Back to Journals » Infection and Drug Resistance » Volume 11
Carbapenem-resistant Acinetobacter baumannii and Pseudomonas aeruginosa: characterization of carbapenemase genes and E-test evaluation of colistin-based combinations
Authors Ramadan RA ![]() , Gebriel MG
, Gebriel MG ![]() , Kadry HM
, Kadry HM ![]() , Mosallam A
, Mosallam A ![]()
Received 5 April 2018
Accepted for publication 2 June 2018
Published 22 August 2018 Volume 2018:11 Pages 1261—1269
DOI https://doi.org/10.2147/IDR.S170233
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Eric Nulens
Raghdaa A Ramadan,1 Manar G Gebriel,1 Heba M Kadry,1 Ahmed Mosallem2
1Medical Microbiology and Immunology Department, Faculty of Medicine, Zagazig University, Zagazig, Sharkia, Egypt; 2Anesthesia and Surgical Intensive Care Department, Faculty of Medicine, Zagazig University, Zagazig, Sharkia, Egypt
Background: Carbapenamase producing Acinetobacter baumannii and Pseudomonas aeruginosa are emerging worldwide limiting the use of carbapenems as effective and safe drugs.
Purpose: To characterize different carbapenemase genes carried by carbapenem-resistant (CR) A. baumannii and P. aeruginosa isolates and to evaluate the in vitro effect of some colistin-based combinations by E-test method in Zagazig University Hospitals ICU isolates.
Methods: CR A. baumannii and P. aeruginosa isolated from the surgical intensive care unit (ICU) were tested for carbapenemase genes by polymerase chain reaction and the effect of colistin/meropenem and colistin/tigecycline combinations was evaluated by E-test.
Results: Genes coding for OXA-23, NDM and GES were detected in 90, 66.7 and 50% of CR A. baumannii, respectively, while genes coding for VIM, GES, NDM and IMP were detected in 50, 40.9, 27.3 and 18.2% of CR P. aeruginosa, respectively. Colistin/tigecycline combination showed synergistic and additive effect in 20% and 60% of A. baumannii isolates, respectively, while colistin/meropenem combination showed synergistic and additive effect in 63.6% and 36.4% of P. aeruginosa, respectively.
Conclusion: Carbapenemase genes carriage accounts for high level carbapenem resistance in our isolates. Colistin/tigecycline and colistin/meropenem combinations can be considered for treatment of severe infections by CR A. baumannii and P. aeruginosa, respectively.
Keywords: meropenem, multidrug resistant, synergy, fractional inhibitory concentration, intensive care unit
Introduction
Carbapenems are well-known β-lactam antibiotics with broad spectrum activity that have long been used for treating severe infections caused by otherwise resistant Gram-negative bacteria (GNB).1 Nevertheless, these agents have been misused to the extent that they are no longer effective due to the high rate of development of different resistant mechanisms that target them.2
Among these resistant mechanisms, acquired carbapenem hydrolyzing enzymes (carbapenemases) emergence and their wide dissemination represented a global health threat which poses serious therapeutic and infection control challenges being associated with elevated mortality rates and increase in hospital stay.3,4 For these reasons, clinicians became highly concerned about rapidly identifying bacteria carrying these genes to reduce or prevent this problem in health care centers.5
Carbapenemases are diverse and include members of β-lactamase molecular classes A, B and D.6 Class D oxacillinases (OXA type) are the most prevalent in Acinetobacter baumannii while in Pseudomonas aeruginosa, carbapenemases are mostly class B (metallo-β-lactamases).7,8
Although the definitions for MDR and XDR (multi- and extensively drug-resistant) GNB do not necessarily require resistance to carbapenems,9 the carbapenem-resistant (CR) phenotype is very common for MDR and particularly for XDR isolates because carbapenemase-encoding genes are carried on mobile genetic elements that usually carry genetic determinants for resistance to other antibiotics.10
The lack of effective antibiotics against CR-GNB and the drying out of the pipeline of new antibiotics forced physicians to recall old antibiotics (ie, polymyxins) back into clinical use.11 Most CR-GNB, including those with XDR profile, show in vitro susceptibility to colistin.12 However, the possibility of emergence of hetero-resistance and the low clinical efficacy of colistin in monotherapy in addition to its high toxicity, led to the utilization of colistin in combination with other antibiotics for desperately ill patients with serious infections especially in intensive care units (ICUs).13
The E-test method for antibiotic combination testing is an easily performed, rapid method that showed good concordance with other methods used for evaluating the effect of antibiotic combinations.14,15 It is thus reliable when results are to be used in guiding combination therapies.16
This study was conducted to characterize different carbapenemase genes carried by CR A. baumannii and P. aeruginosa isolates and to evaluate the in vitro effect of some colistin-based combinations by E-test method in Zagazig University Hospitals ICU isolates.
Materials and methods
Bacterial isolates and primary susceptibility testing
Fifty A. baumannii and 43 P. aeruginosa non-duplicate clinical isolates collected over a 6-month period (July 2017–December 2017) from the surgical ICU of Zagazig University Hospitals were included in the study. Identification was done by Vitek 2 system (Biomerieux, Marcy l’Etoile, France). The designation “A. baumannii” here is used to refer to the pathogenic members of the Acinetobacter baumannii complex.
Isolates were tested for antimicrobial susceptibility by the modified Kirby-Bauer disc diffusion method. Antimicrobial susceptibility included that of carbapenems (imipenem and meropenem), aminoglycosides (amikacin and tobramycin), antipseudomonal penicillin/β-lactamase inhibitor (piperacillin/tazobactam), monobactam (aztreonam), broad spectrum cephalosporins (ceftazidime, cefepime), and antipseudomonal fluoroquinolones (ciprofloxacin, levofloxacin). Minimum inhibitory concentration (MIC) values of meropenem and colistin were detected by broth microdilution method in those isolates found resistant to carbapenems and sub-cultures of these isolates on glycerol broth were stored at –20°C. P. aeruginosa ATCC®27853 was used as a quality control strain (American Type Culture Collection [ATCC], Manassas, VA, USA).
Detection of bla genes responsible for carbapenem resistance in CR isolates by polymerase chain reaction (PCR)
Bacterial DNA was extracted from isolated colonies using QIAamp® DNA Mini kit (Qiagen GmbH, Hilden, Germany). Amplification was done using Taq PCR Master Mix (Qiagen GmbH). Two multiplex PCRs were done; the first was for detection of genes encoding class A carbapenemase (blaKPC and blaGES) and genes encoding class B metallo-beta-lactamases (MBL) (blaIMP and blaVIM) in all isolates using primers and conditions described by Monteiro et al.5 Klebsiella pneumoniae ATCC BAA-1705 was used as the positive control for blaKPC. The other multiplex PCR was for detection of genes encoding class D carbapenemase (OXA-type carbapenemase) (blaOXA-23, blaOXA-24, blaOXA-58, blaOXA-143) in A. baumannii isolates only as described by Woodford et al and Higgins et al.17,18 Detection of (blaNDM) genes encoding New Delhi metallo-beta-lactamases (NDM) was performed in a separate reaction for all isolates as described by Poirel et al.19
Individual MIC values determination by E-test
MIC values of meropenem and colistin against all isolates and MIC values of tigecycline against A. baumannii isolates were determined separately by E-test to be used in fractional inhibitory concentration (FIC) calculation. P. aeruginosa ATCC®27853 was used as a quality control strain. Results were interpreted according to the Clinical and Laboratory Standards Institute 2017,20 except for the MIC values of tigecycline against A. baumannii where the standards specified for the Enterobacteriaceae family by the US Food and Drug Administration were used (susceptible, MIC ≤2 µg/mL, intermediate, MIC is >2 to <8 µg/mL, resistant, MIC ≥8 µg/mL).21
Antimicrobial combination testing
The effect of antimicrobial combinations was evaluated for colistin/meropenem and colistin/tigecycline in A. baumannii isolates and for colistin/ meropenem in P. aeruginosa isolates by “Perpendicular E-test” method. Briefly, E-test strips of the two antimicrobials were placed on the agar at right angles intersecting at the respective MIC values previously determined for each antibiotic separately against the isolate. The agar plates were then incubated at 37°C for 18 hours. The MIC value of each antimicrobial in the combination was read.22
The fractional inhibitory concentration index (∑ FIC) was calculated for each antimicrobial combination according to the following formula: ∑ FIC = FIC [A]+ FIC [B]23
FIC [A]= MIC drug A when tested in combination with drug B/ MIC of drug A alone
FIC [B]= MIC drug B when tested in combination with drug A/ MIC of drug B alone
Interpretation of the ∑ FIC results was done as follows: ≤0.5, synergy; >0.5 to ≤1.0, additively; >1.0 to <4.0, indifference; and ≥4, antagonism.23
When the individual MIC value of an isolate was more than the highest concentration on any of the E-test strips, the strips were placed intersecting at the highest concentration present on this E-test strip. The MIC value used to calculate the FIC index was then considered as one 2-fold dilution above the highest concentration on the E-test strip.14 This was necessary for meropenem in most of the isolates where individual MIC values for meropenem were ≥32 µg/mL.
This study was approved by the institutional review board (IRB) – Faculty of Medicine, Zagazig University. A written informed consent was obtained from each patient or the guardians of unconscious patients.
Results
Thirty out of 50 A. baumannii isolates (60%) and 22 out of 43 P. aeruginosa isolates (51%) tested were CR; they showed resistance to imipenem and meropenem by disc diffusion method as well as resistance to meropenem by broth microdilution (ie, MIC ≥8 µg/mL). These isolates were also resistant to the majority of the tested antibiotic classes expressing an MDR or XDR phenotype.
PCR results showed that all CR A. baumannii isolates harbored at least one of the genes under investigation. The most common resistance gene was blaOXA-23 in 90% of the isolates (27/30) followed by blaNDM in 66.7% of the isolates (20/30), then blaGES in 50% of the isolates (15/30), while the blaKPC, blaIMP, blaVIM, blaOXA-24, blaOXA-58 and blaOXA-143 genes were not detected. On the other hand, all CR P. aeruginosa isolates harbored at least one of the tested genes except a single isolate in which none of the tested genes was detected. They showed high prevalence of class B carbapenemases where 18/22 (81.8%) harbored at least one of the tested MBL genes. Meanwhile blaGES was the only serine carbapenemase gene detected in (40.9%) of P. aeruginosa isolates (Table 1).
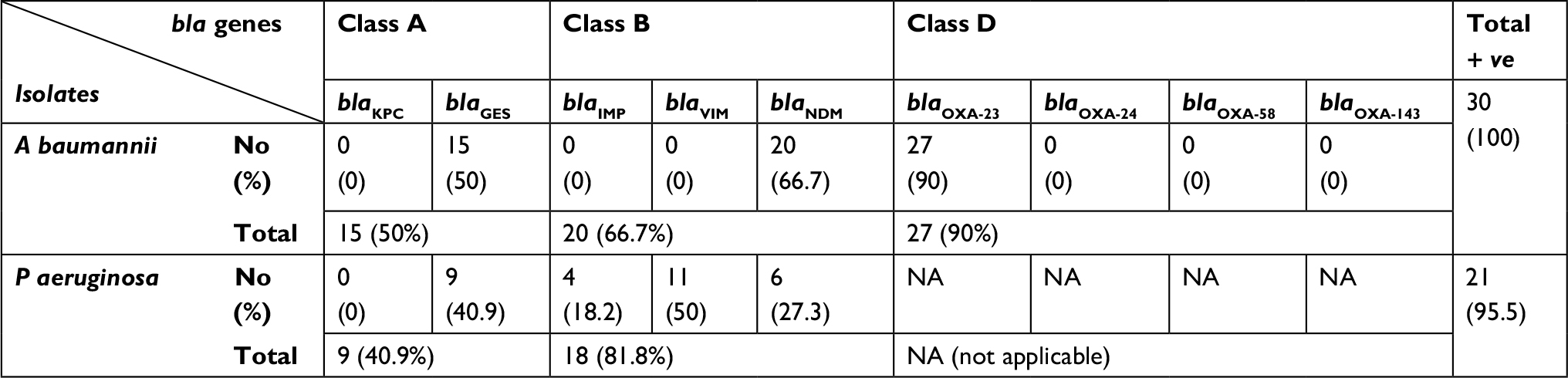 | Table 1 bla genes detected by PCR in carbapenem resistant isolates Abbreviations: A baumannii, Acinetobacter baumannii; P aeruginosa, Pseudomonas aeruginosa; PCR, polymerase chain reaction. |
Meropenem MICs for CR A. baumannii isolates by broth dilution method ranged from 32 to 512 µg/mL (Table 2). By E-test, all meropenem MIC values were ≥32 µg/mL (ie, the maximum value on meropenem strip). All these isolates were colistin-sensitive by broth microdilution, where colistin MIC values ranged from 0.38 to 1.5 µg/mL. However, 15 isolates (50%) were tigecycline-resistant (MIC ≥8 µg/mL), 12 isolates (40%) were intermediate and three isolates (10%) were tigecycline-sensitive (MIC ≤2 µg/mL). MIC for tigecycline by E-test ranged from 2 to –16 µg/mL.
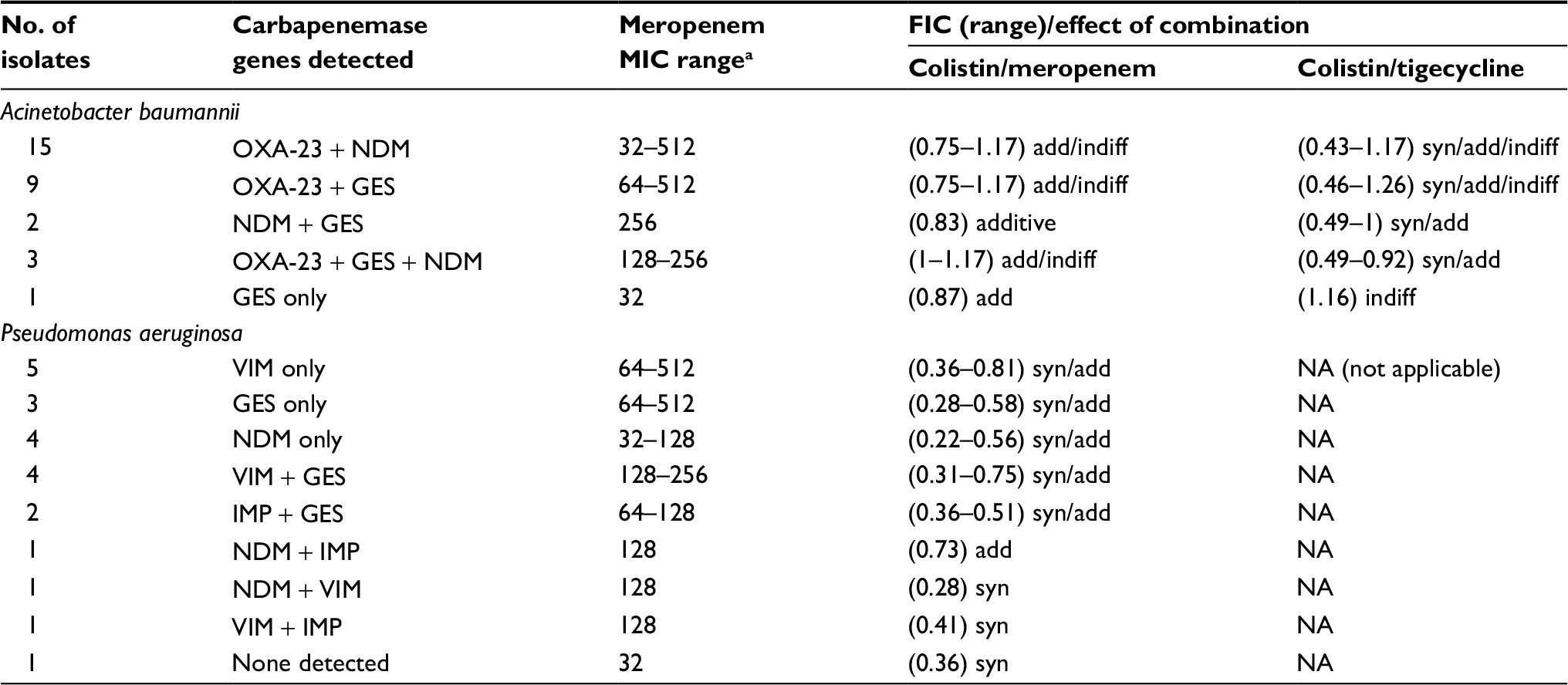 | Table 2 Carbapenemase gene distribution among isolates, the corresponding meropenem MIC range and the effect of antimicrobial combination Notes: aMeasured by broth microdilution method. Abbreviations: FIC, fractional inhibitory concentration; MIC, minimum inhibitory concentration; add, additive; indiff, indifference; syn, synergy. |
Meropenem MICs for CR P. aeruginosa isolates by broth dilution method ranged from 32 to 512 µg/mL (Table 2). By E-test, all meropenem MIC values were ≥32 µg/mL. MICs for colistin ranged from 0.38 to 16 µg/mL. Twenty isolates (90.9%) were colistin-sensitive (MIC ≤2 µg/mL), and two isolates (9.1%) were resistant (MIC ≥4 µg/mL).
In the 30 CR A. baumannii isolates tested, both colistin/meropenem and colistin/tigecycline combination showed additive effects against most of the isolates; 25 (83.3%) and 18 (60%), respectively (Figure 1), but synergism was only detected for colistin/tigecycline combination in six isolates (20%). Meanwhile, no antagonism was detected for either combinations (Table 3).
 | Figure 1 Colistin/tigecycline combination test (Acinetobacter baumannii). Notes: The MIC of colistin was 0.75 µg/mL when tested alone but was 0.25 µg/mL when tested in combination. The MIC of tigecycline was 16 µg/mL when tested alone but was 3 µg/mL when tested in combination. This was an additive effect (isolate no. 12; Table S1). Abbreviation: MIC, minimum inhibitory concentration. |
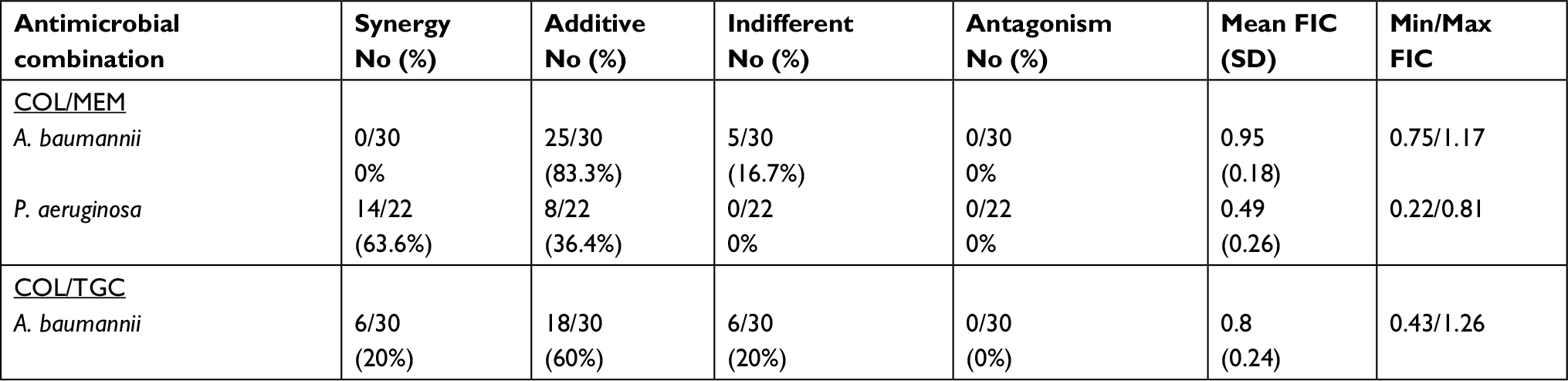 | Table 3 The effect of colistin based combinations against Acinetobacter baumannii and Pseudomonas aeruginosa Abbreviations: COL, colistin; MEM, meropenem; TGC, tigecycline. |
Out of 22 CR P. aeruginosa isolates tested, the colistin/meropenem combination showed synergistic effects against most 14 (63.6%) isolates (Figure 2) and additive effect in eight (36.4%) isolates (Table 3).
 | Figure 2 Colistin/meropenem combination test (Pseudomonas aeruginosa). Notes: The MIC of colistin was 0.75 µg/mL when tested alone but was 0.25 µg/mL when tested in combination. The MIC of meropenem was >32 µg/mL when tested alone but was 2 µg/mL when tested in combination. This was a synergistic effect (isolate no. 1; Table S2). Abbreviation: MIC, minimum inhibitory concentration. |
Discussion
This study depicts the increasingly reported threat of carbapenem resistance in the two frequently isolated Gram negative non-fermentative bacilli in Zagazig University Hospitals (ie, A. baumannii and P. aeruginosa), where 60% of A. baumannii and 43% of P. aeruginosa isolated over a 6-month period were CR. They all showed high level meropenem resistance with MIC values ranging from 32 to 512 µg/mL. In addition, they all exhibited an MDR profile being non-susceptible to at least one agent in at least three of the tested antimicrobial classes. This is attributed to the fact that mobile genetic elements carrying carbapenemase genes also carry genetic determinants of other antimicrobial resistance.
Screening our CR isolates for nine carbapenemase genes previously reported in our region yielded at least a single PCR product in 51/52 isolates which underscores the production of carbapenem-hydrolyzing enzymes as a common mechanism for high level carbapenem resistance displayed by our isolates. In a study in Saudi Arabia only 11 of 39 (18%) CR P. aeruginosa yielded a PCR product for carbapenemase genes.24 Extended spectrum β-lactamases (ESBLs), porin mutations and/or presence of efflux pumps are other carbapenem resistance mechanisms that were implicated in carbapenem resistance in the absence of carbapenemase genes in tested isolates.25
In CR A. baumannii isolates, blaOXA-23 was the most frequent carbapenemase gene and the only OXA-type carbapenemase detected in 27/30 (90%), followed by blaNDM in 20/30 (66.7%) then blaGES in 15/30 (50%). Kateete et al, in another African study, reported blaOXA-23 60% (9/15) as the most frequent OXA-type carbapenemase gene detected in CR A. baumannii followed by blaOXA-58 (2/15, 13%) and blaOXA-24 (1/15, 7%), while blaVIM was the only metallo-β-lactamase detected in 2/15 (13%) in such isolates.26 The blaKPC gene, the metallo-β-lactamases (VIM, IMP) genes as well as other tested OXA-type genes were not detected in our CR A. baumannii isolates.
A. baumannii strains harboring blaOXA-23 have been reported to emerge around the world replacing blaOXA-58 that predominated for a long period among CR A. baumannii isolates and became the most prevalent carbapenemase gene circulating in different Mediterranean countries including Egypt.27,28 The higher carbapenemase activity of OXA-23 and/or horizontal gene transfer might provide a selective advantage for such isolates.29
KPCs and GES-type class A carbapenemases have been described in A. baumannii.30 Only GES-type carbapenemase was reported in Mediterranean countries. In addition, a Turkish study found that A. baumannii strains that belong to ST2, a part of the worldwide distributed clone II group, co-harbor both blaOXA-23 and blaGES-11.31 This applies to the current study where blaGES, and not blaKPC, was detected and coexisted with blaOXA-23 in 12/30 (40%) of CR A. baumannii isolates.
In Egypt NDM-1 and NDM-2 encoding genes have been reported since 2011.32,33 Another Egyptian study identified MBL-encoding genes blaVIM and blaNDM-1 in 86.4% and 59.1% of MBL-producing A. baumannii isolates, respectively.34
In CR P. aeruginosa isolates, blaGES was detected in 40.9% of the isolates (9/22). Metallo-β-lactamase (MBL) genes were detected in 18/22 isolates including blaIMP (18.2%, 4/22), blaVIM (50%, 11/22), and blaNDM (27.3%, 6/22). In a study in Saudi Arabia, two MBL types, VIM and/or IMP, were detected in 12/34 CR P. aeruginosa isolates.24 In Uganda, metallo-β-lactamase genes detected in CR P. aeruginosa were blaIMP1 (36%, 9/25), blaIMP2 (4%, 1/25), blaVIM1 (32%, 8/25), and blaNDM1 (4%, 1/25).26
The KPC enzyme-encoding gene (blaKPC), known to be prevalent among Enterobacteriaceae, was not detected in our isolates but was identified in Pseudomonas (16.9%) and Acinetobacter spp. (11.9%) from another Egyptian study.35
In the past few years, antibiotic combination therapy has been employed as the standard for treating severe infections by CR GNB, particularly those with an XDR profile, justified by the possible additive or synergistic effect of adding drugs and by the broad spectrum activity that suppresses the emergence of resistant subpopulations if one antibiotic is used.36 However, the irrational use of antibiotic combinations can worsen the existing antibiotic resistance problem. Thus, antibiotic combination should be rationally optimized based on preliminary in-vitro or animal studies followed by clinical trials on human subjects. Experts further recommend that combinations should be individualized based on the agent involved, its antibiotic susceptibility pattern, the type of infection and the patient’s characteristics.13
So far, most CR GNB are susceptible to colistin. In the present study all CR A. baumannii and 90.9% of CR P. aeruginosa isolates were colistin-sensitive and this nominates colistin to be a cornerstone drug in combination therapy. In CR A. baumannii, the colistin/meropenem combination showed additive effects in most (83.3%) of the isolates and indifferent effects in only 16.7%. This agrees with a recent Lebanese study where ‘Perpendicular E-tests’ showed additive effects of the colistin/meropenem combination in A. baumannii. Synergy, however, was not detected in either studies but a reduction of colistin MIC in combination means lower doses of colistin can be used in therapy, thus minimizing its toxic effects.16 In CR P. aeruginosa, the colistin/meropenem combination showed better results where synergistic and additive effects were detected in 63.6% and 36.4% of the isolates, respectively. Daoud et al on testing this combination, found that six strains (54.5%) of A. baumannii and three (27.3%) strains of P. aeruginosa showed synergy while five (45.5%) strains of A. baumannii and seven (63.3%) strains of P. aeruginosa showed additive effect, while only one (9%) strain of P. aeruginosa showed antagonism.37
On the other hand, the colistin/tigecycline combination against CR A. baumannii isolates showed synergistic, additive and indifferent effects in six (20%), 18 (60%) and six (20%), respectively. Karaoglan et al found that 96% and 64% of CR A. baumannii isolates were colistin- and tigecycline-sensitive, respectively. Meanwhile, the colistin/tigecycline combination revealed synergistic effects in 12% of such isolates.38 Ni et al in a study on MDR A. baumannii strains, reported that the colistin/tigecycline combination had a synergistic effect in 24.3%.39 In contrast, a recent Turkish study found colistin/tigecycline combination synergistic in 2%, additive in 6%, indifferent in 88%, and antagonistic in 4% of A. baumannii isolates; all were tigecycline- and colistin-sensitive, concluding that the colistin/tigecycline combination is not an optimal treatment choice.40
The synergistic or additive outcome of the colistin-based combination could be due to its ability to disrupt the bacterial outer membrane which facilitates the entry of the other agents into the bacterial cell. In addition, discrepant results of antibiotic combinations in different studies could be due to the different antibiotic selective pressure and genetic makeup of the isolates leading to their differential adaptation in different environments and consequently difference in their response to antimicrobial agents.16
Conclusion
High level carbapenem resistance detected in CR A. baumannii and CR P. aeruginosa in this study is attributed to the carriage of carbapenemase genes in different combinations. Colistin/tigecycline and colistin/meropenem combinations can be considered for treatment of severe infections by CR A. baumannii and P. aeruginosa, respectively, after adequate review of relevant pharmacodynamic and pharmacokinetic data. Moreover, the in vivo efficacy of these combinations needs to be proved by further clinical studies.
Limitations of the study
Primers used in this study amplify all alleles of the GES gene. So, a sequencing step would be helpful to differentiate between alleles of GES gene with ESBL and carbapenemase activities.
Disclosure
The authors report no conflicts of interest in this work.
References
Papp-Wallace KM, Endimiani A, Taracila MA, Bonomo RA. Carbapenems: past, present, and future. Antimicrob Agents Chemother. 2011;55(11):4943–4960. | ||
El-Herte RI, Kanj SS, Matar GM, Araj GF. The threat of carbapenem-resistant Enterobacteriaceae in Lebanon: an update on the regional and local epidemiology. J Infect Public Health. 2012;5(3):233–243. | ||
Walsh TR. Emerging carbapenemases: a global perspective. Int J Antimicrob Agents. 2010;36(Suppl. 3):S8–S14. | ||
Zilberberg MD, Nathanson BH, Sulham K, Fan W, Shorr AF. Multidrug resistance, inappropriate empiric therapy, and hospital mortality in Acinetobacter baumannii pneumonia and sepsis. Crit Care. 2016;20(1):1–10. | ||
Monteiro J, Widen RH, Pignatari AC, Kubasek C, Silbert S. Rapid detection of carbapenemase genes by multiplex real-time PCR. J Antimicrob Chemother. 2012;67(4):906–909. | ||
Shahid M, Sobia F, Singh A, Khan HM. Molecular epidemiology of carbapenem-resistant Enterobacteriaceae from a North Indian Tertiary Hospital. N Z J Med Lab Sci. 2012;66:5–7. | ||
Sarı AN, Biçmen M, Gülay Z. The first report on the outbreak of OXA-24/40-like carbapenemase-producing Acinetobacter baumannii in Turkey. Jpn J Infect Dis. 2013;66(5):439–442. | ||
Potron A, Poirel L, Nordmann P. Emerging broad-spectrum resistance in Pseudomonas aeruginosa and Acinetobacter baumannii: mechanisms and epidemiology. Int J Antimicrob Agents. 2015;45(6):568–585. | ||
Magiorakos AP, Srinivasan A, Carey RB, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–281. | ||
Zavascki AP, Carvalhaes CG, Picão RC, Gales AC. Multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii: resistance mechanisms and implications for therapy. Expert Rev Anti Infect Ther. 2010;8(1):71–93. | ||
Bergen PJ, Landersdorfer CB, Lee HJ, Li J, Nation RL. ‘Old’ antibiotics for emerging multidrug-resistant bacteria. Curr Opin Infect Dis. 2012;25(6):626–633. | ||
Hasanin A, Mukhtar A, El-Adawy A, et al. Ventilator associated pneumonia caused by extensive-drug resistant Acinetobacter species: colistin is the remaining choice. Egypt J Anaesth. 2016;32(3):409–413. | ||
Zavascki AP, Bulitta JB, Landersdorfer CB. Combination therapy for carbapenem-resistant Gram-negative bacteria. Expert Rev Anti Infect Ther. 2013;11(12):1333–1353. | ||
Bonapace CR, White RL, Friedrich LV, Bosso JA. Evaluation of antibiotic synergy against Acinetobacter baumannii: a comparison with Etest, time-kill, and checkerboard methods. Diagn Microbiol Infect Dis. 2000;38(1):43–50. | ||
Cetin ES, Tekeli A, Ozseven AG, Us E, Aridogan BC. Determination of in vitro activities of polymyxin B and rifampin in combination with ampicillin/sulbactam or cefoperazone/sulbactam against multidrug-resistant Acinetobacter baumannii by the E-test and checkerboard methods. Jpn J Infect Dis. 2013;66(6):463–468. | ||
Soudeiha MAH, Dahdouh EA, Azar E, Sarkis DK, Daoud Z. In vitro evaluation of the colistin-carbapenem combination in clinical isolates of A. baumannii using the checkerboard, Etest, and time-kill curve techniques. Front Cell Infect Microbiol. 2017;7:1–10. | ||
Woodford N, Ellington MJ, Coelho JM, et al. Multiplex PCR for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int J Antimicrob Agents. 2006;27(4):351–353. | ||
Higgins PG, Lehmann M, Seifert H. Inclusion of OXA-143 primers in a multiplex polymerase chain reaction (PCR) for genes encoding prevalent OXA carbapenemases in Acinetobacter spp. Int J Antimicrob Agents. 2010;35(3):305. | ||
Poirel L, Walsh TR, Cuvillier V, Nordmann P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn Microbiol Infect Dis. 2011;70(1):119–123. | ||
Clinical and Laboratory Standards Institute. M100 Performance Standards for Antimicrobial Susceptibility Testing. 27th ed; 2017: 224 p. | ||
Yavas¸ S, Yetkın MA, Kayaaslan B, et al. Investigating the in vitro synergistic activities of several antibiotic combinations against carbapenem-resistant Acinetobacter baumannii isolates. Turk J Med Sci. 2016;46(3):892–896. | ||
Sueke H, Kaye SB, Neal T, Hall A, Tuft S, Parry CM. An in vitro investigation of synergy or antagonism between antimicrobial combinations against isolates from bacterial keratitis. Invest Ophthalmol Vis Sci. 2010;51(8):4151–4155. | ||
Pillai SK, Moellering RC, Eliopoulos GM. Antibiotics in laboratory medicine. In: Lorian V, editor. Antibiotics in Laboratory Medicine, 5th edn. Baltimore, MD: Lippincott Williams & Wilkins; 2005:365–440. | ||
Al-Agamy MH, Jeannot K, El-Mahdy TS, et al. Diversity of molecular mechanisms conferring carbapenem resistance to Pseudomonas aeruginosa isolates from Saudi Arabia. Can J Infect Dis Med Microbiol. 2016;2016:4379686. | ||
Chambers H. Carbapenems and monobactams. In: Mandell GL, Bennett JE, Dolin R, editors. Mandell, Douglas and Bennett’s Principles and Practice of Infectious Diseases. 7th edn. Philadelphia, PA: Elsevier Publishers; 2007:341–346. | ||
Kateete DP, Nakanjako R, Namugenyi J, Erume J, Joloba ML, Najjuka CF. Carbapenem resistant Pseudomonas aeruginosa and Acinetobacter baumannii at Mulago Hospital in Kampala, Uganda (2007–2009). Springerplus. 2016;5(1):1308. | ||
Mugnier PD, Poirel L, Naas T, Nordmann P. Worldwide dissemination of the blaOXA-23 carbapenemase gene of Acinetobacter baumannii. Emerg Infect Dis. 2010;16(1):35–40. | ||
Fouad M, Attia AS, Tawakkol WM, Hashem AM. Emergence of carbapenem-resistant Acinetobacter baumannii harboring the OXA-23 carbapenemase in intensive care units of Egyptian hospitals. Int J Infect Dis. 2013;17(12):e1252–e1254. | ||
Djahmi N, Dunyach-Remy C, Pantel A, Dekhil M, Sotto A, Lavigne J-P. Epidemiology of carbapenemase-producing Enterobacteriaceae and Acinetobacter baumannii in Mediterranean countries. Biomed Res Int. 2014;2014(6):11 p–11. | ||
Zarrilli R, Pournaras S, Giannouli M, Tsakris A. Global evolution of multidrug-resistant Acinetobacter baumannii clonal lineages. Int J Antimicrob Agents. 2013;41(1):11–19. | ||
Zeka AN, Poirel L, Sipahi OR, et al. GES-type and OXA-23 carbapenemase-producing Acinetobacter baumannii in Turkey. J Antimicrob Chemother. 2014;69(4):1145–1146. | ||
Hrabák J, Stolbová M, Studentová V, Fridrichová M, Chudácˇková E, Zemlickova H. NDM-1 producing Acinetobacter baumannii isolated from a patient repatriated to the Czech Republic from Egypt, July 2011. Euro Surveill. 2012;17(7):7–9. | ||
Kaase M, Nordmann P, Wichelhaus TA, Gatermann SG, Bonnin RA, Poirel L. NDM-2 carbapenemase in Acinetobacter baumannii from Egypt. J Antimicrob Chemother. 2011;66(6):1260–1262. | ||
Gomaa FAM, Helal ZH, Khan MI. High Prevalence of blaNDM-1, blaVIM, qacE, and qacEΔ1 genes and their association with decreased susceptibility to antibiotics and common hospital biocides in clinical isolates of Acinetobacter baumannii. Microorganisms. 2017;5(2):18. | ||
Fattouh M, Nasr El-Din A, Omar MA. Detection of Klebsiella pneumoniae carbapenemase (KPC) producing gram negative superbugs: an emerging cause of multidrug-resistant infections in General Surgery Department of Sohag University Hospital, Egypt. Int J Curr Microbiol App Sci. 2015;4(5):1–15. | ||
Ahmed A, Azim A, Gurjar M, Baronia AK. Current concepts in combination antibiotic therapy for critically ill patients. Indian J Crit Care Med. 2014;18(5):310–314. | ||
Daoud Z, Mansour N, Masri K. Synergistic combination of carbapenems and colistin against P. aeruginosa and A. baumannii. Open J Med Microbiol. 2013;03(04):253–258. | ||
Karaoglan I, Zer Y, Bosnak VK, Mete AO, Namiduru M. In vitro synergistic activity of colistin with tigecycline or β-lactam antibiotic/β-lactamase inhibitor combinations against carbapenem-resistant Acinetobacter baumannii. J Int Med Res. 2013;41(6):1830–1837. | ||
Ni W, Cui J, Liang B, et al. In vitro effects of tigecycline in combination with colistin (polymyxin E) and sulbactam against multidrug-resistant Acinetobacter baumannii. J Antibiot. 2013;66(12):705–708. | ||
Kaya IA, Guner MD, Akca G, Tuncbilek S, Alhan A, Tekeli E. Evaluation of the synergistic effect of a combination of colistin and tigecycline against multidrug-resistant Acinetobacter baumannii. Pakistan J Med Sci. 2017;33(2):393–397. |
Supplementary materials
 | Table S1 Individual MIC values of tested agents, FIC values of tested combinations and corresponding carbapenemase gene/s for all Acinetobacter baumannii isolates Abbreviations: FIC, fractional inhibitory concentration; MIC, minimum inhibitory concentration; CL, colistin; MEM, meropenem; TGC, tigecycline. |
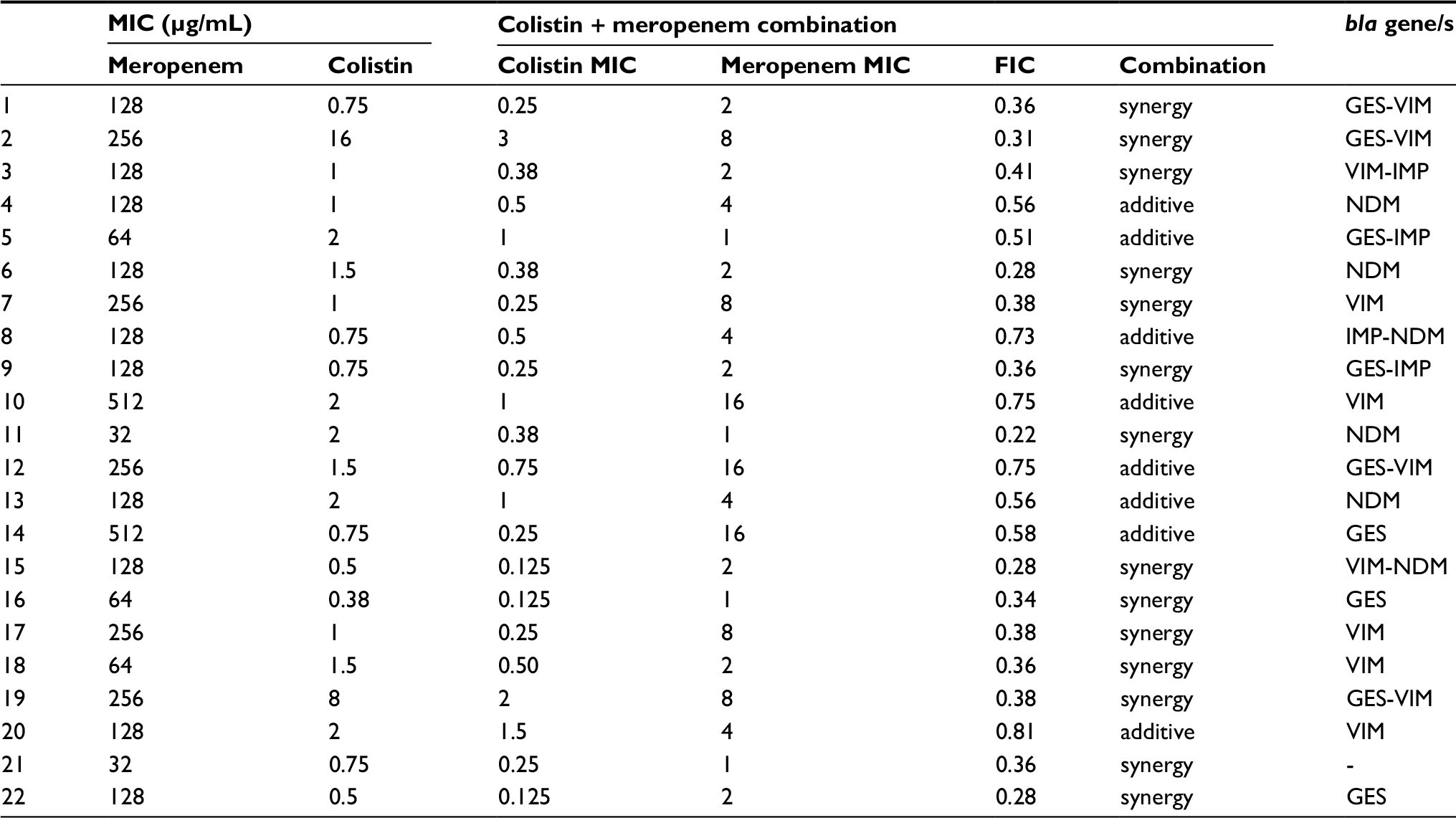 | Table S2 Individual MIC values of tested agents, FIC values of tested combination and corresponding carbapenemase gene/s for all Pseudomonas aeruginosa isolates Notes: Meropenem MIC values >32 µg/mL (highest concentration on E-test strip) were calculated as 64 µg/mL (two-fold dilution above highest concentration on E-test strip). Abbreviations: FIC, fractional inhibitory concentration; MIC, minimum inhibitory concentration. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.