Back to Journals » Diabetes, Metabolic Syndrome and Obesity » Volume 16
Can Antidiabetic Medications Affect Telomere Length in Patients with Type 2 Diabetes? A Mini-Review
Authors Qin B ![]()
Received 2 July 2023
Accepted for publication 7 October 2023
Published 22 November 2023 Volume 2023:16 Pages 3739—3750
DOI https://doi.org/10.2147/DMSO.S428560
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Konstantinos Tziomalos
Baoding Qin
Department of Endocrinology, the Third Affiliated Hospital of Sun Yat-sen University, Guangzhou, Guangdong, People’s Republic of China
Correspondence: Baoding Qin, The Third Affiliated Hospital of Sun Yat-sen University, 600 Tianhe Road, Tianhe District, Guangzhou, Guangdong, People’s Republic of China, Email [email protected]
Abstract: The fight against aging is an eternal pursuit of humankind. The aging rate of patients with type 2 diabetes mellitus (T2DM) is higher than that of healthy individuals. Reducing the aging rate of patients with T2DM and extending their life expectancy are challenges that endocrinologists are eager to overcome. Many studies have shown that antidiabetic medications have potent anti-aging potential. Telomeres are repetitive DNA sequences located at the ends of chromosomes, and telomere shortening is a hallmark of aging. This review summarizes clinical trials that have explored the association between antidiabetic medications and telomere length (TL) in patients with T2DM and explore the mystery of delaying aging in patients with T2DM from the perspective of telomeres. Various antidiabetic medications may have different effects on TL in patients with T2DM. Metformin and sitagliptin may protect telomeres in patients with T2DM, while exogenous insulin may promote telomere shortening in patients with T2DM. The effect of acarbose and glyburide on TL in patients with T2DM is still uncertain due to the absence of evidence from longitudinal studies.
Keywords: type 2 diabetes, antidiabetic medications, telomere length, aging
Introduction
Recent studies have shown that approximately 536.6 million adults worldwide have diabetes mellitus (DM), and its prevalence is increasing worldwide.1,2 DM and its complications seriously endanger the lives and health of patients. In 2019, 6.6 million disability-adjusted life years (DALYs) were associated with T2DM. Furthermore, the annual growth rate of age-standardized DALYs for T2DM was 0.77% from 2000 to 2019.3
Many factors affect human longevity, and telomeres drive the aging process and promote the development of aging-related diseases such as T2DM.4 Telomeres consist of tandem repeat nucleotide sequences 5 “-TTAGGG-3” n at eukaryotic chromosomes’ ends5 and are protective structures that maintain chromosomal integrity.4,5 The process of cell division is accompanied by the shortening of TL, which triggers the Hayflick limit when TL is sufficiently short, and the cell begins to senesce.5
TL shortening is a hallmark of aging6 and is associated with other hallmarks of aging, including cell senescence, stem cell exhaustion, genomic instability, mitochondrial dysfunction, epigenetic dysregulation, loss of protease homeostasis, altered nutrient perception, and inflammation.4 Telomere dysfunction can amplify these mechanisms and promote the occurrence and development of aging and aging-related diseases,4 thereby affecting human lifespans. In 2023, Lo´pez-Otı´n introduced 3 additional hallmarks of aging: disabled macroautophagy, chronic inflammation, and dysbiosis.6 Whether telomeres are associated with these 3 hallmarks and thus have a specific impact on the aging process needs to be further explored.
Telomere shortening is associated with DM and its complications. Specifically, hyperglycemia can increase the oxidative stress level, promoting single-stranded telomere breaks, thereby accelerating telomere shortening.5,7 The level of oxidative stress in islet β cells of patients with T2DM is increased, leading to shorter TL of β cells and reduced insulin secretion.8 A meta-analysis9 involving 9 cohorts (5759 cases and 6518 controls) showed a strong association between shortened TL and T2DM risk. Moreover, a meta-analysis10 of 17 cohorts (5575 cases and 6349 controls) showed that TL was shorter in patients with DM than in non-DM individuals. Relative leukocyte telomere length (rLTL) may also be a predictor of the glycemic progression of T2DM, and rLTL shortening is associated with glycemic progression in T2DM.11 As the disease progresses, patients with T2DM gradually develop various complications.12 Several studies have shown that complications of T2DM are closely related to TL (Table 1). Several scholars have also demonstrated that shorter rLTL is associated with higher all-cause mortality in patients with T2DM.13 Verma et al reported that DM may affect the health and life span of patients with DM by promoting telomere shortening.7
 |
Table 1 Association Between Complications of T2DM and TL |
The anti-aging effect of antidiabetic medications such as metformin has attracted increasing attention. Metformin has a significant cardiovascular protective effect and reduces the incidence of geriatric diseases, the risk of cancer, and all-cause mortality.21,22 Some researchers believe that metformin can delay the progression of aging and aging-related diseases by targeting the key hallmarks of aging.23,24 Studies showing that metformin prolongs the lifespan of mice and Caenorhabditis elegans further support its anti-aging effect.25,26
Multiple clinical studies have reported that a variety of antidiabetic medications, such as sitagliptin, acarbose, glyburide, exogenous insulin, and metformin, are closely related to TL in patients with T2DM (Table 2 and Table 3), indicating that antidiabetic medications may affect the aging process of patients with T2DM. This review summarizes the clinical studies on the relationship between antidiabetic medications and TL in patients with T2DM, hoping to explore the possibility from the camera shots of telomere that antidiabetic medications delay aging, prolong life span, and reduce mortality of patients with T2DM.
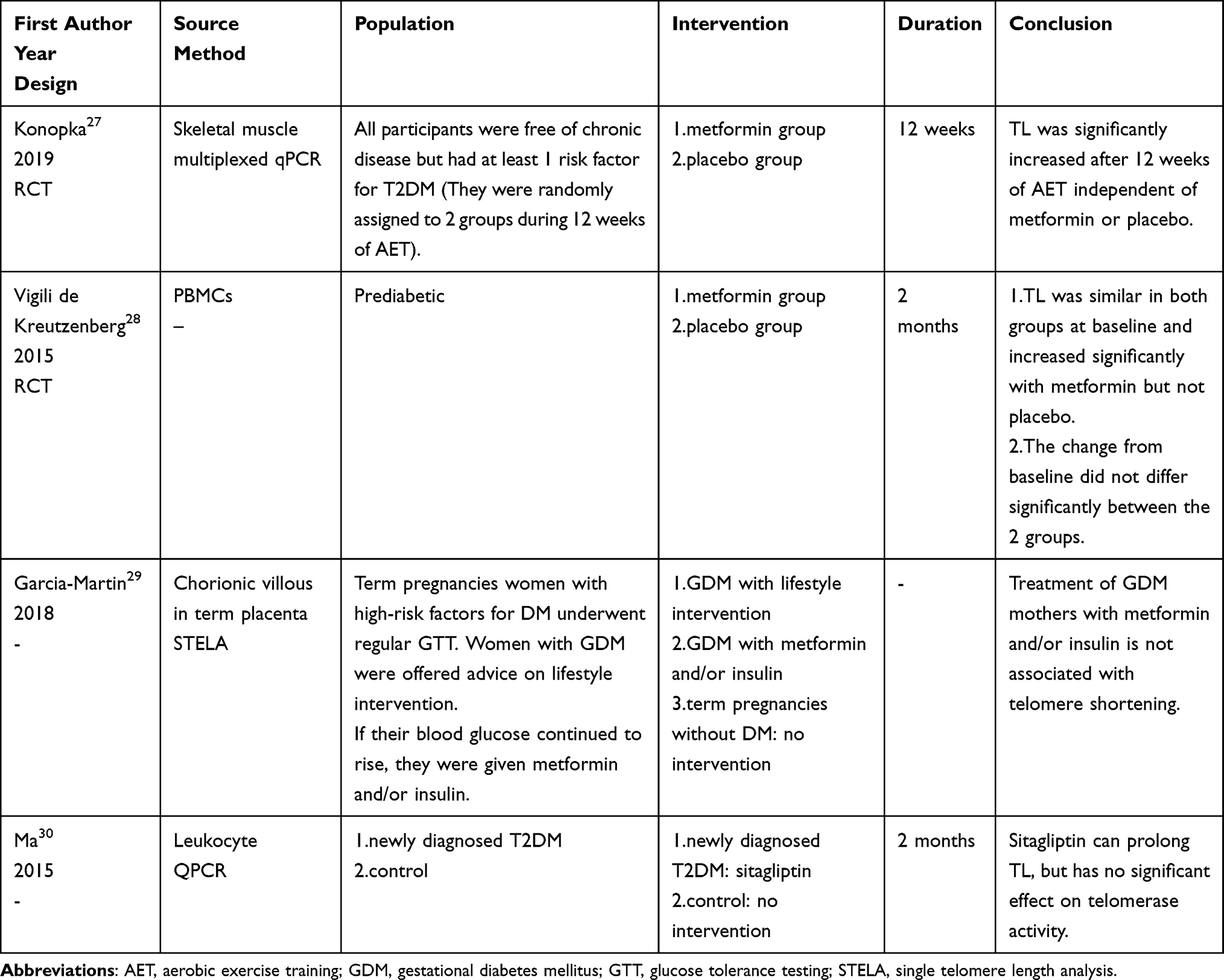 |
Table 2 Effect of Antidiabetic Medications on TL in Patients with T2DM (Intervention Studies) |
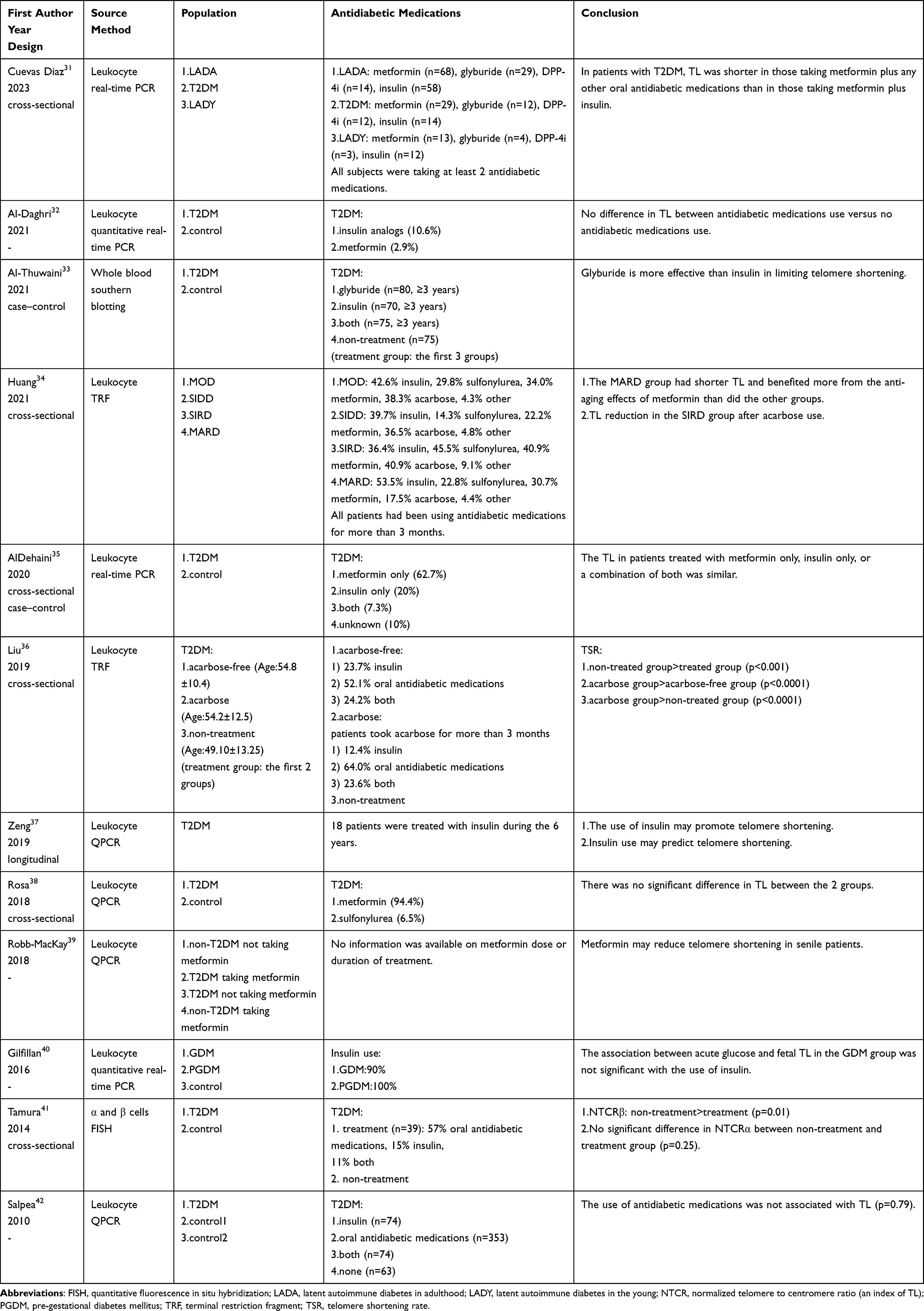 |
Table 3 Effect of Antidiabetic Medications on TL in Patients with T2DM (Non-Interventional Studies) |
Materials and Methods
PubMed and Embase were searched for literature published before May 16, 2023.
The PubMed retrieval strategy was as follows: 1. (“Hypoglycemic Agents” [Mesh] OR “Hypoglycemic Agents” [Pharmacological Action]) AND (“Telomere” [Mesh] OR “Telomere Shortening” [Mesh] OR “Telomere Homeostasis” [Mesh] OR “Telomerase” [Mesh]), 2. The generic names of all common antidiabetic medications and the word “telomere” were connected with “and” respectively, and free-word retrieval was performed.
The Embase retrieval strategy was as follows: (“telomere”/exp OR “telomere length”/exp OR “telomere shortening”/exp OR ‘telomere homeostasis’/exp) AND “antidiabetic agent”/exp.
The literature inclusion criterion was: clinical trial involving patients with T2DM. The literature exclusion criteria were: 1. The main study did not include antidiabetic medications, 2. The main study involved antidiabetic medications; however, no correlation analysis was performed between antidiabetic medications use and TL. One dissertation was found from the references of a review. One randomized controlled trial (RCT) involving patients with prediabetic was included because of its classical design.
Sixteen clinical trials were included in the review (Table 2 and Table 3).
α-Glucosidase Inhibitors
Acarbose is an α-glucosidase inhibitor that controls blood glucose levels by inhibiting carbohydrate digestion in the small intestine.43 Two related studies34,36 showed that the use of acarbose is associated with leukocyte telomere shortening in patients with T2DM independent of glycemic control.
Liu et al36 conducted a cross-sectional analysis of 388 patients with T2DM, showing that among patients who received antidiabetic medications, those taking acarbose for more than 3 months had a higher telomere shortening rate (TSR) than did those not taking acarbose (TSR: −22.14 ± 4.66 bp/year vs −9.29 ± 4.30 bp/year, p<0.0001); furthermore, the glycated hemoglobin (HbA1c), age and gender were similar between the 2 groups. Patients taking acarbose for more than 3 months had a higher TSR than did those not taking antidiabetic medications (TSR: −22.14 ± 4.66 bp/year vs −15.74 ± 6.21 bp/year, p<0.0001) after adjustment for age, and the acarbose group had lower HbA1c (HbA1c: 8.64 ± 2.38% vs 10.57 ± 2.86%, p=0.000), higher age (age: 54.2 ± 12.5 years vs 49.10 ± 13.25 years, p=0.011) and similar sex ratio. This finding suggests that the correlation between the use of acarbose and higher TSR in patients with T2DM is independent of glycemic control.
Huang et al34 analyzed 246 patients with T2DM and classified them as mild obesity-related diabetes (MOD) group, severe insulin-deficient diabetes (SIDD) group, severe insulin-resistant diabetes (SIRD) group or mild age-related diabetes (MARD) group. The 4 groups were statistically different in age (age: MOD: 39.53 ± 9.01 years, SIDD: 37.81 ± 10.65 years, SIRD: 49.45 ± 14.54 years, MARD: 52.28 ± 9.23 years, p<0.001) and had similar sex ratio. The results revealed that among the SIRD group, patients who used acarbose exhibited shorter leukocyte TL compared to those who did not use acarbose (p<0.001). However, in the other 3 subgroups, there was no significant difference in leukocyte TL observed between patients who used acarbose and those who did not use acarbose within each subgroup. Linear regression analysis revealed a significant correlation between the use of acarbose and the leukocyte TL of the SIRD group, even after adjusting for the age and gender of the subjects (r = −0.069, 95% CI: −0.100 to −0.039, p < 0.001), unlike the other 3 groups (p > 0.05). The leukocyte TL was shorter in the MARD group than in the non-MARD group (p=0.0012), but there was no significant difference between the 2 groups after acarbose use. Comparisons of glycemic control among patients using different antidiabetic medications within each subgroup are not provided.
However, the mechanism by which acarbose use is associated with shorter leukocyte TL in patients with T2DM is unclear. Both intestinal flora and oxidative stress hypotheses suggest that acarbose may have anti-aging effects.
Gut Microbiota
The human gut microbiota, composed of bacteria, fungi, viruses, and protozoa that colonize the human gut system, profoundly impacts age-related diseases and aging and can even be an intervention target to delay aging.44 Relevant animal experiments45 have shown that alterations in the gut microbiota can significantly improve aging. A systematic review46 that included 27 studies found significant differences in the gut microbiota among oldest-old, younger-old, and younger adults, suggesting that gut microbiota composition may have an impact on aging and longevity.
Acarbose profoundly affects the gut ecosystem as it can increase the relative abundance of Bifidobacterium and Lactobacillus and reduce the relative abundance of Bacteroides in the intestinal microbiota of patients with T2DM.47 Animal experiments have shown that Bifidobacterium and Lactobacillus have anti-aging effects, reducing the expression of aging biomarkers in mice and improving aging performance, such as cognitive ability and physiological function in older mice.48 Given that these are potential anti-aging bacteria, it is reasonable to assume that acarbose may delay aging in patients with T2DM by increasing the relative abundance of Bifidobacterium and Lactobacillus in the intestine. Animal studies have shown that Lactobacillus can prevent telomere shortening in aging mice.49,50
Oxidative Stress
Oxidative stress refers to an imbalance between reactive oxygen species (ROS) and cellular antioxidant defense, producing excessive ROS and other oxidative substances that can regulate cell signaling pathways and cause cell damage.7,51 Oxidative stress is the main cause of telomere shortening in patients.7 Hyperglycemia can elevate the level of oxidative stress in patients with DM by increasing ROS production in the electron transport chain of the mitochondria.7 In addition, the production of advanced glycation end products, enhanced glucose auto-oxidation, activation of protein kinase (PKC), and activation of the polyol pathway enhance oxidative stress.7 Telomeres are located at the ends of chromosomes and are rich in guanine base pairs that are easily oxidized,5,52 exist as a single strand during mitosis, and are highly susceptible to oxidative stress and single-strand breaks, leading to telomere shortening.5,52
One of the important features of T2DM is insulin resistance (IR).53 Oxidative stress is one of the key factors in inducing IR, and in turn, IR can increase the level of oxidative stress through different mechanisms; therefore, it can be postulated that oxidative stress and IR are mutually causal, leading to a vicious circle.51 Postprandial hyperglycemia, a characteristic of IR, increases oxidative stress in patients with T2DM, whereas acarbose reduces oxidative stress in patients with T2DM by reducing postprandial hyperglycemia.54 Notably, acarbose can also improve IR in patients with T2DM.54 Feeding sucrose-containing food to Zucker rats with T2DM resulted in elevated blood glucose, plasma insulin, and oxidative stress markers, whereas the acarbose intervention largely prevented the increase in oxidative stress markers in these rats, suggesting that acarbose can reduce hyperglycemia-induced oxidative stress.55
However, it is important to note that the 2 hypotheses mentioned above contradict current clinical research findings that acarbose may be related to telomere shortening in patients with T2DM. More prospective, large-sample clinical studies and mechanism-based studies are needed to explore the causal relationship between acarbose and TL.
Dipeptidyl Peptidase-4 Inhibitor
Sitagliptin is a dipeptidyl peptidase-4 inhibitor (DPP-4i) that reduces the inactivation of the endogenous incretin hormones glucagon-like peptide 1 (GLP-1) and glucose-dependent insulinotropic polypeptide (GIP) by inhibiting DPP-4, thereby controlling blood glucose.56 GLP-1 protects cells from apoptosis and promotes β-cell survival.57
In a clinical trial,30 38 patients with newly diagnosed T2DM were treated with sitagliptin for 2 months. After 2 months, their fasting plasma glucose (FPG) (FPG: baseline 11.51 ± 2.32 mmol/L, 1 month 9.78 ± 1.87 mmol/L, 2 months 7.17 ± 2.05 mmol/L, p<0.001) and fasting plasma insulin (FINS) (FINS: baseline 47.58 ± 28.04 mIU/L, 1 month 32.84 ± 21.14 mIU/L, 2 months 18.86 ± 10.64 mIU/L, p<0.001) were significantly lower than those at baseline, the leukocyte TL was significantly longer than that at baseline (telomere length ratio: baseline 1.58 ± 0.57, 1 month 2.51 ± 0.62, 2 months 3.95 ± 0.92, p<0.001). This interventional study suggested that sitagliptin may prolong TL in patients with T2DM.
Khalangot et al58 found that leukocyte TL was negatively related to FPG and 2 h post-load plasma glucose (2hPG) levels in patients with T2DM. That is, blood glucose levels can affect leukocyte TL in patients with T2DM. Ma et al30 suggested that reduced oxidative stress due to good glycemic control is the main reason sitagliptin prolongs leukocyte TL and that sitagliptin may protect cells from destruction and delay the progression of T2DM by prolonging TL.
Telomerase, a ribonucleoprotein complex with reverse transcription functions, plays a key role in maintaining the health and reproduction of species by adding telomeric repeats to the ends of chromosomes to maintain chromosomal integrity.4,5 Preventing telomere erosion is the main function of telomerase.59 Few clinical studies have explored the relationship between antidiabetic medications and telomerase. This study30 analyzed whether sitagliptin affected telomerase activity in patients with T2DM, and the results showed that there was no significant change in telomerase activity before and after sitagliptin intervention (telomerase activity: baseline 0.41 ± 0.22, 1 month 0.42 ± 0.22, 2 months 0.40 ± 0.21, p=0.830), suggesting that sitagliptin may not affect leukocyte TL changes in patients with T2DM by affecting telomerase activity.
Sulfonylurea
Sulfonylureas are second-line antidiabetic medications for T2DM that control blood glucose levels by stimulating β-cells to promote insulin secretion.60 Glyburide is a second-generation sulfonylurea and one of the most commonly used sulfonylureas.33,60
Al-Thuwaini designed a case-control study33 that involved 100 middle-aged individuals without DM and 300 middle-aged patients with T2DM (75 non-treatment patients, 80 using glyburide, 70 using insulin and 75 with both therapies, patients in the latter 3 groups had a history of using antidiabetic medications for more than 3 years). Logistic regression analysis showed that glyburide may have a protective effect on telomeres in patients with T2DM (estimate: 2.42, odds ratio (OR): 2.07, 95% CI: 1.13 to 2.90, p=0.004).
In brief, sulfonylurea drugs bind to sulfonylurea receptors, causing the closure of ATP-depended potassium channels (KATP), which in turn leads to depolarization of β cells. This depolarization results in an increase in intracellular calcium concentration, which promotes endogenous insulin release and exerts glucose-lowering effects.61,62 Al-Thuwaini et al suggested that the prolongation of TL in patients with T2DM with glyburide may be caused by a decrease in oxidative stress levels due to good glycemic control.33 As mentioned earlier, oxidative stress induced by hyperglycemia has been found to accelerate telomere shortening.7 Additionally, some scholars have highlighted that oxidative stress can inhibit the activity of telomerase.63 Glyburide, by promoting endogenous insulin secretion and lowering glucose levels, can effectively reduce oxidative stress.33 As a result, it may help safeguard TL.33
Exogenous Insulin
Insulin therapy is an essential life-saving measure for patients with severe T2DM; however, insulin also has the potential to accelerate telomere shortening in patients with T2DM. In a 6-year longitudinal study37 investigating the effect of insulin on leukocyte TL, 76 patients with T2DM were enrolled, and 64 patients were successfully followed up. Insulin therapy was added to the original antidiabetic regimen for patients with HbA1c > 7.5%, despite taking 3 oral antidiabetic medications. Finally, 18 patients received insulin therapy for poor glycemic control. Of the 64 successfully followed up patients, telomere shortening was found in 45 patients (including 16 insulin-using patients) and telomere lengthening in 19 patients (including 2 insulin-using patients) compared to baseline. Multivariate regression analysis suggested that insulin use was the only independent predictor of leukocyte TL change (β-coefficients: −0.587, 95% CI: −0.198 to −0.085, p < 0.001). Logistic regression analysis showed that only insulin use (OR: 17.355, 95% CI: 2.659 to 35.627, p=0.013) and LDL-C level (OR: 3.493, 95% CI: 1.599 to 10.063, p=0.007) were independent predictors of telomere shortening. The aforementioned case-control study33 found that telomere shortening was 3 times higher in insulin-treated patients with T2DM (logistic regression analysis, estimate: −3.41, p=0.001). A cross-sectional study35 by AlDehaini et al found no significant difference in leukocyte TL among patients treated with metformin alone, insulin alone, or a combination of the both. There are 2 possible reasons for the effects of insulin on TL.
Glycemic Progression
In the longitudinal study37 mentioned above, it was observed that subjects who used exogenous insulin experienced telomere shortening. However, it is important to note that these subjects were also using multiple antidiabetic medications and HbA1c>7.5% at the beginning of the study, indicating that they had a progression of glycemic levels. Telomere shortening is recognized as a biomarker of glycemic progression.11 Therefore, it is plausible to suggest that the telomere shortening observed in subjects using exogenous insulin may not be directly caused by the insulin itself, but rather by the progression of their own glycemic levels.
Weight Gain
Weight gain is an adverse effect of insulin. A retrospective cohort study64 involving 5086 patients with T2DM using insulin suggested that some patients gained 5 kg in weight in the first few years after treatment with insulin. Weight gain is one of the main causes of IR. A study65 involving 1758 middle-aged obese individuals found that a weight gain of 5% in adults was strongly associated with IR. It is reasonable to assume that insulin use may lead to weight gain, which in turn leads to the exacerbation of IR, promotes elevated levels of oxidative stress, and finally leads to telomere shortening.
Biguanides
Metformin is the first-line treatment for T2DM and is recognized as a potent anti-aging drug. Clinical studies suggested that biguanides may prolong TL in patients with T2DM.
Huang et al defined the MARD group in a study34 as older patients with T2DM suffering from moderate metabolic disturbances. This study found that leukocyte TL was significantly shorter in the MARD group than in the non-MARD group (p=0.0012); however, there was no difference in leukocyte TL between the MARD and non-MARD groups under metformin treatment. In the MARD group, a longer leukocyte TL was found in patients using metformin than in those who did not (p<0.001). Linear regression analysis revealed a significant correlation between the use of metformin and the leukocyte TL of the MARD group, even after adjusting for the age and gender of the subjects (r=0.030, 95% CI: 0.010 to 0.051, p=0.004). It has been shown that metformin may reduce all-cause mortality in older patients with DM and reduce the risk of age-related diseases such as cardiovascular disease, cancer, depression, dementia, and frailty in older men with T2DM.66,67 Metformin may exert a significant anti-aging effect in older patients with T2DM.
Vigili de Kreutzenberg et al designed an RCT28 involving 38 prediabetic patients who received metformin (n=19) or placebo interventions for 2 months. Both groups had the same sex ratio and similar age. Peripheral blood mononuclear cells (PBMCs) TL was similar between the 2 groups at baseline. PBMCs TL increased significantly in the metformin group after 2 months (mean T/S ratio: 0.979 ± 0.011 vs 0.951 ± 0.007, p=0.012), whereas it did not change significantly in the control group. It is worth mentioning that the baseline glucose was similar in both groups (glucose: 5.8 ± 0.2 mmol/L vs 5.8 ± 0.2 mmol/L, p-value unknown). However, after 2 months, the metformin group showed a significant decrease in glucose compared to baseline (glucose: 5.3 ± 0.2 mmol/L vs 5.8 ± 0.2 mmol/L, p=0.018), whereas the control group did not exhibit any significant change in glucose compared to baseline. There was no significant difference in PBMCs TL change between the 2 groups after 2 months compared to baseline. This study also showed that metformin increases SIRT1,28 which, in turn, protects against DNA damage.68
SIRT1 is an nicotinamide adenine dinucleotide (NAD(+))-dependent class III histone deacetylase that can alleviate oxidative stress, inflammatory reactions, and apoptosis, regulate transcription factors related to lifespan, and control a variety of metabolic pathways.69,70 SIRT1 is a star target for regulating anti-aging activity and metabolism and is also an inextricable topic in research on aging-related diseases.70
Palacios et al68 studied both loss- and gain-of-function in mouse models of SIRT1 and found that overexpression of SIRT1 in mice reduced telomere erosion associated with cell division and aging of tissues, such as the liver and kidney. In contrast, loss of SIRT1 promotes telomere erosion and destroys telomere integrity, indicating that SIRT1 can positively regulate TL and delay telomere shortening; this effect depends on the telomerase pathway.68 Amano et al71 observed that the NAD(+) precursor nicotinamide mononucleotide (NMN) supplementation maintained TL and inhibited the DNA damage response by regulating the “longevity protein” Sirt1 activity. Another study72 found that SIRT1 may be involved in telomere maintenance, and SIRT1 deficiency may lead to telomere instability. In the above RCT,28 increased SIRT1 gene/protein expression and chromatin accessibility of the SIRT1 promoter were observed in the metformin group compared with baseline, and increased SIRT1 protein expression was observed in the metformin group compared with the control group, suggesting that metformin may prolong TL by increasing SIRT1 expression.
Diman et al73 found that metformin may promote human telomere transcription through the nuclear respiratory factor 1 (NRF1) and adenosine 5′-monophosphate (AMP)–activated protein kinase (AMPK)/peroxisome proliferator-activated receptor γ coactivator 1α (PGC-1α) axis upregulation telomeric repeat-containing RNA (TERRA). Kulkarni et al suggested that the maintenance of telomere homeostasis may involve a protective feedback mechanism mediated by the effects of metformin on mitochondrial and cellular aging.24 However, the mechanisms underlying the effects of metformin on TL remain unclear. It is necessary to explore the mechanism by which metformin delays aging in patients with T2DM from the perspective of telomeres, and the protective effects of metformin on telomeres need to be further explored.
Summary and Perspectives
This review summarizes clinical trials on the effect of antidiabetic medications on TL and the related mechanisms that may be involved. The effects of different antidiabetic medications on TL in patients with T2DM varied. Interventional studies28,30 have shown that sitagliptin and metformin may prolong the TL of patients with T2DM, which is accompanied by a significant decrease in their blood glucose or HbA1c, suggesting that the telomere extension of these patients may be related to the good control of blood glucose by antidiabetic medications. Interestingly, a longitudinal study37 has shown that insulin may accelerate telomere shortening in patients with T2DM. The effect of acarbose and glyburide on TL in patients with T2DM remains uncertain due to the lack of longitudinal research evidence. However, observational studies33,34,36 suggest that the use of acarbose may be associated with telomere shortening and glyburide may protect TL in patients with T2DM. Further research is needed to investigate the relevant mechanism of the effect of antidiabetic medications on TL in patients with T2DM. When recommending treatment for patients with T2DM, especially for patients with different subtypes of T2DM, endocrinologists can re-examine the antidiabetic regimen from the perspective of telomeres and determine the most appropriate antidiabetic medications to achieve the dual goals of effective hypoglycemia and delayed aging.
Sodium-Glucose Transporter 2 inhibitors (SGLT2i) and glucagon-like peptide-1 receptor agonists (GLP-1RA) have been confirmed to provide cardiovascular and renal benefits in patients with T2DM in large clinical trials and even reduce all-cause mortality in patients with T2DM.74 Mitiglinide improves postprandial metabolic disturbances and reduces excessive oxidative stress and inflammatory responses.75 All of these are potential anti-aging drugs or may have certain anti-aging effects; however, to date, no clinical trials have investigated the effect of these commonly used antidiabetic medications on TL. In the future, clinical trials are urgently needed to explore the effects of SGLT2i, GLP-1RA, glinides, and other antidiabetic medications on telomeres and to uncover more anti-aging drugs suitable for patients with T2DM.
It is worth noting that, limited by the small number of large-scale prospective clinical trials, the relevant conclusions need further confirmation. Endocrinologists will continue their pursuit of delaying aging in patients with T2DM. In the future, larger-scale prospective clinical trials and related basic research are needed to further explore this topic to decode the telomere password that connects antidiabetic medications to aging in patients with T2DM.
Acknowledgments
I sincerely thank my teacher, chief physician Yanhua Zhu (Department of Endocrinology, the Third Affiliated Hospital of Sun Yat-sen University), for providing inspiration for the topic selection of this review article.
Disclosure
The author reports no conflicts of interest in this work.
References
1. Sun H, Saeedi P, Karuranga S, et al. IDF diabetes atlas: global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res Clin Pract. 2022;183:109119. doi:10.1016/j.diabres.2021.109119
2. Zhu X, Hu J, Guo H, et al. Effect of metabolic health and obesity phenotype on risk of diabetes mellitus: a population-based longitudinal study. Diabet Metab Synd Obesit. 2021;14:3485–3498. doi:10.2147/DMSO.S317739
3. Chew NWS, Ng CH, Tan DJH, et al. The global burden of metabolic disease: data from 2000 to 2019. Cell Metab. 2023;35:414–428.e3. doi:10.1016/j.cmet.2023.02.003
4. Chakravarti D, LaBella KA, DePinho RA. Telomeres: history, health, and hallmarks of aging. Cell. 2021;184:306–322. doi:10.1016/j.cell.2020.12.028
5. Cheng F, Carroll L, Joglekar MV, et al. Diabetes, metabolic disease, and telomere length. Lancet Diabet Endocrinol. 2021;9:117–126. doi:10.1016/S2213-8587(20)30365-X
6. López-Otín C, Blasco MA, Partridge L, et al. Hallmarks of aging: an expanding universe. Cell. 2023;186:243–278. doi:10.1016/j.cell.2022.11.001
7. Verma AK, Singh P, Al-Saeed FA, et al. Unravelling the role of telomere shortening with ageing and their potential association with diabetes, cancer, and related lifestyle factors. Tissue Cell. 2022;79:101925. doi:10.1016/j.tice.2022.101925
8. Ihara Y, Toyokuni S, Uchida K, et al. Hyperglycemia causes oxidative stress in pancreatic beta-cells of GK rats, a model of type 2 diabetes. Diabetes. 1999;48:927–932. doi:10.2337/diabetes.48.4.927
9. Zhao J, Miao K, Wang H, et al. Association between telomere length and type 2 diabetes mellitus: a meta-analysis. PLoS One. 2013;8:e79993. doi:10.1371/journal.pone.0079993
10. Wang J, Dong X, Cao L, et al. Association between telomere length and diabetes mellitus: a meta-analysis. J Int Med Res. 2016;44:1156–1173. doi:10.1177/0300060516667132
11. Cheng F, Luk AO, Shi M, et al. Shortened leukocyte telomere length is associated with glycemic progression in type 2 diabetes: a prospective and mendelian randomization analysis. Diabetes Care. 2022;45:701–709. doi:10.2337/dc21-1609
12. Kidanie BB, Alem G, Zeleke H, et al. Determinants of diabetic complication among adult diabetic patients in debre markos referral hospital, northwest Ethiopia, 2018: unmatched case control study. Diabet Metab Synd Obesit. 2020;13:237–245. doi:10.2147/DMSO.S237250
13. Cheng F, Luk AO, Wu H, et al. Shortened relative leukocyte telomere length is associated with all-cause mortality in type 2 diabetes- analysis from the Hong Kong diabetes register. Diabetes Res Clin Pract. 2021;173:108649. doi:10.1016/j.diabres.2021.108649
14. Rai S, Badarinath ARS, George A, et al. Association of telomere length with diabetes mellitus and idiopathic dilated cardiomyopathy in a South Indian population: a pilot study. Mutat Res. 2022;874–875:503439. doi:10.1016/j.mrgentox.2021.503439
15. Cheng F, Luk AO, Tam CHT, et al. Shortened relative leukocyte telomere length is associated with prevalent and incident cardiovascular complications in type 2 diabetes: analysis from the Hong Kong diabetes register. Diabetes Care. 2020;43:2257–2265. doi:10.2337/dc20-0028
16. Spigoni V, Aldigeri R, Picconi A, et al. Telomere length is independently associated with subclinical atherosclerosis in subjects with type 2 diabetes: a cross-sectional study. Acta Diabetol. 2016;53:661–667. doi:10.1007/s00592-016-0857-x
17. Adaikalakoteswari A, Balasubramanyam M, Ravikumar R, et al. Association of telomere shortening with impaired glucose tolerance and diabetic macroangiopathy. Atherosclerosis. 2007;195:83–89. doi:10.1016/j.atherosclerosis.2006.12.003
18. Sharma R, Gupta A, Thungapathra M, et al. Telomere mean length in patients with diabetic retinopathy. Sci Rep. 2015;5:18368. doi:10.1038/srep18368
19. Testa R, Olivieri F, Sirolla C, et al. Leukocyte telomere length is associated with complications of Type 2 diabetes mellitus: telomere length and diabetic complications. Diabet Med. 2011;28:1388–1394. doi:10.1111/j.1464-5491.2011.03370.x
20. Tentolouris N, Nzietchueng R, Cattan V, et al. White blood cells telomere length is shorter in males with type 2 diabetes and microalbuminuria. Diabetes Care. 2007;30:2909–2915. doi:10.2337/dc07-0633
21. Khan J, Pernicova I, Nisar K, et al. Mechanisms of ageing: growth hormone, dietary restriction, and metformin. Lancet Diabet Endocrinol. 2023;11:261–281.
22. Sunjaya AP, Sunjaya AF. Targeting ageing and preventing organ degeneration with metformin. Diabetes Metab. 2021;47:101203. doi:10.1016/j.diabet.2020.09.009
23. Chen S, Gan D, Lin S, et al. Metformin in aging and aging-related diseases: clinical applications and relevant mechanisms. Theranostics. 2022;12:2722–2740. doi:10.7150/thno.71360
24. Kulkarni AS, Gubbi S, Barzilai N. Benefits of metformin in attenuating the hallmarks of aging. Cell Metab. 2020;32:15–30. doi:10.1016/j.cmet.2020.04.001
25. Chen J, Ou Y, Li Y, et al. Metformin extends C. elegans lifespan through lysosomal pathway. eLife. 2017;6:e31268. doi:10.7554/eLife.31268
26. Martin-Montalvo A, Mercken EM, Mitchell SJ, et al. Metformin improves healthspan and lifespan in mice. Nat Commun. 2013;4:2192. doi:10.1038/ncomms3192
27. Konopka AR, Laurin JL, Schoenberg HM, et al. Metformin inhibits mitochondrial adaptations to aerobic exercise training in older adults. Aging Cell. 2019;2019:18.
28. Vigilide Kreutzenberg S, Ceolotto G, Cattelan A, et al. Metformin improves putative longevity effectors in peripheral mononuclear cells from subjects with prediabetes. A randomized controlled trial. Nutrit Metabol Card Dis. 2015;25:686–693. doi:10.1016/j.numecd.2015.03.007
29. Garcia-Martin I, Penketh RJA, Janssen AB, et al. Metformin and insulin treatment prevent placental telomere attrition in boys exposed to maternal diabetes. PLoS One. 2018;13:e0208533. doi:10.1371/journal.pone.0208533
30. Ma D, Yu Y, Yu X, et al. The changes of leukocyte telomere length and telomerase activity after sitagliptin intervention in newly diagnosed type 2 diabetes. Diabetes Metab Res Rev. 2015;31:256–261. doi:10.1002/dmrr.2578
31. Cuevas Diaz P, Nicolini H, Nolasco-Rosales GA, et al. Telomere shortening in three diabetes mellitus types in a Mexican sample. Biomedicines. 2023;11:730. doi:10.3390/biomedicines11030730
32. Al-Daghri NM, Abdi S, Sabico S, et al. Gut-derived endotoxin and telomere length attrition in adults with and without type 2 diabetes. Biomolecules. 2021;11:1693. doi:10.3390/biom11111693
33. Al-Thuwaini TM. Association of antidiabetic therapy with shortened telomere length in middle-aged Type 2 diabetic patients. J Diabetes Metab Disord. 2021;20:1161–1168. doi:10.1007/s40200-021-00835-x
34. Huang J, Peng X, Dong K, et al. The association between antidiabetic agents and leukocyte telomere length in the novel classification of type 2 diabetes mellitus. Gerontology. 2021;67:60–68. doi:10.1159/000511362
35. AlDehaini DMB, Al-Bustan SA, Ali ME, et al. Shortening of the leucocytes’ telomeres length in T2DM independent of age and telomerase activity. Acta Diabetol. 2020;57:1287–1295. doi:10.1007/s00592-020-01550-4
36. Liu J, Ge Y, Wu S, et al. Association between antidiabetic agents use and leukocyte telomere shortening rates in patients with type 2 diabetes. Aging. 2019;11:741–755. doi:10.18632/aging.101781
37. Zeng J, Liu H, Ping F, et al. Insulin treatment affects leukocyte telomere length in patients with type 2 diabetes: 6-year longitudinal study. J Diabetes Complications. 2019;33:363–367. doi:10.1016/j.jdiacomp.2019.02.003
38. Rosa ECCC, Dos Santos RRC, Fernandes LFA, et al. Leukocyte telomere length correlates with glucose control in adults with recently diagnosed type 2 diabetes. Diabetes Res Clin Pract. 2018;135:30–36. doi:10.1016/j.diabres.2017.10.020
39. Robb-MacKay C. The Effect of Metformin on Absolute Telomere Length. Lakehead University; 2018.
40. Gilfillan C, Naidu P, Gunawan F, et al. Leukocyte telomere length in the neonatal offspring of mothers with gestational and pre-gestational diabetes. PLoS One. 2016;11:e0163824. doi:10.1371/journal.pone.0163824
41. Tamura Y, Izumiyama-Shimomura N, Kimbara Y, et al. β-cell telomere attrition in diabetes: inverse correlation between HbA1c and telomere length. J Clin Endocrinol Metab. 2014;99:2771–2777. doi:10.1210/jc.2014-1222
42. Salpea KD, Talmud PJ, Cooper JA, et al. Association of telomere length with type 2 diabetes, oxidative stress and UCP2 gene variation. Atherosclerosis. 2010;209:42–50. doi:10.1016/j.atherosclerosis.2009.09.070
43. Mushtaq A, Azam U, Mehreen S, et al. Synthetic α-glucosidase inhibitors as promising anti-diabetic agents: recent developments and future challenges. Eur J Med Chem. 2023;249:115119. doi:10.1016/j.ejmech.2023.115119
44. Haran JP, McCormick BA. Aging, frailty, and the microbiome—how dysbiosis influences human aging and disease. Gastroenterology. 2021;160:507–523. doi:10.1053/j.gastro.2020.09.060
45. Kim KH, Chung Y, Huh J-W, et al. Gut microbiota of the young ameliorates physical fitness of the aged in mice. Microbiome. 2022;10:238. doi:10.1186/s40168-022-01386-w
46. Badal VD, Vaccariello ED, Murray ER, et al. The gut microbiome, aging, and longevity: a systematic review. Nutrients. 2020;12:3759. doi:10.3390/nu12123759
47. Gu Y, Wang X, Li J, et al. Analyses of gut microbiota and plasma bile acids enable stratification of patients for antidiabetic treatment. Nat Commun. 2017;8:1785. doi:10.1038/s41467-017-01682-2
48. Ni Y, Yang X, Zheng L, et al. Lactobacillus and bifidobacterium improves physiological function and cognitive ability in aged mice by the regulation of gut microbiota. Mol Nutr Food Res. 2019;63:1900603. doi:10.1002/mnfr.201900603
49. Lew LC, Hor YY, Jaafar MH, et al. Lactobacilli modulated AMPK activity and prevented telomere shortening in ageing rats. Benef Microbes. 2019;10:883–892. doi:10.3920/BM2019.0058
50. Hor -Y-Y, Ooi C-H, Khoo B-Y, et al. Lactobacillus strains alleviated aging symptoms and aging-induced metabolic disorders in aged rats. J Med Food. 2019;22:1–13. doi:10.1089/jmf.2018.4229
51. Andreadi A, Bellia A, Di Daniele N, et al. The molecular link between oxidative stress, insulin resistance, and type 2 diabetes: a target for new therapies against cardiovascular diseases. Curr Opin Pharmacol. 2022;62:85–96. doi:10.1016/j.coph.2021.11.010
52. Armstrong E, Boonekamp J. Does oxidative stress shorten telomeres in vivo? A meta-analysis. Ageing Res Rev. 2023;85:101854. doi:10.1016/j.arr.2023.101854
53. Han T, Yuan T, Liang X, et al. Sarcopenic obesity with normal body size may have higher insulin resistance in elderly patients with type 2 diabetes mellitus. Diabet Metab Synd Obesit. 2022;15:1197–1206. doi:10.2147/DMSO.S360942
54. Båvenholm PN, Efendic S. Postprandial hyperglycaemia and vascular damage - The benefits of acarbose. Diabet Vascul Dis Res. 2006;3:72–79. doi:10.3132/dvdr.2006.017
55. Rösen P, Osmers A. Oxidative stress in young Zucker rats with impaired glucose tolerance is diminished by acarbose. Horm Metab Res. 2006;38:575–586. doi:10.1055/s-2006-950397
56. Scott LJ. Sitagliptin: a Review in Type 2 Diabetes. Drugs. 2017;77:209–224. doi:10.1007/s40265-016-0686-9
57. Costes S, Bertrand G, Ravier MA. Mechanisms of beta-cell apoptosis in type 2 diabetes-prone situations and potential protection by GLP-1-based therapies. IJMS. 2021;22:5303. doi:10.3390/ijms22105303
58. Khalangot M, Krasnienkov D, Vaiserman A. Telomere length in different metabolic categories: clinical associations and modification potential. Exp Biol Med. 2020;245:1115–1121. doi:10.1177/1535370220931509
59. Rosen J, Jakobs P, Ale-Agha N, et al. Non-canonical functions of telomerase reverse transcriptase – impact on redox homeostasis. Redox Biol. 2020;34:101543. doi:10.1016/j.redox.2020.101543
60. Wilcox T, De Block C, Schwartzbard AZ, et al. Diabetic agents, from metformin to SGLT2 inhibitors and GLP1 receptor agonists. J Am Coll Cardiol. 2020;75:1956–1974. doi:10.1016/j.jacc.2020.02.056
61. Khunti K, Chatterjee S, Gerstein HC, et al. Do sulphonylureas still have a place in clinical practice? Lancet Diabet Endocrinol. 2018;6:821–832. doi:10.1016/S2213-8587(18)30025-1
62. Lv W, Wang X, Xu Q, et al. Mechanisms and characteristics of sulfonylureas and glinides. CTMC. 2020;20:37–56. doi:10.2174/1568026620666191224141617
63. Ahmed W, Lingner J. Impact of oxidative stress on telomere biology. Differentiation. 2018;99:21–27. doi:10.1016/j.diff.2017.12.002
64. Edens MA, Van Dijk PR, Hak E, et al. Determinants of excessive weight gain after the initiation of insulin therapy in type 2 diabetes mellitus: retrospective inception cohort study (ZODIAC 60). Diabetes Res Clin Pract. 2023;200:110719. doi:10.1016/j.diabres.2023.110719
65. Verkouter I, Noordam R, Le Cessie S, et al. The association between adult weight gain and insulin resistance at middle age: mediation by visceral fat and liver fat. JCM. 2019;8:1559. doi:10.3390/jcm8101559
66. Ferri-Guerra J, Aparicio-Ugarriza R, Mohammed YN, et al. Propensity score matching to determine the impact of metformin on all-cause mortality in older veterans with diabetes mellitus. South Med J. 2022;115:208–213. doi:10.14423/SMJ.0000000000001363
67. Wang C-P, Lorenzo C, Habib SL, et al. Differential effects of metformin on age related comorbidities in older men with type 2 diabetes. J Diabetes Complications. 2017;31:679–686. doi:10.1016/j.jdiacomp.2017.01.013
68. Palacios JA, Herranz D, De Bonis ML, et al. SIRT1 contributes to telomere maintenance and augments global homologous recombination. J Cell Biol. 2010;191:1299–1313. doi:10.1083/jcb.201005160
69. Osum M, Serakinci N. Impact of circadian disruption on health; SIRT1 and Telomeres. DNA Repair (Amst). 2020;96:102993. doi:10.1016/j.dnarep.2020.102993
70. Han X, Ding C, Sang X, et al. Targeting Sirtuin1 to treat aging-related tissue fibrosis: from prevention to therapy. Pharmacol Ther. 2022;229:107983. doi:10.1016/j.pharmthera.2021.107983
71. Amano H, Chaudhury A, Rodriguez-Aguayo C, et al. Telomere dysfunction induces sirtuin repression that drives telomere-dependent disease. Cell Metab. 2019;29:1274–1290.e9. doi:10.1016/j.cmet.2019.03.001
72. El Ramy R, Magroun N, Messadecq N, et al. Functional interplay between Parp-1 and SirT1 in genome integrity and chromatin-based processes. Cell Mol Life Sci. 2009;66:3219–3234. doi:10.1007/s00018-009-0105-4
73. Diman A, Boros J, Poulain F, et al. Nuclear respiratory factor 1 and endurance exercise promote human telomere transcription. Sci Adv. 2016;2:e1600031. doi:10.1126/sciadv.1600031
74. Brown E, Heerspink HJL, Cuthbertson DJ, et al. SGLT2 inhibitors and GLP-1 receptor agonists: established and emerging indications. Lancet. 2021;398:262–276. doi:10.1016/S0140-6736(21)00536-5
75. Kitasato L, Tojo T, Hatakeyama Y, et al. Postprandial hyperglycemia and endothelial function in type 2 diabetes: focus on mitiglinide. Cardiovasc Diabetol. 2012;11:79. doi:10.1186/1475-2840-11-79
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Impact of Moderate Physical Activity on Inflammatory Markers and Telomere Length in Sedentary and Moderately Active Individuals with Varied Insulin Sensitivity
Almuraikhy S, Sellami M, Al-Amri HS, Domling A, Althani AA, Elrayess MA
Journal of Inflammation Research 2023, 16:5427-5438
Published Date: 20 November 2023
Mechanisms and Drug-Augmenting Strategies of Mesenchymal Stem Cells for Preserving β-Cell in Type 2 Diabetes
Wang K, Li J, Han C, Li J
Drug Design, Development and Therapy 2026, 20:586404
Published Date: 22 April 2026