Biol Sci. 1980;289(1036):321–331. doi:10.1098/rstb.1980.0049
18. Hwang J, Kim HS. Cell wall recycling-linked coregulation of AmpC and PenB beta-Lactamases through ampD Mutations in Burkholderia cenocepacia. Antimicrob Agents Chemother. 2015;59(12):7602–7610. doi:10.1128/AAC.01068-15
19. Rhodes KA, Schweizer HP. Antibiotic resistance in Burkholderia species. Drug Resist Updat. 2016;28:82–90. doi:10.1016/j.drup.2016.07.003
20. Yigit H, Queenan AM, Anderson GJ, et al. Novel carbapenem-hydrolyzing beta-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob Agents Chemother. 2001;45(4):1151–1161. doi:10.1128/AAC.45.4.1151-1161.2001
21. Munoz-Price LS, Poirel L, Bonomo RA, et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect Dis. 2013;13(9):785–796. doi:10.1016/S1473-3099(13)70190-7
22. Patel G, Bonomo RA. “Stormy waters ahead”: global emergence of carbapenemases. Front Microbiol. 2013;4:48. doi:10.3389/fmicb.2013.00048
23. Ito H, Arakawa Y, Ohsuka S, Wacharotayankun R, Kato N, Ohta M. Plasmid-mediated dissemination of the metallo-beta-lactamase gene blaIMP among clinically isolated strains of Serratia marcescens. Antimicrob Agents Chemother. 1995;39(4):824–829. doi:10.1128/AAC.39.4.824
24. Yong D, Toleman MA, Giske CG, et al. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009;53(12):5046–5054. doi:10.1128/AAC.00774-09
25. Dortet L, Poirel L, Nordmann P. Worldwide dissemination of the NDM-type carbapenemases in Gram-negative bacteria. Biomed Res Int. 2014;2014:249856. doi:10.1155/2014/249856
26. Zhang Y, Wang Q, Yin Y, et al. Epidemiology of Carbapenem-resistant Enterobacteriaceae infections: report from the China CRE Network. Antimicrob Agents Chemother. 2018;62(2):10–128.
27. Zhang R, Liu L, Zhou H, et al. Nationwide surveillance of clinical Carbapenem-resistant Enterobacteriaceae (CRE) Strains in China. EBioMedicine. 2017;19:98–106. doi:10.1016/j.ebiom.2017.04.032
28. Yoon EJ, Jeong SH. Class D beta-lactamases. J Antimicrob Chemother. 2021;76(4):836–864. doi:10.1093/jac/dkaa513
29. Donald HM, Scaife W, Amyes SG, Young HK. Sequence analysis of ARI-1, a novel OXA beta-lactamase, responsible for imipenem resistance in Acinetobacter baumannii 6B92. Antimicrob Agents Chemother. 2000;44(1):196–199. doi:10.1128/AAC.44.1.196-199.2000
30. Evans BA, Amyes SG. OXA beta-lactamases. Clin Microbiol Rev. 2014;27(2):241–263. doi:10.1128/CMR.00117-13
31. Nordmann P, Poirel L, Kubina M, Casetta A, Naas T. Biochemical-genetic characterization and distribution of OXA-22, a chromosomal and inducible class D beta-lactamase from Ralstonia (Pseudomonas) pickettii. Antimicrob Agents Chemother. 2000;44(8):2201–2204. doi:10.1128/AAC.44.8.2201-2204.2000
32. Niumsup P, Wuthiekanun V. Cloning of the class D beta-lactamase gene from Burkholderia pseudomallei and studies on its expression in ceftazidime-susceptible and -resistant strains. J Antimicrob Chemother. 2002;50(4):445–455. doi:10.1093/jac/dkf165
33. Walsh TR, Hall L, MacGowan AP, Bennett PM. Sequence analysis of two chromosomally mediated inducible beta-lactamases from Aeromonas sobria, strain 163a, one a class D penicillinase, the other an AmpC cephalosporinase. J Antimicrob Chemother. 1995;36(1):41–52. doi:10.1093/jac/36.1.41
34. Antonelli A, D’Andrea MM, Montagnani C, et al. Newborn bacteraemia caused by an Aeromonas caviae producing the VIM-1 and SHV-12 beta-lactamases, encoded by a transferable plasmid. J Antimicrob Chemother. 2016;71(1):272–274. doi:10.1093/jac/dkv304
35. Heritier C, Poirel L, Nordmann P. Genetic and biochemical characterization of a chromosome-encoded carbapenem-hydrolyzing ambler class D beta-lactamase from Shewanella algae. Antimicrob Agents Chemother. 2004;48(5):1670–1675. doi:10.1128/AAC.48.5.1670-1675.2004
36. Poirel L, Heritier C, Nordmann P. Chromosome-encoded ambler class D beta-lactamase of Shewanella oneidensis as a progenitor of carbapenem-hydrolyzing oxacillinase. Antimicrob Agents Chemother. 2004;48(1):348–351. doi:10.1128/AAC.48.1.348-351.2004
37. Antonelli A, Di Palo DM, Galano A, et al. Intestinal carriage of Shewanella xiamenensis simulating carriage of OXA-48-producing Enterobacteriaceae. Diagn Microbiol Infect Dis. 2015;82(1):1–3. doi:10.1016/j.diagmicrobio.2015.02.008
38. Hellman LM, Fried MG. Electrophoretic mobility shift assay (EMSA) for detecting protein-nucleic acid interactions. Nat Protoc. 2007;2(8):1849–1861. doi:10.1038/nprot.2007.249
39. Paul S, Stamnes M, Thomas GH, et al. AtrR is an essential determinant of azole resistance in Aspergillus fumigatus. mBio. 2019;10(2):e02563–18. doi:10.1128/mBio.02563-18
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.