Back to Journals » Infection and Drug Resistance » Volume 16
Characterization of ST11 and ST15 Carbapenem-Resistant Hypervirulent Klebsiella pneumoniae from Patients with Ventilator-Associated Pneumonia
Authors Song S, Zhao S, Wang W, Jiang F, Sun J, Ma P, Kang H
Received 20 June 2023
Accepted for publication 24 August 2023
Published 8 September 2023 Volume 2023:16 Pages 6017—6028
DOI https://doi.org/10.2147/IDR.S426901
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Shuang Song,1,2,* Shulong Zhao,2,* Wei Wang,1 Fei Jiang,2 Jingfang Sun,2 Ping Ma,1,2 Haiquan Kang2
1Medical Technology School, Xuzhou Medical University, Xuzhou, People’s Republic of China; 2Department of Clinical Laboratory, Affiliated Hospital of Xuzhou Medical University, Xuzhou, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Haiquan Kang, Department of Laboratory Medicine, Affiliated Hospital of Xuzhou Medical University, Xuzhou, Jiangsu, 221000, People’s Republic of China, Tel +86-516 85802759, Email [email protected] Ping Ma, Medical Technology School, Xuzhou Medical University, Department of Laboratory Medicine, Xuzhou, People’s Republic of China, Tel +86-516 85807017, Email [email protected]
Background: The prevalence of carbapenem-resistant hypervirulent Klebsiella pneumoniae (hv-CRKP) is a serious public threat globally. Here, we performed clinical, molecular, and phenotypic monitoring of hv-CRKP strains isolated from the intensive care unit (ICU) to offer evidence for prevention and control in hospitals.
Methods: Data analysis of ICU patients suffering from ventilator-associated pneumonia (VAP) because of hv-CRKP infection, admitted at the Chinese Teaching Hospital between March 2019 and September 2021 was performed. Patients’ antibiotic-resistance genes, virulence-associated genes, and capsular serotypes of these isolates were detected. Homology analysis of the strains was performed by MLST and PFGE. Six different strains were tested for their virulence traits using the serum killing test and the Galleria mellonella infection assay. For whole genome sequencing, KP3 was selected as a representative strain.
Results: Clinical data of 19 hv-CRKP-VAP patients were collected and their hv-CRKP were isolated, including 10 of ST11-KL64, 4 of ST15-KL112, 2 of ST11-KL47, 1 of ST15-KL19, 1 of ST17-KL140, and 1 of ST48-KL62. Four ST15 and 8 ST11 isolates revealed high homology, respectively. Most strains carried the carbapenemase gene blaKPC-2 (14/19, 73.68%), followed by blaOXA-232 (4/19, 21.05%). All strains were resistant to almost all the antibiotics except polymyxin and tigacycline. Ten patients were treated with polymyxin or tigacycline based on their susceptibility results, and unfortunately 6 patients died. All strains exhibited a hyper-viscous phenotype, and the majority (17/19, 89.47%) of them contained rmpA and rmpA2. The serum killing test showed that KP9 was resistant to normal healthy serum, others were intermediately or highly sensitive. G. mellonella larvae infection assay suggested that the strains in this study were hypervirulent.
Conclusion: This study highlights the dominant strain and molecular epidemiology of hv-CRKP in a hospital in China. We should pay more attention to the effect of hv-CRKP on VAP, strengthen monitoring and control transmission.
Keywords: Klebsiella pneumoniae, hypervirulent, carbapenem-resistant, ventilator-associated pneumonia
Introduction
Klebsiella pneumoniae is a gram-negative opportunistic pathogen that can cause both community and hospital-acquired pneumonia, urinary tract infections, and bacteremia primarily in immunocompromised patients.1 Most experts agree that carbapenems are an effective treatment for serious infections brought on by Enterobacteriaceae that produce extended-spectrum beta-lactamases (ESBLs).2–4 Because of the wide and irrational use of antibiotics, the number of carbapenemase-resistant Enterobacteriaceae (CRE) infections is increasing per year and antibiotic resistance has become an international problem.5,6 During 2005–2020, the China Antimicrobial Surveillance Network (CHINET, http://www.chinets.com) reported that K. pneumoniae resistance rates to imipenem and meropenem have rapidly increased from 3% and 2.9% to 23.2% and 24.2% respectively. There has been a significant increase in respiratory tract infections in intensive care units (ICUs) due to carbapenem-resistant K. pneumoniae (CRKP), which is considered a severe danger to public health as a pathogen associated with hospital-acquired infections (HAIs).7–10 It is reported that 9–27% of ICU patients suffer from ventilator-associated pneumonia (VAP).11 Meanwhile, treatment of CRKP-induced VAP in ICU patients is difficult because of the lack of multiple therapeutic drugs.
Hypervirulent K. pneumoniae (hvKP) is the primary cause of community-acquired illnesses such as meningitis, necrotizing fasciitis, endophthalmitis, liver abscesses, and pneumonia in people of all ages and health statuses. It is associated with severe morbidity and mortality and has been reported to have an increasing incidence rate globally.12–15 When the string test is positive for hvKP, it is defined to have a hyper-viscous phenotype. K1 and K2 are the most frequent capsular loci for hvKP.12 ST23 and ST258 strains account for the majority of hvKP strains.16 Antibiotics usually work well on hvKP, however infections brought on by carbapenem-resistant hvKP (hv-CRKP) have recently been documented increasingly frequently17,18 To the best of our knowledge, the emergence of such a high-virulent, drug-resistant pathogen poses an even more significant challenge to clinical patient safety.
This investigation was conducted on the VAP outbreaks caused by ST11 and ST15 hv-CRKP in a teaching hospital’s Emergency Intensive Care Unit (EICU) in China. Furthermore, the clinical characteristics of patients as well as the outcome, resistance mechanisms, virulence characteristics, and genomic features of these strains were analyzed.
Materials and Methods
Isolates Collection and Identification
From March 2019 to September 2021, 19 hv-CRKP-VAP cases were admitted to the EICU of the Affiliated Hospital of Xuzhou Medical University. 19 non-repeated isolates of hv-CRKP were collected from these patients. Matrix-Assisted Laser Desorption / Ionization Time of Flight Mass Spectrometry (Bruker Daltonics, Billerica, MA, USA) was used to recheck the isolates of K. pneumoniae. All isolates were resistant to at least one carbapenems (imipenem, meropenem, ertapenem) and were identified as CRKP by the VITEK-2 compact automated microbiology analyzer (Biomerieux, Marcy L’Etoile, France). String tests were carried out to see if hvKP had the hypermucoviscosity phenotype. Furthermore, patients’ clinical data including gender, age, isolation date, sampling site, comorbidity, invasive operation, antibiotic treatments, and outcomes were also collected. This investigation was authorized by the Ethics Committee of the Affiliated Hospital of Xuzhou Medical University XYFY2020-KL084.
Testing for Antimicrobial Susceptibility
The minimal inhibitory concentrations of antibiotics, including amikacin, aztreonam, ceftazidime, ciprofloxacin, cefotaxime, cefepime, imipenem, meropenem, polymyxin B, sulfamethoxazole, tigecycline, and piperacillin/tazobactam were determined by VITEK-2 Compact system. The guidelines provided by the Clinical and Laboratory Standards Institute (CLSI, 2021) were followed for the interpretation of the results.19 Pseudomonas aeruginosa ATCC27853 and Escherichia coli ATCC25922 were used as quality control strains.
Detection of Antimicrobial Resistance Genes
For extracting the genomic DNA of all the isolates, the boiling method was applied. The ESBL genes (blaTEM, blaSHV,blaCTX-M-1 group, blaCTX-M-2 group, blaCTX-M-8 group, and blaCTX-M-9 group), carbapenemase genes (blaKPC, blaNDM,blaVIM, blaIMP, and blaOXA-48) and AmpC-lactamase genes (blaCMY and blaDHA) were amplified by polymerase chain reaction (PCR) as described previously and the PCR products were sequenced and then on the sequencing results BLAST (https://blast.ncbi.nlm.nih.gov/Blast.cgi) was run.20,21 The primer sequences are enlisted in Table S1.
Detection of Capsular Serotyping and Virulence-Associated Genes
Wzi gene was amplified by PCR and sequenced as previously described.22 To compare the findings and identify the capsular type of K. pneumoniae, the database (https://bigsdb.pasteur.fr) was consulted. 12 virulence-associated genes including aerobactin, rmpA2, rmpA, magA, kfu, allS, fimH, ybtS, entB, mrkD, iroN, and iucA were also amplified by PCR.23,24 The primer sequences of Wzi and virulent genes are enlisted in Table S1.
Multilocus Sequence Typing (MLST)
gapA, infB, mdh, pgi, phoE, rpoB, and tonB are the seven housekeeping genes (expressed by K. pneumoniae) that were amplified by PCR and sequenced for all isolates by the MLST website’s methodology (https://bigsdb.pasteur.fr/klebsiella/primers-used). The sequencing results were compared with the database to identify sequence types.
Pulsed-Field Gel Electrophoresis (PFGE)
The homology of all strains was analyzed by PFGE assay.25 Genomic DNA was digested with Xbal for 2 h at 37°C and the fragments were separated using the CHEF Mapper XA System (Bio-Rad) for 18h at 14°C. The molecular identifier was determined to be a Salmonella serotype Braenderup strain H9812.26 Bionumerics software was utilized for the unweighted pair-group technique with arithmetic means (UPGMA) for cluster analysis. A greater than 85% similarity was considered as a closely related.20,27
String Test
Isolates of K. pneumoniae were plated on blood agar and grown overnight at 37°C. A single colony was contacted and pushed outward using an inoculation loop. The string test was considered positive and the isolate showed a hypervirulent phenotype if the length of the viscous string was greater than 5 mm.28 This experiment was repeated thrice.
Serum-Killing Assay
Three ST11-KL64 strains (KP3, KP9, KP13) and 3 ST15-KL112 strains (KP6, KP15, KP18) of bacteria were randomly selected for serum killing assay. The test was chosen in accordance with a previously reported study.28,29 Serum was collected from 10 healthy individuals and was kept at −80°C. In phosphate-buffered saline (PBS), the mid-log phase bacteria were suspended in a concentration of 1×106 CFU/mL and then combined with normal serum in a ratio of 1:3. The mixture was immediately plated on Mueller-Hinton agar after being incubated at 37°C for 0, 1, and 3 hours in order to count the number of bacteria. The results were graded on six levels, and an isolate was declared serum sensitive at grade 1 or 2, intermediately sensitive at 3 or 4, and resistant at 5 or 6. The experiment was repeated thrice. For hyper and low virulence control, ATCC43816 and ATCC700603 were employed, respectively.27,30,31
Galleria Mellonella Larvae Infection Assay
Galleria mellonella larvae infection assay was performed to further assess bacterial pathogenicity.20,32–34 The isolates and controls were the same as those in the serum-killing assay. PBS was used to adjust the overnight culture to the concentrations of 1×105 to 1×108 CFU/mL. Galleria mellonella larvae weighing 250—350 mg were used in this experiment. Ten microliter of various concentrations of each bacterium strain suspension was injected in 10 larvae through last proleg. Furthermore, 10 µL PBS was injected into 10 wax moths as an experiment control. Each larva was put onto a clean Petri plate and incubated at 37°C. For three days, the larval survival rates every 12 hours were recorded. This experiment was performed in three replicates.
Whole Genome Sequencing and Analysis
BGI Technology Service Co., Ltd sequenced the whole genome of strain KP3 in Shenzhen. The PacBio Sequel II system was used for third-generation sequencing. The whole genome was submitted to GenBank. BLAST (http://blast.ncbi.nlm.nih.gov) was utilized to compare plasmid sequences and the genes associated with drug resistance and virulence were predicted using ResFinder4.1 and VFDB 2019 databases. Map plasmid genome circles were assessed via the Gcview website.
Results
Patient’s Clinical Features and CR-hvKP Isolates
Nineteen isolates of hv-CRKP-VAP were collected from different patients during their stay at the EICU between March 2019 to September 2021. Before strain isolation, all the patients underwent invasive operations and received antibiotic treatment. The mortality rate was 73.68% (14/19). The isolates were acquired from different specimens including sputum (11/19, 57.89%), blood (4/19, 21.05%), bone marrow (1/19, 5.26%), cerebrospinal fluid (1/19, 5.26%), fibro bronchoscope lotion (1/19, 5.26%), and hydrothorax (1/19, 5.26%). The specifics of each clinical feature are listed in Table 1.
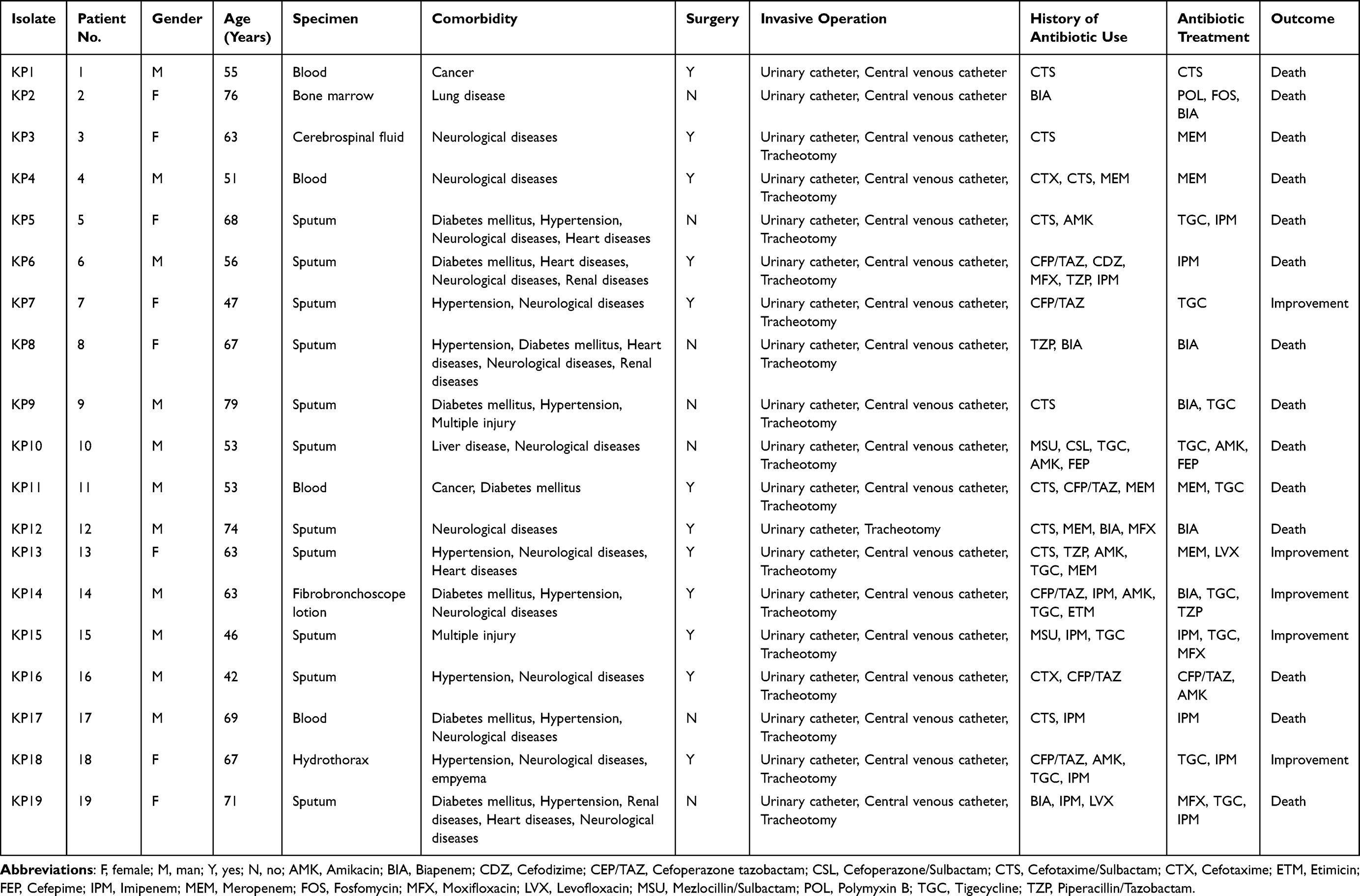 |
Table 1 Patient’s and Hv-CRKP Isolate’s Clinical Features |
Antibiotic Sensitivity and Genetic Characteristics of Resistance Genes
Except for tigecycline and colistin, all the organisms were highly resistant to the tested antibiotics (Table 2). Carbapenemase genes were detected from all 19 isolates, including blaKPC-2 (14/19, 73.68%), blaOXA-232 (4/19, 21.05%), and blaNDM-1 (2/19, 10.53%). KP19 contained both blaKPC-2 and blaNDM-1. Seven strains (36.84%) harbored the blaCTX-M-1 group and the profitable products were identified as blaCTX-M-107. The blaCTX-M-65 variant was present in 12 (63.16%) of the strains that owned the blaCTX-M-9 group. The ratios of blaSHV and blaTEM expression by CR-hvKP bacteria were 100% (19/19) and 73.68% (14/19), respectively. No isolate expressed blaVIM,blaIMP,blaCMY,blaDHA,blaCTX-M-2, and blaCTX-M-8 groups (Figure 1).
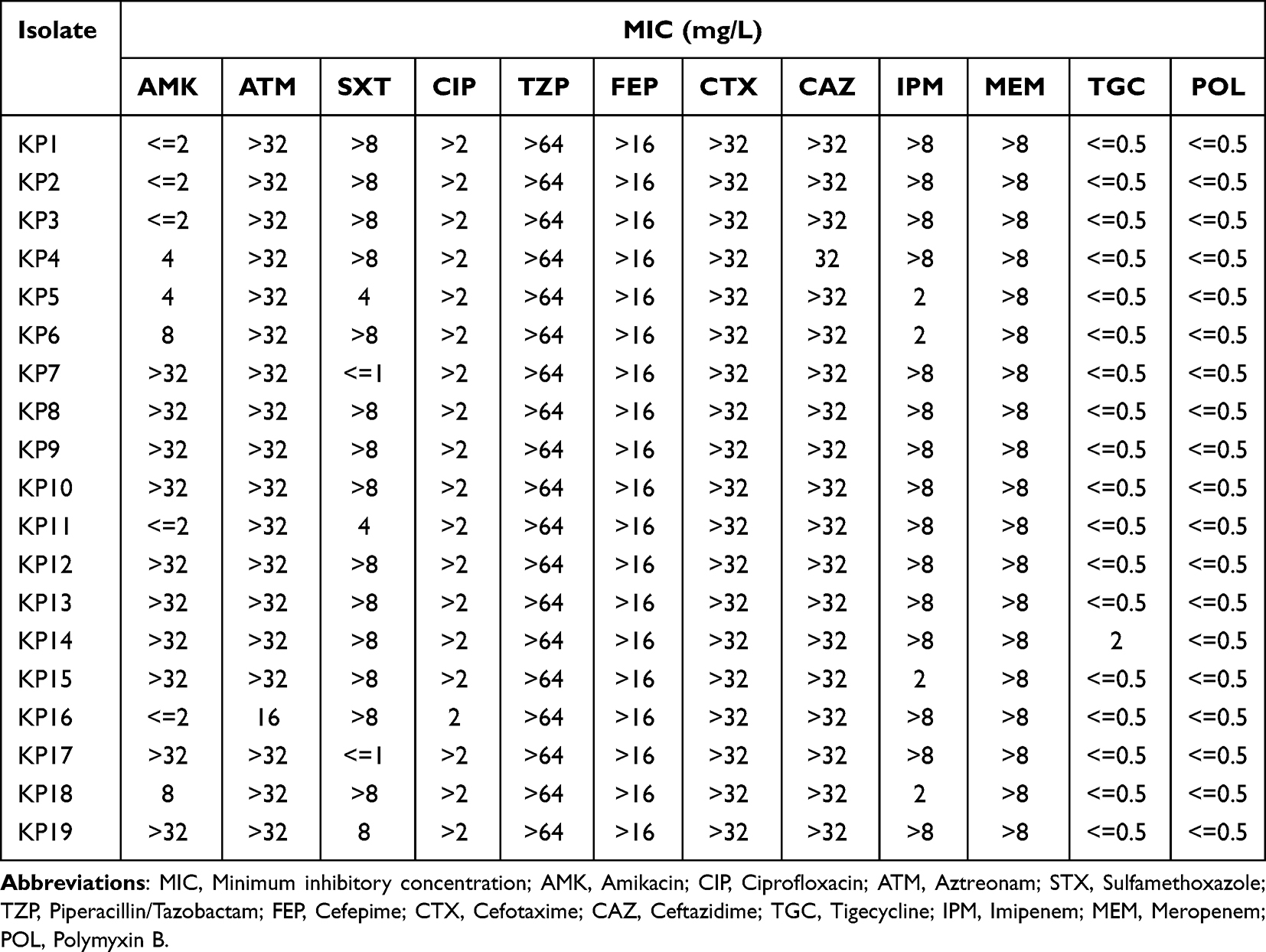 |
Table 2 Antibiotic Resistance Characteristics |
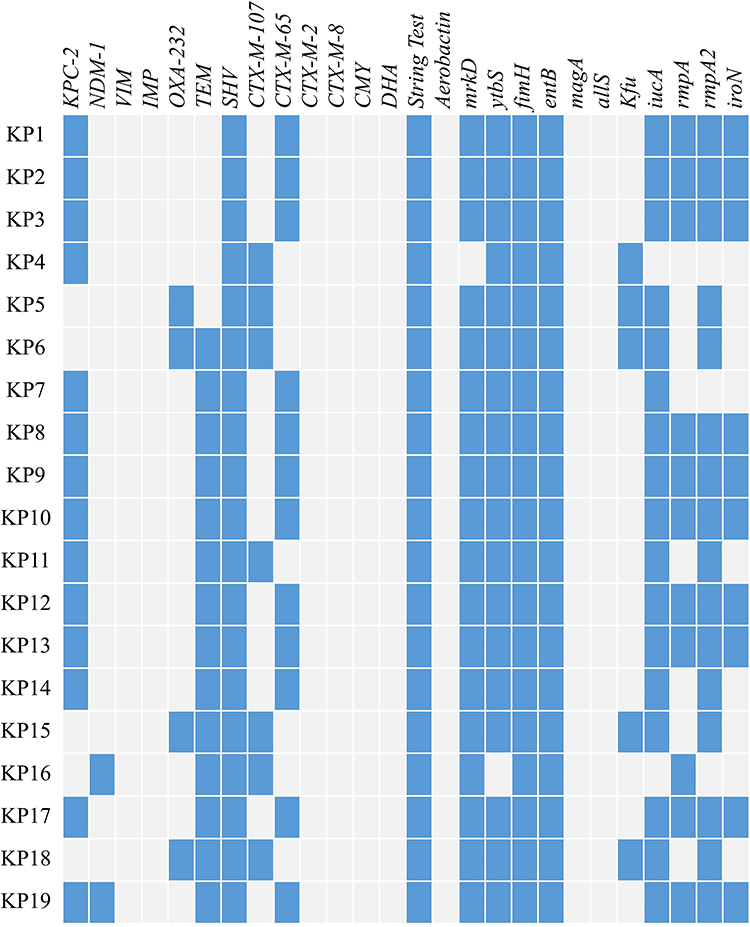 |
Figure 1 19 hv-CRKP isolates’ antimicrobial resistance genes, virulence-related genes, and string test results. The blue box represents the presence of genes or a positive string test result. |
Virulence-Associated Genes and Capsular Serotyping
Figure 1 displays the outcomes of 12 virulence-related genes. The fimH and entB genes were present in all 19 isolates. The following genes were mrkD (18/19, 94.74%), ytbS (18/19, 94.74%), iucA (17/19, 89.47%), rmpA2 (16/19, 84.21%), rmpA (11/19, 57.89%), iroN (10/19, 52.63%), and kfu (5/19, 26.32%). No isolate in this study expressed aerobactin, magA, and allS. Six distinct capsular serotypes for strains were found by PCR and sequencing of the Wzi gene, including KL64 (10/19, 52.63%), KL112 (4/19, 21.05%), KL47 (2/19, 10.53%), KL19 (1/19, 5.26%), KL62 (1/19, 5.26%), and KL140 (1/19, 5.26%) (Figure 2).
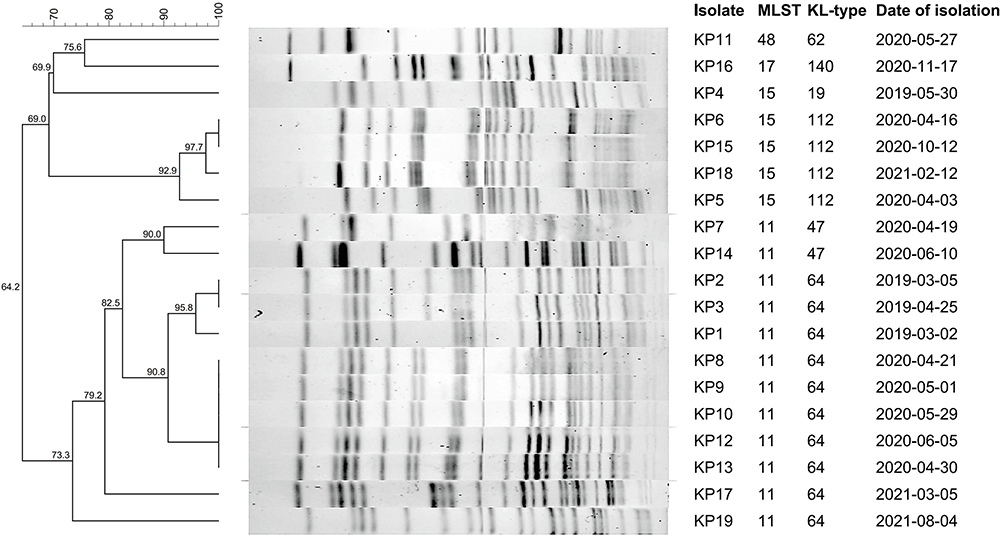 |
Figure 2 PFGE results, MLST type, capsular serotype, and date of isolate for 19 hv-CRKP isolates. Similarity >85% was considered closely related. |
MLST and PFGE
In total four STs were identified, where ST11 (12/19, 63.16%) accounted for the majority of the strains, followed by ST15 (5/19, 26.32%), ST17 (1/19, 5.26%), and ST48 (1/19, 5.26%). In the PFGE experiment, 4 ST15 and 8 ST11 isolates indicated high homology (>85%), respectively (Figure 2).
Virulence-Related Characteristics
The string test revealed that all strains were positive (Figure 1). KP9 and the hypervirulence standard strain ATCC43816 are resistant to healthy, normal serum (grade 6). KP15 and KP18 strains were moderately sensitive (grade 4) to healthy serum, however, KP3, KP6, and KP13 as well as the classical K. pneumoniae isolate ATCC700603 were all susceptible to healthy serum (grade 1 or 2) (Figure 3). To further evaluate the virulence of these strains in-vivo, G. mellonella larvae were infected with the experimental strains and their survival rate was calculated after 72h of incubation. When the concentration of bacteria suspension was 1×106 CFU/mL, the survival rate of classic K. pneumoniae isolates ATCC700603 was 73.33%, and that of hypervirulence control ATCC43816 was 6.67%, whereas, the survival rates of KP3, KP6, and KP13 were 16.67%, 6.67%, and 13.33%, respectively at 72h. KP9 and KP15 had a survival rate of 10% after 72h. Survival of G. mellonella reached 0% with KP18 at 24h (Figures 4 and S1). The result of the G. mellonella larvae infection assay suggested the strains in this study were hypervirulent.
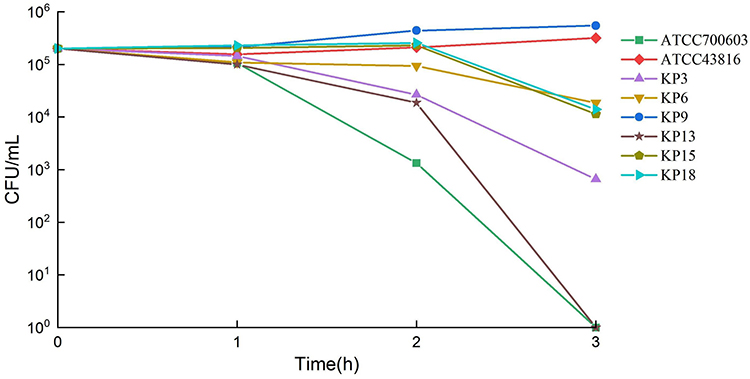 |
Figure 3 Serum killing assay of 6 selected isolates. ATCC43816 and ATCC700603 were hypervirulence and low virulence controls, respectively. |
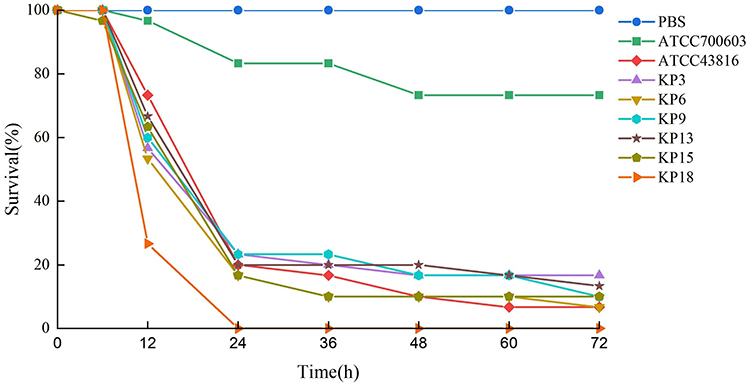 |
Figure 4 Survival rate of G. mellonella larvae infected with 1×106 CFU/mL of each selected isolate. ATCC43816 and ATCC700603 were hypervirulence and low virulence controls, respectively. PBS was the experimental control. |
Whole Genome Sequencing and Analysis
The genome sequence identified in this investigation was submitted to GenBank with the accession number CP116491, CP116492, CP116493, and CP116494. KP3 was discovered to have a 5521665bp chromosome and three plasmids (218592bp, 100630bp, and 85038bp). Plasmid 1 contained the virulence genes rmpA, rmpA2, and iucABCD, was quite similar to the virulence plasmid pLVPK (AY378100), and had a coverage of 91% and consistency of 99.05% (Figure 5A). Plasmid 3 carried blaKPC-2, had 99.98% homology with NTUH-K2044-CR plasmid unnamed2 (MZ475710.1) and 100% with pKPC-CR-hvKP-C789 (CP034417.1), phvKP12-KPC (CP103319.1) and pJX2-2 (CP064248.1), and query coverages of 80–100% (Figure 5B).
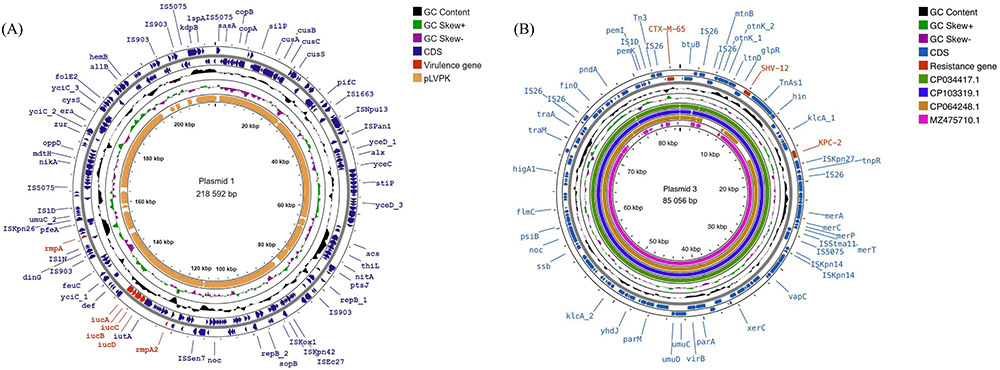 |
Figure 5 Two plasmid circle maps were generated with the GCview website. (A) Plasmid 1 carried rmpA, rmpA2, and iucABCD and was highly similar to pLVPK. The white regions indicate absence. (B) Plasmid 3 carried blaKPC-2 and shared a high identity with CP034417.1, CP103319.1, CP064248.1, and MZ475710.1 by comparison. |
Discussion
This study retrospectively investigated 19 hv-CRKP-VAP cases from March 2019 to September 2021 enrolled at the EICU of a Chinese teaching hospital and analyzed the outbreak characteristics of ST11-KL64 and ST15-KL112 hv-CRKP strain. Following the analysis, 13 risk variables associated with CRE infection were identified, including infection acquired from ICU, invasive procedures, mechanical ventilation, medical devices, CRE exposure, carbapenem use, β-lactam use, cephalosporin use, glycopeptide use, quinolone use, demographic patient characteristics, underlying disease or condition, and exposure to hospital care.35 To our knowledge, outbreaks of HAIs due to CRKP are very commonly reported, especially in VAP due to intubation. PFGE assays revealed that the homology of the eight kpc-2-producing ST11-KL64 CR-hvKP strains was >90%. These strains were isolated from elderly patients hospitalized in the EICU between March 2019 to June 2020, most of whom had neurological diseases. All patients underwent invasive procedures and had a morbidity and mortality rate of 73.7%, suggesting that the consequences of hv-CRKP-induced VAP are severe. Recently, infections and outbreaks caused by this pathogen have been reported in many regions.24,36,37 This pathogen requires more attention from clinicians and infection control physicians. Since the first report of K. pneumoniae carrying OXA-232 in Shanghai, in 2017, its epidemics have been reported in China.38–40 In this study, the four strains of OXA-232-producing ST15-KL112 hv-CRKP had >92% homology. Different from the KPC-2-producing CRE strains (MIC ≥ 16ug/mL), these strains showed intermediary sensitivity to imipenem with MIC values of 2ug/mL, possibly because of more available drug options. 50% of patients with OXA-232-producing strains improved after treatment compared with the KPC-2-producing CRE strain. A deadly VAP epidemic induced by ST11 hv-CRKP was documented in a 2017 article and occurred in a Chinese hospital.18 The stains were highly transmissible, multidrug-resistant, and hypervirulent at the same time. Genomic analyses revealed that the virulence plasmid acquired by the hv-CRKP strains was the classic ST11 CRKP stains. Simply, hv-CRKP strains have high virulence and multidrug resistance, and therefore possess a great challenge to clinical treatment and are a severe threat to public health. Fortunately, no homologous strains in VAP patients were detected after July 2020 following intervention with stringent infection control measures.
hv-CRKP consists of two evolutionary mechanisms, one in which CRKP strains acquire virulence genes or virulence plasmids, and the other in which hvKP strains acquire chromosomal or plasmid-encoded drug resistance genes.16 The strains in this investigation fall in the former type. There are three main mechanisms responsible for K. pneumoniae’s resistance to carbapenem: enzyme production, efflux pumps, and pore protein mutations.41 In China, KPC-2 is the most common carbapenemase for CRKP, and ST11 is the most common sequence type.10,24,36 This study comprises the majority of the KPC-2-producing ST11 K. pneumoniae strains. The hvKP strains are frequently used to describe the positive strains of the “string test”. However, it has also been reported that these lack hypervirulent phenotypes. Therefore, the string test cannot be the only indicator of the hvKP strain.42 A study suggested that rmpA, rmpA2, iucA, iroB, and peg-344 are the most accurate molecular markers to define hvKP because these genes are all present on virulence plasmids.43 Capsular polysaccharide, siderophore system, lipopolysaccharide, outer membrane protein, and efflux pump are currently recognized hvKP pathogenic components, however, the capsule has been identified as a major virulence component.44 The rmpA and rmpA2 genes increase muco-viscosity and pathogenicity and are linked to high capsule productivity.42 Here, all strains had a hyper-viscous phenotype and most of them contained rmpA or rmpA2 except KP4 and KP7, indicating that other mechanisms may also affect the hyper-mucoviscosity.45 Aerobactin is encoded by the iucABCD gene, and its corresponding receptor is expressed by the iutA. iucA encodes a IucA ortholog that is thought to bind ahLys onto the primary carboxylates of citrate to produce aerobactin siderophore.44,46 Aerobactin is considered the critical siderophore system of hvKP, expressed in >90% of its strains but only in 6% of classic K. pneumoniae.44 No isolate from this study expressed aerobactin, further indicating that these strains were produced by CRKP obtaining virulence genes or virulence plasmids. The blaKPC-2 and iucABCD were also identified on plasmids by whole genome sequencing of the KP3 strain.
In vitro serum killing assays and in vivo G. mellonella larvae infection assays were conducted to assess the virulence of these strains. Six strains were selected randomly for the virulence test and all displayed high virulence in G. mellonella larvae. However, the serum survival rate of KP3, KP6, and KP13 were low. Such inconsistency in the in vitro and in vivo virulence results are not unique. Shen P also reported a similar situation.47 The underlying mechanisms behind these differences have yet to be determined. According to a study, humans and G. mellonella have different levels of K. pneumoniae virulence.48 For the virulence investigation, ST11-KL64 and ST15-KL112 hv-CRKP were chosen. The results revealed that ST11-KL64 included both rmpA and rmpA2, while ST15-KL112 only contained rmpA2, but the findings from serum killing assays and G. mellonella larvae infection assays were identical. The G. mellonella larvae infection model, however, has been proposed as a preliminary discriminant between K. pneumoniae strains with comparable virulence. According to Tang, the LD80 of K1 and K57 was 1.0×106 CFU/mL while the LD80 of K2, K5, and K20 was 1.0×107 CFU/mL after 96 hours of inoculation, indicating that K1 and K57 were more virulent than K2, K5, and K20.49 Insua also distinguished between K1 and K2 pathogenicity using G. mellonella larvae.50 Although the G. mellonella infection model has many benefits, it has been reported that the murine model is better at differentiating between hypervirulent and standard K. pneumoniae.51 Nevertheless, one study showed that G. mellonella larvae are a crucial infection model in assessing the infection process, antioxidant response, and extent of cell damage of the K. pneumoniae pathogen. Furthermore, G. mellonella larvae may be appropriate models for antibiotic susceptibility investigations as well as host interactions with resistant K. pneumoniae.52
This study illustrates that hv-CRKP has caused an outbreak in hospitals, and clarified the clinical and molecular characteristics of VAP caused by ST11 and ST15 hv-CRKP. It offers suggestions for clinical prevention, control, diagnosis, and therapy. However, this study has some limitations. Retrospective analysis prevents us from gathering more detailed clinical information and taking environmental samples to understand the route of the outbreak more accurately. Unfortunately, only a few strains were used in this single-center investigation and only the entire genome of one strain was read, sequencing the genomes of all the strains would allow for a deeper understanding of the genetic traits. In addition, we did not use the murine model to assess virulence, and using only G. mellonella larvae may not reveal the real hypervirulent natures of the studied strains.
Conclusions
Our findings show the prevalence of hv-CRKP in Chinese hospitals, with ST11-blaKPC-2 being the most common. After acquiring a virulence plasmid, the strain can infect patients across many systems, which significantly raises the mortality rate. The transmission mechanism of virulence plasmids, as well as variances in drug resistance and virulence among different subtypes, are concerns that should be addressed in the future. As a result, it provides substantial data support for infection control.
Data Sharing Statement
The following datasets are included in this study: NCBI GenBank NO.: CP116491, CP116492, CP116493, and CP116494.
Ethical Approval and Consent to Participate
The medical ethics committee of The Affiliated Hospital of Xuzhou Medical University granted this study ethical approval (XYFY2020-KL084).
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This research was supported by the six talent peaks project of Jiangsu Province (WSN-091), the Scientific Research Project of Jiangsu Provincial Health Commission (Z2021009), and the Open project of Jiangsu Province Key Laboratory (XZSYSKF2020030).
Disclosure
There are no relevant conflicts of interest reported by any of the authors.
References
1. Zhang Y, Zeng J, Liu W, et al. Emergence of a hypervirulent carbapenem-resistant Klebsiella pneumoniae isolate from clinical infections in China. J Infect. 2015;71(5):553–560. doi:10.1016/j.jinf.2015.07.010
2. Schwaber MJ, Carmeli Y. Carbapenem-resistant Enterobacteriaceae: a potential threat. JAMA. 2008;300(24):2911–2913. doi:10.1001/jama.2008.896
3. Ripabelli G, Sammarco ML, Salzo A, Scutellà M, Felice V, Tamburro M. New Delhi metallo-β-lactamase (NDM-1)-producing Klebsiella pneumoniae of sequence type ST11: first identification in a hospital of central Italy. Lett Appl Microbiol. 2020;71(6):652–659. doi:10.1111/lam.13384
4. Ripabelli G, Sammarco ML, Scutellà M, Felice V, Tamburro M. Carbapenem-resistant KPC- and TEM-producing Escherichia coli ST131 isolated from a hospitalized patient with urinary tract infection: first isolation in Molise Region, Central Italy, July 2018. Microb Drug Resist. 2020;26(1):38–45. doi:10.1089/mdr.2019.0085
5. Hu FP, Guo Y, Zhu DM, et al. Resistance trends among clinical isolates in China reported from CHINET surveillance of bacterial resistance, 2005–2014. Clin Microbiol Infect. 2016;22(Suppl 1):S9–S14. doi:10.1016/j.cmi.2016.01.001
6. Munoz-Price LS, Poirel L, Bonomo RA, et al. Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect Dis. 2013;13(9):785–796. doi:10.1016/S1473-3099(13)70190-7
7. Ding Y, Wang Y, Hsia Y, Sharland M, Heath PT. Systematic review of carbapenem-resistant Enterobacteriaceae causing neonatal sepsis in China. Ann Clin Microbiol Antimicrob. 2019;18(1):36. doi:10.1186/s12941-019-0334-9
8. de Man TJB, Yaffee AQ, Zhu W, et al. Multispecies outbreak of Verona integron-encoded metallo-ß-lactamase-producing multidrug resistant bacteria driven by a Promiscuous Incompatibility Group A/C2 plasmid [published correction appears in Clin Infect Dis. 2021 Mar 15;72(6):1108]. Clin Infect Dis. 2021;72(3):414–420. doi:10.1093/cid/ciaa049
9. Zhou H, Zhang K, Chen W, et al. Epidemiological characteristics of carbapenem-resistant Enterobacteriaceae collected from 17 hospitals in Nanjing district of China. Antimicrob Resist Infect Control. 2020;9(1):15. doi:10.1186/s13756-019-0674-4
10. Shao C, Wang W, Liu S, Zhang Z, Jiang M, Zhang F. Molecular epidemiology and drug resistant mechanism of carbapenem-resistant Klebsiella pneumoniae in elderly patients with lower respiratory tract infection. Front Public Health. 2021;9:669173. doi:10.3389/fpubh.2021.669173
11. Chen S, Zou D. Prognosis of hospital-acquired pneumonia/ventilator-associated pneumonia with Stenotrophomonas maltophilia versus Klebsiella pneumoniae in intensive care unit: a retrospective cohort study. Clin Respir J. 2022;16(10):669–676. doi:10.1111/crj.13537
12. Russo TA, Marr CM. Hypervirulent Klebsiella pneumoniae. Clin Microbiol Rev. 2019;32(3):e00001–19. doi:10.1128/CMR.00001-19
13. Chew KL, Lin RTP, Teo JWP. Klebsiella pneumoniae in Singapore: hypervirulent infections and the carbapenemase threat. Front Cell Infect Microbiol. 2017;7:515. doi:10.3389/fcimb.2017.00515
14. Zou Q, Li Y. Hypervirulent Klebsiella pneumoniae. N Engl J Med. 2021;385(9):833. doi:10.1056/NEJMicm2101602
15. Decré D, Verdet C, Emirian A, et al. Emerging severe and fatal infections due to Klebsiella pneumoniae in two university hospitals in France. J Clin Microbiol. 2011;49(8):3012–3014. doi:10.1128/JCM.00676-11
16. Choby JE, Howard-Anderson J, Weiss DS. Hypervirulent Klebsiella pneumoniae - clinical and molecular perspectives. J Intern Med. 2020;287(3):283–300. doi:10.1111/joim.13007
17. Wu C, Zhou Y, Ai W, et al. Co-occurrence of OXA-232, RmtF-encoding plasmids, and pLVPK-like virulence plasmid contributed to the generation of ST15-KL112 hypervirulent multidrug-resistant Klebsiella pneumoniae. Front Microbiol. 2023;14:1133590. doi:10.3389/fmicb.2023.1133590
18. Gu D, Dong N, Zheng Z, et al. A fatal outbreak of ST11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: a molecular epidemiological study. Lancet Infect Dis. 2018;18(1):37–46. doi:10.1016/S1473-3099(17)30489-9
19. CLSI. Performance Standards for Antimicrobial Susceptibility Testing, M100.
20. Zhao Y, Zhang X, Torres VVL, et al. An outbreak of carbapenem-resistant and hypervirulent Klebsiella pneumoniae in an intensive care unit of a major teaching hospital in Wenzhou, China. Front Public Health. 2019;7:229. doi:10.3389/fpubh.2019.00229
21. Kang H, Zheng W, Kong Z, et al. Disease burden and molecular epidemiology of carbapenem-resistant Klebsiella pneumonia infection in a tertiary hospital in China. Ann Transl Med. 2020;8(9):605. doi:10.21037/atm.2020.03.122
22. Brisse S, Passet V, Haugaard AB, et al. Wzi gene sequencing, a rapid method for determination of capsular type for Klebsiella strains. J Clin Microbiol. 2013;51(12):4073–4078. doi:10.1128/JCM.01924-13
23. Zhou C, Wu Q, He L, et al. Clinical and molecular characteristics of carbapenem-resistant hypervirulent Klebsiella pneumoniae isolates in a Tertiary Hospital in Shanghai, China. Infect Drug Resist. 2021;14:2697–2706. doi:10.2147/IDR.S
24. Xu M, Fu Y, Fang Y, et al. High prevalence of KPC-2-producing hypervirulent Klebsiella pneumoniae causing meningitis in Eastern China. Infect Drug Resist. 2019;12:641–653. doi:10.2147/IDR.S191892
25. Tenover FC, Arbeit RD, Goering RV, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33(9):2233–2239. doi:10.1128/jcm.33.9.2233-2239.1995
26. Hunter SB, Vauterin P, Lambert-Fair MA, et al. Establishment of a universal size standard strain for use with the PulseNet standardized pulsed-field gel electrophoresis protocols: converting the national databases to the new size standard. J Clin Microbiol. 2005;43(3):1045–1050. doi:10.1128/JCM.43.3.1045-1050.2005
27. Su S, Zhang J, Zhao Y, et al. Outbreak of KPC-2 carbapenem-resistant Klebsiella pneumoniae ST76 and carbapenem-resistant K2 hypervirulent Klebsiella pneumoniae ST375 strains in Northeast China: molecular and virulent characteristics. BMC Infect Dis. 2020;20(1):472. doi:10.1186/s12879-020-05143-y
28. Zhang Y, Jin L, Ouyang P, et al. Evolution of hypervirulence in carbapenem-resistant Klebsiella pneumoniae in China: a multicentre, molecular epidemiological analysis. J Antimicrob Chemother. 2020;75(2):327–336. doi:10.1093/jac/dkz446
29. Siu LK, Fung CP, Chang FY, et al. Molecular typing and virulence analysis of serotype K1 Klebsiella pneumoniae strains isolated from liver abscess patients and stool samples from noninfectious subjects in Hong Kong, Singapore, and Taiwan. J Clin Microbiol. 2011;49(11):3761–3765. doi:10.1128/JCM.00977-11
30. Cai Z, Jia T, Pu M, et al. Clinical and molecular analysis of ST11-K47 carbapenem-resistant hypervirulent Klebsiella pneumoniae: a strain causing liver abscess. Pathogens. 2022;11(6):657. doi:10.3390/pathogens11060657
31. Yan R, Lu Y, Zhu Y, et al. A sequence type 23 hypervirulent Klebsiella pneumoniae strain presenting carbapenem resistance by acquiring an IncP1 blaKPC-2 plasmid. Front Cell Infect Microbiol. 2021;11:641830. doi:10.3389/fcimb.2021.641830
32. Singkum P, Suwanmanee S, Pumeesat P, Luplertlop N. A powerful in vivo alternative model in scientific research: galleria mellonella. Acta Microbiol Immunol Hung. 2019;66(1):31–55. doi:10.1556/030.66.2019.001
33. Feng Y, Lu Y, Yao Z, Zong Z. Carbapenem-resistant hypervirulent Klebsiella pneumoniae of sequence type 36. Antimicrob Agents Chemother. 2018;62(7):e02644–17. doi:10.1128/AAC.02644-17
34. Yuan Y, Li Y, Wang G, et al. blaNDM-5 carried by a hypervirulent Klebsiella pneumoniae with sequence type 29. Antimicrob Resist Infect Control. 2019;8(1):140. doi:10.1186/s13756-019-0596-1
35. van Loon K, Voor In ‘t Holt AF, Vos MC. A systematic review and meta-analyses of the clinical epidemiology of carbapenem-resistant Enterobacteriaceae. Antimicrob Agents Chemother. 2017;62(1):e01730–17. doi:10.1128/AAC.01730-17
36. Ouyang P, Jiang B, Peng N, et al. Characteristics of ST11 KPC-2-producing carbapenem-resistant hypervirulent Klebsiella pneumoniae causing nosocomial infection in a Chinese hospital. J Clin Lab Anal. 2022;36(6):e24476. doi:10.1002/jcla.24476
37. Zhang X, Ouyang J, He W, et al. Co-occurrence of rapid gene gain and loss in an interhospital outbreak of carbapenem-resistant hypervirulent ST11-K64 Klebsiella pneumoniae. Front Microbiol. 2020;11:579618. doi:10.3389/fmicb.2020.579618
38. Yin D, Dong D, Li K, et al. Clonal dissemination of OXA-232 carbapenemase-producing Klebsiella pneumoniae in Neonates. Antimicrob Agents Chemother. 2017;61(8):e00385–17. doi:10.1128/AAC.00385-17
39. Li X, Ma W, Qin Q, et al. Nosocomial spread of OXA-232-producing Klebsiella pneumoniae ST15 in a teaching hospital, Shanghai, China. BMC Microbiol. 2019;19(1):235. doi:10.1186/s12866-019-1609-1
40. Shu L, Dong N, Lu J, et al. Emergence of OXA-232 carbapenemase-producing Klebsiella pneumoniae that carries a pLVPK-like virulence plasmid among elderly patients in China. Antimicrob Agents Chemother. 2019;63(3):e02246–18. doi:10.1128/AAC.02246-18
41. Suay-García B, Pérez-Gracia MT. Present and future of carbapenem-resistant Enterobacteriaceae (CRE) Infections. Antibiotics. 2019;8(3):122. doi:10.3390/antibiotics8030122
42. Catalán-Nájera JC, Garza-Ramos U, Barrios-Camacho H. Hypervirulence and hypermucoviscosity: two different but complementary Klebsiella spp. phenotypes? Virulence. 2017;8(7):1111–1123. doi:10.1080/21505594.2017.1317412
43. Russo TA, Olson R, Fang CT, et al. Identification of biomarkers for differentiation of hypervirulent Klebsiella pneumoniae from Classical K. pneumoniae. J Clin Microbiol. 2018;56(9):e00776–18. doi:10.1128/JCM.00776-18
44. Paczosa MK, Mecsas J. Klebsiella pneumoniae: going on the offense with a strong defense. Microbiol Mol Biol Rev. 2016;80(3):629–661. doi:10.1128/MMBR.00078-15
45. Zhu J, Wang T, Chen L, Du H. Virulence factors in hypervirulent Klebsiella pneumoniae. Front Microbiol. 2021;12:642484. doi:10.3389/fmicb.2021.642484
46. Li C, Pan D, Li M, et al. Aerobactin-mediated iron acquisition enhances biofilm formation, oxidative stress resistance, and virulence of Yersinia pseudotuberculosis. Front Microbiol. 2021;12:699913. doi:10.3389/fmicb.2021.699913
47. Shen P, Berglund B, Chen Y, et al. Hypervirulence markers among non-ST11 strains of carbapenem- and multidrug-resistant Klebsiella pneumoniae isolated from patients with bloodstream infections. Front Microbiol. 2020;11:1199. doi:10.3389/fmicb.2020.01199
48. McLaughlin MM, Advincula MR, Malczynski M, Barajas G, Qi C, Scheetz MH. Quantifying the clinical virulence of Klebsiella pneumoniae producing carbapenemase Klebsiella pneumoniae with a Galleria mellonella model and a pilot study to translate to patient outcomes. BMC Infect Dis. 2014;14:31. doi:10.1186/1471-2334-14-31
49. Tang Y, Hu R. Galleria mellonella infection model for evaluating the virulence of Klebsiella pneumoniae. Chin J Microbiol Immunol. 2018;38(8):611–615. Chinese. doi:10.3760/cma.j.issn.0254-5101.2018.08.009
50. Insua JL, Llobet E, Moranta D, et al. Modeling Klebsiella pneumoniae pathogenesis by infection of the wax moth Galleria mellonella. Infect Immun. 2013;81(10):3552–3565. doi:10.1128/IAI.00391-13
51. Russo TA, MacDonald U, Papasian CJ. The galleria mellonella infection model does not accurately differentiate between hypervirulent and classical Klebsiella pneumoniae. mSphere. 2020;5(1):e00850–19. doi:10.1128/mSphere.00850-19
52. Sugeçti S. Pathophysiological effects of Klebsiella pneumoniae infection on Galleria mellonella as an invertebrate model organism. Arch Microbiol. 2021;203(6):3509–3517. doi:10.1007/s00203-021-02346-y
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Analysis of Risk Factors and Mortality of Patients with Carbapenem-Resistant Klebsiella pneumoniae Infection
Wu C, Zheng L, Yao J
Infection and Drug Resistance 2022, 15:2383-2391
Published Date: 3 May 2022
Characterization of Hypervirulent and Carbapenem-Resistant K. pneumoniae Isolated from Neurological Patients
Zhou Q, Wu C, Zhou P, Zhang J, Xiong Z, Zhou Y, Yu F
Infection and Drug Resistance 2023, 16:403-411
Published Date: 24 January 2023
Hypervirulent Klebsiella pneumoniae
Chen J, Zhang H, Liao X
Infection and Drug Resistance 2023, 16:5243-5249
Published Date: 11 August 2023
Molecular Characterization and Risk Factors of Carbapenem-Resistant Hypervirulent Klebsiella pneumoniae Isolated from Chinese Tertiary Hospital
Zhu R, Li J, Lian S, Li Y, Cai M, Cao Y, Xu X
Infection and Drug Resistance 2025, 18:83-92
Published Date: 6 January 2025
A Comprehensive Overview of Klebsiella Pneumoniae: Resistance Dynamics, Clinical Manifestations, and Therapeutic Options
Jin SS, Wang WQ, Jiang YH, Yu YT, Wang RL
Infection and Drug Resistance 2025, 18:1611-1628
Published Date: 25 March 2025