Back to Journals » Breast Cancer: Targets and Therapy » Volume 18
Characterization and Cytotoxicity of West Java Coffee Extracts, Chlorogenic Acid, Caffeine on MDA-MB-231, MCF-7 Cell Lines, and Their Impact on MYC and AXIN2 Wnt Pathway Gene Expression
Authors Aborziza M, Zuhrotun A ![]() , Novianto A, Amalia R
, Novianto A, Amalia R ![]() , Fakih TM, Muchtaridi M
, Fakih TM, Muchtaridi M ![]()
Received 2 March 2026
Accepted for publication 29 April 2026
Published 26 May 2026 Volume 2026:18 606305
DOI https://doi.org/10.2147/BCTT.S606305
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Pranela Rameshwar
Mohamed Aborziza,1– 3 Ade Zuhrotun,4 Agung Novianto,5 Riezki Amalia,5,6 Taufik Muhammad Fakih,2,7 Muchtaridi Muchtaridi2,8
1Doctoral Program of Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, 45363, Indonesia; 2Department of Pharmaceutical Analysis and Medicinal Chemistry, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, 45363, Indonesia; 3Department of Medicinal Chemistry, Faculty of Pharmacy, Elmergib University, Alkhoms, 40414, Libya; 4Department of Biological Pharmacy, Faculty of Pharmacy, Universitas Padjadjaran, Sumedang, 45363, Indonesia; 5Laboratory of Translational Pharmaceutical Research, Faculty of Pharmacy, Universitas Pad-Jadjaran, Sumedang, 45363, Indonesia; 6Department of Pharmacology and Clinical Pharmacy, Faculty of Pharmacy, Universitas Pad-jadjaran, Sumedang, 45363, Indonesia; 7Department of Pharmacy, Faculty of Mathematics and Natural Sciences, Universitas Islam Bandung, Bandung, 40116, Indonesia; 8Research Collaboration Centre for Radiopharmaceuticals Theranostic, National Research and Innovation Agency (BRIN), Jl. Raya Bandung-Sumedang, Sumedang 45363, Indonesia
Correspondence: Muchtaridi Muchtaridi, Department of Pharmaceutical Analysis and Medicinal Chemistry, Faculty of Pharmacy, Universitas Padjadjaran, Jl. Raya Bandung Sumedang KM 21, Sumedang, 45363, Indonesia, Tel +62 813-9449-5569, Email [email protected]
Background: Coffee bean extracts have been widely investigated for their potential anticancer properties, particularly due to bioactive compounds such as chlorogenic acid and caffeine. Breast cancer development involves complex genetic and signaling alterations, including dysregulation of the Wnt/β-catenin pathway.
Purpose: To determine the phytochemical composition and quantify caffeine and chlorogenic acid in West Java coffee bean extracts. The study assessed their effects on the viability of MDA-MB-231 and MCF-7 breast cancer cells, and examined impacts on the Wnt pathway, focusing on MYC and AXIN2 expression.
Methods: Phytochemical screening and HPLC analysis are used to measure chlorogenic acid and caffeine in green and roasted coffee extracts. Cytotoxic activity and IC50 values in MDA-MB-231 and MCF-7 breast cancer cells were assessed with the WST-8 assay. Gene expression of MYC and AXIN2 was analyzed by q-RT-PCR to assess modulation of the Wnt pathway. Cytotoxic selectivity was tested in HEK293 cells. Flow cytometry analyzes apoptosis and the cell cycle. Molecular mechanisms were explored using docking, 200-ns molecular dynamics simulations, and MM-PBSA binding free energy calculations targeting Tankyrase-1 and Estrogen Receptor Alpha.
Results: Roasting and extraction methods affected phytochemicals, with higher temperatures reducing chlorogenic acid levels. Coffee extracts showed cytotoxicity against MDA-MB-231 (IC50 258.4– 637 μg/mL) and MCF-7 (IC50 190.7– 686 μg/mL) cells, and minimal toxicity to HEK293 cells—Green Arabica and Robusta extracts, via maceration, notably downregulated MYC and slightly reduced AXIN2. Flow cytometry indicated increased late apoptosis and cell-cycle arrest. Docking analysis found that chlorogenic acid binds more strongly to Tankyrase-1 than ERα. MD and MM-PBSA confirmed stable, energetically favorable binding to Tankyrase-1 complex.
Conclusion: Coffee bean extracts, especially chlorogenic acid–rich green coffee, show selective antiproliferative effects in breast cancer cells and may act through Wnt signaling rather than estrogen receptors. More in vivo and mechanistic studies are needed to confirm their potential as complementary or preventive anticancer agents.
Keywords: breast cancer, chlorogenic acid, Wnt signaling pathway, Tankyrase-1 inhibition, molecular simulation, apoptosis
Introduction
Breast cancer is the top cancer in women and the leading cause of cancer death. In 2022, the WHO reported 2.3 million diagnoses and 670,000 deaths worldwide.1 The rise and death rates of cancers like breast cancer led to new drugs targeting cancer cells with minimal damage to healthy cells.2
Most current anticancer drugs are highly toxic, damaging both cancerous and healthy cells. Efforts continue to develop new drugs that target cancer cells while sparing normal cells, through chemical synthesis or the isolation of natural resources.3–5 Natural compounds from herbs, vegetables, fruits, seeds, flowers, spices, and nuts are vital for the development of broad-spectrum drugs. Studies show that about 50% of prescribed medicines are from natural compounds or their analogs. Additionally, 74% of commonly used drugs contain natural ingredients, and nearly 60% of new anticancer drugs are derived from natural products.6,7 Several natural anti-cancer medicines include vinca alkaloids (vincristine, vinblastine, vinorelbine, vindesine), taxanes (docetaxel, paclitaxel), anthracyclines (idarubicin, daunorubicin, epirubicin, doxorubicin, podophyllotoxin and derivatives (irinotecan, topotecan).8–13
Coffee is rich in polyphenols and alkaloids, including phenolic acids such as chlorogenic acid, coumarin, ferulic acid, and sinapic acid, as well as caffeine. Among these, chlorogenic acid is particularly abundant and has been linked to potent antioxidant and anticancer properties.14–17 Studies have found that green coffee bean extract can protect normal kidney cells (HEK293) from toxic damage by reducing oxidative stress and inflammation. While some coffee by-products have shown cytotoxic effects on certain cell lines under specific conditions, the primary impact is renoprotective, as they improve kidney function in animal models by scavenging free radicals and reducing cell death.18 Coffee bean extracts have shown promising effects against breast cancer cell lines, including MDA-MB-231 and MCF-7.19 Bioactive compounds such as caffeine and caffeic acid have been reported to inhibit cell growth, with studies specifically demonstrating their suppressive effects on MDA-MB-231 and MCF-7 cells.20–22 Beyond its well-known role as a stimulant, caffeine exhibits anticancer potential against triple-negative breast cancer (TNBC) by inducing apoptosis, modulating telomerase activity, disrupting cellular metabolism, and sensitizing cancer cells to chemotherapy through interference with DNA repair.23 At higher concentrations, caffeine induces morphological changes, such as cell rounding and detachment, further indicating its cytotoxic effects on hormone receptor-positive MCF-7 and hormone receptor-negative MDA-MB-231 cells.24,25
The Wnt pathway is vital for breast cancer cell proliferation, metastasis, and the maintenance of stemness, all of which are crucial for tumor growth and spread.4,5 In triple-negative breast cancer (TNBC), the Wnt/β-catenin pathway is linked to resistance against chemotherapy and targeted therapies. It interacts with other pathways, such as receptor tyrosine kinase (RTK) signaling, to promote survival and metastasis.9,10 The Wnt/β-catenin pathway plays a crucial role in regulating key processes in breast cancer cells, including proliferation, differentiation, and apoptosis, as observed in the MCF-7 cell line. Activation begins when Wnt ligands bind to receptors, stabilizing β-catenin, which then moves to the nucleus to activate growth genes. In MCF-7 cells, Wnt signaling can be modulated by drugs such as lobaric acid and dendrosomal nanocurcumin, which suppress the pathway, inducing apoptosis and inhibiting cell growth.26 WNT signaling influences the immune microenvironment and immune cell infiltration in breast cancer, thereby affecting the response to immunotherapy. Although crucial, no WNT inhibitors are approved yet. Research continues to develop pathway-targeting drugs to enhance therapy outcomes.12,13
Chlorogenic acid (CGA) blocks the Wnt/β-catenin signaling pathway in breast cancer cells mainly by decreasing the expression of the coreceptor LRP6. This process suppresses epithelial-mesenchymal transition (EMT), diminishes cancer cell migration and invasion, and hampers tumor growth. Additionally, caffeine can elevate the BAX/BCL2 mRNA ratio, indicating apoptosis, and, when combined with coffee extracts, reduces levels of cytoplasmic and nuclear β-catenin, a critical element of the Wnt pathway.27 The antioxidant properties of coffee bean extract may help modulate signaling pathways, such as the Wnt pathway, by reducing oxidative stress, which is known to influence cancer progression.21 This compound in coffee bean extract modulates the Wnt pathway, notably inhibiting the Wnt/β-catenin pathway that is upregulated in various cancers, including breast cancer. MYC and AXIN2 are general Wnt target genes often used as an indicator of canonical Wnt activity.27 Some studies suggest that drinking coffee is connected to a lower risk of certain types of breast cancer. However, the exact mechanisms, including how Wnt signaling is involved, remain unclear.10,28
After years of focusing mainly on endocrine and cytotoxic therapies, we are now seeing targeted and pathway-specific treatments becoming part of therapeutic targets. This exciting progress has been made possible by our growing understanding of the detailed molecular workings of signaling pathways involved in initiation, invasion, and migration. Truly understanding these complex mechanisms inside cells is crucial for developing therapies that target specific pathways. This highlights the ongoing need for research to understand the effects of natural products and therapeutic substances on intracellular pathways.
The anti-breast cancer effects of coffee bean extracts remain insufficiently characterized, particularly regarding their underlying molecular mechanisms. This study aimed to evaluate the cytotoxic activity, induction of apoptosis, and cell cycle inhibition of green- and medium-roasted Arabica and Robusta coffee bean extracts from West Java, prepared by maceration and decoction, in MDA-MB-231 and MCF-7 breast cancer cell lines. Additionally, some in vitro studies investigated the modulation of Wnt/β-catenin–related genes, specifically MYC and AXIN2, to elucidate potential involvement of signaling pathways. To further clarify the molecular basis of these effects, computational analyses, including molecular docking, molecular dynamics simulations, and MM-PBSA binding free energy calculations, were performed targeting Tankyrase-1 and Estrogen Receptor Alpha. By integrating in vitro and in silico approaches, this study aims to provide mechanistic insight into the anticancer potential of chlorogenic acid–rich coffee extracts.
Materials and Methods
Chemicals and Reagents
The following reagents were obtained: Dulbecco’s Modified Eagle’s Medium (DMEM), fetal bovine serum (FBS), and antibiotics (100 U/mL penicillin and 0.1 mg/mL streptomycin) from Sigma-Aldrich; WST-8 from Dojindo Laboratories; and dimethyl sulfoxide (DMSO) from Phytotecs Lab. Chlorogenic acid was purchased from Chengdu Biopurity Phytochemicals Ltd. GENEzol™ Reagent from Geneaid; Nuclease-Free Water from Promega; ReverTra Ace qPCR RT Master Mix and gDNA Remover from Toyobo; SYBR no-ROX kit (Bioline Meridian); ACTB, MYC and AXIN2 primers from Integrated DNA Technologies (IDT); FITC Annexin V Apoptosis Detection Kit I, and PI/RNase staining buffer from BD Pharmingen; Additional reagents, including aquadest, 70% ethanol, Dragendorff reagent, Mayer reagent, NaOH solution, magnesium, 5N HCl, FeCl3 solution, dilute HCl, 1% gelatin solution, anisaldehyde–sulfuric acid, Liebermann–Burchard reagent, chloroform, isopropanol, were provided by the Pharmaceutical Biology Laboratory, Faculty of Pharmacy, Universitas Padjadjaran. The Central Laboratory of Universitas Padjadjaran, Indonesia, supplied caffeine, methanol, and 0.1% trifluoroacetic acid.
Plant Materials
Coffee bean samples were collected from farms in West Java, Indonesia. Arabica coffee beans were obtained from Manglayang Mountain on 19 October 2023, while Robusta coffee beans were collected from Puntang Mountain, Mekarjaya, on 16 December 2023. Seed specimens were authenticated at the Herbarium, Department of Biology, Universitas Padjadjaran, Indonesia, and identified as Coffea arabica L. and Coffea canephora Pierre ex A. Froehner (synonyms: Coffea bourbonica Pharm. ex Wehmer and Coffea robusta L. Linden). Both Arabica and Robusta beans, in green and medium-roasted forms, were subsequently ground into medium particle-size powder.
Decoction Extraction Method
The extraction was performed with minor modifications as described by Parnomo (2021). Water extraction was performed using a 5:1 water-to-coffee ratio at 100°C for 30 minutes, followed by filtration. The filtrate was then dried using a freeze dryer.29
Maceration Extraction Method
The maceration extraction method, as described by Hartanti & Arif (2023), was followed with slight modifications. It used a 5:1 (v/w) ethanol-to-coffee ratio at room temperature and dried the mixture with a rotary evaporator to produce a thick extract.30
Phytochemical Screening
Identification of group compounds contained in the coffee extract, such as alkaloids, quinones, flavonoids, saponins, Polyphenols, Tannins, and Terpenoids, was carried out based on the modified method of Praherdiono & Adi 2008.31
HPLC with UV/VIS Detector Analysis
The HPLC analysis was performed according to the method described in the reference, with adjustments made as necessary. It used an HPLC Waters Alliance e2695 with a UV-Vis 2489 detector set at 272 nm for caffeine and 326 nm for chlorogenic acid (CA). Separation was performed using an Eclipse Plus C18 column (250 mm × 4.6 mm, 5 μm) with a 10 μL injection, a column temperature of 25 °C, and a data rate of 10 Hz. Mobile phases were methanol/water (60:40) for caffeine and methanol/water (40:60) with 0.1% TFA for CA. The mobile phase was degassed and delivered at 1.0 mL/min.32
Cell Culture
The MDA-MB-231 and MCF-7 breast cancer cell lines, along with the HEK 293 normal kidney cell line, were obtained from the European Collection of Authenticated Cell Cultures (ECACC). Cells were routinely maintained in Dulbecco’s Modified Eagle’s Medium–High Glucose (DMEM) supplemented with 10% fetal bovine serum and antibiotics (100 U/mL penicillin and 0.1 mg/mL streptomycin), and cultured under standard conditions of 5% CO2.33
Determining IC50 Through Cell Viability Assay
Cells were seeded in 96-well plates, incubated for 24 h, and then treated with coffee bean extracts at concentrations ranging from 0 to 1000 µg/mL (three replicates per concentration) for an additional 24 h. Subsequently, 10 µL of WST-8 reagent was added to each well, and the plate was incubated for 24 h. Absorbance was measured at 450 nm with a reference wavelength of 620 nm using a Tecan Infinite Pro spectrophotometer.33 Cell viability was expressed as the ratio of absorbance in treated cells to that in untreated control cells. Absolute IC50 values were calculated using GraphPad Prism version 10.4.1 (p < 0.05).
Quantitative RT-PCR
The breast cancer cells were seeded and cultured to achieve 60–70% confluence, treated with samples and standards at concentrations of 100 µg/mL and 200 µg/mL (the concentrations chosen depend on the lowest concentration that causes IC50 in the cytotoxicity test), (three replicates per concentration), and incubated for 8 hours. Then, RNA was extracted using the GENEzol reagent, and cDNA was synthesized using the ReverTra Ace qPCR RT Master Mix and gDNA Remover. Quantitative PCR (qRT-PCR) was performed using the SYBR no-ROX according to the manufacturer’s protocol. Expression quantification was performed using the Livak method.34 The primers used were human AXIN2 (forward 5′-TCCCAGACTCAGTGGGAAGA-3′, reverse 5′-GCCTGGTGTTGGAAGAGACA-3′), human MYC (forward 5′-CCTCGGATTCTCTGCTCTCC-3′, reverse 5′-TTTCTTCCTCATCTTCTTGTTCCTC-3′), and housekeeping gene human ACTB (forward 5′-AAGATCAAGATCATTGCTCC3′, reverse 5′-TCCTAACAGCGCCTAGAAGC-3′).
Apoptosis Analysis
The cells were seeded and cultured to achieve 60–70% confluence, treated with samples and standards at concentrations of 100 µg/mL and 200 µg/mL (the concentrations chosen depend on the lowest concentration that causes IC50 in the cytotoxicity test), (three replicates per concentration), and then incubated for 24 hours. The cells were washed with PBS and detached using 0.125% trypsin/EDTA by incubation at 37 °C for 3 minutes. The cells were then collected and stained with Annexin V-FITC/PI (BD Pharmingen) according to the manufacturer’s instructions. The stained cells were then quantified using flow cytometry with a Beckton Dickinson Accuri C6 and analyzed via BD Accuri C6 Software relative to the cell control.35
Cell Cycle Analysis
The cells were treated with the same procedure as the Apoptosis assay. The collected cells were resuspended in cold 70% ethanol and incubated for 24 hours at 4°C. The cells were then treated with RNase (200 μg/mL) for 30 minutes at 37°C and subsequently stained with propidium iodide (50 μg/mL). The DNA content was analyzed by flow cytometry using a Beckton Dickinson Accuri C6. The proportions of apoptotic cells (2n but < 4n) and cells in the G2/M phase (4n) were determined using BD Accuri C6 Software relative to the cell control.36
Statistical Analysis
Statistical analysis was performed using one-way ANOVA in GraphPad Prism version 10.4.1. The results are considered significant when p < 0.05.
Molecular Docking Analysis
Molecular docking simulations were performed to investigate the interaction of chlorogenic acid with Tankyrase-1 (PDB ID: 3UDD) and Estrogen Receptor Alpha (PDB ID: 3ERT). The three-dimensional crystal structures were retrieved from the Protein Data Bank (https://www.rcsb.org).37 Protein preparation was conducted using AutoDock Tools version 1.5.7, which included removal of co-crystallized water molecules, addition of polar hydrogens, and assignment of Kollman united atom charges.38 The native ligands were retained temporarily to define the active site grid coordinates and were subsequently removed prior to docking. The three-dimensional structure of chlorogenic acid was downloaded from PubChem (CID: 1794427)39 and energy-minimized using Chem3D version 22.0 before conversion to PDBQT format.40 Docking simulations were performed using AutoDock Vina version 1.2.3.41 The grid box was centered at the coordinates of the native ligand binding site with dimensions adjusted to fully cover the catalytic pocket. The exhaustiveness parameter was set to 8, and the maximum number of binding modes was set to 9. The best docking pose was selected based on the lowest binding affinity (kcal/mol) and interaction relevance with key residues. Protein–ligand interactions were visualized and analyzed using Discovery Studio Visualizer version 21.1.0.42
Molecular Dynamics Simulation
Molecular dynamics simulations were carried out using GROMACS version 2024.3 to evaluate the structural stability of the docked complexes.43,44 The GROMOS96 54a7 force field was applied for protein parameterization, and ligand topology parameters were generated using the PRODRG server with manual correction of charges to ensure compatibility. Each protein–ligand complex was placed in a cubic simulation box with a minimum distance of 1.0 nm between the protein surface and box edge. The system was solvated using the SPC water model under periodic boundary conditions. Counter ions (Na⁺ or Cl−) were added to neutralize the system charge. Energy minimization was. Counterions (Na⁺ or Cl−) were added to neutralize the system’s charge. Energy minimization was performed using the steepest descent algorithm until the maximum force fell below 1000 kJ/mol/nm. The system was equilibrated under NVT ensemble for 100 ps at 300 K using the V-rescale thermostat, followed by NPT equilibration for 100 ps at 1 bar pressure using the Parrinello–Rahman barostat. Long-range electrostatic interactions were treated using the Particle Mesh Ewald (PME) method with a cutoff of 1.0 nm. The LINCS algorithm was applied to constrain all bond lengths, allowing a 2-fs integration time step. Production MD simulations were conducted for 200 ns. Structural stability was assessed through root mean square deviation (RMSD) and root mean square fluctuation (RMSF) analyses using built-in GROMACS tools.
MM-PBSA Binding Free Energy Calculation
Binding free energy calculations were performed using the g_mmpbsa tool version 1.6 integrated with GROMACS 2024.3.45,46 Snapshots were extracted from the last 50 ns of the equilibrated trajectories at 100 ps intervals for energy analysis. The MM-PBSA method was used to compute van der Waals, electrostatic, polar solvation, and solvent-accessible surface area (SASA) energy components. The polar solvation energy was calculated using the Poisson–Boltzmann equation, while the nonpolar solvation energy was estimated based on SASA. The total binding free energy was determined by summing molecular mechanics energy contributions and solvation energies. Standard deviation values were calculated to evaluate energetic stability across sampled frames. Negative total binding free energy values indicated favorable complex formation. Energy decomposition results were compared between the Tankyrase-1 and Estrogen Receptor Alpha complexes to assess their relative thermodynamic stability.
Results
Affects the Extraction Method on the Percentage of Green and Roasted Coffee Extract Yield
Table 1 summarizes the extraction yields of various coffee samples and the extraction methods employed in this study, showing that the yield of roasted coffee extract was slightly higher than that of green coffee extract, and that the decoction method yielded more extract than the maceration method. The extract yield was calculated using the following formula:

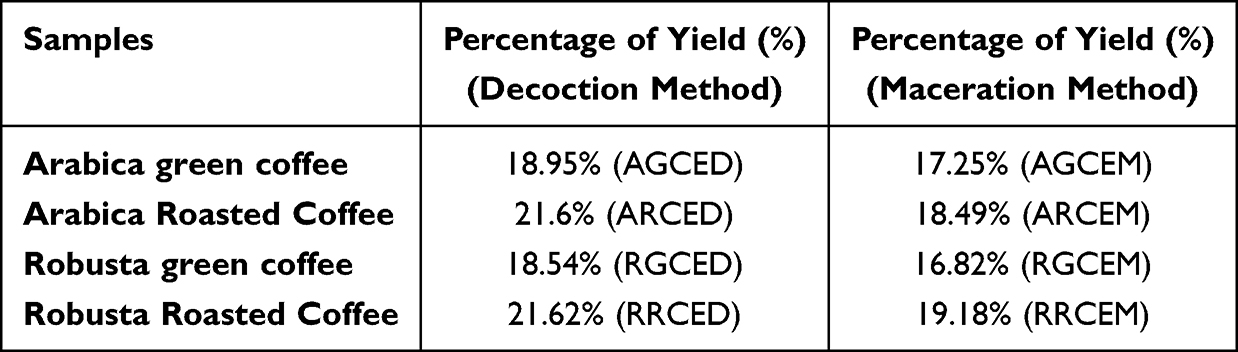 |
Table 1 Percentage of Extract Yield of West Java Arabica and Robusta Coffee Beans |
Phytochemical Screening
The compounds tested on powder and extracts of coffee bean samples included alkaloids, quinones, flavonoids, polyphenols, saponins, tannins, monoterpenoids, sesquiterpenoids, triterpenoids, and steroids. The qualitative tests used the reagents listed in Table 2. Depending on the density of requirements, Flavonoids, Saponins, Monoterpenoids, and Sesquiterpenoids showed slightly lower requirement density in extracts than in raw coffee powder, except in green coffee bean extract, where the decoction method did not reveal the requirements. Showed a negative result in Tannins, Triterpenoids, and Steroids tests for both samples of coffee powder and samples of coffee extract.
 |
Table 2 Phytochemical Screening Qualitative Results of Coffee Bean Extracts and Powder |
Determination of Standard Solution Using HPLC
Caffeine and chlorogenic acid are two of the most essential compounds in coffee, with wide-ranging medical applications and effects on various cancer cell models. The standard levels of CF and CA are outlined in Table 3 and shown in Figure 1. The analyses were identified by comparing the retention times of the coffee samples with those of the standards CF and GA.
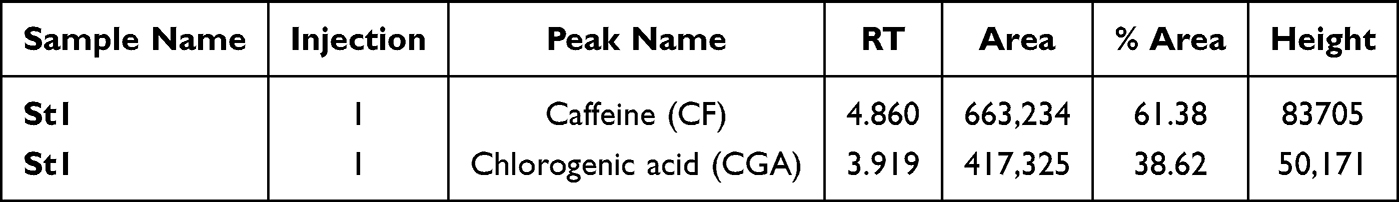 |
Table 3 Peak Names: Caffeine and Chlorogenic Acid (CGA) Standards |
 |
Figure 1 The HPLC chromatogram of caffeine standard (RT 3.919) and chlorogenic acid standard (RT 4.860). |
Caffeine (CF) Content in Coffee Bean Extract Samples
The experimental results for the caffeine analysis in coffee samples, as well as the concentrations of caffeine in percentage (%) and mg/L in coffee bean extract samples, are reported in Table 4, Figures 2 and 3.
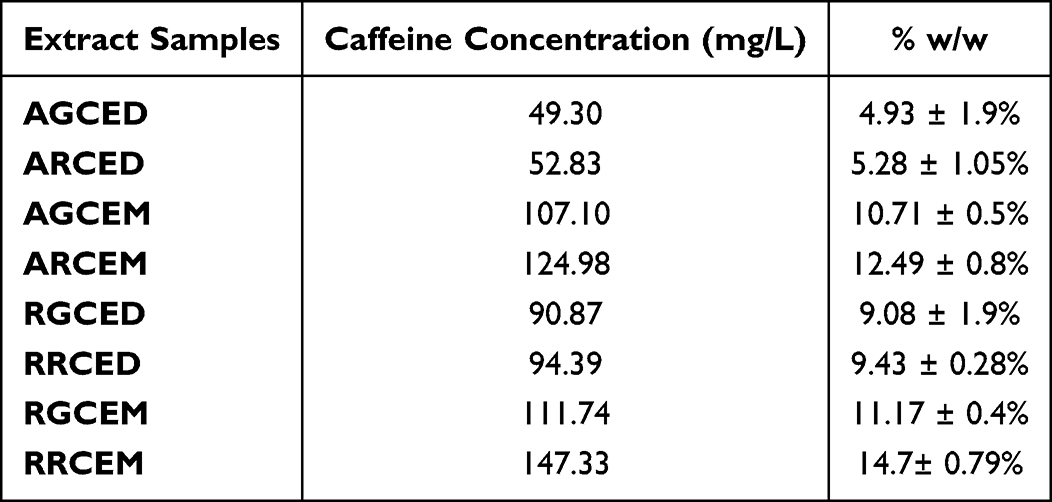 |
Table 4 The Concentration of Caffeine (CF) in Coffee Bean Extract Samples |
 |
Figure 2 HPLC graph illustrating chlorogenic acid (CGA) and caffeine (CF) contents in Arabica coffee extract samples: (A) Arabica green coffee extract utilizing the decoction method (AGCED), (B) Arabica roasted coffee extract employing the decoction method (ARCED), (C) Arabica green coffee extract prepared via the maceration method (AGCEM), and (D) Arabica roasted coffee extract using the maceration method (ARCEM). |
 |
Figure 3 HPLC graph illustrating chlorogenic acid and caffeine levels in Robusta coffee extract samples. (A) Green Robusta coffee extracted via the decoction method (RGCED). (B) Roasted Robusta coffee extracted via decoction method (RRCED). (C) Green Robusta coffee extracted via the maceration method (RGCEM). (D) Roasted Robusta coffee extracted via the maceration method (RRCEM). |
Chlorogenic Acid (CA) Content in Coffee Bean Extract Samples
The experimental results for the CA analysis in coffee samples and the concentrations of CA in percentage (%) and mg/L in coffee bean extract samples are reported in Table 5, Figures 2 and 3.
 |
Table 5 The Concentration of Chlorogenic Acid (CA) in Coffee Bean Extract Samples |
Cytotoxicity of Coffee Bean Extracts on MDA-MB-231 and MCF-7 Breast Cancer Cell Lines
Coffee extracts and standards (chlorogenic acid and caffeine) were tested for cytotoxic effects on non-hormone-related factors MDA-MB-231 breast cancer cells and hormone-related factors MCF-7 breast cancer cells using the WST-8 method. Cell viability was assessed after 24 hours of treatment with various concentrations of the compound. The results, presented in Table 6 and Figure 4, indicate that the viability of the human breast carcinoma cell lines decreased significantly in a concentration-dependent manner.
 |
Table 6 IC50 Cytotoxicity Values of Coffee Bean Extracts and Standard on MDA-MB-231 and MCF-7 Breast Cancer Cells |
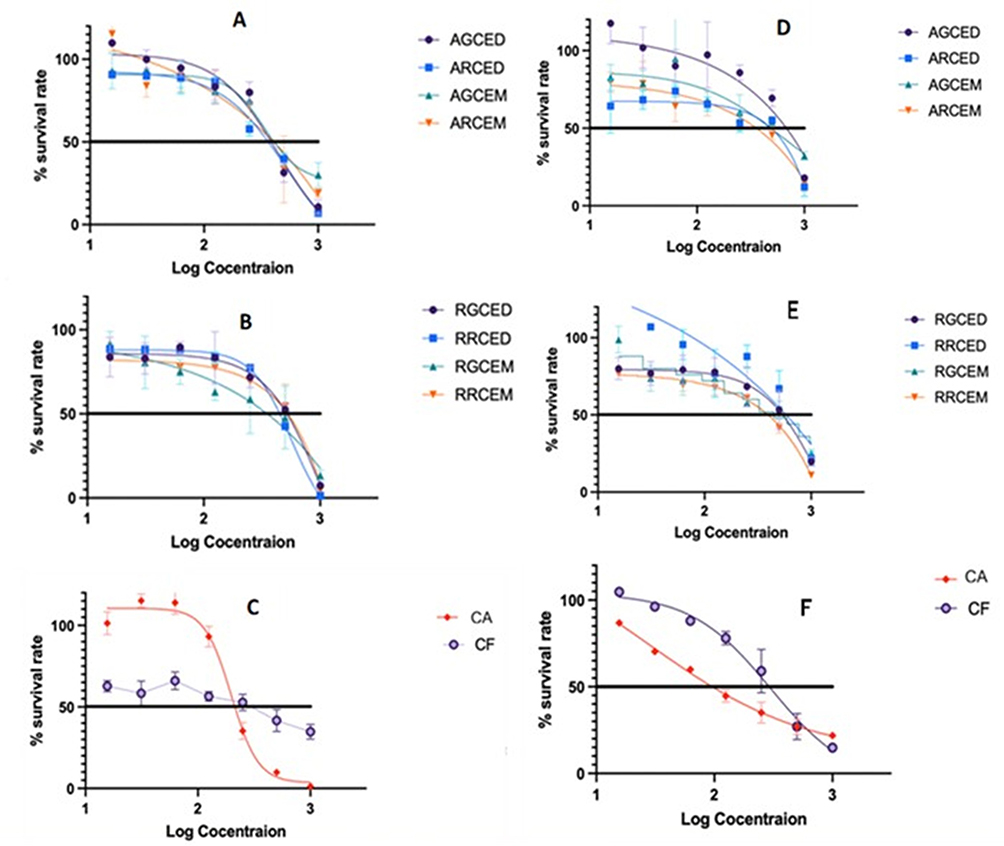 |
Figure 4 The cytotoxicity IC50 values of Arabica coffee extract (A), Robusta coffee extract (B), and CA and CF standards (C) on MDA-MB-231 breast cancer cells, as well as the IC50 values of Arabica coffee extract (D), Robusta coffee extract (E), and CA and CF standards (F) on MCF-7 breast cancer cells at various concentrations during a 24-hour treatment, are presented. Data are expressed as mean ± standard error of the mean (SEM) (n=3). |
Both Arabica and Robusta coffee extracts had cytotoxicity effects less than those of the standards. The cytotoxicity of chlorogenic acid on MCF-7 is the strongest, with an IC50 of 35.02 µg/mL. Arabica roasted coffee extract with the maceration method (ARCEM) exhibited the most potent cytotoxicity effect on MDA-MB-231 cells, with an IC50 value of 258.4 µg/mL, whereas Robusta roasted coffee extract with the decoction method (RRCED) showed cytotoxicity on MCF-7 cells, with an IC50 value of 190.7.
qRT-PCR MYC and AXIN2 Gene Expression
The effect of chlorogenic acid, caffeine, and coffee bean extracts on MYC and AXIN2 gene expression in MDA-MB-231 and MCF-7 cells is shown in Figure 5. Coffee bean extracts significantly decreased MYC gene expression levels. Treatment with coffee bean extract reduced the MYC level range (from 0.77-fold to 0.15-fold) at 100 µg/mL and (from 0.32-fold to 0.05-fold) at 200 µg/mL in MDA-MB-231 cells. In MCF-7 cells, at 100 µg/mL, the range was (from 0.76-fold to 0.17-fold), and at 200 µg/mL, it decreased further (from 0.27-fold to 0.08-fold). The AXIN2 gene level range decreased by (1.33-fold to 0.48-fold) at 100 ppm and by (0.76-fold to 0.34-fold) at 200 µg/mL in MDA-MB-231 cells. In MCF-7 cells, AXIN2 levels decreased from (0.78-fold to 0.28-fold) at 100 µg/mL to (0.41-fold to 0.28-fold) at 200 µg/mL, compared to the control at the same concentrations. The most notable effects on MYC and AXIN2 gene expression were observed with Arabica green coffee extract obtained by maceration (AGCEM). For MYC, this extract caused a 0.25-fold decrease at 100 µg/mL and a 0.05-fold reduction at 200 µg/mL in MDA-MB-231 cells, and a decrease to 0.22-fold at 100 µg/mL and 0.08-fold at 200 µg/mL in MCF-7 cells. For AXIN2, the decreases were 0.48-fold at 100 µg/mL and 0.34-fold at 200 µg/mL in MDA-MB-231 cells, and 0.44-fold at 100 µg/mL and 0.29-fold at 200 µg/mL in MCF-7 cells. The best results with Robusta coffee beans came from Robusta green coffee extract via maceration (RGCEM), which decreased MYC expression by 0.15-fold at 100 µg/mL and 0.07-fold at 200 µg/mL in MDA-MB-231 cells, and by 0.17-fold at 100 µg/mL and 0.12-fold at 200 µg/mL in MCF-7 cells. For AXIN2, the reductions were 0.87-fold at 100 µg/mL and 0.36-fold at 200 µg/mL in MDA-MB-231, and 0.43-fold at 100 µg/mL and 0.28-fold at 200 µg/mL in MCF-7 cells (Figure 5A–F, all p < 0.05).
 |
Figure 5 The impact of coffee bean extracts, chlorogenic acid, and caffeine on the Wnt signaling pathways, specifically examining: (A) MYC gene expression in MDA-MB-231 cells; (B) AXIN2 gene expression in MDA-MB-231 cells; (C) MYC gene expression in MCF-7 cells; and (D) AXIN2 gene expression in MCF-7 cells, as measured by qRT-PCR following an 8-hour treatment. Cells were exposed to coffee bean extracts and standard compounds at concentrations of 100 µg/mL and 200 µg/mL. B-Actin was employed for normalization of relative gene expression. The data are presented as means with standard error of the mean (SEM). Statistical significance was defined as * P<0.05. Notes: (ns means P value not less than 0.05). The more symbols (*) are used, the lower the value of P becomes (less than 0.05). * P <0.05, ** P <0.005, *** P = 0.0001, **** P <0.0001. |
Cytotoxic Effects of Coffee Bean Extracts (AGCEM and RGCEM) on HEK 293 Kidney Normal Cells
Cytotoxicity was assessed using WST-8 to determine the IC50 of Arabica and Robusta extracts; the Robusta extracts showed the greatest inhibitory effect in the qRT-PCR test. Cell viability was assessed after 24 hours at various concentrations ranging from 0 to 2000 µg/mL. As shown in Figure 6, the results indicate that the selected samples are not cytotoxic to normal cells within the tested range, with an IC50 value exceeding 2000 µg/mL.
 |
Figure 6 The IC50 values for cytotoxicity of Arabica green coffee extract prepared via the maceration method (AGCEM) and Robusta green coffee extract prepared via the maceration method (RGCEM) on HEK 239 normal kidney cell lines. |
Effects of Green Coffee Bean Extracts (AGCEM and RGCEM) and Standards (CA and CF) on Cell Cycle Distribution
The effects of coffee bean extract, chlorogenic acid, and caffeine on cell cycle distribution and apoptosis in MCF-7 and MDA-MB-231 cells were examined after 24 hours of exposure to 100 µg/mL and 200 µg/mL concentrations of the extract (Figure 7A and B). In MCF-7 cells, coffee bean extracts induced a notable shift in cell cycle progression with increasing concentration, increasing the number of cells in the G1 and S phases. It decreased in the G2/M phase as the concentration rose. Conversely, caffeine exhibited effects opposite to those of chlorogenic acid: the number of cells increased in G1 and G2/M phases and decreased in the S phase with higher caffeine levels (p < 0.05). In MDA-MB-231 cells, Arabica green coffee bean extract prepared by maceration (AGCEM) showed effects similar to caffeine: the number of cells in the G1 phase declined, while counts in the S and G2/M phases increased with increasing concentration. Meanwhile, Robusta green coffee bean extract prepared via the maceration method (RGCEM) exhibited effects similar to those of chlorogenic acid; the number of cells in the G1 and G2/M phases decreased, whereas those in the S phase increased with increasing concentration (P < 0.05).
 |
Figure 7 Cell cycle analysis of MDA-MB-231 and MCF-7 cancer cell lines treated with Arabica and Robusta green coffee bean extracts prepared via the maceration method, as well as chlorogenic acid (CA) and caffeine (CF) at concentrations of 100 µg/mL and 200 µg/mL. The study examines the distribution of cells undergoing apoptosis and occupying various phases of the cell cycle. (A). The values are expressed as the mean ± standard error of the mean of three determinations. When indicated, they demonstrated a statistically significant difference (P < 0.05) compared with the cell control (B). |
Effects of Green Coffee Bean Extracts (AGCEM and RGCEM) and Standards (CA and CF) on Early and Late Apoptosis
Apoptotic Activity in Flow Cytometry Analysis: The apoptotic activity of green coffee bean extract, chlorogenic acid, and caffeine (at 100 and 200 µg/mL) was tested against MDA-MB-231 and MCF-7 cancer cell lines using flow cytometry (Figure 8A and B) shows the proportions of viable (Q1), early apoptotic (Q2), late apoptotic (Q3), and necrotic (Q4) cells. Generally, the percentage of living cells decreased as the concentrations of coffee bean extracts, chlorogenic acid, and caffeine increased, with a smaller decrease observed in MCF-7 cells than in MDA-MB-231 cells. The substances demonstrated concentration-dependent apoptotic activity, affecting both the early and late stages of apoptosis. Cells undergoing apoptosis were more often in the late phase than in the early phase. The effects of coffee bean extract, chlorogenic acid, and caffeine on the early and late apoptotic fractions in MCF-7 cells were significantly increased with increasing concentration, with more pronounced effects in MDA-MB-231 cells.
 |
Figure 8 The apoptosis study of MDA-MB-231 and MCF-7 cells involved exposure to Arabica and Robusta green coffee bean extracts prepared by maceration, along with chlorogenic acid (CA) and caffeine (CF) at concentrations of 100 µg/mL and 200 µg/mL. The analysis included the distribution of cells undergoing early and late apoptosis, as well as live cells not undergoing apoptosis, compared with the cell control, using flow cytometric analysis with Annexin-V FITC and propidium iodide (A). The values are presented as the mean ± standard error of the mean from three determinations and, where indicated, demonstrate a statistically significant difference (P < 0.05) relative to the cell control (B). Notes: (ns means P value not less than 0.05). The more symbols (*) are used, the lower the value of P becomes (less than 0.05). ** P <0.005, *** P = 0.0001, **** P <0.0001. |
Molecular Docking Analysis
These apoptotic findings are further supported by the molecular docking analysis, which revealed the potential molecular targets underlying the pro-apoptotic effects of chlorogenic acid (Table 7). In Tankyrase-1 (PDB ID: 3UDD), the native ligand demonstrated a strong binding affinity of −12.27 kcal/mol with a nanomolar inhibition constant, whereas chlorogenic acid showed a lower binding energy of −8.08 kcal/mol with a micromolar Ki value. Although weaker than the native ligand, chlorogenic acid formed multiple conventional hydrogen bonds with key residues, including TYR1213, GLY1196, GLY1211, and GLY1185, indicating stable anchoring within the catalytic pocket. Additional hydrophobic interactions with HIS1201, ALA1191, and ILE1192 contributed to complex stabilization. The predominance of hydrogen bonding interactions suggests that chlorogenic acid relies on polar contacts to maintain binding stability. Tankyrase-1 is known to regulate Wnt/β-catenin signaling, which plays a critical role in breast cancer cell survival and proliferation. Inhibition of Tankyrase-1 may promote β-catenin degradation, thereby suppressing pro-survival transcriptional programs. This molecular mechanism is consistent with the increased proportion of late apoptotic cells observed in flow cytometry analysis, particularly in MDA-MB-231 cells. The stronger apoptotic response in MDA-MB-231 cells compared with MCF-7 cells may reflect differences in Wnt pathway dependence between these subtypes. Therefore, partial inhibition of Tankyrase-1 by chlorogenic acid could mechanistically explain the concentration-dependent induction of apoptosis observed experimentally.
 |
Table 7 Molecular Docking Results of Chlorogenic Acid and Native Ligands Against Tankyrase-1 and Estrogen Receptor Alpha |
Similarly, docking analysis against Human Estrogen Receptor Alpha (PDB ID: 3ERT) provides insight into the differential response between ER-positive MCF-7 and ER-negative MDA-MB-231 cells. The native ligand 4-hydroxytamoxifen exhibited a binding affinity of −11.41 kcal/mol, whereas chlorogenic acid showed a weaker affinity of −6.07 kcal/mol with a micromolar Ki. Despite this lower affinity, chlorogenic acid formed several conventional hydrogen bonds with critical residues, including HIS524, GLU419, GLU353, and GLY420, as well as a hydrophobic interaction with MET421. Interaction with GLU353 is particularly noteworthy because this residue is essential for ligand recognition and receptor activation. Although chlorogenic acid does not bind as strongly as tamoxifen, its ability to interact within the ligand-binding domain suggests potential modulatory effects on ER signaling. The weaker apoptotic effect observed in MCF-7 cells compared to MDA-MB-231 cells may be attributed to the moderate binding strength toward ERα, resulting in partial rather than complete antagonism. In contrast, the greater apoptosis in MDA-MB-231 cells may be more closely associated with non–ER-mediated mechanisms, such as inhibition of the Wnt pathway. The higher percentage of late apoptosis indicates that sustained signaling disruption eventually drives cells beyond early apoptotic checkpoints. These combined in vitro and in silico findings suggest that chlorogenic acid exerts multitarget activity, contributing to apoptosis induction through Tankyrase-1–related pathways and partial modulation of estrogen receptor signaling.
Molecular Dynamics Simulation Analysis
The molecular dynamics simulation results of the Tankyrase-1–chlorogenic acid complex demonstrate an initial equilibration phase during the first 20 ns of the 200 ns trajectory. During this stage, the RMSD increases from approximately 0.2 nm to around 0.8 nm, indicating structural adaptation after ligand binding. Following equilibration, the RMSD stabilizes within a range of approximately 0.9–1.3 nm for most of the simulation period. Moderate fluctuations observed between 60 and 120 ns reflect conformational flexibility rather than structural instability. A slight increase toward the final phase of the trajectory does not suggest ligand dissociation but indicates normal dynamic motion within the protein structure. The absence of an abrupt structural deviation confirms the preservation of the global protein architecture throughout the simulation. The RMSF profile further reveals that most residues fluctuate by less than 0.15 nm, suggesting rigidity across the catalytic domain. Higher peaks are localized in loop or surface-exposed regions rather than within the core binding pocket. Importantly, residues previously identified in docking interactions exhibit relatively low fluctuations, indicating persistent ligand engagement. The stability of these interaction sites supports continuous hydrogen bonding and hydrophobic contacts during the simulation. Such controlled flexibility reflects a dynamically stable protein–ligand complex. These observations indicate that chlorogenic acid maintains stable occupancy of the Tankyrase-1 catalytic pocket, which may contribute to apoptosis induction.
The dynamic behavior of the Estrogen Receptor Alpha–chlorogenic acid complex is illustrated in Figure 9, which shows a comparatively low RMSD profile throughout the 200 ns simulation. After a brief equilibration period, the RMSD stabilizes between approximately 0.18 and 0.30 nm, indicating high structural stability. Slight deviations observed around 60 to 100 ns correspond to conformational adjustments within the ligand-binding domain. Throughout the simulation, the RMSD remains below 0.45 nm, confirming the absence of major structural perturbations. This limited deviation suggests that chlorogenic acid binding does not disrupt the global receptor conformation. The RMSF analysis shows that most residues fluctuate by less than 0.15 nm within the structured core region. Higher fluctuation peaks are mainly detected at terminal regions, which are inherently more flexible. Residues critical for ligand recognition exhibit relatively low fluctuation values, supporting stable positioning within the binding pocket. The maintained compactness of the receptor structure indicates persistent receptor–ligand interaction. This dynamic stability aligns with the moderate docking affinity previously observed. The absence of excessive flexibility suggests that chlorogenic acid exerts a modulatory rather than disruptive effect on ERα structure. These structural findings provide molecular-level support for the comparatively weaker apoptotic response observed in ER-positive breast cancer cells.
 |
Figure 9 RMSD and RMSF profiles of Tankyrase-1 (3UDD) and Estrogen Receptor Alpha (3ERT) in complex with chlorogenic acid during 200 ns simulation. |
MM-PBSA Binding Free Energy Calculation
The MM-PBSA results presented in Table 8 indicate that chlorogenic acid exhibits favorable binding energetics toward Tankyrase-1. The total binding free energy of −43.977 kJ/mol indicates spontaneous, thermodynamically stable complex formation. The van der Waals contribution (−103.023 kJ/mol) represents the dominant favorable interaction component, indicating that hydrophobic contacts play a crucial role in stabilizing the complex. Electrostatic interactions also contribute negatively (−21.971 kJ/mol), supporting the presence of stable polar interactions within the binding pocket. However, the polar solvation energy shows a positive value (94.956 kJ/mol), reflecting an unfavorable desolvation penalty upon complex formation. This effect is partially compensated by the negative SASA energy (−13.939 kJ/mol), which accounts for nonpolar solvation contributions. The balance between favorable intermolecular interactions and solvation penalties ultimately results in a negative total binding energy. The dominance of van der Waals interactions aligns with the docking analysis that identified multiple hydrophobic contacts within the catalytic pocket. The electrostatic contribution further supports the persistence of hydrogen-bonding interactions throughout the simulation. Although the standard deviation values indicate some energetic fluctuation, the overall energy profile remains favorable. These energetic characteristics are consistent with the stable RMSD and RMSF behavior observed during molecular dynamics simulation. The strong binding affinity toward Tankyrase-1 supports its proposed role in modulating apoptosis-related signaling pathways.
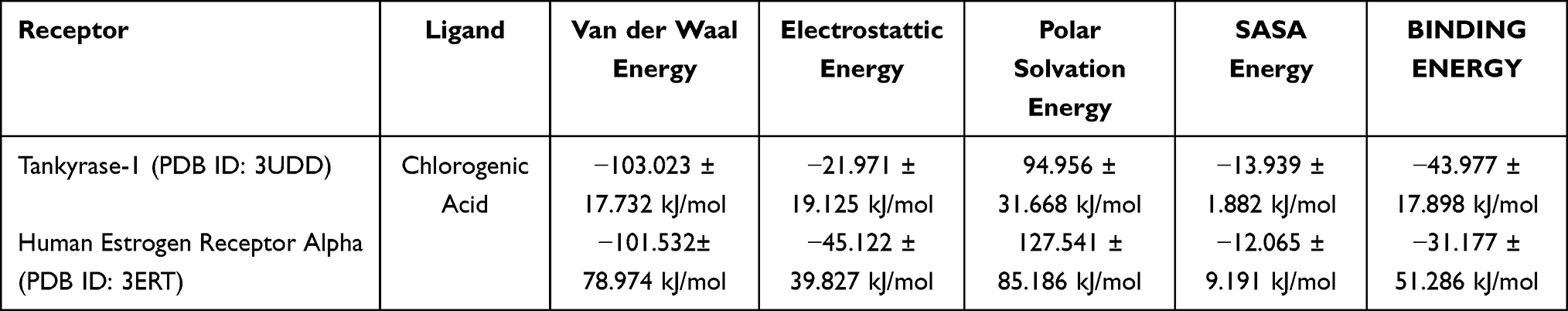 |
Table 8 MM-PBSA Binding Free Energy Decomposition of Chlorogenic Acid in Complex with Tankyrase-1 (3UDD) and Estrogen Receptor Alpha (3ERT) |
In contrast, the MM-PBSA analysis of the Estrogen Receptor Alpha complex shows a less favorable binding free energy of −31.177 kJ/mol. Although still negative, this value indicates weaker overall binding than that of the Tankyrase-1 complex. The van der Waals energy (−101.532 kJ/mol) remains a major stabilizing factor, similar in magnitude to that observed in the Tankyrase-1 system. Notably, the electrostatic contribution is more pronounced (−45.122 kJ/mol), suggesting stronger polar interactions within the ERα binding pocket. However, this favorable electrostatic term is counterbalanced by a significantly higher polar solvation energy (127.541 kJ/mol), thereby reducing the net binding strength. The SASA energy (−12.065 kJ/mol) provides only minor compensation for this solvation penalty. The relatively large standard deviation observed in the total binding energy indicates greater energetic fluctuation during the simulation. This variability may reflect dynamic adjustments within the ligand-binding domain. Despite stable structural behavior in the RMSD analysis, the energetic profile indicates a weaker thermodynamic driving force than that of Tankyrase-1. The lower binding affinity for ERα is consistent with the previously obtained moderate docking score. These findings provide molecular-level support for the comparatively weaker apoptotic response observed in ER-positive breast cancer cells. The MM-PBSA data therefore reinforce the hypothesis that chlorogenic acid may exert stronger functional influence through Tankyrase-1 modulation than through direct ERα inhibition.
Discussion
Coffee, a popular beverage made from beans worldwide, has been studied for its effects on cancer cells.35 Extracts from coffee beans exhibit anti-cancer effects by inducing apoptosis, promoting cellular senescence, and generating reactive oxygen species, all of which slow down the growth and spread of cancer cells. The key compounds that make a difference include caffeine, chlorogenic acids, kahweol, cafestol, trigonelline, lignin, and melanoidins. These compounds act in various ways, including inhibiting the cell cycle, reducing inflammation, and serving as potent antioxidants; however, the metastatic mechanisms of coffee bean extract remain poorly understood.47 This study examined two coffee types from West Java (Arabica and Robusta). We used green and medium-roasted coffees, extracted using two methods, to investigate the impact of extraction and roasting on phytochemical levels, specifically chlorogenic acid and caffeine. Results show decoction with water yields higher extract amounts than other methods with non-polar solvents. Specific yields were 81.26 ± 19.23 mg/g with methanol at 100 °C, 19.12 ± 1.28 mg/g with dichloromethane at 120°C, and 1.76 ± 0.93 mg/g with n-hexane at 120 °C.48 In coffee roasting, CF slightly increased, but CA degraded with brewing temperature. CA levels dropped in roasted coffee, while melanoidins and Maillard products formed.49
The results of the present study have demonstrated that ethanol and boiling water extract of the coffee beans, which determine phytochemical compounds of the extract and the level of chlorogenic acid and caffeine, cytotoxic apoptotic response was not dependent on the estrogen receptor, as similar results were obtained in cell lines possessing (MCF-7) and lacking (MDA-MB-231) this receptor and arrested the growth of proliferating cells from two breast cancer cell lines, which was associated with the induction of apoptosis as measured by cell cycle profiling. We recently reported that this extract contains high levels of phenolic compounds, including phenolic acids such as chlorogenic acid, as well as alkaloids such as caffeine. Many of these are known to inhibit the growth of breast cancer cells.50 Chlorogenic acid (CA) has been found to exhibit significant cytotoxic action against MCF-7 cells, with an inhibitory concentration (IC50) of 127 µM. When CA interacts with cisplatin, an anticancer agent, it enhances the cytotoxic activity by 2.5-fold compared to cisplatin alone.22 CA triggers the production of reactive oxygen species (ROS), which reduce mitochondrial membrane potential (MMP), activate the caspase-dependent apoptotic pathway, and arrest the cell cycle.51 Additionally, CA inhibits cell migration by decreasing N-cadherin levels and increasing C-cadherin levels. Studies have also shown that CA-mediated translocation of protein kinase C (PKC) in the tested MDA-MB-231 and MCF-7 cells alters the cell cycle, resulting in decreased cellular viability (IC50 values of 75.88 µg/mL and 52.5 µg/mL, respectively).11
Researchers found that CA inhibited the growth of MCF-7 cells (IC50 = 350 µM) and caused cell death in a concentration-dependent manner. When combined with hesperidin (100 µM), CA (350 µM) significantly changed the appearance of MCF-7 cells and resulted in 68% cell death. Additionally, CA and hesperidin reduced the expression of various genes, including transcription factor A, mitochondrial and nuclear respiratory factor 1, mitochondrial DNA, and ATP-synthesis-related genes ATP5PB and mtATP6.16,17 HPLC analysis revealed high chlorogenic acid in green coffee extracts, which decreased in roasted coffee as CF levels increased. Coffee extracts, especially those rich in caffeine, show potential anticancer effects against TNBC by inhibiting growth, inducing apoptosis, and suppressing metastasis through the modulation of cell signaling, cell cycle regulation, and programmed cell death.52 A recent HPLC/DAD analysis of five commercial coffee samples reported an average concentration of 8.35 ± 6.13 mg of CF per gram of methanol extract.53 Green coffee beans contained 0.97 ± 0.09 mg/g CA and 3.13 ± 0.33 mg/g CF. Roasted beans had 1.30 ± 0.13 mg/g CA and 1.00 ± 0.02 mg/g CF. Roasting reduced CA and phytochemicals, but caffeine increased.54,55 During roasting, CA degrades thermally, stabilizing alkaloids at high temperatures. As a result, CF and trigonelline become the primary metabolites in roasted coffee beans.56,57 The level of CA can indicate the quality of green coffee beans, helping to differentiate between low-quality varieties (9.1 g CA/100 g) and commercial ones (10.4 g/100 g).58 Dried green coffee beans contain carbohydrates (59–62%), chlorogenic acids (7–10%), aliphatic acids (2%), caffeine (1–2%), trigonelline (1%), and free amino acids (<1%). Roasting reduces carbs, CA, and amino acids but increases alkaloids (mainly caffeine) and aliphatic acids.59
Experiments evaluated the effects of coffee extracts on cancer cells. The MTT assay measured cytotoxicity in MDA-MB-231 and MCF-7 breast cancer cells, followed by analyzing MYC and AXIN2 gene expression. The more effective Arabica and Robusta extracts were tested for cytotoxicity in HEK293 normal kidney cells, and their mechanisms of apoptosis and cell cycle arrest were analyzed. Cytotoxicity was assessed after a 24-hour incubation with varying concentrations of the extract. Figure 4 shows a significant, concentration-dependent decrease in the viability of MDA-MB-231 and MCF-7 cells. Similar effects were observed in colon (CHT116), brain (T98G), and bone (U2OS) cancer cell lines.
Data indicate that green and roasted coffee extracts, chlorogenic acid, and caffeine inhibit the viability of breast cancer cell lines. Ayelén D. Nigra’s research on Brazilian coffee showed IC50s greater than 500 µg/mL for both green and dark coffee. Cell viability in the non-tumor MCF 10A line was unaffected except at high concentrations 1000 μg/mL).33 Different coffee constituents may have varying effects on ER+ and ER- breast cancer subtypes. Research has shown that caffeine doses significantly inhibit the proliferation and overall cell count of MDA-MB-231 breast cancer cells, with the greatest inhibition observed at 5 mmol/L.19 In contrast, chlorogenic acid is highly cytotoxic to MCF-7 cells. Coffee extracts can suppress the growth, migration, and invasion of TNBC cells, likely by inhibiting enzymes such as MMP-3, which is essential for invasion. They also modulate signaling pathways, such as the AKT and β-catenin pathways, which are involved in cell growth and metastasis.60,61 Green coffee samples, selected by qRT-PCR, exhibited low cytotoxicity in normal kidney cells (HEK293) at concentrations exceeding 2000 µg/mL (Figure 6). Roasted coffee contains fewer bioactive compounds due to high-temperature processing. We assessed cell viability after treatment with roasted coffee extracts, which are more commonly consumed. Both coffee types showed antitumor effects on MDA-MB-231 and MCF-7 breast cancer cells; green coffee extract was less cytotoxic to normal cells than roasted coffee. Thus, green coffee extract was selected for further analysis.
Further experiments investigated whether the cytotoxicity of coffee bean extracts on MDA-MB-231 and MCF-7 cells was associated with apoptosis and cell cycle activity. Arabica and Robusta extracts were evaluated using flow cytometry (Figures 7A, 7B, 8A and 8B), following analysis of MYC and AXIN2 gene expression via qRT-PCR (Figure 5). The MYC gene encodes a transcription factor that regulates cell growth, proliferation, differentiation, and apoptosis.62 The AXIN2 gene plays a crucial role in the Wnt signaling pathway, which is important for cell growth and differentiation.63 Analysis of MYC and AXIN2 expression in coffee extracts by qRT-PCR reveals that MYC expression in the treated sample is significantly lower than in the control (Figure 5), indicating downregulation. Since MYC is often overexpressed in cancers, reduced levels suggest potential anti-cancer effects by decreasing cell proliferation, which benefits conditions with uncontrolled growth. The AXIN2 expression is also lower in the treated sample, indicating downregulation that may affect the Wnt pathway and influence cell proliferation, differentiation, and apoptosis. Numerous studies have highlighted the potential anti-cancer properties of coffee extracts and compounds, including chlorogenic acid and caffeine, with various mechanisms underlying their effects.64,65 Our findings on the cytotoxic activity of coffee extracts against ER-negative breast cancer cells, along with the underlying molecular mechanisms, suggest that coffee may have potential as an anticancer agent. Efforts to identify specific target pathways showed that cancer cell characteristics influence how anticancer agents interact. ER-negative cells exposed to coffee exhibit greater sensitivity, primarily via the Wnt signaling pathway. Green coffee extracts affected the MYC and AXIN2 genes more than roasted coffee, chlorogenic acid, or caffeine alone, indicating a more efficient interaction with cancer cells and a lesser impact on normal cells. These hold promise for overcoming a lack of specificity, but further research is needed to confirm the therapeutic potential of coffee extracts.
To further elucidate the molecular basis underlying the observed downregulation of MYC and AXIN2, molecular docking analysis was performed targeting Tankyrase-1, a key regulator of the Wnt/β-catenin pathway. Docking results demonstrated that chlorogenic acid exhibited favorable binding affinity toward Tankyrase-1, forming multiple hydrogen bonds and hydrophobic interactions within the catalytic pocket. In contrast, binding affinity toward Estrogen Receptor Alpha was comparatively weaker, suggesting that the cytotoxic response observed in both ER-positive and ER-negative cells was not primarily mediated through estrogen receptor inhibition. Since Tankyrase-1 regulates AXIN stability and β-catenin signaling, its inhibition may suppress Wnt-driven transcription factors such as MYC. This finding is consistent with the qRT-PCR data showing significant downregulation of MYC and AXIN2 expression following treatment. The stronger docking interaction with Tankyrase-1 provides a mechanistic explanation for the higher sensitivity observed in MDA-MB-231 cells. Molecular dynamics simulation further confirmed that chlorogenic acid maintained structural stability within the Tankyrase-1 binding pocket throughout the 200 ns trajectory, as indicated by stable RMSD and controlled RMSF fluctuations. In comparison, the ERα complex displayed structural stability but showed weaker thermodynamic binding strength in MM-PBSA analysis. The binding free energy decomposition revealed that the Tankyrase-1 complex possessed a more favorable total binding energy, dominated by van der Waals and electrostatic contributions. Although electrostatic interactions were also present in the ERα complex, the higher polar solvation penalty reduced its overall binding stability. These computational findings align with the apoptosis and cell cycle data, suggesting that chlorogenic acid exerts stronger functional modulation through Wnt pathway regulation rather than direct estrogen receptor antagonism. The integration of docking, molecular dynamics, and MM-PBSA analyses, therefore, provides molecular-level support for the experimentally observed anticancer effects of green coffee extracts.
The results of this study demonstrate that a coffee bean extract inhibits the growth of proliferating cells from two breast cancer cell lines, an effect associated with apoptosis induction, as shown by cell cycle analysis. This cytotoxic apoptotic response was not dependent on the estrogen receptor, as similar results were observed in cell lines with and without the MCF-7 receptor, but not in the MDA-MB-231 cell line.33 To determine whether the growth inhibitory effects of coffee bean extract, chlorogenic acid, and caffeine were caused by specific disruptions of cell cycle-related events, the DNA contents of both MCF-7 and MDA-MB-231 cells were measured using flow cytometric analysis. In MCF-7 cells, this analysis showed that both Arabica and Robusta green coffee bean extracts caused an accumulation of cells in the G1 phase, similar to caffeine effects, and in the S-phase, identical to chlorogenic acid effects, thus inhibiting the transition of cells into the G2/M phase with increasing concentrations from 100 µg/mL to 200 µg/mL. In MDA-MB-231 cells, both extracts, along with chlorogenic acid and caffeine, initially caused cell cycle arrest at G1 and G2/M phases. However, at the higher concentration of 200 µg/mL, specifically for Arabica green coffee bean extract and caffeine, the percentage of cells in the G1 phase decreased. In contrast, those in G2/M and S phases increased. Conversely, with Robusta green coffee bean extract and chlorogenic acid, the percentages of cells in G1 and G2/M phases decreased, while the proportion in the S phase increased. The results of the cell cycle analysis indicate that treating cells with green coffee extract at 1000 μg/mL increased the S-phase and decreased the G2/M population in both breast cancer cell lines (MDA-MB-231 and MCF7). In contrast, this was not observed in the cell cycle population distribution of MCF-10A, a standard breast cell line.66
To determine whether the cytotoxic effects of coffee bean extracts were related to apoptosis, annexin V-FITC/PI double staining was used to quantify apoptotic cells by flow cytometry. Phosphatidylserine (PS) is a key biomarker of early apoptosis. It translocates from the cytosolic side of the membrane to the extracellular domain, which is detected by annexin V-FITC staining. As apoptosis progresses, the cell membrane is compromised, allowing PI to enter the nucleus and stain DNA. The MDA-MB-231 cell line showed greater sensitivity than the MCF-7 cell line to apoptosis induced by coffee bean extracts at concentrations of 100 µg/mL, particularly in early apoptosis. When the concentration was increased to 200 µg/mL, MCF-7 cells exhibited higher sensitivity than MDA-MB-231 cells, especially in late apoptosis.
Our current findings on the cytotoxic activity of West Java coffee bean extracts against breast cancer cells, along with molecular mechanism profiles, provide additional evidence of their anticancer potential. This new evidence was obtained through our efforts to identify more specific target cells for coffee bean extracts, caffeine, and chlorogenic acid. The characteristics of cancer cells can influence how selectively an anticancer agent interacts with them. When breast cancer cells exhibit more sensitive and specific responses to coffee bean extract, it becomes more likely that the extract will interact more effectively with target cell pathways, such as the Wnt signaling pathway, and less so with normal cells. This discovery is expected to reduce the limitations of coffee bean extracts related to non-specificity. However, the data obtained remain incomplete, and much work remains to be done to demonstrate the therapeutic relevance of coffee bean extracts.
Conclusions
The results of this study indicate that both roasting processes and extraction methods significantly influence the phytochemical composition of coffee beans, particularly chlorogenic acid content, which decreases at higher temperatures. Green coffee extracts, especially those obtained through maceration, demonstrated cytotoxic activity against MDA-MB-231 and MCF-7 breast cancer cell lines while exhibiting relatively low toxicity toward normal HEK293 cells. Gene expression analysis revealed significant downregulation of MYC and reduced AXIN2 expression, suggesting modulation of the Wnt signaling pathway. Flow cytometry further confirmed increased late apoptosis and cell-cycle arrest, supporting the extracts’ antiproliferative and pro-apoptotic activities. Molecular docking analysis indicated that chlorogenic acid interacts favorably with Tankyrase-1, a key regulator of the Wnt/β-catenin pathway, while exhibiting comparatively weaker binding toward Estrogen Receptor Alpha. Molecular dynamics simulations demonstrated stable ligand accommodation within the Tankyrase-1 binding pocket over 200 ns, supported by controlled structural fluctuations. MM-PBSA energy calculations further revealed a more favorable binding free energy for the Tankyrase-1 complex compared to ERα. These computational findings provide mechanistic support for the observed downregulation of Wnt-related genes and induction of apoptosis. Collectively, the integration of in vitro and in silico analyses suggests that green coffee extracts, particularly chlorogenic acid–rich fractions, may exert anticancer effects predominantly through modulation of Wnt signaling rather than estrogen receptor–dependent pathways. Although promising, further mechanistic validation and in vivo studies are required to confirm their therapeutic relevance and potential application as complementary or preventive anticancer agents.
Acknowledgment
The authors would like to thank the Pharmaceutical Biology Laboratory, the Translational Pharmaceutical Research Laboratory, and the Central Laboratory of Universitas Padjadjaran, the places where the research was conducted. The authors also thank Universitas Padjadjaran for supporting this research and the APC for publication, the research facility, and funding.
Funding
The authors would like to express their sincere gratitude to Rector of Universitas Padjadjaran, for providing valuable support and facilities that contributed to the successful completion of this research. This publication charge is funded by Universitas Padjadjaran through the Indonesian Endowment Fund for Education (LPDP) on behalf of the Indonesian Ministry of Higher Education, Science and Technology and managed under the EQUITY Program (Contract No. 4303/B3/DT.03.08/2025 and 3927/UN6.RKT/HK.07.00/2025).
Disclosure
The authors report no conflicts of interest in this work.
References
1. Schneider KA, Chittenden A, Shannon KM. Counseling About Cancer: Strategies for Genetic Counseling. John Wiley & Sons; 2023.
2. Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–25. doi:10.3322/caac.21492
3. Herawati IE, Lesmana R, Levita J, Cytotoxicity SA. Apoptosis, migration inhibition, and autophagy-induced by crude ricin from ricinus communis seeds in A549 lung cancer cell lines. Med Sci Monit Basic Res. 2022;28:e936683. doi:10.12659/MSMBR.936683
4. Xu X, Zhang M, Xu F, Jiang S. Wnt signaling in breast cancer: biological mechanisms, challenges and opportunities. Mol Cancer. 2020;19(1):1–35. doi:10.1186/s12943-020-01276-5
5. Serafino A, Sferrazza G, Colini Baldeschi A, et al. Developing drugs that target the Wnt pathway: recent approaches in cancer and neurodegenerative diseases. Expert Opin Drug Discov. 2017;12(2):169–186. doi:10.1080/17460441.2017.1271321
6. Yulia M, Ningtyas KR, Suharyatun S, Rahmawati W, Suhandy D. UV spectroscopy for discrimination of two arabica coffee cultivars in West Java Indonesia: a feasibility study. IOP Conf Ser Earth Environ Sci. 2022;1027(1):012015. doi:10.1088/1755-1315/1027/1/012015
7. Figueiredo LP, Borém FM, Almeida MR, et al. Raman spectroscopy for the differentiation of Arabic coffee genotypes. Food Chem. 2019;288:262–267. doi:10.1016/j.foodchem.2019.02.093
8. Rico, Darma R, Salman D, Mahyuddin. Problems identification of Arabica coffee commodities on traditional farming in Indonesia: a review. IOP Conf Ser Earth Environ Sci. 2021;886(1):012069. doi:10.1088/1755-1315/886/1/012069
9. Merikhian P, Eisavand MR, Farahmand L. Triple-negative breast cancer: understanding Wnt signaling in drug resistance. Cancer Cell Int. 2021;21(1):1–8. doi:10.1186/s12935-021-02107-3
10. Sferrazza G, Corti M, Brusotti G, et al. Nature-derived compounds modulating Wnt/β-catenin pathway: a preventive and therapeutic opportunity in neoplastic diseases. Acta Pharm Sin B. 2020;10(10):1814–1834. doi:10.1016/j.apsb.2019.12.019
11. Lu H, Tian Z, Cui Y, Liu Z, Ma X. Chlorogenic acid: a comprehensive review of the dietary sources, processing effects, bioavailability, beneficial properties, mechanisms of action, and future directions. Compr Rev Food Sci Food Saf. 2020;19(6):3130–3158. doi:10.1111/1541-4337.12620
12. Vasan N, Baselga J, Hyman DM. A view on drug resistance in cancer. Nature. 2019;575:7782):299–309. doi:10.1038/s41586-019-1730-1
13. Bai X, Ni J, Beretov J, Graham P, Li Y. Cancer stem cell in breast cancer therapeutic resistance. Cancer Treat Rev. 2018;69:152–163. doi:10.1016/j.ctrv.2018.07.004
14. Science E. Phytochemical screening and antioxidant test of Arabika roasted coffee bean extract (Coffea arabica L.) from agam regency phytochemical screening and antioxidant test of Arabika roasted. In
15. Cory H, Passarelli S, Szeto J, Tamez M, Mattei J. The role of polyphenols in human health and food systems: a mini-review. Front Nutr. 2018;5:370438. doi:10.3389/fnut.2018.00087
16. Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73(1):17–48. doi:10.3322/caac.21763
17. Amano Y, Honda H, Nukada Y, et al. Safety pharmacological evaluation of the coffee component, caffeoylquinic acid, and its metabolites, using ex vivo and in vitro profiling assays. Pharmaceuticals. 2019;12(3):110. doi:10.3390/ph12030110
18. Unis A, Abdelbary A. Green coffee bean extract attenuates gentamicin induced acute nephrotoxicity in rats. Bol Latinoam y del Caribe Plantas Med y Aromat. 2022;21(2):256–267. doi:10.37360/blacpma.22.21.2.16
19. Rosendahl AH, Perks CM, Zeng L, et al. Caffeine and caffeic acid inhibit growth and modify estrogen receptor and insulin-like growth factor i receptor levels in human breast cancer. Clin Cancer Res. 2015;21(8):1877–1887. doi:10.1158/1078-0432.CCR-14-1748
20. Zeng L, Holly JMP, Perks CM. Effects of physiological levels of the green tea extract epigallocatechin-3-gallate on breast cancer cells. Front Endocrinol. 2014;5(MAY):1–10. doi:10.3389/fendo.2014.00061
21. Xu R, Kang Q, Ren J, Li Z, Xu X. Antitumor molecular mechanism of chlorogenic acid on inducting genes GSK-3 β and APC and inhibiting gene β -Catenin. J Anal Methods Chem. 2013;2013:1–7. doi:10.1155/2013/951319
22. Gupta A, Atanasov AG, Li Y, Kumar N, Bishayee A. Chlorogenic acid for cancer prevention and therapy: current status on efficacy and mechanisms of action. Pharmacol Res. 2022;186:106505. doi:10.1016/j.phrs.2022.106505
23. Zhang -J-J, Xu Q-J, Schmidt C, et al. Elucidating the multimodal anticancer mechanism of an organometallic Terpyridine Platinum(II) N-Heterocyclic carbene complex against triple-negative breast cancer in vitro and in vivo. J Med Chem. 2023;66(6):3995–4008. doi:10.1021/acs.jmedchem.2c01925
24. Amira QH, Bowolaksono A, Dwiranti A. Caffeine as a modulator of redox balance and migration in MDA-MB-231 triple-negative breast cancer cells. Curr Res Biosci Biotechnol. 2025;6(2):15–21.
25. Amira QH, Irdianto SA, Fadilah F, et al. Caffeine modulates cell death and telomerase activity in triple-negative breast cancer cells. Int J Mol Cell Med. 2025;14(2):606–619. doi:10.22088/IJMCM.BUMS.14.2.606
26. Kalın ŞN, Bayındırlı KN, Toraman E, et al. Lobaric acid exhibits anticancer potential by modulating the Wnt/β-catenin signaling pathway in MCF-7 cells. Pharmacol Res Perspect. 2025;13(4). doi:10.1002/prp2.70142
27. Lustig RH. Hypothalamic obesity: the sixth cranial endocrinopathy. Endocrinologist. 2002;12(3):210–217. doi:10.1097/00019616-200205000-00008
28. Villota H, Santa-González GA, Uribe D, et al. Modulatory effect of chlorogenic acid and coffee extracts on Wnt/β-Catenin pathway in colorectal cancer cells. Nutrients. 2022;14(22):1–19. doi:10.3390/nu14224880
29. Parnomo T. Effect of Arabica coffee bean extract (Coffea arabica) as a growth inhibitor of Enterococcus faecalis ATCC 29212. J Drug Delivery Ther. 2021;11(3):89–96. doi:10.22270/jddt.v11i3.4820
30. Hartanti EA, Arif M. Copyright © 2023 Universitas Pakuan. Stand Semanggi (Marsilea crenata C Presl) Leaves from Benowo Dist Surabaya Stand Herb Raw Mater Burhan. FITOFARMAKA J Ilmiah Farmasi. 2023;13(1):70–76.
31. Praherdiono H, Adi E. Panduan Praktikum Multimedia. 2008;4.
32. AAl SA. Determination of caffeine and chlorogenic acid (CGA) in the methanolic extracts coffee (C. arabica. L) to seeds and peels (Unroasted and Roasted) cultivars grown in yemen by high performance liquid chromatography (HPLC). Bioequiv Bioavailab Int J. 2023;7(1):1–19. doi:10.23880/beba-16000180
33. Nigra AD, De Almeida Bauer Guimarães D, Prucca CG, Freitas-Silva O, Teodoro AJ, Gil GA. Antitumor effects of freeze-dried robusta coffee (Coffea canephora) extracts on breast cancer cell lines. Oxid Med Cell Longev. 2021;2021. doi:10.1155/2021/5572630
34. Amalia R, Abdelaziz M, Puteri MU, et al. TMEPAI/PMEPA1 inhibits Wnt signaling by regulating β-catenin stability and nuclear accumulation in triple negative breast cancer cells. Cell Signal. 2019;59:24–33. doi:10.1016/j.cellsig.2019.03.016
35. Bauer D, Abreu J, Jordão N, da Rosa JS, Freitas-Silva O, Teodoro A. Effect of roasting levels and drying process of coffea canephora on the quality of bioactive compounds and cytotoxicity. Int J Mol Sci. 2018;19(11):3407. doi:10.3390/ijms19113407
36. Liu G, Chu H. Andrographolide inhibits proliferation and induces cell cycle arrest and apoptosis in human melanoma cells. Oncol Lett. 2018;15(4):5301–5305. doi:10.3892/ol.2018.7941
37. Berman HM, Westbrook J, Feng Z, et al. The Protein Data Bank. Nucleic Acids Res. 2000;28:235–242. doi:10.1093/nar/28.1.235
38. Forli W, Halliday S, Belew R, Olson A. AutoDock Version 4.2. Citeseer. 2012.
39. Kim S, Chen J, Cheng T, et al. PubChem 2025 update. Nucleic Acids Res. 2025;53(D1):D1516–D1525. doi:10.1093/nar/gkae1059
40. Brown T. ChemDraw. Sci Teach. 2014;81(2):1.
41. Eberhardt J, Santos-Martins D, Tillack AF, Forli S. AutoDock Vina 1.2.0: new docking methods, expanded force field, and python bindings. J Chem Inf Model. 2021;61(8):3891–3898. doi:10.1021/acs.jcim.1c00203
42. BIOVIA. Dassault Systèmes BIOVIA, Discovery Studio Modeling Environment, Release 2017. In: Dassault Systèmes. San Diego(Calif); 2017.
43. Pronk S, Páll S, Schulz R, et al. GROMACS 4.5: a high-throughput and highly parallel open source molecular simulation toolkit. Bioinformatics. 2013;29(7):845–854. doi:10.1093/bioinformatics/btt055
44. Abraham MJ, Murtola T, Schulz R, et al. GROMACS: high performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX. 2015;1–2:19–25. doi:10.1016/j.softx.2015.06.001
45. Kumari R, Kumar R, Consortium OSDD, Lynn A. g_mmpbsa —A GROMACS tool for high-throughput MM-PBSA calculations. J Chem Inf Model. 2014;54:1951–1962. doi:10.1021/ci500020m
46. Wang C, Greene D, Xiao L, Qi R, Luo R. Recent developments and applications of the MMPBSA method. Front Mol Biosci. 2018;4. doi:10.3389/fmolb.2017.00087.
47. Korkmaz ED, Temizci B. Coffee bean-derived extracellular vesicles exhibit anti-cancer effects on neuroblastoma cells. J High Sch Sci. 2025;9(3):306–322.
48. Ahmad R, Ahmad N, Al-Anaki WS, Ismail FA, Al-Jishi F. Solvent and temperature effect of accelerated solvent extraction (ASE) coupled with ultra-high-pressure liquid chromatography (UHPLC-PDA) for the determination of methyl xanthines in commercial tea and coffee. Food Chem. 2020;311:126021. doi:10.1016/j.foodchem.2019.126021
49. Montenegro J, Dos Santos LS, de Souza RGG, et al. Bioactive compounds, antioxidant activity and antiproliferative effects in prostate cancer cells of green and roasted coffee extracts obtained by microwave-assisted extraction (MAE). Food Res Int. 2021;140:110014. doi:10.1016/j.foodres.2020.110014
50. Shah S, Kumar A. Coffee. Const Health Benefits. 2020;7051(5):22–38. doi:10.9734/BJI/2020/v24i530115
51. Abrankó L, Clifford MN. An unambiguous nomenclature for the acyl-quinic acids commonly known as chlorogenic acids. J Agric Food Chem. 2017;65(18):3602–3608. doi:10.1021/acs.jafc.7b00729
52. Pascua SM, McGahey GE, Ma N, Wang JJ, Digman MA. Caffeine and cisplatin effectively targets the metabolism of a triple-negative breast cancer cell line assessed via phasor-FLIM. Int J Mol Sci. 2020;21(7):2443. doi:10.3390/ijms21072443
53. Córdoba N, Moreno FL, Osorio C, Velásquez S, Ruiz Y. Chemical and sensory evaluation of cold brew coffees using different roasting profiles and brewing methods. Food Res Int. 2021;141:110141. doi:10.1016/j.foodres.2021.110141
54. Martínez-López S, Sarriá B, Baeza G, Mateos R, Bravo-Clemente L. Pharmacokinetics of caffeine and its metabolites in plasma and urine after consuming a soluble green/roasted coffee blend by healthy subjects. Food Res Int. 2014;64:125–133. doi:10.1016/j.foodres.2014.05.043
55. Gómez-Juaristi M, Martínez-López S, Sarria B, Bravo L, Mateos R. Bioavailability of hydroxycinnamates in an instant green/roasted coffee blend in humans. Identification of novel colonic metabolites. Food Funct. 2018;9(1):331–343. doi:10.1039/C7FO01553D
56. Stegmüller S, Beißmann N, Kremer JI, Mehl D, Baumann C, Richling E. A New UPLC-qTOF approach for elucidating furan and 2-methylfuran metabolites in human urine samples after coffee consumption. Molecules. 2020;25(21):5104. doi:10.3390/molecules25215104
57. Acidri R, Sawai Y, Sugimoto Y, et al. Phytochemical profile and antioxidant capacity of coffee plant organs compared to green and roasted coffee beans. Antioxidants. 2020;9(2):93. doi:10.3390/antiox9020093
58. Angeloni G, Guerrini L, Masella P, Innocenti M, Bellumori M, Parenti A. Characterization and comparison of cold brew and cold drip coffee extraction methods. J Sci Food Agric. 2019;99(1):391–399. doi:10.1002/jsfa.9200
59. Bøhn SK, Blomhoff R, Paur I. Coffee and cancer risk, epidemiological evidence, and molecular mechanisms. Mol Nutr Food Res. 2014;58(5):915–930. doi:10.1002/mnfr.201300526
60. Farghadani R, Naidu R. The anticancer mechanism of action of selected polyphenols in triple-negative breast cancer (TNBC). Biomed Pharmacother. 2023;165:115170. doi:10.1016/j.biopha.2023.115170
61. Nwafor E, Lu P, Zhang Y, Liu R, Peng H, Xing B. Translational Oncology Chlorogenic acid: potential source of natural drugs for the therapeutics of fibrosis and cancer. Transl Oncol. 2022;15(1):101294. doi:10.1016/j.tranon.2021.101294
62. Madden SK, de Araujo AD, Gerhardt M, Fairlie DP, Mason JM. Taking the Myc out of cancer: toward therapeutic strategies to directly inhibit c-Myc. Mol Cancer. 2021;20(1):1–18. doi:10.1186/s12943-020-01291-6
63. Marraccini P. Gene expression in coffee. Progr Bot. 2020;(December 2020):43–111. doi:10.1007/124_2020_42
64. Giovannucci EL. Coffee and Solid Tumors. Nutraceut Hum Heal Food Supplement Paradig. 2020;23:105.
65. Ismail T, Donati-zeppa S, Akhtar S, Turrini E, Sestili P, Fimognari C. review AC. Expert Opin Drug Metab Toxicol. 2021;17:69–85. doi:10.1080/17425255.2021.1839412
66. Kanlaya R, Subkod C, Nanthawuttiphan S, Thongboonkerd V. Caffeine causes cell cycle arrest at G0/G1 and increases of ubiquitinated proteins, ATP and mitochondrial membrane potential in renal cells. Comput Struct Biotechnol J. 2023;21(September):4552–4566. doi:10.1016/j.csbj.2023.09.023
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Caffeic Acid Phenethyl Ester as a DHODH Inhibitor and Its Synergistic Anticancer Properties in Combination with 5-Fluorouracil in a Breast Cancer Cell Line
Amalia E, Diantini A, Endang Prabandari E, Waluyo D, Subarnas A
Journal of Experimental Pharmacology 2022, 14:243-253
Published Date: 23 July 2022
The Potent Novel CDK4/6 Inhibitor TQB3616 in Hormone Receptor Positive Breast Cancer: Preclinical Characterization with in vitro and Human Tumor Xenograft Models
Hu W, Wang L, Luo J, Zhang J, Li N
Breast Cancer: Targets and Therapy 2023, 15:899-912
Published Date: 8 December 2023
Exploring the Potential of Chaihu-Danggui Tang in Breast Cancer Treatment Based on Network Pharmacology, Molecular Docking, and Experimental Validation

Liu Y, Zhang J, Lai Y, Wu C, Liu D, Liang R, Chen G, Jiang X
Breast Cancer: Targets and Therapy 2025, 17:385-401
Published Date: 10 May 2025
Quercetin: A Natural Ally in Combating Breast Cancer
Wu ZY, Qiu KY, Gai YJ, Wu JH, Zhou BX, Shi QF
International Journal of Nanomedicine 2025, 20:9155-9177
Published Date: 19 July 2025
Novel Insights into the Oncogenic Role and Clinical Significance of ORC6L in Breast Cancer
Jiang X, Wang S, Wei Q, Yang Y, Chen Y, Chen S
Breast Cancer: Targets and Therapy 2026, 18:586715
Published Date: 20 April 2026