")
Back to Journals » Infection and Drug Resistance » Volume 16
Characterization of Hypervirulent and Carbapenem-Resistant K. pneumoniae Isolated from Neurological Patients
Authors Zhou Q, Wu C, Zhou P, Zhang J, Xiong Z, Zhou Y, Yu F
Received 21 October 2022
Accepted for publication 16 January 2023
Published 24 January 2023 Volume 2023:16 Pages 403—411
DOI https://doi.org/10.2147/IDR.S392947
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Suresh Antony
Qingping Zhou,1,* Chunyang Wu,2,* Peiyao Zhou,3 Ji Zhang,1 Zhanghua Xiong,4 Ying Zhou,5 Fangyou Yu5
1Department of Neurology, Jiangxi Provincial People’s Hospital, The First Affiliated Hospital of Nanchang Medical College, Nanchang, 330006, People’s Republic of China; 2Department of Respiratory Medicine, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou, 325000, People’s Republic of China; 3Department of Laboratory Medicine, The First Affiliated Hospital of Wenzhou Medical University, Wenzhou, 325000, People’s Republic of China; 4Department of Clinical Laboratory Medicine, Jiangxi Provincial People’s Hospital, The First Affiliated Hospital of Nanchang Medical College, Nanchang, 330006, People’s Republic of China; 5Department of Clinical Laboratory Medicine, Shanghai Pulmonary Hospital, Tongji University School of Medicine, Shanghai, 200082, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Ying Zhou, Email [email protected]
Background: Patients with neurological disorders were easier to develop severe intracranial infections caused by hypervirulent and carbapenem-resistant K. pneumoniae, leading to a distressing clinical outcome. In this study, eight hv-CRKP were isolated from neurological patients, to clarify the resistant and virulent features.
Methods: We tested the susceptibility of common antibiotics in these isolates to feature the antibiotic-resistant phenotypes. We also detected the key virulence factors, including mucoviscosity, siderophores production, biofilm formation in vitro, and further evaluated the virulence potential with serum killing model. We also used whole-genome sequencing (WGS) to investigate the molecular mechanisms.
Results: We observed that ST11-KL64 hv-CRKP (6/8) has an overwhelming epidemic dominance in these hypervirulent and carbapenem-resistant K. pneumoniae. Though the acquirement of virulence plasmid made no influence to the maintain of multidrug-resistant phenotype of these isolates, only the ST11-KL64 strains fully exhibited the hypervirulent features. Compared with ST11-KL47 and ST15-KL24 strains, ST11-KL64 hv-CRKP were more advantages in productions of capsule polysaccharide, biofilm, and siderophores. The virulence potential of ST11-KL64 hv-CRKP was further confirmed by using serum killing model. Previous studies have demonstrated that IncFII plasmid could act as a helper plasmid to mobile the non-conjugative IncFIB/IncHIB virulence plasmids. We could only observe the co-existence of IncFII resistance plasmid and IncFIB/IncHIB virulence plasmids in ST11-KL64 isolates. The co-existence of such two plasmids facilitated the formation of ST11-KL64 hv-CPKP, which then become nosocomial epidemic under the antibiotic stress.
Conclusion: Overall, we observed the ST11-KL64 hv-CRKP dominated in the isolates from neurological patients, and required most clinical attention.
Keywords: Klebsiella pneumoniae, KPC-2, virulence plasmid, carbapenem resistance, hypervirulent
Introduction
Hypervirulent K. pneumoniae (hvKP) has arisen as a superbug during the 1990s, causing pyogenic liver abscesses, brain abscesses, and other invasive syndromes.1–3 Patients with hvKP infections generally have poor outcomes due to the infection’s fast progression.3 Recently, an increasing number of K. pneumoniae strains have been found that combine both hypervirulent and resistance characteristics, resulting in hypervirulent and carbapenem-resistant K. pneumoniae (hv-CRKP) with disastrous clinical outcomes.1,4–6
Since 2010, hv-CRKP has spread globally.1 The clinical situation raises a number of alarming issues. The fast dissemination and epidemics of hv-CRKP in medical settings constitute a significant challenge to infection control.7,8 A high level of resistance and pathogenicity frequently leads to poor clinical outcomes. Additionally, hv-CRKP seem to be able to colonize the gut.1 These strains are highly contagious and have the potential to cause severe infection, such as metastasis to distant sites, most commonly the eye, lung, and central nervous system (CNS).3
A large study of over 800 patients with K. pneumoniae liver abscesses in Taiwan, South Korea, and the United States found that 12% of patients had metastatic disease,9 though rates as high as 28% have been reported.10 The CNS is the most common metastatic site for hv-CRKP, occurring in roughly one-third of patients with metastatic disease.3,9,11 Patients with neurological disorders, where the brain barrier is disrupted, are more likely to develop intracranial infections caused by K. pneumoniae. Numerous cases of CNS disease, such as meningitis, brain abscess, and epidural abscess, have been reported.12–14 Previous research has shown that KL1-hvKP can cause central nervous system complications from pyogenic liver abscess regardless of the host’s disease conditions, with 70% of patients experiencing long-term disability due to neurological deficits.11 Although the clinical risk of hv-CRKP to the neurological patients has been demonstrated, the resistance and hypervirulent features of these isolates identified in neurological patients remain unclear.
In this study, we isolated hypervirulence and carbapenem-resistant K. pneumoniae from patients diagnosed with neurological diseases, aiming to clarify the resistant and virulent features, and clinical hazards of these organisms. We tested the susceptibility of common antibiotics in these isolates to feature the antibiotic-resistant phenotypes. To characterize the virulence in detail, we detected the key virulence factors, including mucoviscosity, siderophores production, biofilm formation in vitro, and further evaluated the virulence potential with serum killing model. We also used whole-genome sequencing (WGS) to investigate the molecular mechanisms underlying multidrug resistance and hypervirulent phenotypes. Moreover, we also analyzed the plasmids diversity to determine the potential transmission of these high-risk determinants. Overall, our goal was to describe the specific hypervirulence and carbapenem-resistant K. pneumoniae from neurological patients, to figure out the lineage requiring most clinical attention, and emphasize the possible risk of such strains.
Materials and Methods
Bacterial Strains
We have obtained 13 isolates from patients diagnosed with neurological diseases, including 8 hv-CRKP and 5 CRKP. Eight K. pneumoniae strain exhibited typical hypervirulent and carbapenem-resistant features, harboring carbapenem-resistant genes and hypervirulent determinants. K. pneumoniae NUTH-K2044 (ST23-KL1) was used as a virulence-positive control strain.
Antimicrobial Susceptibility Test
The MIC (minimum inhibitory concentration) of the K. pneumoniae strains was determined using broth microdilution by the Clinical and Laboratory Standards Institute.15 For MIC determination, E. coli ATCC 25922 was used as a quality control strain. CLSI2022-M100-ED31was used to determine the interpretative breakpoints.
Whole-Genome Sequencing and Bioinformatics Analysis
The genomic DNA of these hv-CRKP was extracted using a commercial DNA extraction kit (Qiagen, Germany) and the genome sequencing was then performed by the Illumina NovaSeq 6000 platform.16 The multilocus sequence typing (MLST), serotype, resistance and virulence determinants were determined by the Kleborate (version 0.3.0) (https://github.com/katholt/Kleborate/). Plasmid replicons were identified using the PlasmidFinder database using the minimum coverage and minimum identities of 90% (https://cge.cbs.dtu.dk/services/PlasmidFinder/).
String Test
A positive string test for hypermucoviscosity is defined as a viscous filament of greater than 5 mm length when stretching a K. pneumoniae colony with a bacteriological loop on an agar plate.1
Quantitative Siderophore Production and Biofilm Formation Assay
To determine the iron-chelating capabilities of bacterial supernatants, the chrome azurol S (CAS) assay was performed as previously described.17 In brief, stationary-phase iron-chelated cultures (10 mL) were dropped on CAS plates, and siderophore production was determined by the orange halos after 48 hours at 37°C incubation.
Static biofilm production was quantified using crystal violet in 96-well flat-bottom polystyrene microtiter plates, and the extent of biofilm formation was determined as previously described. The image was taken for visual observation and for quantitation, OD 595 was detected through a microplate reader.
Serum Killing Model
The assay was carried out exactly as described in the published experiment.18 In brief, 25ul of a bacterial suspension containing 1.5×106 CFU/mL was mixed with 75ul of healthy human serum and incubated at 35°C. The baseline and 3-hour mixtures were diluted and inoculated overnight on nutrient agar plates. The number of colonies was counted at each timepoint. Sero-sensitivity survival curves were created based on the survival rate at each timepoint.
Results
ST11-KL64 Strains Dominated Among hv-CRKP Isolated in Neurological Patients
Eight hv-CRKP were collected from the patients diagnosed with neurological disorders in three hospitals, and most of them (6/8) also suffered with bloodstream infections. According to the MLST and serotype analysis, we found that ST11-KL64 hv-CRKP (6/8) has an overwhelming epidemic dominance in these hypervirulent and carbapenem-resistant K. pneumoniae (Table 1). Further core single-nucleotide polymorphism (SNP) distance analysis showed that the 6 ST11-KL64 isolates differed by an average of 51 core SNPs (range 43–68), indicating no clonal expansion (Figure S1). Another two isolates belonged to ST11-KL47, and ST15-KL24 (Table 2), which were usually recognized classical multidrug-resistant (MDR) K. pneumoniae.19 Hence, we speculated the formation of these eight hv-CRKP followed such pattern: CRKP (most were ST11-KL64 isolates) obtained a virulence plasmid.1
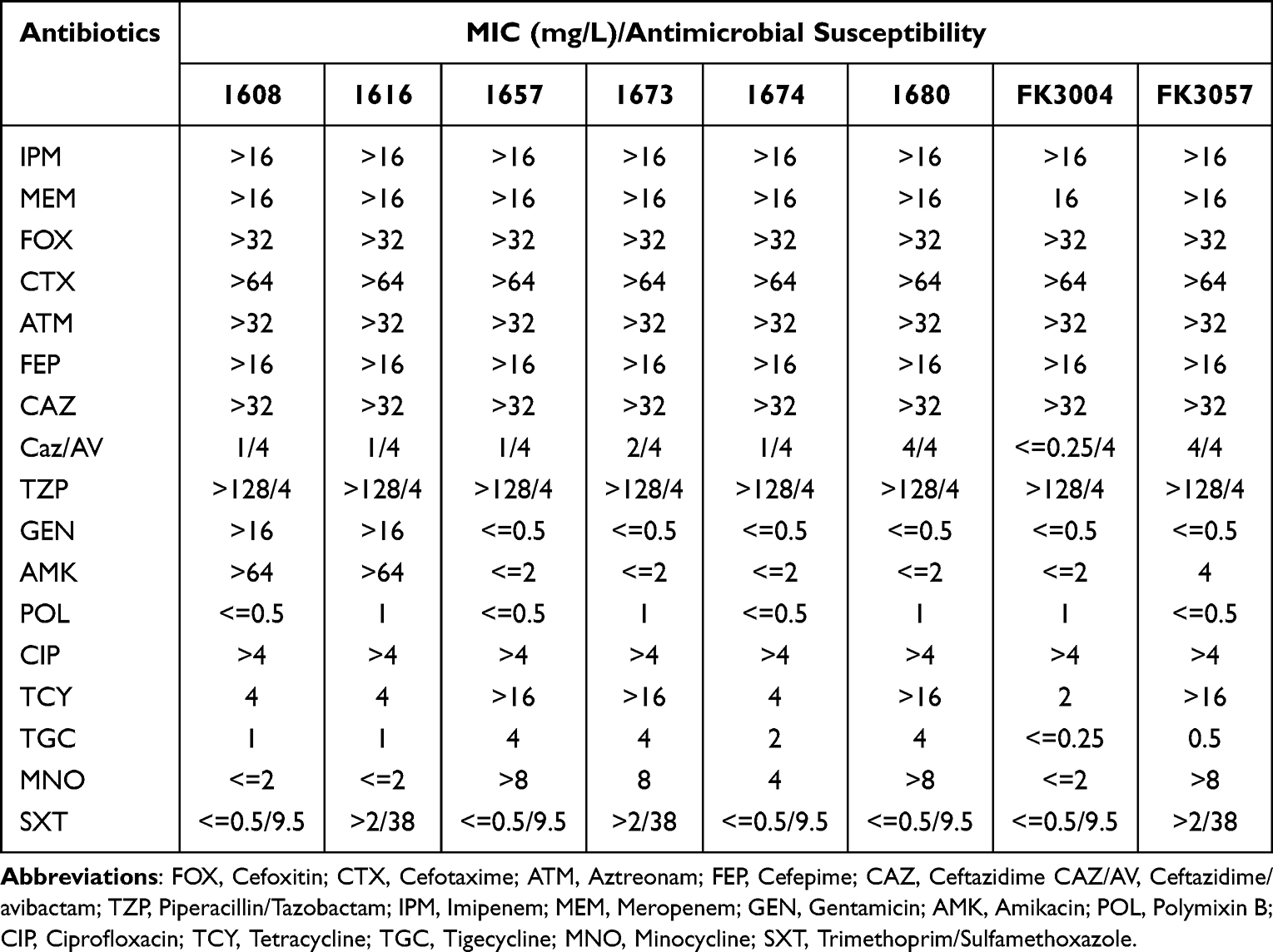 |
Table 1 Antimicrobial Drug Susceptibility Profiles |
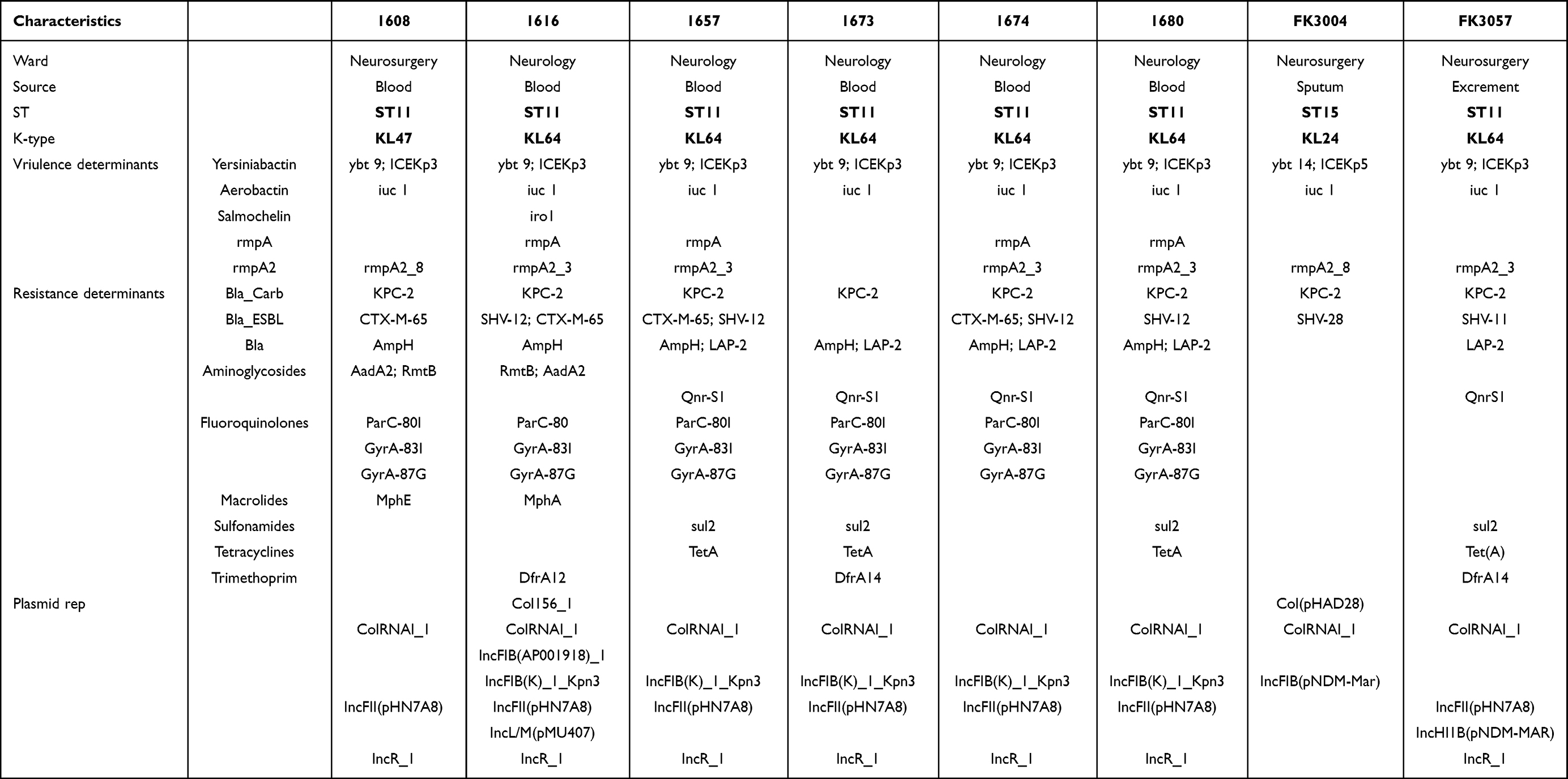 |
Table 2 General Features and Antimicrobial Resistance Genes and Virulence Determinants of in hv-CRKP |
Hv-CRKP Isolated in Neurological Patients All Exhibited Typical MDR Phenotype
In order to clarify the antibiotic-resistant phenotype of these hv-CRKP, we tested the susceptibility of 17 antibiotics in these strains (Table 2). We found all these K. pneumoniae posed similar multi-drug-resistant features; they exhibited high-level resistance to all β-lactam antibiotics and carbapenems, but remained susceptible to tigecycline, polymyxin, and ceftazidime/avibactam. Notably, K. pneumoniae 1608 and 1616 also resistant to aminoglycoside antibiotics, which were usually considered as effective therapeutic options for CRE (Carbapenem-Resistant Enterobacteriaceae). These antibiotic-resistant phenotype of K. pneumoniae 1608 (ST11-KL47) and 1616 (ST11-KL64) were not common in hv-CRKP. All these results indicated that the acquirement of virulence plasmid make no influence to the maintain of MDR phenotype.
Characterization of Virulence Phenotype of hv-CRKP Isolated in Neurological Patients
To protect themselves from the activities of bactericidal compounds, Klebsiella pneumoniae strains produce a capsule polysaccharide coating. In this study, we discovered that half of the hv-CRKP, all of which were ST11-KL64, had a particularly thick capsule due to increased capsule polysaccharide production, resulting in a hypermucoviscous phenotype (Figure 1A). Moreover, we also found these hypermucoviscous hv-CRKP also produced almost equivalent biofilm with ST23-KL1 hvKP NTUH-K2044 (Figure 1B). Quantitative siderophore assays indicated that except the ST15 isolate FK3004, all hv-CRKP produced comparable siderophores with NTUH-K2044 (Figure 1C). All these results showed that the expressions of hypervirulence-specific factors differed among ST11-KL64, ST11-KL47, and ST15-KL24 isolates. The ST11-KL64 hv-CRKP were more advantageous in the production of capsule polysaccharide, biofilm, and siderophores. We further evaluated the virulence potential of these isolates by using serum resistance in vitro. Only two ST11-KL64 hv-CRKP (1616 and FK3057) could perfect resistance the serum killing (Figure 1D), with almost 100% isolates survived after incubated with healthy human serum at 35°C for 3h. Notably, for the ST11-KL47 isolate 1608 and ST15-KL24 isolate FK3004, the survival rates were extremely low with 8% and 0.25%, respectively. Although the K. pneumoniae 1680 belonged to ST11-KL64 type and produced high capsule polysaccharide and siderophore, these hypervirulent factors do no help for the 1680 to anti the serum killing; only after 2h incubation, the surviving number of 1680 strain became 0.
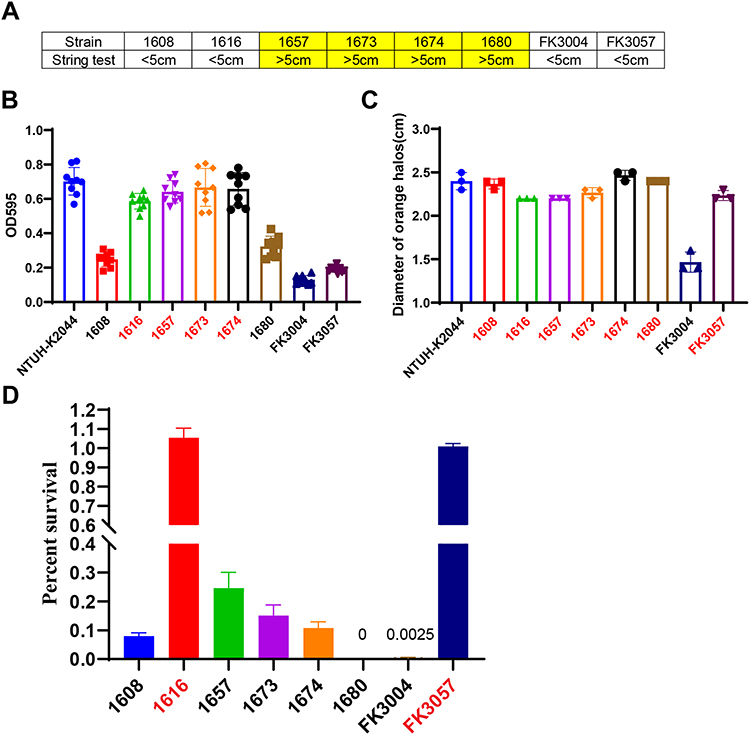 |
Figure 1 The virulence phenotype of eight hv-CRKP. (A) String test (B) biofilm formation. (C) Siderophores production determined by CAS agar plate. (D) Serum killing model, the survival rates were counted at 3 hours after incubated with the serum. |
The Variance of Resistance and Virulence Determinants Among hv-CRKP Isolated in Neurological Patients
To further investigate the related mechanism that mediated the multidrug resistant and hypervirulent characteristic of these K. pneumoniae, we used WGS to deeply describe the genomic information of these bacteria. All isolates harboring blaKPC-2 gene, which make the major contribution to the MDR phenotype (Table 1). Moreover, the aadA2 and rmtB identified in both 1608 and 1616, which conferred higher-level resistance to aminoglycoside antibiotics (Table 1). Among all these ST11 isolates including KL47 and KL64 types, we could find more than 10 elements mediating resistance to β-Lactams, aminoglycosides, fluoroquinolones, macrolides, etc. However, in the ST15 FK3004 isolates, we only observed two resistant genes blaKPC-2 and blaSHV-28 (Table 1), which only contributed to the resistance to β-Lactams and carbapenems, and supported with the antimicrobial susceptibility results (Table 2). Although ST15 K. pneumoniae were recognized the second major MDR-KP in China,19 it attained fewer resistance genes than ST11 strains.
As we all know, iron acquisition was a critical factor contributing to the high pathogenicity of hvKP.1,20 Among these three isolates, only 1616 harbored three siderophores (yersiniabactin, salmochelin, and aerobactin), another seven isolates only carried yersiniabactin and aerobactin (Table 1). However, the presence of salmochelin did not increase the production of siderophores, compared with other ST11 isolates (Figure 1C). Notably, the yersiniabactin gene in ST15 isolate FK3004 was typed as ybt-15 located on ICEKp5-like mobile genetic element, while which identified in ST11 strains were ybt-9 on ICEKp3. Such difference may explain why the high siderophores production absence in ST15 strain FK3004 (Figure 1C). In addition to the aerobactin, thick capsule due to high polysaccharide production in the capsule, which mediated by RmpA/RmpA2 also contributed to the hvKp phenotype. According to the WGS analysis, K. pneumoniae 1657, 1674, and 1680 all co-harbored rmpA and rmpA2 genes, which result in the hypermucoviscous phenotype (Table 1 and Figure 1A). However, the rmpA and rmpA2 were not the essential elements for the capsule polysaccharide production, since the 1673 was hypermucoviscous but loosed rmpA and rmpA2 (Table 1 and Figure 1A). Notably, the rmpA2 gene identified in ST11-KL47 isolate (1608) and ST15-KL24 isolate (FK3004) were truncated. All these results indicated difference of hypervirulence phenotype among these isolates were attributed to the difference of key virulence determinants.
The Plasmid Diversity in hv-CRKP Isolated in Neurological Patients
As plasmids are frequently transmitted between bacteria and have spread globally, which contributed significantly to the dissemination threats of resistant and virulent factors, we made analysis of the diversity of plasmids identified in these isolates. We found more than three plasmid replicons in these eight isolates (Table 1), indicating that CRKP strains, like ST11 or ST15 strains, could keep several plasmids with low fitness cost; this pattern can easily spread and cause an outbreak in the hospital.
Among these plasmids, IncFII plasmids served as the major vehicles of the blaKPC gene,21,22 and the IncFIB/IncHIB plasmids usually dominated in virulence plasmids,23 these plasmids could be detected in all six ST11-KL64 isolates, while the ST11-KL47 and ST15-KL24 isolates did not obtain (Table 1). Previous studies have demonstrated that the IncFII conjugative plasmid not only be self-transferred to other recipients, but also could act as a helper plasmid to mobile the non-conjugative IncFIB/IncHIB virulence plasmids.5,24 The oriT (origin of transfer site) region was the key element to recruit the conjugative process, which shared high identity between IncFII plasmids and IncFIB/IncHIB virulence plasmids.5,24 The co-existence of IncFII blaKPC-2 plasmid and virulence plasmid contributed to the transmission of both resistant and hypervirulent phenotype in ST11-KL64 isolates.
Discussion
Infections caused by the hypervirulent and carbapenem-resistant K. pneumoniae usually resulting in distressing clinical outcomes, especially for the patients suffering with neurological disorders, who were easier to develop serious intracranial infections. In this study, we isolated 8 hv-CRKP from the neurological patients, characterized the resistance and virulence phenotypes, and molecular features, and observed the ST11-KL64 hv-CRKP dominated, and required most clinical attention.
All these hv-CRKP could be categorized into three group: ST11-KL64, ST11-KL47, and ST15-KL24, which all were recognized as typical MDR K. pneumoniae. Most hypervirulent ST15 isolates were typed as KL19,25 while what we obtained was typed as KL24. Notably, not like ST11 isolates showed resistance to most common antibiotics, the ST15 isolate FK3004 only resistant to β-Lactams, carbapenems, and fluoroquinolones, since it just harbored two resistant genes blaKPC-2 and blaSHV-28. When these strains obtained virulence plasmids, only the ST11-KL64 strain fully exhibited the hypervirulent features. Although yersiniabactin, and aerobactin could be detected in the FK3004(ST15-KL24), it did not pose high siderophores production. ICEKp is an integrative conjugative element (ICE) that mobilizes the ybt locus, which encodes biosynthesis of the siderophore yersiniabactin and its receptor.20,26 Yersiniabactin is an important bacterial virulence factor because it provides mechanisms for removing iron from host transport proteins.20 The ICEKp population structure is made up of numerous sub-lineages, each with its own set of cargo genes and the yersiniabactin synthesis locus ybt.20 In this study, the yersiniabactin gene in ST15 isolate FK3004 was typed as ybt-15 located on ICEKp5 elements, different from that identified in ST11 strains, which could be explained by the presence of balancing selection for the encoded traits. Since the ybt and associated cargo genes were essential to the virulence phenotype, such variance among ST15 and ST11 isolates may result in different virulence phenotype. In addition, with the truncated rmpA2, the ST15 hv-CRKP produced less capsule polysaccharide, and only survive 025% strains under the serum killing, resulting in lower pathogenic.
Although the first carbapenem-resistant hypervirulent K. pneumoniae ST11 outbreak clone reported in eastern China was typed as ST11-KL47,7 ST11-KL64 strains appeared to be more resistant to harsh conditions and virulent, with evidence that pLVPK-like virulence plasmids were more frequently discovered among ST11-KL64 lineages.27 In this study, the ST11-KL47 hv-CRKP (1608) produced significantly less biofilm than ST11-KL64 isolates and NTUH-K2044 hypervirulent strain. Moreover, under the serum killing, only 8% ST11-KL47 isolates survived, while two ST11-KL64 isolates could survive 100%. Previous studies have observed that patients infected with ST11-KL64 isolates died at a significantly higher rate than those infected with ST11-KL47 isolates.27 The results of our phenotypic assays, which revealed that ST11-KL64 was more virulent than ST11-KL47, support this finding. The capsule type is thought to be an important determinant of K. pneumoniae pathogenicity, but we cannot rule out the possibility that the increased virulence is due to other chromosomal and plasmid variations.
The IncFIB/IncHIB virulence plasmid typed as a mobilizable plasmid, which just contain a MOB (mobility) module (oriT positive) and need the MPF (mating pair formation) of a coresident conjugative plasmid to become transmissible by conjugation.5,24 According to previous demonstration, the conjugative IncFII blaKPC plasmid seemed to act as a driving force in virulence plasmid mobilization. Notably, we found IncFII resistance plasmids and the IncFIB/IncHIB virulence plasmids could be co-detected in all six ST11-KL64 isolates, while these plasmid absence in ST11-KL47 and ST15-KL24 isolates. The co-existence of such virulence plasmid and blaKPC plasmid facilitate the formation and dissemination of ST11-KL64 hv-CPKP. The antibiotic stress further aggravated the epidemic of such hv-CRKP strains in hospitals. Moreover, as ST11-KL64 hv-CPKP could maintain multiple plasmids with low fitness cost, which do a great assistance for such strain to gain more hypervirulent and resistant phenotype.
Altogether, our study characterized 8 hv-CRKP isolated from neurological patients, and observed that the ST11-KL64 hv-CPKP emerged as dominated high-risk lineages, with enhanced MDR and hypervirulent phenotype. In comparison to ST11-KL47 and ST15-KL24 hv-CRKP, these ST11-KL64 strains are not only more resistant to stress, but also act as “super-spreaders” of virulence plasmids, allowing them to spread to more CRKP strains. Customized surveillance and stricter infection-control measures were urgently required to prevent the spread of such high-risk hv-CRKP.
Ethics Statement
The research protocol was approved by the Ethics Committee of Shanghai Pulmonary Hospital. We confirm that all adult participants gave their informed consent. Guidelines outlined in the Declaration of Helsinki were followed.
Acknowledgments
We thank the authority of NTUH-K2044 by Jin-Town Wang from National Taiwan University Hospital.
Funding
This study was supported by the [Jiangxi Provincial Health and Family Planning Commission Science and Technology Program] under Grant [number 20195061]. This study was supported by grants from Shanghai Pulmonary Hospital Development of Discipline-Department of Clinical Laboratory Medicine. This study was also supported by the [National Natural Science Foundation of China] under Grant [number 82202564], and [Shanghai Sailing Program] under Grant [number 22YF1437500].
Disclosure
The authors report that there are no competing interests to declare in this work.
References
1. Lan P, Jiang Y, Zhou J, Yu Y. A global perspective on the convergence of hypervirulence and carbapenem resistance in Klebsiella pneumoniae. J Glob Antimicrob Resist. 2021;25:26–34. doi:10.1016/j.jgar.2021.02.020
2. Nordmann P, Cuzon G, Naas T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect Dis. 2009;9(4):228–236. doi:10.1016/S1473-3099(09)70054-4
3. Choby JE, Howard-Anderson J, Weiss DS. Hypervirulent Klebsiella pneumoniae - clinical and molecular perspectives. J Intern Med. 2020;287(3):283–300. doi:10.1111/joim.13007
4. Yang X, Dong N, Chan EW, Zhang R, Chen S. Carbapenem resistance-encoding and virulence-encoding conjugative plasmids in Klebsiella pneumoniae. Trends Microbiol. 2021;29(1):65–83. doi:10.1016/j.tim.2020.04.012
5. Tian D, Liu X, Chen W, et al. Prevalence of hypervirulent and carbapenem-resistant Klebsiella pneumoniae under divergent evolutionary patterns. Emerg Microbes Infect. 2022;11(1):1936–1949. doi:10.1080/22221751.2022.2103454
6. Bialek-Davenet S, Criscuolo A, Ailloud F, et al. Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg Infect Dis. 2014;20(11):1812–1820. doi:10.3201/eid2011.140206
7. Gu D, Dong N, Zheng Z, et al. A fatal outbreak of st11 carbapenem-resistant hypervirulent Klebsiella pneumoniae in a Chinese hospital: a molecular epidemiological study. Lancet Infect Dis. 2018;18(1):37–46. doi:10.1016/S1473-3099(17)30489-9
8. Zhao Y, Zhang X, Torres V, et al. An outbreak of carbapenem-resistant and hypervirulent Klebsiella pneumoniae in an intensive care unit of a major teaching hospital in Wenzhou, China. Front Public Health. 2019;7:229. doi:10.3389/fpubh.2019.00229
9. Siu LK, Yeh KM, Lin JC, Fung CP, Chang FY. Klebsiella pneumoniae liver abscess: a new invasive syndrome. Lancet Infect Dis. 2012;12(11):881–887. doi:10.1016/S1473-3099(12)70205-0
10. Lederman ER, Crum NF. Pyogenic liver abscess with a focus on Klebsiella pneumoniae as a primary pathogen: an emerging disease with unique clinical characteristics. Am J Gastroenterol. 2005;100(2):322–331. doi:10.1111/j.1572-0241.2005.40310.x
11. Fang CT, Lai SY, Yi WC, Hsueh PR, Liu KL, Chang SC. Klebsiella pneumoniae genotype k1: an emerging pathogen that causes septic ocular or central nervous system complications from pyogenic liver abscess. Clin Infect Dis. 2007;45(3):284–293. doi:10.1086/519262
12. Patel PK, Russo TA, Karchmer AW. Hypervirulent Klebsiella pneumoniae. Open Forum Infect Dis. 2014;1(1):ofu028. doi:10.1093/ofid/ofu028
13. Saccente M. Klebsiella pneumoniae liver abscess, endophthalmitis, and meningitis in a man with newly recognized diabetes mellitus. Clin Infect Dis. 1999;29(6):1570–1571. doi:10.1086/313539
14. Kuramochi G, Takei SI, Sato M, Isokawa O, Takemae T, Takahashi A. Klebsiella pneumoniae liver abscess associated with septic spinal epidural abscess. Hepatol Res. 2005;31(1):48–52. doi:10.1016/j.hepres.2004.09.006
15. CLSI. Performance standards for antimicrobial susceptibility testing. In: Clinical and Laboratory Standards Institute. Wayne, PA: CLSI; 2022:M100–M128.
16. Zhou Y, Ai W, Cao Y, et al. The co-occurrence of ndm-5, mcr-1, and fosa3-encoding plasmids contributed to the generation of extensively drug-resistant Klebsiella pneumoniae. Front Microbiol. 2022;12. doi:10.3389/fmicb.2021.811263
17. Tian D, Wang W, Li M, et al. Acquisition of the conjugative virulence plasmid from a cg23 hypervirulent Klebsiella pneumoniae strain enhances bacterial virulence. Front Cell Infect Microbiol. 2021;11:752011. doi:10.3389/fcimb.2021.752011
18. Hughes C, Phillips R, Roberts AP. Serum resistance among Escherichia coli strains causing urinary tract infection in relation to o type and the carriage of hemolysin, colicin, and antibiotic resistance determinants. Infect Immun. 1982;35(1):270–275. doi:10.1128/iai.35.1.270-275.1982
19. Wang M, Earley M, Chen L, et al. Clinical outcomes and bacterial characteristics of carbapenem-resistant Klebsiella pneumoniae complex among patients from different global regions (crackle-2): a prospective, multicentre, cohort study. Lancet Infect Dis. 2021;22:401–412. doi:10.1016/S1473-3099(21)00399-6
20. Lam M, Wick RR, Wyres KL, et al. Genetic diversity, mobilisation and spread of the yersiniabactin-encoding mobile element icekp in Klebsiella pneumoniae populations. Microb Genom. 2018;4(9). doi:10.1099/mgen.0.000196
21. Chen L, Mathema B, Chavda KD, Deleo FR, Bonomo RA, Kreiswirth BN. Carbapenemase-producing Klebsiella pneumoniae: molecular and genetic decoding. Trends Microbiol. 2014;22(12):686–696. doi:10.1016/j.tim.2014.09.003
22. Fu P, Tang Y, Li G, Yu L, Wang Y, Jiang X. Pandemic spread of bla((kpc-2)) among Klebsiella pneumoniae st11 in China is associated with horizontal transfer mediated by incfii-like plasmids. Int J Antimicrob Agents. 2019;54(2):117–124. doi:10.1016/j.ijantimicag.2019.03.014
23. Tian D, Wang M, Zhou Y, Hu D, Ou HY, Jiang X. Genetic diversity and evolution of the virulence plasmids encoding aerobactin and salmochelin in Klebsiella pneumoniae. Virulence. 2021;12(1):1323–1333. doi:10.1080/21505594.2021.1924019
24. Xu Y, Zhang J, Wang M, et al. Mobilization of the nonconjugative virulence plasmid from hypervirulent Klebsiella pneumoniae. Genome Med. 2021;13(1):119. doi:10.1186/s13073-021-00936-5
25. Cienfuegos-Gallet AV, Zhou Y, Ai W, Kreiswirth BN, Yu F, Chen L. Multicenter genomic analysis of carbapenem-resistant Klebsiella pneumoniae from bacteremia in China. Microbiol Spectr. 2022;10(2). doi:10.1128/spectrum.02290-21
26. Lin TL, Lee CZ, Hsieh PF, Tsai SF, Wang JT. Characterization of integrative and conjugative element icekp1-associated genomic heterogeneity in a Klebsiella pneumoniae strain isolated from a primary liver abscess. J Bacteriol. 2008;190(2):515–526. doi:10.1128/JB.01219-07
27. Zhou K, Xiao T, David S, et al. Novel subclone of carbapenem-resistant Klebsiella pneumoniae sequence type 11 with enhanced virulence and transmissibility, China. Emerg Infect Dis. 2020;26(2):289–297. doi:10.3201/eid2602.190594
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.