Back to Journals » Infection and Drug Resistance » Volume 16
Carbapenem-Resistant and ESBL-Producing Enterobacterales Emerging in Central Texas
Authors Parker JK ![]() , Gu R, Estrera GA, Kirkpatrick B, Rose DT, Mavridou DAI
, Gu R, Estrera GA, Kirkpatrick B, Rose DT, Mavridou DAI ![]() , Mondy KE, Davies BW
, Mondy KE, Davies BW ![]()
Received 3 January 2023
Accepted for publication 24 February 2023
Published 2 March 2023 Volume 2023:16 Pages 1249—1261
DOI https://doi.org/10.2147/IDR.S403448
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 5
Editor who approved publication: Prof. Dr. Héctor Mora-Montes
Jennifer K Parker,1 Richard Gu,1 Gregory A Estrera,1 Betsy Kirkpatrick,2 Dusten T Rose,3 Despoina AI Mavridou,1,4 Kristin E Mondy,5 Bryan W Davies1,4
1Department of Molecular Biosciences, The University of Texas at Austin, Austin, TX, USA; 2Austin Public Health, City of Austin, Austin, TX, USA; 3Department of Pharmacy, Ascension Seton, Dell Seton Medical Center at The University of Texas, Austin, TX, USA; 4John Ring LaMontagne Center for Infectious Diseases, The University of Texas at Austin, Austin, TX, USA; 5Department of Internal Medicine, Dell Medical School, The University of Texas at Austin, Austin, TX, USA
Correspondence: Bryan W Davies, Email [email protected]
Purpose: Carbapenem-resistant Enterobacterales (CRE) are subject to intense global monitoring in an attempt to maintain awareness of prevalent and emerging resistance mechanisms and to inform treatment and infection prevention strategies. CRE and extended-spectrum beta-lactamase (ESBL)-producing Enterobacterales are not usually examined collectively in regards to their shared pool of resistance determinants. Here, we genetically and phenotypically assess clinical isolates of CRE and extended-spectrum beta-lactamase (ESBL)-producing Enterobacterales in the growing region of Central Texas, where CRE are emergent and occurrence of non-carbapenemase-producing-CRE (non-CP-CRE) infections is increasing.
Methods: CRE (n=16) and ESBL-producing Enterobacterales (n=116) isolates were acquired from a regional hospital in Central Texas between December 2018 and January 2020. Isolates were assessed genetically and phenotypically using antibiotic susceptibility testing, targeted PCR, and whole genome sequencing.
Results: CRE infections are increasing in incidence in Central Texas, and Klebsiella pneumoniae is causing the majority of these infections. Moreover, K. pneumoniae sequence type (ST) 307 is commonly found among both non-CP-CRE and EBSL-producing strains. Isolates carry similar plasmids harboring the gene for the ESBL CTX-M-15 and belong to the global lineage, rather than the Texas lineage, of ST307. Antibiotic resistance profiles, sequence data, and clinical records suggest that porin mutations may promote the transition of ST307 isolates from ESBL-producing to non-CP-CRE. In addition to antibiotic resistance mechanisms, several CRE isolates harbor active colicinogenic plasmids, which might influence the competitiveness of these bacteria during patient colonization.
Conclusion: K. pneumoniae of the global ST307 lineage is circulating in Central Texas and is responsible for both non-CP CRE and ESBL-producing Enterobacterales infections. Enhanced surveillance is needed to understand the possible routes for the emergence of non-CP-CRE from EBSL-producing strains.
Keywords: beta-lactam, Klebsiella, colicin, genetics, carbapenemase
Introduction
With the prevalence of carbapenem-resistant Enterobacterales (CRE) increasing worldwide,1,2 long-term management of these challenging resistant strains would benefit from a better understanding of their geographical distribution and spread. It is important to differentiate carbapenemase-producing CRE (CP-CRE) from non-carbapenemase-producing CRE (non-CP-CRE). Acquisition of a single carbapenemase is sufficient for a strain to become CP-CRE, while the resistance mechanism(s) of non-CP-CRE are varied and often difficult to identify.3,4 Alarmingly, of all CRE isolates cataloged by the Centers for Disease Control and Prevention (CDC) Antibiotic Resistance Laboratory Network (ARLN) in the United States in 2017, 68% were non-CP-CRE.5 The most common CRE species, in order, are Klebsiella pneumoniae, Enterobacter cloacae species complex, Serratia marcescens, Escherichia coli, and Klebsiella aerogenes.2
In addition to CRE, another highly prevalent group of beta-lactam-resistant Enterobacterales that causes significant clinical burden, despite remaining susceptible to carbapenems, are the extended-spectrum beta-lactamase (ESBL)-producing Enterobacterales.2,6 Although CRE and ESBL-producing strains often share similar genetic backgrounds, carry some of the same antibiotic-resistance determinants, and endure the same antibiotic pressures from treatment with beta-lactams, they are not always considered collectively when examining the epidemiology of multidrug-resistant Enterobacterales. Further elucidation of the potential relationships between the resistance profiles of CRE and ESBL-producing strains and of the contribution of these relationships to the dissemination of resistance is necessary.7
To increase our understanding of infections caused by CRE and ESBL-producing Enterobacterales, we examined the genotypic, phenotypic, and clinical features of CRE (n=16) and ESBL-producing strains (n=116) from Central Texas from December 2018 to January 2020; this strain collection included both CP-CRE and non-CP-CRE isolates. Since Central Texas is a growing area of the United States, CRE are emergent here, and isolates from this region have not previously been analyzed in depth. The purpose of this study is to provide the first detailed characterization of CRE isolates from Central Texas, contextualized with analysis of ESBL-producing strains isolated from the same population during the same time period. Incidentally, plasmids with colicin determinants were discovered in some CRE isolates during this process; therefore, we also sought to provide a first characterization of colicin production by CRE isolates in vitro.
Materials and Methods
Bacterial Isolate Collection and Initial Screening
Bacterial isolates cultured as part of routine clinical diagnostics were acquired from a regional hospital in Central Texas. Isolates were obtained from patients both within the hospital and from other facilities (eg long-term care facilities) within the same region (from which isolates/samples were submitted to the hospital for testing) from December 2018 to January 2020. Species identification and antibiotic susceptibility testing were conducted on the VITEK 2 system using the GN ID card and the AST-GN69 and AST-XN06 susceptibility cards, respectively (BioMérieux). All isolates designated “ESBL” by VITEK analysis (n=116) or meeting the CDC definition of CRE (n=16) were included in this study. CRE is defined by the CDC as “Enterobacterales that test resistant to at least one of the carbapenem antibiotics”.8 CRE isolates (Klebsiella spp. and E. coli only) were submitted to the Texas Department of State Health Services (DSHS) for additional analyses per CDC recommendations.9 At DSHS they were tested for carbapenemase production (mCIM method).10 MICs for select aminoglycosides, monobactams, carbapenems, cephalosporins, fluoroquinolones, tigecycline, polymyxins, piperacillin/tazobactam, and trimethoprim/sulfamethoxazole were also determined by broth microdilution (BMD). PCRs for important carbapenemase genes (blaKPC, blaOXA-48, blaNDM, blaIMP, and blaVIM) and colistin-resistance genes (mcr-1 and mcr-2) were also performed by DSHS. For a few of the CRE isolates, either the VITEK or the DSHS report was unavailable.
Local CRE Incidence
Data on incidence and resistance mechanisms of local CRE cases since they were first recorded in the area were compiled. All CRE cases in Travis County, Texas are reported to Austin Public Health, and reporting facilities are encouraged to submit these isolates to DSHS labs for the additional analyses detailed above. Case counts, by carbapenemase presence/type when available, were compiled for the years 2014–2020.
Isolate Clinical Data
Patient clinical data collected included age, gender, dates of admission and discharge, and treatment outcome. Date of isolate collection, sample source, initial antibiotic treatment and susceptibility were also recorded, along with any other relevant details (eg, previous history of infection). Isolates were identified as causing either a healthcare-associated infection (HAI) or a community-acquired infection (CAI) based on National Healthcare Safety Network (NHSN) definitions, which generally designate an isolate as HAI when acquired >48 hours after hospitalization.11
Whole Genome Sequencing and Analysis
Whole genome sequencing (WGS) by GENEWIZ was initially conducted on seven diverse (by species and resistance phenotype) CRE isolates. Subsequently, one additional CRE isolate and five additional ESBL isolates were sequenced for sequence type confirmation, as described in further detail in the results. Library preparation was performed using the NEBNext Ultra DNA Library Prep for Illumina (NEB). Pooled libraries were sequenced on a MiSeq System (Illumina) with 250 bp paired-end reads. Raw reads were trimmed to remove adapters, low quality ends (<Q30), and short reads (<30 bp) using BBDuk.12 Trimmed reads were assembled to appropriate reference chromosomes and plasmids (GenBank accessions CP010128, CP010140, CP011991, CP015504, CP018348, CP019002, CP031655, CP031756, CP034396, CP045782, CP048326, CP050378, CP052241, CP052243, CP052268, CP053035, CP068803, MH745929, MK862125, MN542378), and remaining unassembled reads were de novo assembled, all using Geneious Prime 2021.1.1 (https://www.geneious.com). Sequence types were determined by analysis of assembled contigs at PubMLST, for standard multilocus sequence typing (MLST) per species,13 and at BIGSdb-Pasteur (Institut Pasteur)14 using the K. pneumoniae 629-gene strict core genome MLST (scgMLST) scheme.15 Capsular type was also analyzed at BIGSdb-Pasteur based on the wzi gene sequence.16 Antibiotic resistance determinants were identified in all resulting contigs using the Resistance Gene Identifier (RGI) component of the Comprehensive Antibiotic Resistance Database (CARD).17 Plasmid replicons were identified using PlasmidFinder 2.0.1.18 Raw sequence reads from WGS are deposited in the Sequence Read Archive (SRA) of the National Center for Biotechnology Information (NCBI) (BioProject ID PRJNA777127). Complete colicin plasmid assemblies for MDR006_pCol006 and MDR119_pCol119 are also deposited at NCBI (accessions ON209486 and ON209487, respectively).
PCR Amplification of Carbapenemase, Beta-Lactamase, and gyrA Genes
CRE isolates lacking DSHS PCR data on the mechanism of resistance were analyzed for the presence of the carbapenemase genes blaKPC, blaOXA-48 and blaNDM by PCR. Subsets of CRE and ESBL-producing isolates (all available isolates of K. pneumoniae and rare species and a convenience sample of E. coli) were also analyzed by PCR for the non-carbapenemase beta-lactamase genes blaSHV, blaTEM, blaOXA-1-like, and blaCTX-M-1-like. The quinolone resistance-determining region (QRDR) of gyrA from all available K. pneumoniae isolates was PCR-amplified and Sanger sequenced. All primers used here were previously published19–21 (Supplemental Table 1).
Assessment of Colicin Activity
Based on WGS results, two CRE isolates for which colicin plasmids were detected were assessed for in vitro colicin production. Clinical strains (MDR_006 and MDR_119) and E. coli K-12 strains from the Keio collection22 were grown overnight in LB medium at 37°C. Competitions were carried out on 0.8% w/v LB agar medium, while 1.5% w/v LB agar medium was used for determination of colicin toxicity from spent culture supernatants. When needed, solid or liquid media were supplemented with mitomycin C (at a final concentration of 0.1 μg mL−1, unless otherwise indicated). For the competition experiments, cells were harvested from overnight cultures, washed twice with LB medium, and resuspended and normalized for cell density before spotting. Competition plates were incubated statically at 37°C for 16 hours. For the growth inhibition experiment, strains were grown overnight in the presence of mitomycin C. Cell suspensions were normalized and supernatants were then collected, filtered and placed in 10 mm wells carved on plates with lawns of the indicated bacteria on their surface. After 16 hours of incubation, the zone of growth inhibition around the wells was measured at four points to compare the toxicities of the culture supernatants. Supernatants grown under the indicated conditions were precipitated with trichloroacetic acid (TCA) and separated by SDS-PAGE.
 |
Figure 1 CRE cases per year in Travis County, Texas. Colors indicate, when known, the beta-lactamases responsible for resistance to carbapenems (for CP-CRE) or the lack thereof (for non-CP-CRE). |
Results and Discussion
Local CRE Trends
Annual CRE cases in Travis County increased 850% from four in 2014 to 38 in 2020 (Figure 1). Significant numbers of both CP-CRE and non-CP-CRE are reported. Notably, incidence of non-CP-CRE cases has been increasing steadily since 2017. Most recently (2020), for cases where isolates were screened for the presence of carbapenemases, non-CP-CRE accounted for 54% of the tested strains. In this study period (2014–2020), the predominant species of CRE in Travis County was K. pneumoniae, accounting for 51% of isolates (data not shown). For comparison, recent surveillance (2015–2020) of CRE in the Greater Houston region of Texas also found a large increase (216%) in CRE incidence during this timeframe (from 374 isolates in 2015 to 1181 isolates in 2018), although incidence has decreased since 2018.23 The Greater Houston region’s dominant CRE species was also K. pneumoniae (84.8% of isolates). Where the carbapenemase status was known, 21% of CRE isolates from the Greater Houston region were non-CP-CRE.
 |
Figure 2 CRE and ESBL-producing Enterobacterales isolates collected during the study period. Isolate resistance phenotypes are indicated by species. Whole genome sequencing (WGS) was initially conducted for seven diverse isolates, as indicated here. Subsequent WGS was performed for six additional K. pneumoniae isolates to confirm their suspected sequence type. Figure created with BioRender.com. |
Isolate Overview and Antibiotic Resistance
132 clinical isolates from the order Enterobacterales with either a CRE or an ESBL phenotype were included in this study (Figure 2). All species studied belong to the family Enterobacteriaceae, with the exception of Proteus spp., which belong to Morganellaceae.24 K. pneumoniae was the most common CRE species (69% of CRE isolates), while E. coli was the most common ESBL-producing species (79% of ESBL-producing isolates).
 |
Figure 3 Colicinogenic plasmids identified in CRE isolates are active in vitro. (A) MDR_006 kills other E. coli bacteria. MDR_006 was spotted onto LB agar (at 0.8% w/v) without or with mitomycin C (at a final concentration of 0.1 μg mL−1) next to the E. coli K-12 strain BW25113,22 with each strain at a different dilution (1 vs 10−4, respectively). Plates were incubated at 37 °C overnight. The left image shows a schematic representation of the competition conditions. (B) pCol006 harbored by MDR006 encodes a Colicin E1-like toxin. LB agar (at 1.5% w/v) plates were swabbed with E. coli BW25113, its btuB or tsx mutant (as obtained from the Keio collection),22 and four wells were carved into each plate. Wells were filled with culture supernatant (spent media) either produced by MDR_006 or by E. coli BW25113 after overnight growth in the presence of 0.1 μg mL−1 mitomycin C. This panel contains a schematic representation of the growth inhibition assay (left; top), representative images of the obtained growth inhibition zones for all strain-spent media combinations (left; bottom), and quantification of the growth inhibition zones (right). (C) MDR_119 releases cloacin toxin when it is exposed to DNA damage. The panel shows an SDS-PAGE analysis of TCA-precipitated culture supernatants of MDR_119 after overnight growth in the absence (lane 1) or presence (lane 2) of mitomycin C (at a final concentration of 0.5 μg mL−1). Molecular weight markers (M) are on the left, the red arrow indicates the position of the bands originating from the cloacin toxin. |
Clinical data for the 16 CRE isolates is reported in Supplemental Table 2. The median age for patients with CRE infections is 58 years old, and the majority of these individuals have an association with long-term acute care (LTAC) facilities. Specifically, only isolates MDR_018 (CP-CRE E. coli) and MDR_007 (non-CP-CRE K. pneumoniae) are community-acquired. Two patients with K. pneumoniae infections (CP-CRE MDR_009 and non-CP-CRE MDR_007) had a documented history of a previous infection caused by an ESBL-producing strain.
Varied antibiotic resistance profiles were evident among both CRE and ESBL-producing isolates. Of the 16 CRE isolates, 12 (75%) were resistant or intermediately susceptible to all carbapenem antibiotics tested in vitro but were susceptible to at least one alternative drug (Table 1). Of these, only one (MDR_007) was a non-CP-CRE isolate; this isolate was also resistant to polymyxins, having colistin and polymyxin B MICs >4 μg mL−1 (the EUCAST breakpoint for colistin resistance is MIC >2 μg mL−1). The CRE strain closest to being pan-drug resistant is isolate MDR_103, as it is only susceptible to polymyxin antibiotics. The resistance profiles of ESBL-producing isolates to non-carbapenem antibiotics differed depending on the bacterial species (Supplemental Table 3).
 |
Table 1 CRE Isolates (n=16), Carbapenemase Presence/Types, and Antimicrobial Susceptibility |
Detection of Beta-Lactamase Genes in ESBL-Producing and CRE Isolates
Initial PCR-based screening for key groups of non-carbapenemase beta-lactamase genes (blaSHV, blaTEM, blaCTX-M-1-like, and blaOXA-1-like) in select isolates is shown in Supplemental Table 4. blaSHV was detected only in K. pneumoniae isolates, where it was present in 100% of isolates tested. blaTEM, blaCTX-M-1-like, and blaOXA-1-like were detected in 66%, 62%, and 43% of ESBL-producing isolates tested, respectively (Supplemental Table 4). Notably, none of the CRE E. coli carried CTX-M enzymes. By contrast, blaCTX-M-1-like genes were detected among several of the CRE K. pneumoniae, including all of the non-CP-CRE isolates (n=3) and the single CP-CRE isolate with an NDM carbapenemase.
CP-CRE and Non-CP-CRE Mechanisms of Resistance
In Central Texas, like in most other locations, the resistance mechanisms of non-CP-CRE are poorly understood compared to those of CP-CRE. The acquisition of plasmid-borne carbapenemases is a straightforward means of acquiring carbapenem resistance, and the presence of carbapenemase genes and their identities can be easily confirmed. By contrast, resistance mechanisms of non-CP-CRE are more cryptic. According to tests performed at DSHS during routine clinical diagnostic procedures, of the 16 available CRE isolates, the majority (n=12) were CP-CRE (Table 1). These included eight K. pneumoniae and four E. coli isolates. As also seen in countywide trends in Figure 1, the most common carbapenemase in our isolate collection is KPC (n=9) (Table 1). OXA-48 was identified once for each species, and NDM was found in one strain of K. pneumoniae (Table 1). The four non-CP-CRE isolates were all Klebsiella spp., either K. pneumoniae (n=3) or K. aerogenes (n=1).
To gain a more thorough understanding of the genetic factors promoting carbapenem resistance in Central Texas, WGS was initially carried out for seven CRE isolates (Table 1, Figure 2). This panel consisted of K. pneumoniae (n=4), E. coli (n=2), and Klebsiella aerogenes (n=1) strains, which were selected to include CP-CRE (n=4) and non-CP-CRE (n=3) (Figure 2). Key findings from analysis of WGS contigs, including results from screens with CARD, PubMLST, and PlasmidFinder are summarized in Supplemental Table 5.
WGS confirmed the presence of CP alleles in the four analyzed CP-CRE strains. Of the three KPC-expressing isolates analyzed by WGS, two harbored genes for KPC-2 and one for KPC-3, and in all three cases the blaKPC genes were located on a plasmid. By contrast, the OXA-48 carbapenemase of the E. coli isolate (MDR_019) was chromosomal. Interestingly, the outer membrane porin OmpK36 sequence of MDR_119 (one of the CP-CRE containing a blaKPC-2 gene) is identical to the CARD reference sequence for the OmpK36 protein knockout model (NCBI Reference Sequence YP_005228001).25 This model designation implies that this porin variant is not functional and could also contribute to antibiotic resistance.
For the non-CP-CRE K. pneumoniae isolates with WGS (MDR_007 and MDR_049), several beta-lactam resistance determinants were identified (Supplemental Table 5), which are discussed in further detail below. In addition, the two major porins, OmpK35 and OmpK36, and the minor porin, OmpK37, were investigated for deficiencies that could contribute to carbapenem resistance (Table 1). For both isolates, their OmpK35 protein sequences were identical to each other, and to the functional sequence from K. pneumoniae strain ATCC 11296 (NCBI Reference Sequence WP_004141771),26 suggesting OmpK35 is functional in these strains. The OmpK36 sequence of MDR_007 is identical to NCBI Reference Sequence WP_004149145 and is also likely functional. However, OmpK36 from MDR_049 has a premature stop codon at residue 125. This probably renders OmpK36 nonfunctional, which could contribute to MDR_049 resistance. OmpK37 sequences in both isolates are identical to each other (and match NCBI Reference Sequence WP_002902433), and have the mutations I70M and I128M (the latter is located in Loop 327), which have been previously reported in other isolates from South Texas.28 They also contain additional, previously detected, insertions and mutations in the Loop 5 and Loop 6 regions.27 Since there is no information in the literature about the functional effects of these particular variants, analysis of OmpK37 does not provide further insight into the resistance profiles of MDR_007 and MDR_049. Lastly, although the colistin and polymyxin MIC values for MDR_007 are >4 μg mL−1, this strain does not harbor known mcr genes and does not have any mutations to mgrB, phoPQ, or ccrB that would suggest a genetic cause for resistance.
The non-CP-CRE K. aerogenes strain (MDR_129) did not have any obvious beta-lactam resistance determinants based on CARD RGI analysis of WGS. However, independent BLAST searches identified a chromosomal AmpC sequence that has been previously reported.29 Recent work on a hospital outbreak of non-CP-CRE K. aerogenes implicated mutations in AmpD, a regulator of AmpC, and OmpK36 as drivers of carbapenem resistance in this species.30 Using the same carbapenem-susceptible strain (KCTC 2190/ATCC 13048) for all reference sequences,31 we found that MDR_129 has wild-type AmpD and OmpK35, but harbors a mutant allele of OmpK36. OmpK36 in MDR_129 contained three individual residue changes (D189G, D205E, N279D) along with seven residue changes within a designated hypervariable region. All of these mutations were documented together in several non-CP-CRE K. aerogenes isolates from the aforementioned hospital outbreak,30 however their role in resistance remains unclear.
The genetic factors and conditions required for the transition of strains to non-CP-CRE are not well-defined. Evidence provided here suggests that non-CP-CRE isolates MDR_049 and MDR_129 likely have defects in porin function, which can be selected for during carbapenem treatment of infections caused by ESBL-producing bacteria. Despite the limited clinical data available, we also know that the patient from whom MDR_007 was isolated had previously suffered from an infection caused by an ESBL-producing strain. Therefore, it is plausible that this individual was exposed to carbapenems during treatment of this prior infection. It was recently shown that the use of imipenem during the treatment of infections caused by ESBL-producing strains can lead to outer membrane remodeling (especially through OmpK36), resulting in a CRE phenotype post-treatment.32 Overall, this suggests that MDR_007 may have undetected porin defects that make it carbapenem resistant and hints at the possibility of antibiotic-treated ESBL-producing strains providing a reservoir for non-CP-CRE strains.
K. pneumoniae ST307 Background is Present Among Both CRE and ESBL Isolates
Non-CP-CRE K. pneumoniae isolates MDR_007 and MDR_049 belong to the emerging ST307 sequence type and appear to be clonal based on analysis of assembled contigs by scgMLST using 629 K. pneumoniae loci.15 The closest allelic profile match for both MDR_007 and MDR_049 was scgST-12390. Furthermore, the capsular type, based on analysis of wzi,16 was the same for both isolates (allele 173). Like other ST307 strains, both MDR_007 and MDR_049 have the chromosomal GyrA S83I and ParC S80I mutations, which are conserved in this ST and are associated with fluoroquinolone resistance.33 However, neither of these strains have the GyrA D87N mutation that is present in the Texas lineage of ST307 as defined by Wyres et al, indicating they most likely belong to the global ST307 lineage.33 ST307, along with ST258, was first identified in the United States as a carrier of blaKPC-2 during clonal spread of CP-CRE in hospitals in the Houston, Texas area.34 While ST307 has recently been described as an emerging, globally-disseminated, high-risk clone of K. pneumoniae,35 in the United States it is still found predominantly in the southern region, where it is mainly localized in the Houston area.36 Our observations suggest ST307 is present more broadly across Texas. Additional phylogenomics analysis would be necessary to clarify the precise evolutionary relationship between the ST307 isolates identified here and the numerous sublineages of the global ST307 lineage described by Wyres et al.33
We hypothesized that non-CP-CRE K. pneumoniae ST307 isolates like MDR_007 and MDR_049 carrying the plasmid that encodes CTX-M-15 (Supplemental Table 5), may originate from ESBL-producing precursors. To investigate this hypothesis, we screened CRE (n=10) and ESBL-producing (n=17) K. pneumoniae isolates for evidence of this genotype. First, we sequenced the gyrA QRDR region of these strains; ST307 isolates should have the S83I mutation in this region. We also performed PCRs for blaCTX-M-1-like, blaTEM, and blaOXA-1-like beta-lactamase genes. All three of these resistance determinants are present on IncFIB(K)/IncFII(K) replicon plasmids containing the blaCTX-M-15 gene that we identified by WGS (Supplemental Table 5). Using this analysis, we categorized all tested isolates based on their CRE/ESBL status, gyrA mutations, beta-lactamase gene presence/absence, and antibiotic susceptibility phenotype (Table 2).
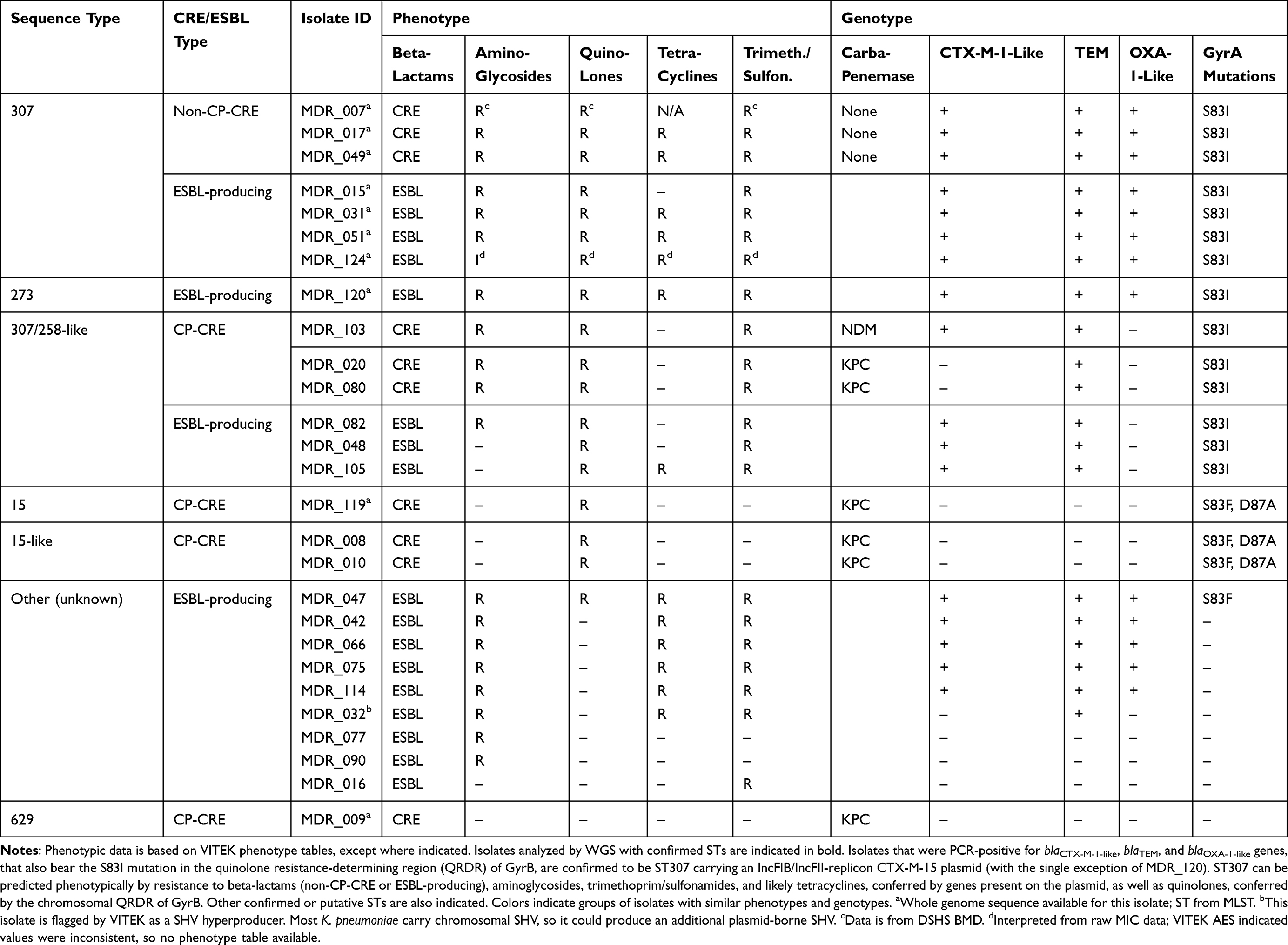 |
Table 2 Antibiotic Susceptibility Phenotype versus Genotype for 27 Klebsiella pneumoniae Isolates (All K. pneumoniae Except for MDR_053, MDR_055, and MDR_104, for Which the DNA Extraction Proved to Be Challenging) |
We identified eight strains that could be related, based on these criteria, including both non-CP-CRE (n=3) and ESBL-producing isolates (n=5) (Table 2). The additional GyrA D87N mutation is absent in all eight strains, indicating that, provided they are ST307, they belong to the global rather than the Texas ST307 lineage.33 Moreover, all strains have near identical resistance profiles: they are resistant to aminoglycosides, quinolones, tetracyclines, and trimethoprim/sulfonamides except for MDR_015 (which is susceptible to tetracyclines) and MDR_007 (where the tetracycline MIC is not available). With the exception of quinolones, where resistance is chromosomally encoded for ST307 due to the mutations found on GyrA, resistance to the other three antibiotic classes can be attributed, at least in part, to genes usually encoded on the plasmid. This plasmid does not always carry tetA,28,33 which may be why MDR_015 remains tetracycline susceptible.
Whole genomes of two of these eight isolates had been sequenced previously (CRE isolates MDR_007 and MDR_049) and confirmed to be ST307, as discussed above. To confirm the sequence type of the remaining six isolates, a second round of WGS was conducted (Figure 2). Of these six isolates, all except one (MDR_120) were ST307, with the closest scgST allelic profile 12,390, and capsular type wzi allele 173, as shown for MDR_007 and MDR_049. This shows that all non-CP-CRE K. pneumoniae collected in this study (n=3) and at least four ESBL-producing K. pneumoniae (21%) belonged to a clonal lineage of ST307.
Together, these data suggest that in vitro resistance to beta-lactams, as well as four other classes of antibiotics (Table 2), is linked to ST307 when combined with carriage of a CTX-M-encoding plasmid. This genotype can be found among both non-CP-CRE and ESBL-producing isolates. This suggests that ST307 expressing CTX-M-15 is a locally prevalent ESBL-producing strain which, given the right conditions, may transition from ESBL to CRE status. We recognize that a caveat to this conclusion is the small sample size (10 CRE and 17 ESBL-producing isolates) analyzed here, which was due to limitations in isolate availability in a location where CRE K. pneumoniae are emergent. That said, our results provide the motivation for a larger study, aiming to further clarify the relationships between ESBL-producing and non-CP-CRE strains.
CRE Isolates Carry Active Colicinogenic Plasmids
In addition to antibiotic resistance genes, our WGS analysis provided information on other characteristics of these CRE strains, including the presence of toxins involved in interbacterial competition. An example of such toxins are colicins, a class of bacteriocins produced by E. coli and other enterobacterial species, which kill closely related strains.37 Colicins bind to surface receptors of prey bacteria, and this allows their cellular entry and lethal activity. While not commonly searched for in clinical studies, colicins are generally known to play an important role in the fitness of E. coli strains in the presence of other bacteria.38 As such, their presence in clinical strains could influence the success of these pathogenic species during patient colonization. Colicinogenic plasmids were identified in three of the seven CRE isolates (43%) that were analyzed by WGS. These were the CP-CRE E. coli isolate MDR_006 (harboring pCol006), the CP-CRE K. pneumoniae isolate MDR_119 (harboring pCol119), and the non-CP-CRE K. aerogenes MDR_129 (harboring pCol129); these plasmids did not contain any antimicrobial resistance genes (Supplemental Table 5). Here, we investigated further the colicinogenic activity of MDR_006 and MDR_119.
MDR_006, which contains a colicin gene similar to toxins from the E series, spontaneously killed prey E. coli in vitro (Figure 3A). This activity increased in the presence of mitomycin C, which is known to induce colicin expression for toxins of this family through DNA damage.39–41 Further activity testing of cell supernatant isolated from MDR_006 grown in the presence of mitomycin C against E. coli btuB and tsx receptor mutants demonstrated that Col006 requires the BtuB receptor to enter and kill prey bacteria (Figure 3B), as observed for the majority of colicins of the E series.42
Col119 and Col129 are identical to each other and similar to cloacin.43 Lacking a suitable prey strain to test the killing activity of these isolates, we isolated cell supernatants of MDR_119 grown with and without mitomycin C, and analyzed their protein content by SDS-PAGE. We found that the supernatant from the strain that was DNA damaged was significantly richer in protein (Figure 3C), in agreement with induction of cloacin expression and release of cellular contents through cell lysis.44,45 We also detected a major band at ~60 kDa when mitomycin C was used (Figure 3C, lane 2), which correlates with the predicted molecular weight of Col119. Considering that when colicinogenic strains are DNA damaged, the expressed toxin becomes the major protein of the cell and is then released during cell lysis,37 this indicates that pCol119 is also an active colicinogenic plasmid. With multidrug-resistant strains harboring active colicinogenic plasmids, it can be speculated that these toxins facilitate pathogens clearing their niche of commensals during infection or, alternatively, play a role in competition between different pathogens that coexist in clinical environments.
Conclusion
Because CRE are emerging in Central Texas, it is important to understand local genetic and phenotypic trends to optimize current infection treatment and prevention strategies and attempt to anticipate evolution of resistance in clinical strains. To the best of our ability, given the limited number of 16 CRE isolates observed in the course of this study, we have provided the first analysis of local CRE as contextualized among the 116 ESBL-producing Enterobacterales collected during the same time period. Our results revealed that Klebsiella pneumoniae caused the majority of CRE infections and that sequence type (ST) 30735 is common among both non-CP-CRE and EBSL-producing isolates. Our data also suggests that porin mutations may promote the transition of circulating ST307 isolates from ESBL-producing to CRE. Finally, we demonstrate that several CRE isolates are capable of producing colicin toxins, which could influence the competitiveness of these bacteria in hospital settings. The suggested genetic relationship between non-CP-CRE and ESBL-producing K. pneumoniae warrants further monitoring going forward.
Ethical Approval
This research study (Protocol Number 2019-09-0047) has an Institutional Review Board (IRB) Exempt status based on 45 CFR 46.101(b)(4), as determined by the Office of Research Support and Compliance at The University of Texas at Austin. The collection and use of protected health information (PHI) was also approved during this review. All researchers analyzing isolate and clinical data were blinded to patient identity.
Acknowledgments
We appreciate assistance from Jenny Harder at Ascension Seton in providing the clinical isolates and associated phenotypic data.
Funding
Research reported in this publication was supported by the following: The University of Texas at Austin Faculty Support Funds (PIs Davies and Mavridou), and Tito’s Handmade Vodka (PI Davies).
Disclosure
The authors declare that there are no conflicts of interest.
References
1. Logan LK, Weinstein RA. The epidemiology of carbapenem-resistant Enterobacteriaceae: the impact and evolution of a global menace. J Infect Dis. 2017;215:S28–S36. doi:10.1093/infdis/jiw282
2. Castanheira M, Deshpande LM, Mendes RE, Canton R, Sader HS, Jones RN. Variations in the occurrence of resistance phenotypes and carbapenemase genes among. Open Forum Infect Dis. 2019;6:S23–S33. doi:10.1093/ofid/ofy347
3. Bouganim R, Dykman L, Fakeh O, et al. The clinical and molecular epidemiology of noncarbapenemase-producing carbapenem-resistant. Open Forum Infect Dis. 2020;7:ofaa299. doi:10.1093/ofid/ofaa299
4. Suay-García B, Pérez-Gracia MT. Present and future of carbapenem-resistant. Antibiotics. 2019;8:435–456.
5. Woodworth KR, Walters MS, Weiner LM, et al. Vital signs: containment of novel multidrug-resistant organisms and resistance mechanisms - United States, 2006-2017. MMWR Morb Mortal Wkly Rep. 2018;67:396–401. doi:10.15585/mmwr.mm6713e1
6. Bezabih YM, Sabiiti W, Alamneh E, et al. The global prevalence and trend of human intestinal carriage of ESBL-producing Escherichia coli in the community. J Antimicrob Chemother. 2021;76:22–29. doi:10.1093/jac/dkaa399
7. Tian X, Sun S, Jia X, Zou H, Li S, Zhang L. Epidemiology of and risk factors for infection with extended-spectrum β-lactamase-producing carbapenem-resistant Enterobacteriaceae: results of a double case-control study. Infect Drug Resist. 2018;11:1339–1346. doi:10.2147/IDR.S173456
8. CDC (Centers for Disease Control and Prevention). CRE technical information; 2019. Available from: https://www.cdc.gov/hai/organisms/cre/technical-info.html.
9. CDC AR Laboratory Network. Guidance for testing CRE & CRPA in state and local public health laboratories; 2020.
10. Pierce VM, Simner PJ, Lonsway DR, et al. Modified carbapenem inactivation method for phenotypic detection of carbapenemase production among Enterobacteriaceae. J Clin Microbiol. 2017;55:2321–2333. doi:10.1128/JCM.00193-17
11. NHSN (National Healthcare Safety Network). Identifying Healthcare-associated Infections (HAI) for NHSN Surveillance; 2022.
12. Bushnell B. BBTools. Available from: sourceforge.net/projects/bbmap/.
13. Jolley KA, Bray JE, Maiden MCJ. Open-access bacterial population genomics: bIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018;3:124.
14. Jolley KA, Maiden MC. BIGSdb: scalable analysis of bacterial genome variation at the population level. BMC Bioinform. 2010;11(1):595. doi:10.1186/1471-2105-11-595
15. Hennart M, Guglielmini J, Maiden MCJ, Jolley KA, Criscuolo A, Brisse S. A dual barcoding approach to bacterial strain nomenclature: genomic taxonomy of Klebsiella pneumoniae strains. Molecular Biology and Evolution. 2022;39 7 :msac135 doi:10.1093/molbev/msac135.
16. Brisse S, Passet V, Haugaard AB, et al. wzi gene sequencing, a rapid method for determination of capsular type for Klebsiella strains. J Clin Microbiol. 2013;51(12):4073–4078. doi:10.1128/JCM.01924-13
17. McArthur AG, Waglechner N, Nizam F, et al. The comprehensive antibiotic resistance database. Antimicrob Agents Chemother. 2013;57(7):3348–3357. doi:10.1128/AAC.00419-13
18. Carattoli A, Zankari E, García-Fernández A, et al. In silico detection and typing of plasmids using plasmidfinder and plasmid multilocus sequence typing. Antimicrob Agents Chemother. 2014;58(7):3895–3903. doi:10.1128/AAC.02412-14
19. Dallenne C, Da Costa A, Decré D, Favier C, Arlet G. Development of a set of multiplex PCR assays for the detection of genes encoding important beta-lactamases in Enterobacteriaceae. J Antimicrob Chemother. 2010;65:490–495. doi:10.1093/jac/dkp498
20. Brisse S, Milatovic D, Fluit AC, et al. Comparative in vitro activities of ciprofloxacin, clinafloxacin, gatifloxacin, levofloxacin, moxifloxacin, and trovafloxacin against Klebsiella pneumoniae, Klebsiella oxytoca, Enterobacter cloacae, and Enterobacter aerogenes clinical isolates with alterations in GyrA and ParC proteins. Antimicrob Agents Chemother. 1999;43:2051–2055. doi:10.1128/AAC.43.8.2051
21. Poirel L, Walsh TR, Cuvillier V, Nordmann P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn Microbiol Infect Dis. 2011;70(1):119–123. doi:10.1016/j.diagmicrobio.2010.12.002
22. Baba T, Ara T, Hasegawa M, et al. Construction of Escherichia coli K-12 in-frame, single-gene knockout mutants: the Keio collection. Mol Syst Biol. 2006;2. doi:10.1038/msb4100050
23. Tadese BK, Darkoh C, DeSantis SM, Mgbere O, Fujimoto K. Clinical epidemiology of carbapenem-resistant Enterobacterales in the greater Houston region of Texas: a 6-year trend and surveillance analysis. J Glob Antimicrob Resist. 2022;30:222–227. doi:10.1016/j.jgar.2022.06.019
24. Adeolu M, Alnajar S, Naushad S, Gupta R. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int J Syst Evol Microbiol. 2016;66:5575–5599. doi:10.1099/ijsem.0.001485
25. Liu P, Li P, Jiang X, et al. Complete genome sequence of Klebsiella pneumoniae subsp. pneumoniae HS11286, a multidrug-resistant strain isolated from human sputum. J Bacteriol. 2012;194:1841–1842. doi:10.1128/JB.00043-12
26. Sugawara E, Kojima S, Nikaido H. Klebsiella pneumoniae major porins OmpK35 and OmpK36 allow more efficient diffusion of β-lactams than their Escherichia coli Homologs OmpF and OmpC. J Bacteriol. 2016;198:3200–3208. doi:10.1128/JB.00590-16
27. Doménech-Sánchez A, Hernández-Allés S, Martínez-Martínez L, Benedí VJ, Albertí S. Identification and characterization of a new porin gene of Klebsiella pneumoniae: its role in beta-lactam antibiotic resistance. J Bacteriol. 1999;181:2726–2732. doi:10.1128/JB.181.9.2726-2732.1999
28. Black CA, So W, Dallas SS, et al. Predominance of non-carbapenemase producing carbapenem-resistant Enterobacterales in South Texas. Front Microbiol. 2021;11:623574. doi:10.3389/fmicb.2020.623574
29. Preston KE, Radomski CC, Venezia RA. Nucleotide sequence of the chromosomal ampC gene of Enterobacter aerogenes. Antimicrob Agents Chemother. 2000;44:3158–3162. doi:10.1128/AAC.44.11.3158-3162.2000
30. Malek A, McGlynn K, Taffner S, et al. Next-generation-sequencing-based hospital outbreak investigation yields insight into Klebsiella aerogenes population structure and determinants of carbapenem resistance and pathogenicity. Antimicrob Agents Chemother. 2019;63. doi:10.1128/AAC.02577-18
31. Lavigne JP, Sotto A, Nicolas-Chanoine MH, et al. Membrane permeability, a pivotal function involved in antibiotic resistance and virulence in Enterobacter aerogenes clinical isolates. Clin Microbiol Infect. 2012;18:539–545. doi:10.1111/j.1469-0691.2011.03607.x
32. Tian X, Wang Q, Perlaza-Jiménez L, et al. First description of antimicrobial resistance in carbapenem-susceptible Klebsiella pneumoniae after imipenem treatment, driven by outer membrane remodeling. BMC Microbiol. 2020;20:218. doi:10.1186/s12866-020-01898-1
33. Wyres KL, Hawkey J, Hetland MAK, et al. Emergence and rapid global dissemination of CTX-M-15-associated Klebsiella pneumoniae strain ST307. J Antimicrob Chemother. 2019;74:577–581. doi:10.1093/jac/dky492
34. Castanheira M, Farrell SE, Wanger A, Rolston KV, Jones RN, Mendes RE. Rapid expansion of KPC-2-producing Klebsiella pneumoniae isolates in two Texas hospitals due to clonal spread of ST258 and ST307 lineages. Microb Drug Resist. 2013;19:295–297. doi:10.1089/mdr.2012.0238
35. Peirano G, Chen L, Kreiswirth BN, Pitout JDD. Emerging antimicrobial-resistant high-risk Klebsiella pneumoniae clones ST307 and ST147. Antimicrob Agents Chemother. 2020;64. doi:10.1128/AAC.01148-20
36. van Duin D, Arias CA, Komarow L, et al. Molecular and clinical epidemiology of carbapenem-resistant Enterobacterales in the USA (CRACKLE-2): a prospective cohort study. Lancet Infect Dis. 2020;20:731–741. doi:10.1016/S1473-3099(19)30755-8
37. Cascales E, Buchanan SK, Duché D, et al. Colicin biology. Microbiol Mol Biol Rev. 2007;71:158–229. doi:10.1128/MMBR.00036-06
38. Majeed H, Gillor O, Kerr B, Riley MA. Competitive interactions in Escherichia coli populations: the role of bacteriocins. ISME J. 2011;5:71–81. doi:10.1038/ismej.2010.90
39. Butala M, Zgur-Bertok D, Busby SJ. The bacterial LexA transcriptional repressor. Cell Mol Life Sci. 2009;66:82–93. doi:10.1007/s00018-008-8378-6
40. Janion C. Inducible SOS response system of DNA repair and mutagenesis in Escherichia coli. Int J Biol Sci. 2008;4:338–344. doi:10.7150/ijbs.4.338
41. Gillor O, Vriezen JAC, Riley MA. The role of SOS boxes in enteric bacteriocin regulation. Microbiology. 2008;154:1783–1792. doi:10.1099/mic.0.2007/016139-0
42. Kim YC, Tarr AW, Penfold CN. Colicin import into E. coli cells: a model system for insights into the import mechanisms of bacteriocins. Biochim Biophys Acta. 2014;1843:1717–1731. doi:10.1016/j.bbamcr.2014.04.010
43. Hagan EC, Mobley HL. Uropathogenic Escherichia coli outer membrane antigens expressed during urinary tract infection. Infect Immun. 2007;75:3941–3949. doi:10.1128/IAI.00337-07
44. Pugsley AP, Rosenbusch JP. Release of colicin E2 from Escherichia coli. J Bacteriol. 1981;147:186–192. doi:10.1128/jb.147.1.186-192.1981
45. Pugsley AP, Goldzahl N, Barker RM. Colicin E2 production and release by Escherichia coli K12 and other Enterobacteriaceae. J Gen Microbiol. 1985;131:2673–2686. doi:10.1099/00221287-131-10-2673
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.