Back to Journals » Journal of Pain Research » Volume 11
Caffeic acid phenethyl ester attenuates neuropathic pain by suppressing the p38/NF-κB signal pathway in microglia
Authors Cheng H, Zhang Y, Lu W, Gao X, Xu C, Bao H
Received 22 February 2018
Accepted for publication 26 July 2018
Published 1 November 2018 Volume 2018:11 Pages 2709—2719
DOI https://doi.org/10.2147/JPR.S166274
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Michael Überall
Hao Cheng, Yong Zhang, Weiping Lu, Xianzhong Gao, Chenjie Xu, Hongguang Bao
Department of Anesthesiology, Nanjing First Hospital, Nanjing Medical University, Nanjing 210006, China
Background: Management of neuropathic pain is still a clinical challenge. Evidence has accumulated indicating that propolis is effective in attenuating neuropathic pain; however, the mechanism is not fully understood. Our present study investigated the effects and the possible mechanism of caffeic acid phenethyl ester (CAPE), the main ingredient of propolis, in improving neuropathic pain via its inhibition on p38/NF-κB signal pathway in microglia.
Materials and methods: Chronic constriction injury (CCI) mice model and the microglial cell line BV-2 were used to investigate the effects and the mechanism of CAPE. Cell signaling was measured by real-time PCR, Western blotting and immunofluorescence assay.
Results: CAPE relieved neuropathic pain behaviors induced by CCI in mice. CAPE also inhibited CCI-induced activation of microglia. Furthermore, CAPE suppressed the phosphorylation of p38 mitogen-activated protein kinase, inhibited the translocation of NF-κB and decreased the expression of proinflammatory cytokines tumor necrosis factor-α, IL-1β and IL-6.
Conclusion: CAPE was found to be an effective and safe drug candidate for alleviating neuropathic pain by its powerful inhibition on the P38/NF-κB signal pathway.
Keywords: caffeic acid phenethyl ester, NF-κB, neuropathic pain, microglia
Introduction
Neuropathic pain is caused by damage or disease affecting the somatosensory nervous system and its treatment has remained a clinical challenge.1 Studies have shown that microglia play a key role in the development and maintenance of neuropathic pain.2 Activation of microglia synthesizes and releases inflammatory factors, including tumor necrosis factor-alpha (TNF-α), IL-1β and IL-6, which further activates microglia. This positive feedback loop causes central sensitization and aggravates neuropathic pain.3 Moreover, studies show that minocycline, the inhibitor of microglia, reduced neuropathic pain,4,5 but its clinical use is limited by severe side effects. Therefore, a safer and more effective inhibitor of microglia for neuropathic pain treatment is urgently needed.
Mitogen-activated kinase (MAPK) pathways are important for inflammatory responses in neuropathic pain, especially the p38 MAPK. Studies show that spinal microglia p38 MAPK is activated after nerve injury, and it has been shown to activate the transcription factor NF-κB, leading to the upregulation of TNF-α, IL-1β and IL-6 expression.6 Accumulating evidence has proven that inhibiting p38 can suppress microglial activation and alleviate pain-related behaviors in animal models,7,8 thereby acting as an important analgesic target.
Caffeic acid phenethyl ester (CAPE) is the main ingredient of propolis, which has been widely used in traditional Chinese medicine to treat various diseases.9 It has antioxidative, antitumor, anti-inflammatory and many other pharmacological effects.10 For instance, Tolba et al reported that CAPE exerted therapeutic effects on atherosclerosis and Alzheimer’s disease.11 CAPE also ameliorated lipopolysaccharide (LPS)-induced microglial activation and motor incoordination.12 However, the mechanism by which CAPE treats neuropathic pain is still largely unknown.
Hereby, we hypothesize that CAPE may attenuate chronic constrictive injury (CCI)-induced neuropathic pain via inhibition of microglial activity by suppressing the p38/NF-κB signal pathway. This study may provide new insights into the mechanism of CAPE and the application of its clinical analgesic effect.
Materials and methods
Ethics statement
All procedures were strictly performed in accordance with the regulations of the ethics committee of the International Association for the Study of Pain and the Guide for the Care and Use of Laboratory Animals (The Ministry of Science and Technology of China, 2006). All animal experiments were approved by the Nanjing Medical University Animal Care and Use Committee, and were designed to minimize suffering and the number of animals used.
Animals
Adult male CD-1 mice weighing 18–22 g were provided by the Experimental Animal Center at Nanjing Medical University, Nanjing, China. All animals were housed under controlled temperature (22°C±2°C) and a 12-hour light/dark cycle (lights on at 8:00 a.m.). The animals had free access to food and water. All animals were allowed to acclimatize to these conditions for at least 2 days before starting the experiments.
Neuropathic pain model
CCI model: Mice were injected intraperitoneally (i.p.) with chloral hydrate (300 mg/kg) and fixed in a prone position. The left common sciatic nerve of each mouse was exposed at the mid-thigh level. In the sciatic nerve close to the bifurcation, four ligatures (5-0 chronic gut) were tied loosely around the sciatic nerve. The distance between the ligatures was 1 mm.
Drugs and reagents
CAPE was purchased from Sigma (St Louis, MO, USA). Antibodies for ionized calcium-binding adapter molecule 1 (IBA-1) were purchased from Abcam (Cambridge, MA, USA). Antibodies for phospho-p38 MAPK (Thr180/Tyr182), p38 MAPK, phospho-NF-κB p65 (Ser536), NF-κB p65 and secondary antibodies were purchased from Cell Signaling Technology (Beverly, MA, USA). MTT was purchased from Sunshine Biotechnology (Nanjing, China). FBS was purchased from Gibco. Antibody for glyceraldehyde 3-phosphate dehydrogenase (GAPDH), LPS, dimethyl sulfoxide and other reagents were purchased from Sigma.
Assessment of pain behaviors
Rats were housed in a clear plexiglass box whose bottom was made of barbed wire. Von Frey Hairs test was used to (Woodland Hills, LA, USA) vertically stimulate the plantar surface of the hind paw of each mouse for three times. The average of the threshold was measured.
Western blotting
The spinal cord segments at L1–L6 were rapidly removed and homogenized in RIPA lysis buffer after deep anesthesia with chloral hydrate. The protein concentrations were determined by Bradford method and 30–50 µg of proteins was loaded and separated by SDS-PAGE electrophoresis. The protein was transferred to a polyvinylidene fluoride membrane. The membranes were blocked with 5% BSA for 1 hour at room temperature and incubated overnight at 4°C with the primary antibodies, including p-p65 (1:1,000), p65 (1:1,000), p-p38 (1:1,000), p38 (1:1,000) and IBA-1 (1:1,000). GAPDH (1:8,000) was used as a loading control. The polyvinylidene fluoride membrane was washed three times for 10 minutes each with TBST and then incubated with the corresponding secondary antibody. Data were analyzed by Quantity One-4.6.5 (Bio-Rad Laboratories, Berkeley, CA, USA).
Immunofluorescence assay
Mice were injected i.p. with chloral hydrate (300 mg/kg) and perfused with saline and then perfused with 4% araformaldehyde. The L4 and/or L5 lumbar segments were then dissected and fixed in 4% paraformaldehyde. Tissues were cut into 25 µm thickness using a cryostat. Each group of sections was incubated with IBA-1 (1:100) overnight and then washed and incubated with the secondary antibody for 2 hours. Samples was observed by immunofluorescence microscope and analyzed by Image Pro Plus 6.0 software (Media Cybernetics Inc., Rockville, MD, USA).
Real time-polymerase chain reaction
According to the manufacturer’s protocol, the spinal cord tissue and the total cellular RNA were extracted by TRIzol method and their purity was measured. Reverse transcription of RNA into cDNA, cDNA were used as a template for PCR amplification. The sense and antisense primers (TNF-α, IL-1β, IL-6 and GAPDH) were as follows: TNF-alpha: 5′-TACTGAACTTCGGGGTGATTGGTCC-3′ and 5′-CAGCCTTGTCCCTTGAAGAGAAC-3′; IL-1β: 5′-GCACTACAGGCTCCGAGATGAAC-3′ and 5′-TTGTCGTTGCTTGGTTCTCCTTGT-3′; IL-6: 5′-CCGGAGAGGAGACTTCACAG-3′ and 5′-GGAAATTGGGGTAGGAAGGA-3′; GAPDH: 5′-ACCACAGTCCATGCCATCAC-3′ and 5′-CACCACCCTGTTGCTGTAGCC-3′.
BV2 cells culture
BV2 cell lines were purchased from American Type Culture Collection. They were inoculated into culture flasks in appropriate concentrations, with DMEM containing 10% FBS, 100 U/mL penicillin and 100 µg/mL streptomycin. Then, they were kept at 37°C in humidified 5% CO2 overnight, and the next morning, the medium was changed to serum-free medium and then the cells were treated with LPS (1 µg/mL) with or without CAPE (100 µM) for 12 hours. We used dimethyl sulfoxide (0.1%) as a control.
NF-κB activation assay
BV-2 cells were plated in cell culture dishes and treated with LPS (1 µg/mL) for 12 hours with or without CAPE (100 µM). Then 4% paraformaldehyde was added to the cells and they were fixed at room temperature for 30 minutes. After blocking with 1% BSA for 1 hour at room temperature and incubating overnight at 4°C with the primary antibodies, the cells were incubated with the corresponding secondary antibody for 1 hour. Finally, anti-fluorescence annihilation mount solution was added. Coverslips were stained with 1 µg/mL DAPI for 1 minute. Confocal microscopy was performed with Olympus FV1000 confocal system.
Statistical analyses
Statistical analysis was carried out using Student’s two-tailed t-test for comparison between two groups and with two-way ANOVA followed by Dunnett’s test when the data involved three or more groups. Results are expressed as mean ± standard error of the mean. P<0.05 was considered statistically significant. All analyses were performed with GraphPad Prism Version 5.01 (GraphPad Software Inc., San Diego, CA, USA).
Results
CAPE attenuated CCI-induced neuropathic pain
To investigate the effects of CAPE on CCI mice, Von Frey Hairs test was performed. After 14 days of CCI surgery, the mechanical threshold was marked decreased in CCI-treated mice (Figure 1A). Pain-related behaviors were greatly ameliorated by a single dose of CAPE (5 and 25 mg/kg, i.p.), as shown in Figure 1B. Hence, a dose of 25 mg/kg of CAPE was selected for subsequent experiments, considering its best effect. Furthermore, continuous administration of CAPE (25 mg/kg, i.p., for 7 days) greatly ameliorated the mechanical threshold induced by CCI (Figure 1C). There was no significant difference in pain-related behaviors between the control group and the CAPE-treated groups in naïve mice (Figure 1D).
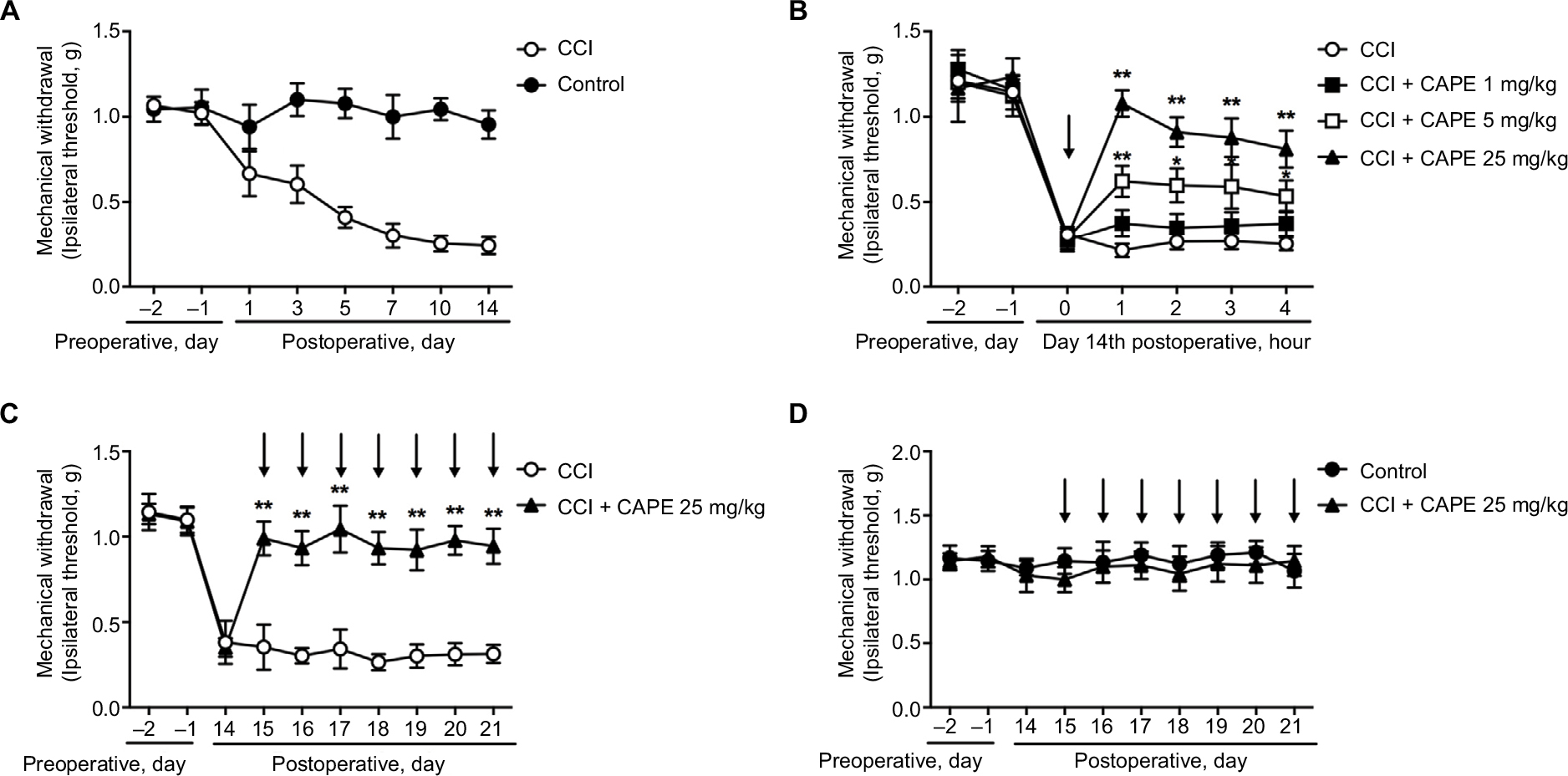 | Figure 1 CAPE attenuated CCI-induced neuropathic pain. Notes: (A) Mechanical allodynia was significantly induced after CCI surgery. (B) Single administration of CAPE (1, 5, 25 mg/kg, i.p.) significantly attenuated CCI-induced mechanical allodynia (n=6). (C) Consecutive administration of CAPE (25 mg/kg, i.p.) significantly attenuated CCI-induced mechanical allodynia (n=6). (D) No significant difference was seen on CAPE (25 mg/kg, i.p.) treatment in naïve rats. Mechanical pain threshold was tested by Von Frey Hairs. n=6 in each group. Two-way ANOVA revealed a significant difference at **P<0.01 vs CCI. Drug administration is indicated by arrows. Abbreviations: CAPE, caffeic acid phenethyl ester; CCI, chronic constrictive injury; i.p., intraperitoneal. |
CAPE inhibited CCI-induced microglia activation
Numerous studies have demonstrated the critical role of microglia in the development of neuropathic pain.13 Therefore, we evaluated the effect of CAPE in activation of microglia induced by CCI. Our results showed that CAPE (25 mg/kg, i.p.) significantly inhibited the upregulation of microglia marker IBA-1 in spinal cord after CCI (Figure 2A). Furthermore, immunofluorescence of IBA-1 in the dorsal horn showed CAPE inhibited the activation of microglia (Figure 2B–E).
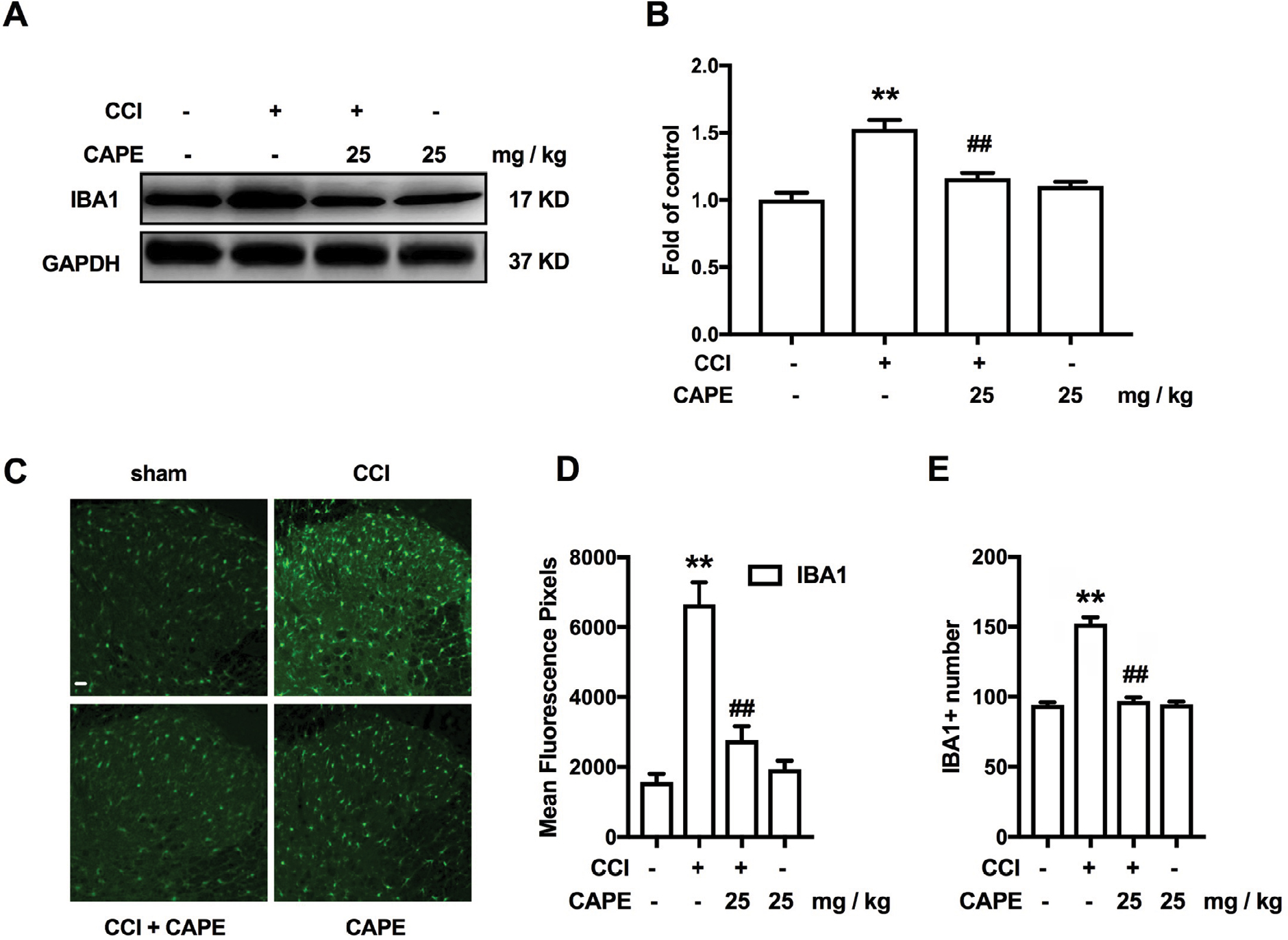 | Figure 2 CAPE significantly inhibited CCI-induced activation of microglia. Notes: (A, B) Effects of CAPE (25 mg/kg, i.p.) on the expression of IBA-1 in the spinal cord. (C-E) Confocal images and immunofluorescence analysis data showing IBA-1 in the dorsal horns. Quantification of immunofluorescence was represented as the mean fluorescence pixels in the superficial dorsal horns. CAPE was consecutively administered daily from day 15 to day 21 after CCI operation. The lumbar spines (L1–L6) were collected and analyzed 60 minutes after the last drug administration. n=4. Two-way ANOVA revealed a significant difference at **P<0.01 vs sham; and ##P<0.01 vs CCI. Scale bar 50μm, original magnification ×200. Abbreviations: CAPE, caffeic acid phenethyl ester; CCI, chronic constrictive injury; IBA-1, ionized calcium-binding adapter molecule 1; i.p., intraperitoneal. |
CAPE inhibited CCI-induced p38 MAPK and NF-κB p65 phosphorylation
Western blotting revealed that CAPE dramatically decreased CCI-induced phosphorylation of p38 MAPK after continuous administration of CAPE (25 mg/kg, i.p.), as shown in Figure 3A. We next analyzed the effects of CAPE on NF-κB. CAPE decreased the phosphorylation of p65 induced by CCI (Figure 3B). Immunofluorescence assay showed that the phosphorylated p38 was localized primarily with microglia (Figure 4A). Moreover, CCI caused a significantly increased phosphorylation of p38 and p65 in microglia, whereas CAPE could suppress these effects (Figure 4A and B), in line with the Western blot data.
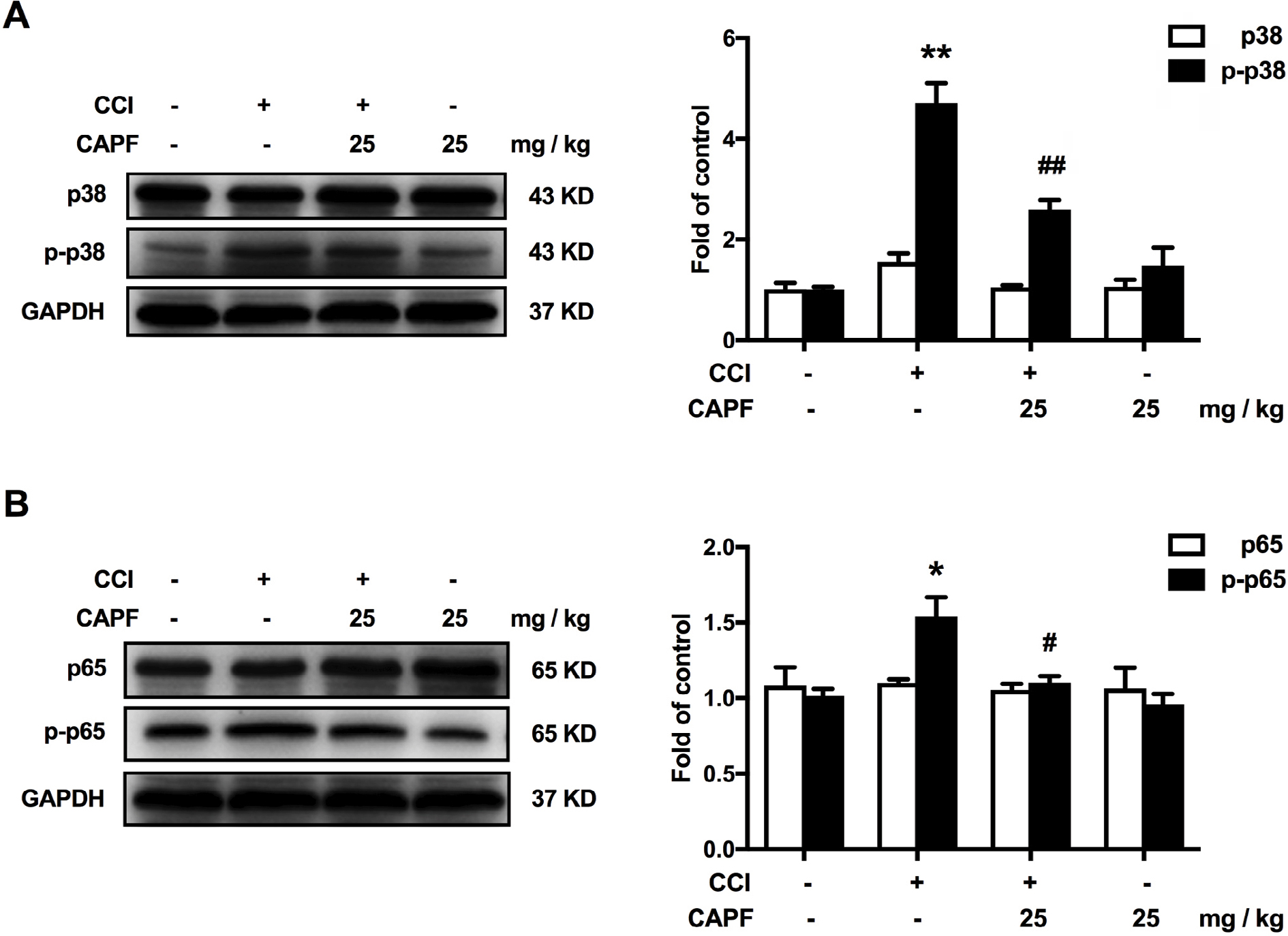 | Figure 3 CAPE significantly attenuated CCI-induced microglia activation by inhibiting p38 MAPK/NF-κB in spinal cord. Notes: (A) CAPE significantly inhibited CCI-induced phosphorylation of p38. (B) CAPE significantly inhibited CCI-induced phosphorylation of p65. CAPE (25 mg/kg, i.p.) was administered daily from day 15 to day 21 after CCI operation. The lumbar spines (L1–L6) were collected and analyzed 60 minutes after the last administration. n=4.Two-way ANOVA revealed a significant difference at *P<0.05 and **P<0.01 vs sham; #P<0.05 and ##P<0.01 vs CCI. Abbreviations: CAPE, caffeic acid phenethyl ester; CCI, chronic constrictive injury; i.p., intraperitoneal. |
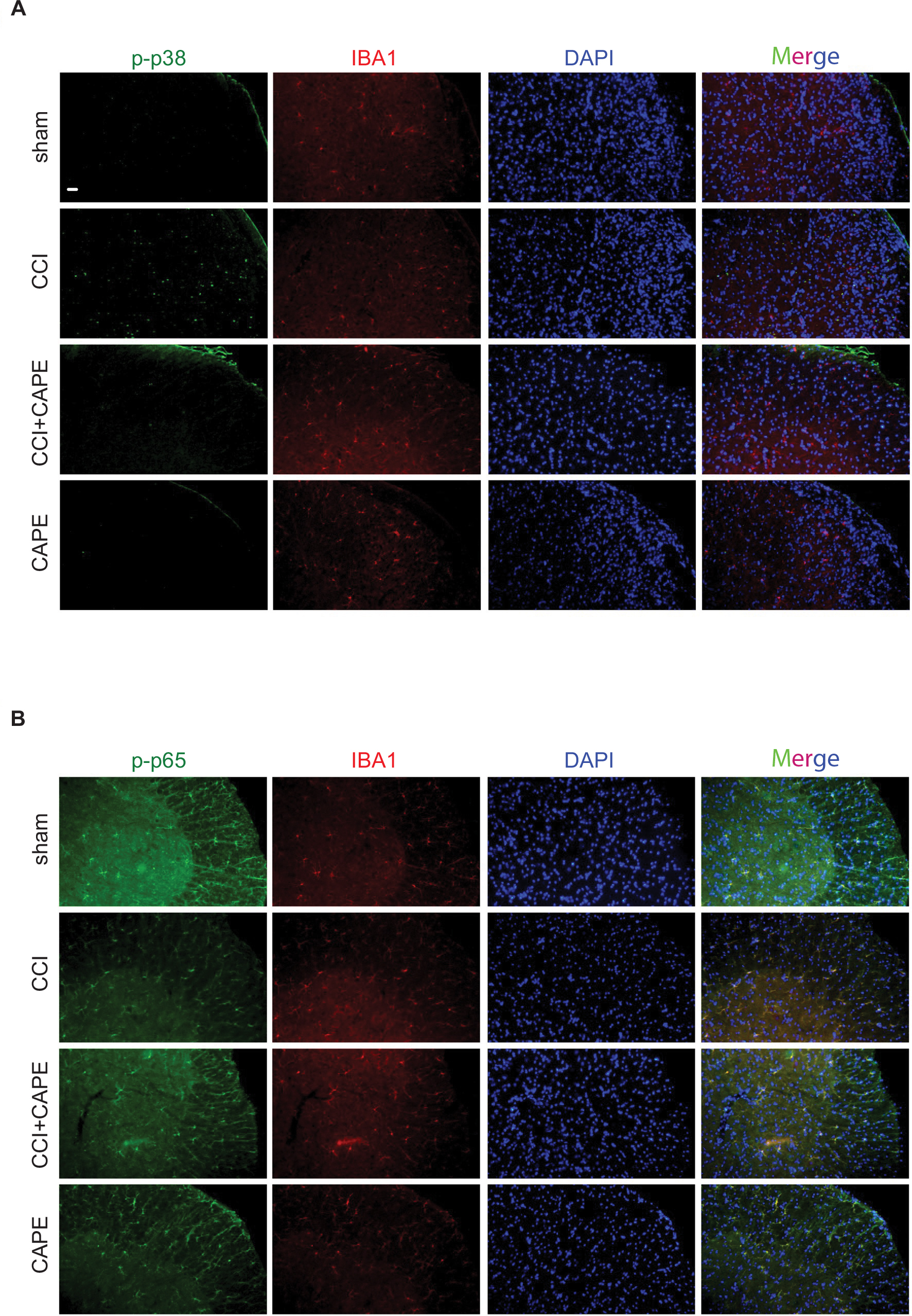 | Figure 4 CAPE suppressed CCI-induced phosphorylation of p38 and p65 in the spinal cords. Notes: (A, B) Confocal images show the cellular distribution of phosphorylated p38 and p65 in the superficial dorsal horns. CAPE was given once a day for 7 days from postoperative day 14. One hour after the final administration, spinal samples were collected (n=4). Abbreviations: CAPE, caffeic acid phenethyl ester; CCI, chronic constrictive injury; TNF, tumor necrosis factor. |
CAPE suppressed CCI-induced upregulated expression of proinflammatory cytokines in the spinal cord
We further examined the impact of CAPE on proinflammatory cytokines after CCI. Our quantitative real-time PCR analysis data demonstrated that, pro-inflammatory cytokines (TNF-α, IL-1β and IL-6) mRNA levels were notably increased in CCI mice, whereas administration of CAPE suppressed them effectively (Figure 5A–D).
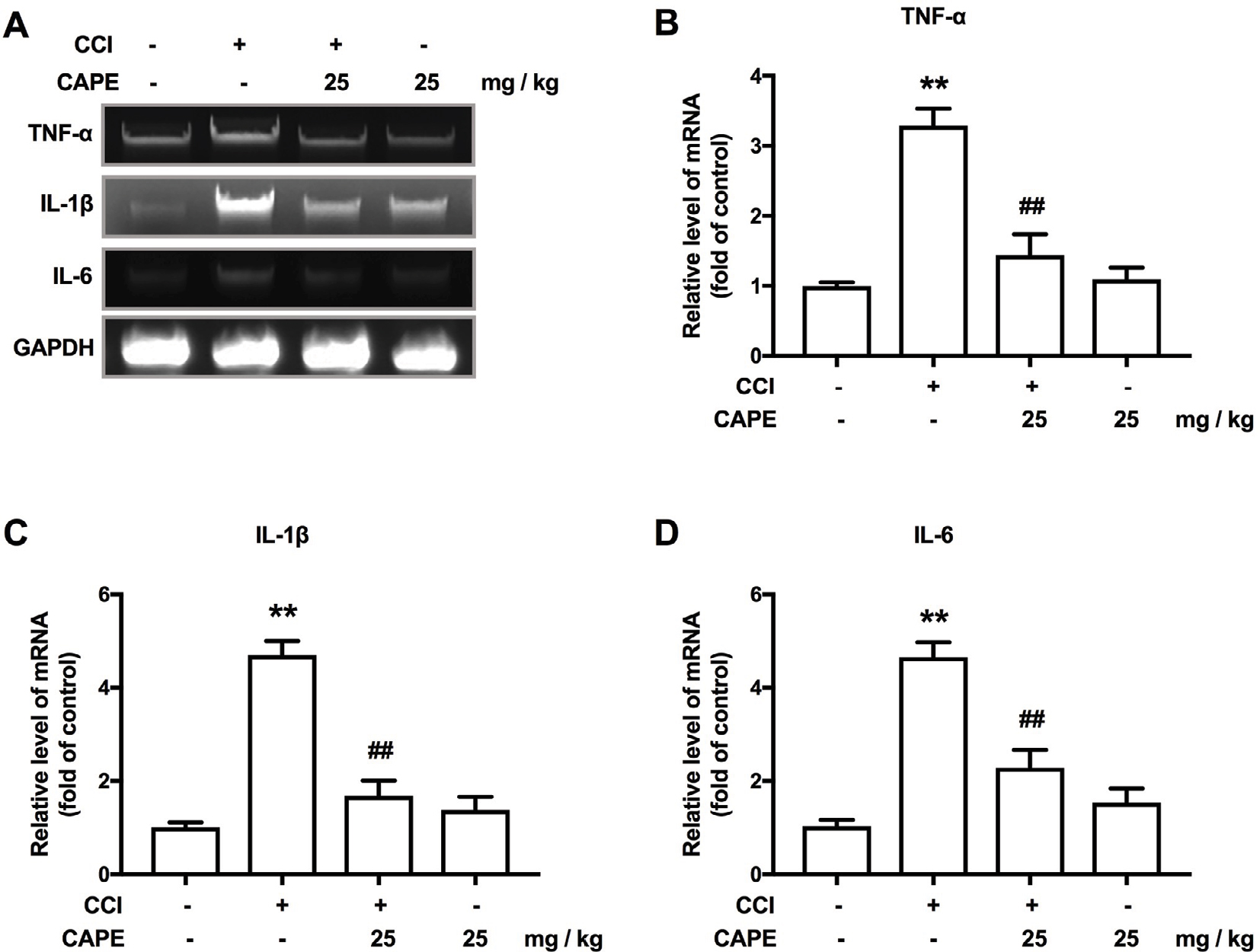 | Figure 5 CAPE inhibited CCI-induced upregulated expression of proinflammatory cytokines in the spinal cord. Notes: (A) CAPE inhibited the CCI-induced upregulation of TNF-α, IL-1β and IL-6 in the spinal cord. (B) CAPE inhibited the CCI-induced upregulation of TNF-α in the spinal cord. (C) CAPE suppressed the CCI-induced upregulation of IL-1β in the spinal cord. (D) CAPE inhibited the CCI-induced upregulation of IL-6 in the spinal cord. The mRNA levels of TNF-α, IL-1β and IL-6 were determined using real-time quantitative PCR. GAPDH was used as an invariant control. n=4. Two-way ANOVA revealed a significant difference at **P<0.01 vs sham; ##P<0.01 vs CCI. Abbreviations: CAPE, caffeic acid phenethyl ester; CCI, chronic constrictive injury; DMSO, dimethyl sulfoxide; LPS, lipopolysaccharide; TNF, tumor necrosis factor. |
CAPE suppressed LPS-induced microglial activation by inhibiting p38 MAPK and NF-κB signaling
To investigate the effects of CAPE on LPS-induced microglial activation in vitro, microglial BV-2 cells were used to confirm the role of CAPE on p38 and NF-κB. As shown in Figure 6, LPS increased the phosphorylation of p38 MAPK and promoted the translocation of NF-κB from the cytosol to the nucleus. Compared with the LPS-treated group, CAPE significantly reduced p38 MAPK phosphorylation and p65 translocation (Figure 6A–C). MTT assay indicated that the different doses of CAPE did not affect cell proliferation (Figure 6D).
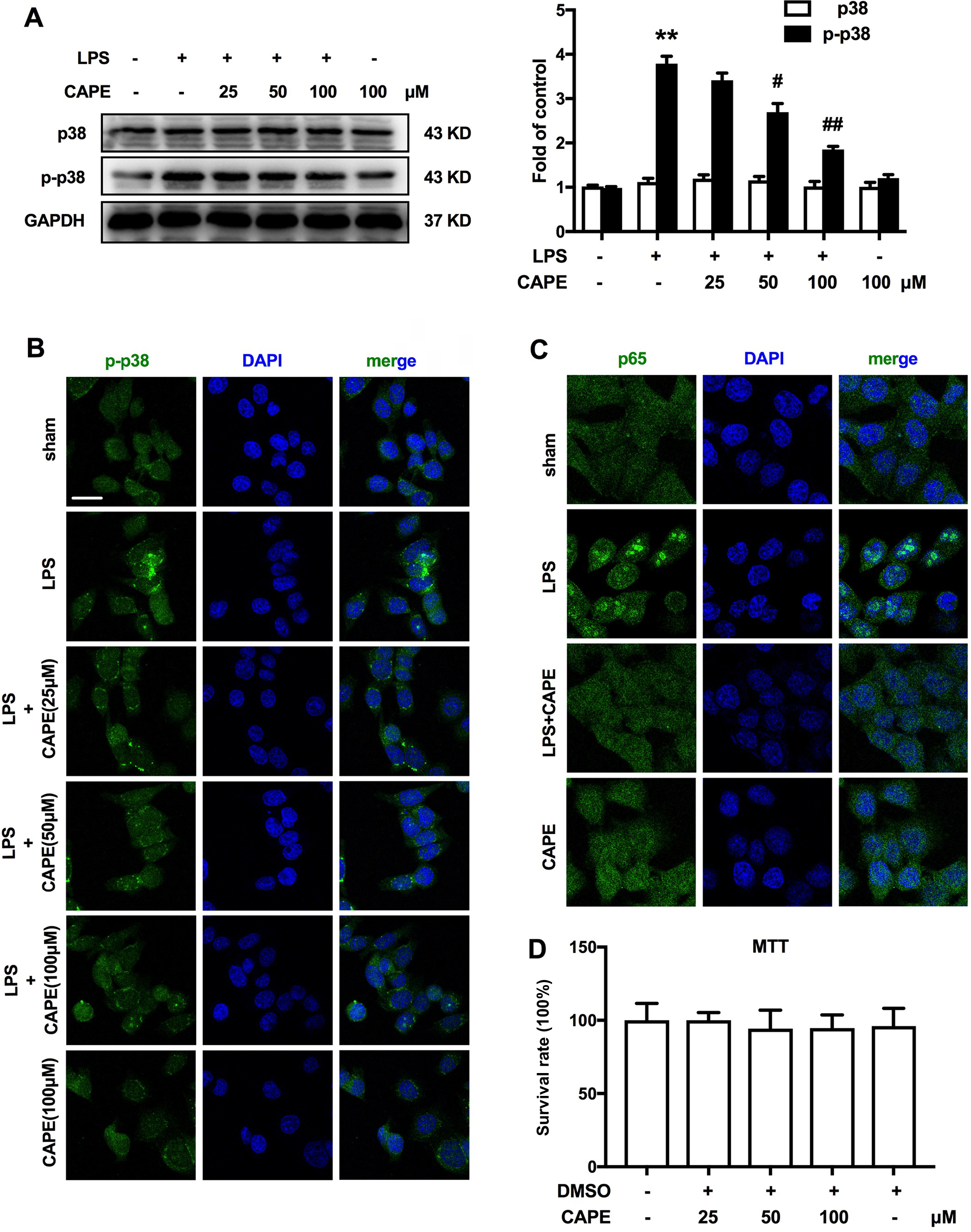 | Figure 6 CAPE suppressed LPS-induced microglia activation by inhibiting p38 MAPK/NF-κB. Notes: (A, B) Pre-administration of CAPE (25, 50, 100 µM) significantly inhibited the LPS-induced phosphorylation of p38 MAPK. (C) CAPE (100 µM) inhibited the translocation of NF-κB from the cytosol to the nucleus after LPS treatment for 6 hours in BV-2 cells. (D) MTT experiments showed that different doses of CAPE did not affect cell proliferation. n=4. Two-way ANOVA revealed a significant difference at *P<0.05 and **P<0.01 vs control; #P<0.05 and ##P<0.01 vs LPS-treated group. Scale bar 20μm, original magnification ×800. Abbreviations: CAPE, caffeic acid phenethyl ester; LPS, lipopolysaccharide; TNF, tumor necrosis factor. |
CAPE suppressed LPS-induced upregulated expression of proinflammatory cytokines in BV-2 cells
We further examined the effects of CAPE on proinflammatory cytokines after LPS treatment. LPS treatment induced robust activation in BV-2 cells, characterized by increased mRNA expression of the proinflammatory cytokines TNF-α, IL-1β and IL-6 (Figure 7A–D). Compared with the LPS-treated group, co-incubation with CAPE significantly reduced these effects (Figure 7A–D).
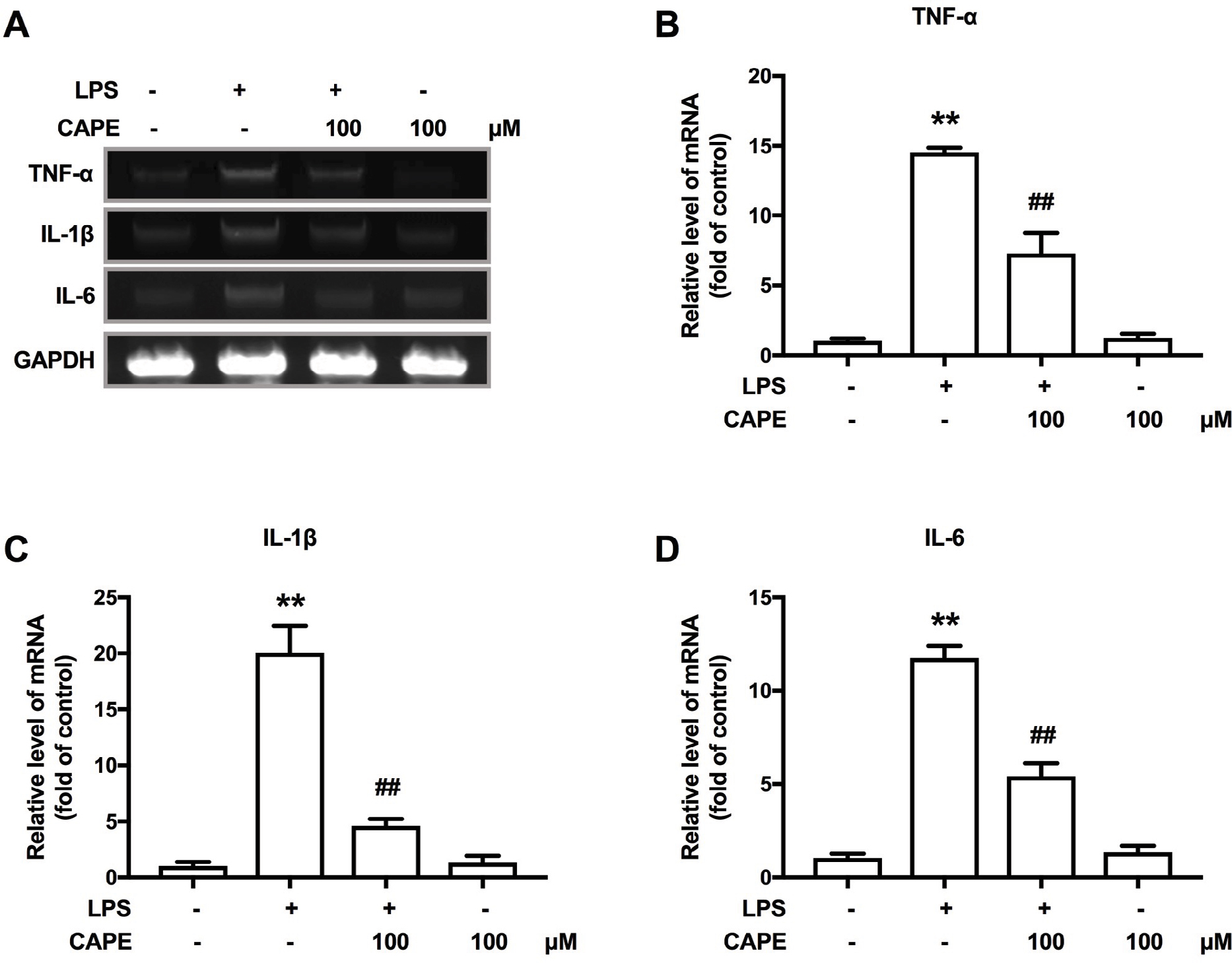 | Figure 7 CAPE inhibited LPS-induced upregulated expression of pro-inflammatory cytokines in BV-2 cells. Notes: (A) CAPE inhibited the LPS-induced upregulation of TNF-α, IL-1β and IL-6 in BV-2 cells. (B) CAPE inhibited the LPS-induced upregulation of TNF-α in BV-2 cells. (C) CAPE suppressed the LPS-induced upregulation of IL-1β in BV-2 cells. (D) CAPE inhibited the LPS-induced upregulation of IL-6 in BV-2 cells. The mRNA levels of TNF-α, IL-1β and IL-6 were determined using RT-PCR. GAPDH was used as an invariant control. n=4. Two-way ANOVA revealed a significant difference at *P<0.05 and **P<0.01 vs control; #P<0.05 and ##P<0.01 vs LPS-treated group. Abbreviations: CAPE, caffeic acid phenethyl ester; LPS, lipopolysaccharide; RT-PCR, real-time quantitative PCR; TNF, tumor necrosis factor. |
Discussion
In this study, our major findings were as follows: 1) CAPE significantly attenuated the development of CCI-induced mechanical allodynia in mice; 2) CAPE obviously inhibited CCI-induced microglial activation; 3) CAPE markedly inhibited CCI-induced upregulated expression of p-p38, p-p65 and proinflammatory cytokine in the spinal cord; and 4) CAPE also inhibited LPS-induced upregulated p38 MAPK phosphorylation, NF-κB nuclear translocation and proinflammatory cytokine expression in BV2 cells.
Numerous evidence indicates that microglia play a crucial role in the development and maintenance of neuropathic pain.14 Activation of microglia synthesizes and releases inflammatory factors (such as TNF-α, IL-1β and IL-6), which enhances central sensitization and nerve injury–induced persistent pain.13 Our results showed that CAPE could significantly inhibit CCI-induced activation of spinal microglia (Figure 2), being consistent with the findings of Tsai et al. They found that CAPE inhibited microglial activation after LPS injection i.p. in mice.12 In accordance with the results above, behavior tests showed that CAPE attenuated CCI-induced neuropathic pain (Figure 1B and C) effectively. These results suggest that microglial inhibition by CAPE is efficient in attenuation of neuropathic pain.
A study showed that p38 MAPK plays an important role in the process of neuropathic pain15 and activates spinal microglia after nerve injury.16 Further studies confirmed that intrathecal injection of the p38 inhibitor SB203580 significantly alleviated behaviors associated with neuropathic pain in animal models.17 Similarly, we herein found that inhibiting p-p38 by CAPE significantly inhibited the activation of microglia and reduced pain in CCI mice (Figures 1, 3 and 4). Previous studies found that p38 regulated the transcriptional activity of NF-κB.18 Activation of p38 leads to the nuclear translocation of NF-κB p65 and activation of downstream signaling pathways, leading to the development of pain.19,20 Consistent with the above studies, our research found that NF-κB p65 phosphorylation was increased in CCI mice, which was effectively reversed by CAPE treatment (Figures 3 and 4). These results suggest that CAPE can attenuate CCI-mediated activation of NF-κB, which has been implicated as the first signal to trigger the synthesis of inflammatory cytokines in microglia.
However, it must be mentioned that we could not exclude that the analgesic effect of CAPE may be partly due to inhibition of reactive oxygen species (ROS) production. Dozens of studies suggest that ROS are critically involved in the generation of pain in various painful conditions, including neuropathic and inflammatory pain.21,22 Studies have reported that vitamin E and vitamin C, which are known as powerful antioxidants, have antinociceptive effects.23 A recent study demonstrates that vitamin C+E treatment reduces p38, but not ERK (p42/p44) phosphorylation in tissue extracts from the spinal cord and dorsal root ganglion of spared nerve injury-treated animals.24 CAPE is a potent scavenger of ROS.25 Therefore, CAPE attenuates CCI-induced neuropathic pain partly via inhibition of ROS production.
In other species, CAPE also showed excellent therapeutic effects. Altuğ et al found that CAPE could provide neuroprotective effects on cerebral ischemic injury in a rabbit model of focal permanent middle cerebral artery occlusion.26 Kart et al reported that CAPE improved cisplatin-induced hepatotoxicity in rabbits.27 They believed that the role of CAPE was mainly through its antioxidant function. In addition, Tan et al’s study found a strong myocardial protective effect on acute myocardial ischemia–reperfusion injury in rabbits due to its anti-inflammatory effects.28 Our study found that CAPE could significantly alleviate neuropathic pain in CCI mice, but whether it has the same effect in humans remains to be studied. Moreover, our study only initially explored the inhibitory effect of CAPE on P38, and its specific binding site needs further study.
In addition, CAPE, a promising component of propolis, is a small lipid-soluble compound with multiple beneficial biological activities. It has been used safely since ancient times as a dietary supplement and for the treatment of various ailments.29 Studies have shown that CAPE exhibits significant beneficial biological efficacy in various target organs, such as heart,30 brain,31 skeletal muscles,32 and breast,33 with almost no reported adverse effects. Therefore, CAPE may be safe and effective for inhibiting neuropathic pain.
Conclusion
In summary, our results demonstrated that CAPE attenuates CCI-induced neuropathic pain via inhibiting p38 MAPK/NF-κB signaling and proinflammatory cytokine expression in the microglia. Our study suggests that CAPE may be a potential drug candidate for neuropathic pain treatment.
Disclosure
The authors report no conflicts of interest in this work.
References
Murnion BP. Neuropathic pain: current definition and review of drug treatment. Aust Prescr. 2018;41(3):60–63. | ||
Malcangio M. Microglia and chronic pain. Pain. 2016;157(5):1002–1003. | ||
Zhao H, Alam A, Chen Q, et al. The role of microglia in the pathobiology of neuropathic pain development: what do we know? Br J Anaesth. 2017;118(4):504–516. | ||
Guasti L, Richardson D, Jhaveri M, et al. Minocycline treatment inhibits microglial activation and alters spinal levels of endocannabinoids in a rat model of neuropathic pain. Mol Pain. 2009;5:35p. | ||
Rojewska E, Popiolek-Barczyk K, Jurga AM, Makuch W, Przewlocka B, Mika J. Involvement of pro- and antinociceptive factors in minocycline analgesia in rat neuropathic pain model. J Neuroimmunol. 2014;277(1-2):57–66. | ||
Ji RR, Suter MR. p38 MAPK, microglial signaling, and neuropathic pain. Mol Pain. 2007;3:33p. | ||
Wu J, Xu Y, Pu S, Jiang W, du D. p38/MAPK inhibitor modulates the expression of dorsal horn GABA(B) receptors in the spinal nerve ligation model of neuropathic pain. Neuroimmunomodulation. 2011;18(3):150–155. | ||
Xu JT, Xin WJ, Wei XH, et al. p38 activation in uninjured primary afferent neurons and in spinal microglia contributes to the development of neuropathic pain induced by selective motor fiber injury. Exp Neurol. 2007;204(1):355–365. | ||
Lima Cavendish R, de Souza Santos J, Belo Neto R, et al. Antinociceptive and anti-inflammatory effects of Brazilian red propolis extract and formononetin in rodents. J Ethnopharmacol. 2015;173:127–133. | ||
Armutcu F, Akyol S, Ustunsoy S, Turan FF. Therapeutic potential of caffeic acid phenethyl ester and its anti-inflammatory and immunomodulatory effects (review). Exp Ther Med. 2015;9(5):1582–1588. | ||
Tolba MF, Azab SS, Khalifa AE, Abdel-Rahman SZ, Abdel-Naim AB. Caffeic acid phenethyl ester, a promising component of propolis with a plethora of biological activities: a review on its anti-inflammatory, neuroprotective, hepatoprotective, and cardioprotective effects. IUBMB Life. 2013;65(8):699–709. | ||
Tsai CF, Kuo YH, Yeh WL, et al. Regulatory effects of caffeic acid phenethyl ester on neuroinflammation in microglial cells. Int J Mol Sci. 2015;16(3):5572–5589. | ||
Tsuda M. Microglia in the spinal cord and neuropathic pain. J Diabetes Investig. 2016;7(1):17–26. | ||
Aldskogius H, Kozlova EN. Microglia and neuropathic pain. CNS Neurol Disord Drug Targets. 2013;12(6):768–772. | ||
Kawasaki Y, Xu ZZ, Wang X, et al. Distinct roles of matrix metalloproteases in the early- and late-phase development of neuropathic pain. Nat Med. 2008;14(3):331–336. | ||
Jin SX, Zhuang ZY, Woolf CJ, Ji RR. p38 mitogen-activated protein kinase is activated after a spinal nerve ligation in spinal cord microglia and dorsal root ganglion neurons and contributes to the generation of neuropathic pain. J Neurosci. 2003;23(10):4017–4022. | ||
Tsuda M, Mizokoshi A, Shigemoto-Mogami Y, Koizumi S, Inoue K. Activation of p38 mitogen-activated protein kinase in spinal hyperactive microglia contributes to pain hypersensitivity following peripheral nerve injury. Glia. 2004;45(1):89–95. | ||
Saha RN, Jana M, Pahan K. MAPK p38 regulates transcriptional activity of NF-kappaB in primary human astrocytes via acetylation of p65. J Immunol. 2007;179(10):7101–7109. | ||
Dejardin E, Deregowski V, Chapelier M, et al. Regulation of NF-kappaB activity by I kappaB-related proteins in adenocarcinoma cells. Oncogene. 1999;18(16):2567–2577. | ||
Chen ZJ. Ubiquitin signalling in the NF-kappaB pathway. Nat Cell Biol. 2005;7(8):758–765. | ||
Kim HK, Park SK, Zhou JL, et al. Reactive oxygen species (ROS) play an important role in a rat model of neuropathic pain. Pain. 2004;111(1-2):116–124. | ||
Gao X, Kim HK, Chung JM, Chung K. Reactive oxygen species (ROS) are involved in enhancement of NMDA-receptor phosphorylation in animal models of pain. Pain. 2007;131(3):262–271. | ||
Riffel AP, de Souza JA, Santos MC, et al. Systemic administration of vitamins C and E attenuates nociception induced by chronic constriction injury of the sciatic nerve in rats. Brain Res Bull. 2016;121:169–177p. | ||
Lu R, Kallenborn-Gerhardt W, Geisslinger G, Schmidtko A. Additive antinociceptive effects of a combination of vitamin C and vitamin E after peripheral nerve injury. PLoS One. 2011;6(12):e29240. | ||
Serarslan G, Altuğ E, Kontas T, Atik E, Avci G. Caffeic acid phenethyl ester accelerates cutaneous wound healing in a rat model and decreases oxidative stress. Clin Exp Dermatol. 2007;32(6):709–715. | ||
Altuğ ME, Serarslan Y, Bal R, et al. Caffeic acid phenethyl ester protects rabbit brains against permanent focal ischemia by antioxidant action: a biochemical and planimetric study. Brain Res. 2008;1201:135–142. | ||
Kart A, Cigremis Y, Karaman M, Ozen H. Caffeic acid phenethyl ester (CAPE) ameliorates cisplatin-induced hepatotoxicity in rabbit. Exp Toxicol Pathol. 2010;62(1):45–52. | ||
Tan J, Ma Z, Han L, et al. Caffeic acid phenethyl ester possesses potent cardioprotective effects in a rabbit model of acute myocardial ischemia–reperfusion injury. Am J Physiol Heart Circ Physiol. 2005;289(5):H2265–H2271. | ||
Ishida Y, Gao R, Shah N, et al. Anticancer activity in honeybee propolis: functional insights to the role of caffeic acid phenethyl ester and its complex with γ-cyclodextrin. Integr Cancer Ther. 2018;17(3):867–873. | ||
Mansour HH, Tawfik SS. Early treatment of radiation-induced heart damage in rats by caffeic acid phenethyl ester. Eur J Pharmacol. 2012;692(1-3):46–51. | ||
Zhao J, Pati S, Redell JB, Zhang M, Moore AN, Dash PK. Caffeic acid phenethyl ester protects blood–brain barrier integrity and reduces contusion volume in rodent models of traumatic brain injury. J Neurotrauma. 2012;29(6):1209–1218. | ||
Ozyurt H, Ozyurt B, Koca K, Ozgocmen S. Caffeic acid phenethyl ester (CAPE) protects rat skeletal muscle against ischemia-reperfusion-induced oxidative stress. Vascul Pharmacol. 2007;47(2–3):108–112. | ||
Omene C, Kalac M, Wu J, Marchi E, Frenkel K, O’Connor OA. Propolis and its active component, caffeic acid phenethyl ester (CAPE), modulate breast cancer therapeutic targets via an epigenetically mediated mechanism of action. J Cancer Sci Ther. 2013;5(10):334–342. |
 © 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2018 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.