Back to Journals » Journal of Pain Research » Volume 19
Bidirectional Mendelian Randomization Analysis of 338 Cerebrospinal-Fluid Metabolites and Cluster-Headache Risk
Authors Yu D, Yang X, Zhou J, Chen W, Song J, Yu W, Huang S
Received 27 June 2025
Accepted for publication 26 December 2025
Published 8 January 2026 Volume 2026:19 550160
DOI https://doi.org/10.2147/JPR.S550160
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Rune Häckert Christensen
Danhua Yu,1,* Xuewei Yang,1,* Jinli Zhou,1,* Weiwei Chen,1 Jinhui Song,1 Weifei Yu,1 Shaokang Huang2
1Department of Neurology, Yiwu Central Hospital, Yiwu, Zhejiang Province, People’s Republic of China; 2Department of Orthopedics, Shanghai Changhai Hospital, Shanghai, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Danhua Yu, Email [email protected]
Background: Cluster headache (CH) is a rare but highly disabling primary headache disorder characterized by severe unilateral attacks and autonomic symptoms. The metabolic mechanisms underlying CH remain poorly understood.
Objective: To investigate the potential causal effects of cerebrospinal fluid (CSF) metabolite levels on CH risk, and to explore possible reverse causal effects of CH on CSF metabolites, using a bidirectional Mendelian randomization (MR) approach.
Methods: We performed a bidirectional two-sample Mendelian randomization (MR) analysis integrating genome-wide association study (GWAS) data for 338 cerebrospinal fluid (CSF) metabolites and CH (1,833 cases and 498,515 controls from FinnGen release 12). Genetic instruments were selected at P < 1× 10−5 (LD r2 < 0.01). The primary causal estimates were derived using the inverse-variance weighted (IVW) method under a random-effects model, complemented by MR-Egger, weighted median, and MR-PRESSO sensitivity tests. Multiple testing correction was performed using both Bonferroni and false discovery rate (FDR) approaches.
Results: In the forward MR analysis, 11 CSF metabolites were significantly associated with CH risk (P< 0.05). The strongest associations were observed for orotate (β = 0.53, 95% CI: 0.23– 0.82, P = 0.0006), betaine (β = 0.47, 95% CI: 0.16– 0.79, P = 0.0035), and 5-oxoproline (β = 0.57, 95% CI: 0.17– 0.97, P = 0.0053). In the reverse MR analysis, eight metabolites, including lysine (β = 0.015, P = 0.029) and kynurenine (β = 0.025, P = 0.020), were nominally associated with genetic liability to CH. Sensitivity analyses showed no evidence of directional pleiotropy or heterogeneity (all P > 0.05).
Conclusion: This bidirectional MR study provides the first genetic evidence linking central metabolic alterations to CH susceptibility. While these results highlight potential metabolic biomarkers and mechanistic pathways, the findings remain preliminary due to modest statistical power and should be replicated in larger and ethnically diverse cohorts.
Keywords: cluster headache, cerebrospinal fluid metabolites, Mendelian randomization, oxidative stress
Introduction
Cluster headache (CH) is a severe primary headache disorder classified under trigeminal autonomic cephalalgias.1 It is characterized by recurrent attacks of excruciating, unilateral pain, typically localized to the orbital or temporal region, accompanied by ipsilateral autonomic symptoms such as conjunctival injection, lacrimation, nasal congestion, and ptosis.2 Although relatively rare—affecting approximately 0.1% of the general population—CH is among the most debilitating and painful conditions known, with a striking male predominance and peak onset in early adulthood.3 The underlying pathophysiology of CH remains incompletely understood, though hypothalamic dysfunction, particularly in the posterior region, plays a central role.4 Additional mechanisms involving the trigeminal-autonomic reflex, circadian rhythm disturbances, and altered neuropeptide signaling—especially of calcitonin gene-related peptide (CGRP)—further implicate complex neurovascular and neuroinflammatory pathways.5
Recent evidence has highlighted metabolic alterations within the central nervous system in CH and related primary headache disorders. Neuroimaging and magnetic resonance spectroscopy (MRS) studies have demonstrated abnormal hypothalamic energy metabolism, elevated lactate accumulation, and disturbed glutamate–GABA homeostasis during both active and remission phases of CH.6 Similar neuroenergetic and neurotransmitter imbalances have also been reported in migraine, trigeminal neuralgia, and other chronic orofacial pain syndromes, suggesting that metabolic dysregulation may represent a shared pathophysiological mechanism across primary headache conditions.7 Despite these insights, studies directly characterizing cerebrospinal fluid (CSF) metabolite profiles in CH remain limited.
Cerebrospinal fluid, which circulates within the central nervous system, offers a unique window into brain metabolism and homeostasis.8 CSF metabolomics—the comprehensive profiling of small-molecule metabolites—has emerged as a powerful approach for biomarker discovery and mechanistic exploration in neurological and psychiatric diseases.9 These metabolites encompass diverse biochemical classes, including amino acids, neurotransmitters and their precursors (eg, glutamate, GABA), energy metabolism intermediates (eg, lactate, pyruvate), lipid derivatives, purines and pyrimidines, as well as markers of oxidative stress and neuroinflammation. CSF metabolomic signatures have been increasingly recognized as valuable biomarkers in disorders such as Alzheimer’s disease, Parkinson’s disease, and schizophrenia.10,11 However, their role in the pathophysiology of CH remains largely unexplored, representing a critical gap in current research.
Mendelian randomization (MR) is a genetic epidemiological approach that leverages germline genetic variants as instrumental variables to assess the causal effects of an exposure on an outcome.12,13 By mimicking the conditions of a randomized controlled trial, MR minimizes confounding and reverse causation inherent in traditional observational studies.14,15 One of the key strengths of MR lies in its ability to exploit the fixed nature of genetic variants established at conception, thereby allowing for the evaluation of long-term, unconfounded effects of exposures such as biomarkers, behaviors, or metabolic traits.16 With the growing availability of genome-wide association study (GWAS) summary statistics for both CSF metabolites and headache phenotypes, MR offers a robust framework to evaluate whether perturbations in central metabolic profiles causally influence CH risk.10,11 In addition to traditional “forward MR,” which estimates the effect of an exposure on a disease outcome, “reverse MR” can be employed to test whether the disease process itself causally influences levels of a specific biomarker or metabolic trait, thus helping to elucidate directionality and bidirectional relationships.
In this study, we aim to systematically evaluate the potential causal relationships between genetically predicted CSF metabolite levels and the risk of cluster headache using a two-sample MR design. By integrating genetic instruments for CSF metabolites with publicly available GWAS data for CH, we seek to identify novel metabolic biomarkers and elucidate underlying biological mechanisms, ultimately contributing to improved diagnostic and therapeutic strategies for this disabling condition.
Methods
Study Design
This study employed a two-sample MR framework to assess the potential causal relationship between CSF metabolites and the risk of CH (Figure 1). MR analysis relies on three core assumptions: (1) the genetic variants used as instrumental variables (IVs) are robustly associated with the exposure (CSF metabolite levels); (2) these variants are not associated with confounders of the exposure–outcome relationship; and (3) they influence the outcome (CH risk) exclusively through the exposure, not via alternative pathways (ie, no horizontal pleiotropy).17
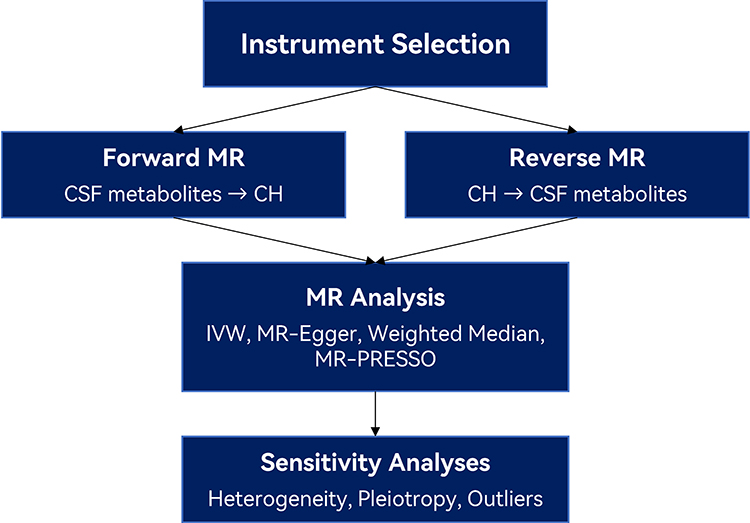 |
Figure 1 Overview of Bidirectional Mendelian Randomization Workflow. Schematic representation of the study design and analytical workflow. Genetic instruments were selected for cerebrospinal fluid (CSF) metabolites and cluster headache (CH) based on genome-wide association study (GWAS) data. Forward Mendelian randomization (MR) analysis was performed to assess the causal effect of genetically predicted CSF metabolite levels on CH risk, while reverse MR analysis tested whether CH genetic liability influenced CSF metabolite concentrations. For both directions, multiple MR methods were applied, including inverse-variance weighted (IVW), MR-Egger, weighted median, and MR-PRESSO. Sensitivity analyses were conducted to assess heterogeneity, horizontal pleiotropy, and potential outliers to ensure the robustness of the causal estimates. |
To assess potential bidirectional causality, reverse MR analyses were also performed, treating CH as the exposure and CSF metabolites as outcomes. This bidirectional design allows clarification of the directionality of observed associations and helps rule out reverse causation. As this two-sample MR analysis was based on publicly available summary-level GWAS data, individual-level clinical information—such as diagnostic confirmation details, imaging parameters, or follow-up durations—was not accessible.
Data Sources
Genetically predicted levels of 338 CSF metabolites(Table S1) were obtained from a metabolome-wide GWAS conducted in cognitively normal individuals from the Wisconsin Alzheimer’s Disease Research Center (WADRC) and the Wisconsin Registry for Alzheimer’s Prevention (WRAP). Metabolite prediction models were constructed using elastic net and other machine learning algorithms, including least absolute shrinkage and selection operator (LASSO), ridge regression, and random forest regression, to optimize predictive accuracy across diverse metabolite classes.
Only metabolites with a cross-validated predictive performance of R2 > 0.025 were retained as exposures for MR analysis. This threshold was adopted in accordance with prior large-scale CSF metabolomic GWAS studies, where it was used to ensure sufficient instrument strength while maximizing metabolite coverage.18 Although this represents a modest level of variance explained, it balances the trade-off between instrument validity and the inclusion of a comprehensive CSF metabolic profile, particularly given the typically lower heritability of CSF metabolite traits compared to plasma-based measures.
Summary-level genetic association data for CH were derived from the FinnGen consortium (Release 12), a large-scale biobank integrating genetic data with nationwide electronic health records in Finland (Table 1). The dataset included 1,833 individuals diagnosed with cluster headache (ICD-10 code G44.0) and 498,515 controls. GWAS analyses were conducted using logistic regression models, adjusting for age, sex, genotyping batch, and population structure (first 10 genetic principal components).
 |
Table 1 Data Source |
Instrument Selection
Independent single nucleotide polymorphisms (SNPs) strongly associated with each exposure (P < 1 × 10−5) were selected as instrumental variables. SNPs were clumped for linkage disequilibrium (LD r2 < 0.01, window size = 10,000 kb), and harmonization was performed to align effect alleles between exposure and outcome datasets.
Mendelian Randomization Analyses
A two-sample MR framework was implemented to investigate potential causal relationships between genetically predicted CSF metabolite levels and the risk of CH, as well as the reverse direction (CH influencing CSF metabolites). All MR analyses were performed using summary-level data.
The primary analysis used the inverse-variance weighted (IVW) method under a random-effects model. IVW provides an efficient estimate of the causal effect when all genetic instruments satisfy the core MR assumptions and there is no unbalanced horizontal pleiotropy. Causal estimates were reported as odds ratios (ORs) for binary outcomes and beta coefficients for continuous outcomes, along with 95% confidence intervals (CIs) and P-values.
To account for multiple comparisons across numerous metabolites, Bonferroni correction was applied based on the total number of tested metabolite–phenotype pairs. Associations with adjusted P-values < 0.05 were considered statistically significant. False discovery rate (FDR)–adjusted P-values (q-values) were also reported as a complementary measure of significance.
Sensitivity Analyses
To evaluate the robustness and validity of the causal estimates, several complementary MR methods were employed. MR-Egger regression was applied to assess directional pleiotropy by estimating both a causal slope and an intercept term; a non-zero intercept indicates the presence of unbalanced horizontal pleiotropy and potential violation of MR assumptions. The weighted median estimator was used to obtain consistent causal estimates even when up to 50% of the genetic instruments are invalid, assuming that the majority of the weight comes from valid instruments. MR-PRESSO (Pleiotropy RESidual Sum and Outlier) was implemented to detect and correct for horizontal pleiotropy by identifying outlier variants; the global test was used to assess overall pleiotropic distortion, and outlier-adjusted causal estimates were reported when applicable. In addition, a leave-one-out sensitivity analysis was performed, wherein each instrumental SNP was sequentially excluded to determine whether any single SNP disproportionately influenced the overall IVW estimate. Given the relatively small number of CH cases, power to detect small causal effects was limited; this should be considered when interpreting marginal associations.
Bidirectional MR Analyses
To explore potential reverse causality, reverse MR analyses were conducted using cluster headache as the exposure and each CSF metabolite as the outcome. Independent SNPs strongly associated with CH (P < 1 × 10−5, LD r2 < 0.01) were selected as instruments. The same MR methods (IVW, MR-Egger, weighted median, and MR-PRESSO) were applied to assess whether genetically predicted CH risk influenced CSF metabolite levels.
Statistical Analysis and Software
All analyses were conducted in R (version 4.3.1) using publicly available packages. Instrument selection, harmonization, and MR implementation were performed with the TwoSampleMR package, while horizontal pleiotropy and outlier detection were assessed using MR-PRESSO.
All summary statistics were aligned to ensure consistent effect alleles, and ambiguous palindromic SNPs were excluded where strand alignment was uncertain. Visualizations, including scatter plots, funnel plots, and leave-one-out influence diagnostics, were generated using ggplot2 and customized R scripts.
Results
Forward MR Analyses
In the forward two-sample MR analysis, 11 CSF metabolites showed statistically significant associations with CH risk (PIVW < 0.05) (Figure 2, Table 2 and Table S2). These findings provide preliminary genetic evidence for the involvement of central metabolic alterations in CH pathogenesis.
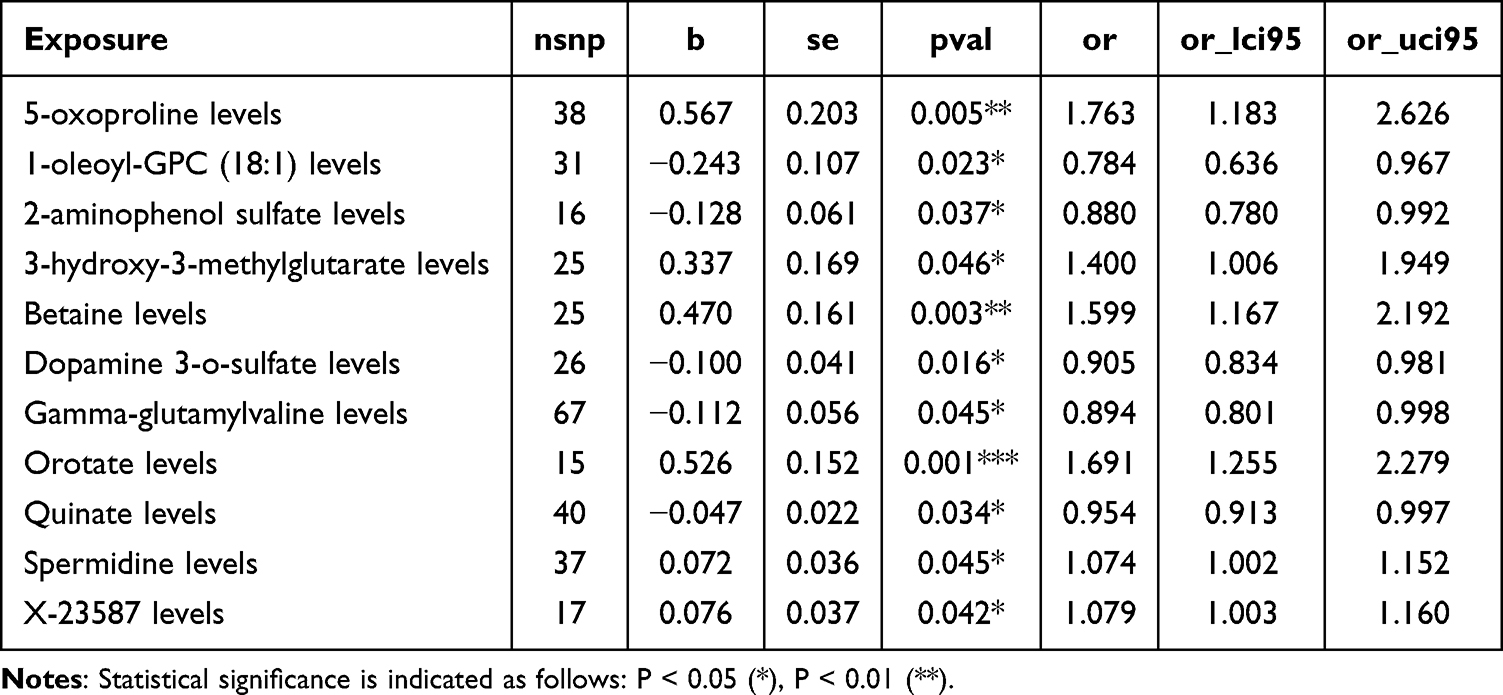 |
Table 2 Forward MR Analysis |
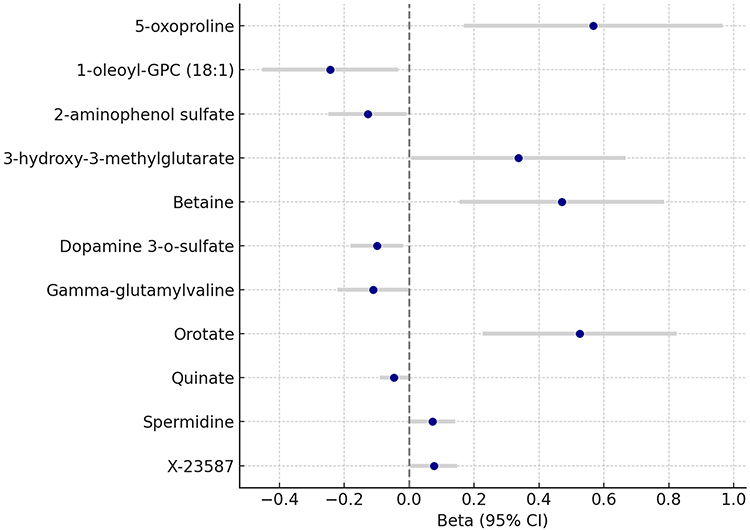 |
Figure 2 Mendelian Randomization Estimates of CSF Metabolites on Cluster Headache Risk. Forest plot showing the causal effects of 11 cerebrospinal fluid (CSF) metabolites on cluster headache (CH) risk based on forward two-sample Mendelian randomization (MR) analysis. Beta coefficients and 95% confidence intervals (CIs) were derived using the inverse-variance weighted (IVW) method. Positive effect sizes indicate that higher genetically predicted metabolite levels are associated with increased CH risk, while negative values suggest a protective effect. All associations shown reached nominal statistical significance (P_IVW < 0.05). |
Among amino acid–related metabolites, 5-oxoproline (β = 0.567, 95% CI: 0.168 to 0.966, P = 0.0053) and spermidine (β = 0.072, 95% CI: 0.002 to 0.142, P = 0.0446) were positively associated with CH risk, while gamma-glutamylvaline (β = −0.112, 95% CI: −0.222 to −0.002, P = 0.0454) and dopamine 3-O-sulfate (β = −0.100, 95% CI: −0.181 to −0.019, P = 0.0157) were negatively associated. These associations suggest potential roles for oxidative stress and neurotransmitter metabolism in CH. Lipid metabolism also emerged as relevant, with 1-oleoyl-GPC (18:1), a lysophosphatidylcholine, inversely associated with CH (β = −0.243, 95% CI: −0.453 to −0.034, P = 0.0229), implicating membrane signaling or inflammation. Energy and mitochondrial metabolites, including 3-hydroxy-3-methylglutarate (β = 0.337, 95% CI: 0.006 to 0.667, P = 0.0461) and orotate (β = 0.526, 95% CI: 0.227 to 0.824, P = 0.0006), showed strong positive associations, consistent with hypotheses of bioenergetic disruption in headache pathophysiology. Betaine, a methyl donor, was significantly associated with increased CH risk (β = 0.470, 95% CI: 0.155 to 0.785, P = 0.0035), indicating potential involvement of methylation-related pathways. Additionally, quinate (β = −0.047, 95% CI: −0.091 to −0.003, P = 0.0344) and 2-aminophenol sulfate (β = −0.128, 95% CI: −0.249 to −0.008, P = 0.0371) were inversely associated with CH, and one uncharacterized metabolite, X-23587, also demonstrated a positive association (β = 0.076, 95% CI: 0.003 to 0.148, P = 0.0415).
Reverse MR Analyses
In the reverse MR analysis, genetically predicted risk of CH was evaluated for its potential causal effect on CSF metabolite levels (Figure 3). Using CH-associated genetic variants as instrumental variables, eight CSF metabolites demonstrated statistically significant associations (PIVW < 0.05), suggesting possible downstream metabolic consequences or biomarkers of disease activity (Table 3). Notably, none of these metabolites overlapped with those identified in the forward MR analysis.
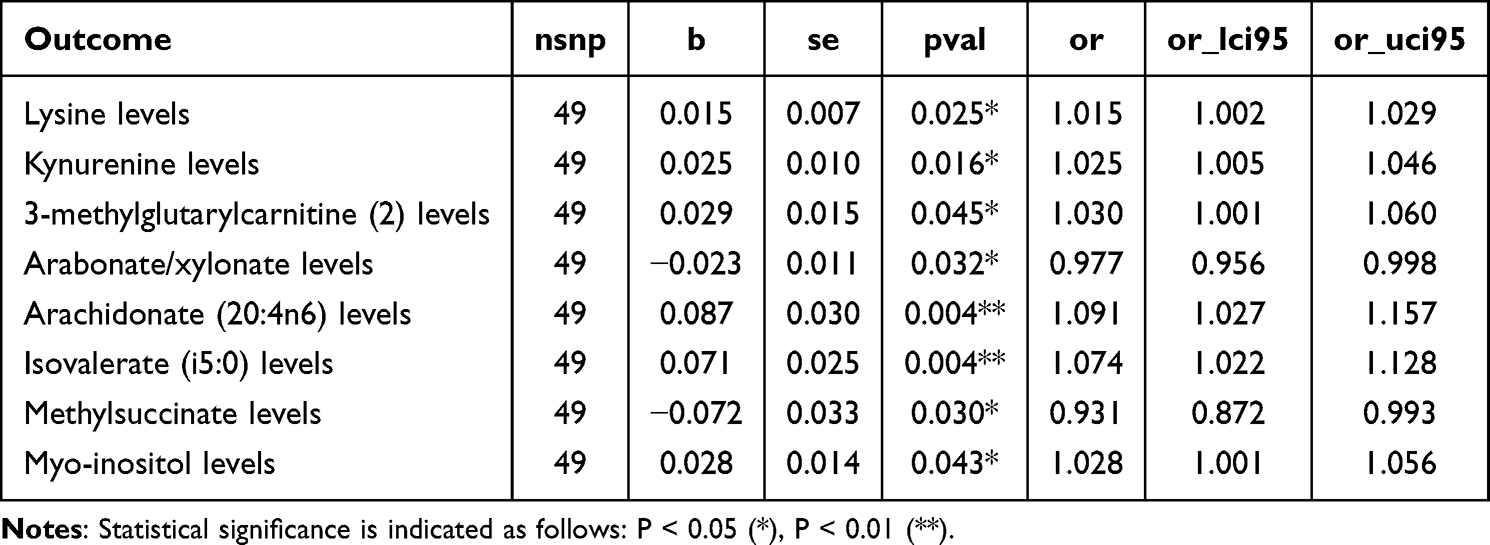 |
Table 3 Reverse MR Analysis |
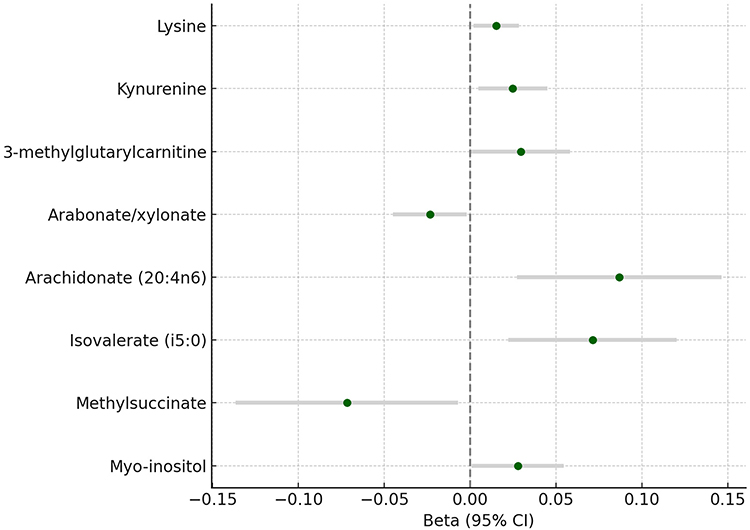 |
Figure 3 Reverse Mendelian Randomization Estimates: Effect of Cluster Headache on CSF Metabolites. Forest plot illustrating the causal effects of genetically predicted cluster headache (CH) liability on cerebrospinal fluid (CSF) metabolite levels based on reverse two-sample Mendelian randomization (MR) analysis. Beta coefficients and 95% confidence intervals (CIs) were estimated using the inverse-variance weighted (IVW) method. Positive effect sizes indicate that increased CH genetic liability is associated with higher CSF metabolite levels, while negative values indicate a potential reduction. All associations shown reached nominal significance (P_IVW < 0.05). No overlap was observed between these metabolites and those identified in the forward MR analysis. |
Several metabolites exhibited nominally significant associations in the reverse MR analysis, indicating potential downstream effects of genetically predicted CH on CSF metabolite levels. Lysine (β = 0.015, 95% CI: 0.002 to 0.028, P = 0.0245) and kynurenine (β = 0.025, 95% CI: 0.005 to 0.045, P = 0.0159) were positively associated with CH genetic liability. Arachidonate (20:4n6) (β = 0.087, 95% CI: 0.027 to 0.146, P = 0.0044) and isovalerate (i5:0) (β = 0.071, 95% CI: 0.022 to 0.120, P = 0.0044) also showed positive associations. In contrast, methylsuccinate (β = −0.072, 95% CI: −0.137 to −0.007, P = 0.0299) and arabonate/xylonate (β = −0.023, 95% CI: −0.045 to −0.002, P = 0.0323) were negatively associated with CH. 3-methylglutarylcarnitine (β = 0.029, 95% CI: 0.001 to 0.058, P = 0.0454) and myo-inositol (β = 0.028, 95% CI: 0.001 to 0.055, P = 0.0432) were also positively associated.
Sensitivity Analyses
In the forward MR analysis, multiple sensitivity tests were performed to examine the robustness of the causal estimates and to evaluate the core assumptions of Mendelian Randomization. Heterogeneity was assessed using Cochran’s Q statistic under both IVW and MR-Egger models. No substantial heterogeneity was observed for most metabolites, with Q P-values consistently above 0.20 (Table 4 and Table S3). Directional pleiotropy was evaluated using MR-Egger intercepts, all of which were non-significant (P > 0.05), indicating no evidence of unbalanced horizontal pleiotropy (Table 4 and Table S3).
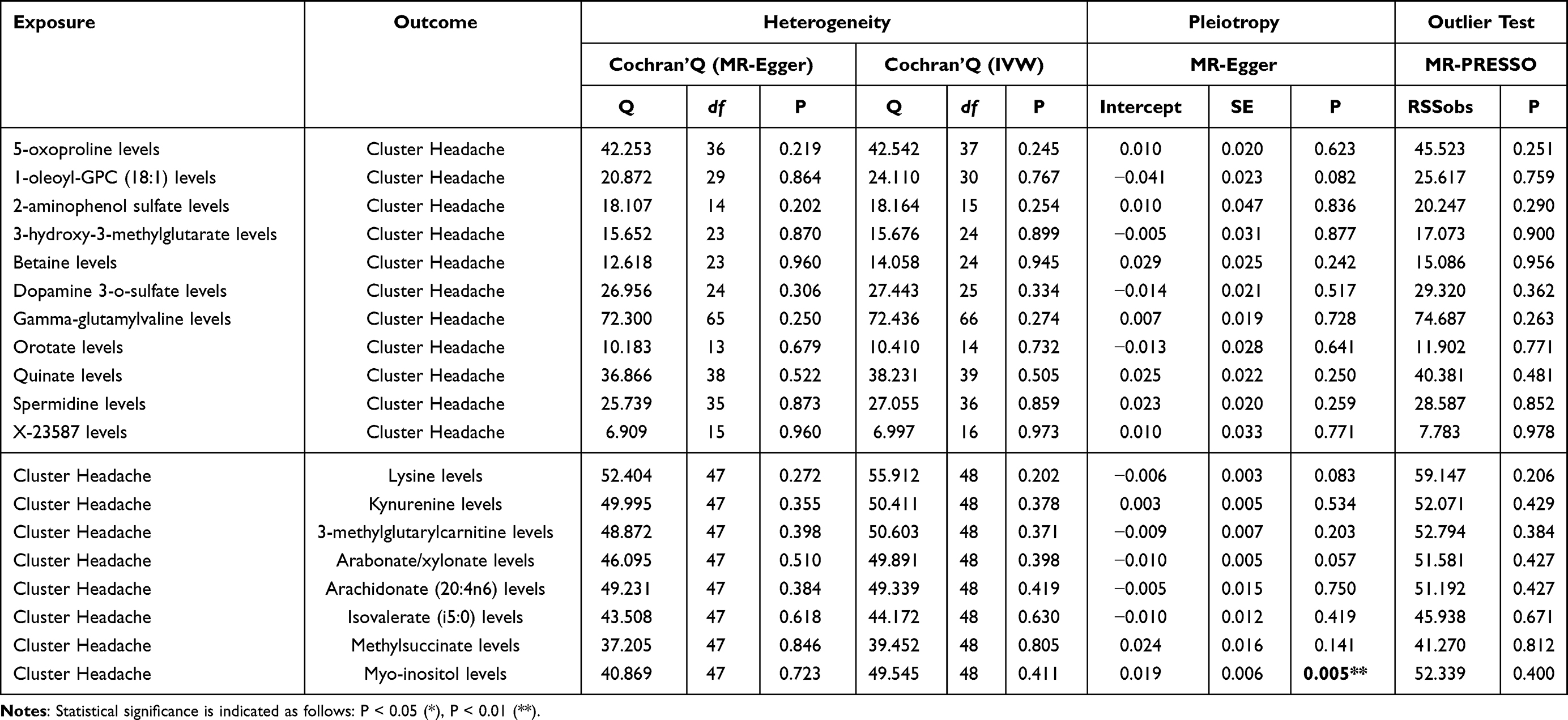 |
Table 4 Bidirectional Sensitivity Test |
Further assessment using MR-PRESSO confirmed the stability of the findings. The MR-PRESSO global test was non-significant for all traits (eg, betaine: P = 0.956; orotate: P = 0.771), and no outlier-corrected causal estimates were produced, suggesting an absence of influential variants (Table 4 and Table S4). All causal estimates (eg, 5-oxoproline: β = 0.567, P = 0.008; betaine: β = 0.470, P < 0.001; orotate: β = 0.526, P = 0.001) were therefore retained from the raw models. Together, these results confirm that the reported forward MR associations are robust and unlikely to be biased by pleiotropy or outlier effects.
In the reverse MR analysis, sensitivity analyses were similarly conducted. Cochran’s Q test did not indicate notable heterogeneity in any of the CSF metabolites tested, and all MR-Egger intercept tests were non-significant (P > 0.05), with the exception of myo-inositol, which showed a borderline significant intercept (P = 0.005), suggesting potential pleiotropic bias for this trait. Despite this, the MR-PRESSO global test for myo-inositol remained non-significant (P = 0.400), and no outliers were detected for any metabolite. All reverse MR estimates, including lysine (β = 0.015, P = 0.029), kynurenine (β = 0.025, P = 0.020), and arachidonate (β = 0.087, P = 0.006) were derived from raw models without evidence of outlier-driven distortion.
Discussion
In this bidirectional two-sample MR study, we systematically evaluated the potential causal relationships between CSF metabolite levels and the risk of CH, as well as the reverse influence of CH liability on CSF metabolic profiles. Our findings provide novel genetic evidence implicating central metabolic alterations in the pathophysiology of CH and offer new insights into potential disease mechanisms.
In the forward MR analysis, we identified 11 CSF metabolites significantly associated with CH risk. Notably, metabolites related to oxidative stress (5-oxoproline, spermidine), neurotransmission (dopamine 3-O-sulfate, gamma-glutamylvaline), membrane lipid metabolism (1-oleoyl-GPC [18:1]), mitochondrial energy metabolism (3-hydroxy-3-methylglutarate, orotate), and methyl group transfer (betaine) were implicated. These associations are biologically plausible given that oxidative stress and mitochondrial dysfunction have been long proposed as contributing factors to primary headache disorders.19 Our identification of 5-oxoproline, a marker of disrupted glutathione metabolism, complements previous studies implicating oxidative imbalance in migraine and tension-type headache, suggesting that impaired antioxidant defenses may also contribute to CH.20,21 Likewise, the observed associations with mitochondrial metabolites, such as orotate and 3-hydroxy-3-methylglutarate, are consistent with previous findings linking bioenergetic dysregulation to migraine, thereby reinforcing the hypothesis that mitochondrial dysfunction represents a shared pathogenic feature across primary headache syndromes.22,23
In contrast, the reverse MR analysis revealed no overlap between metabolites identified as causal risk factors and those influenced by CH liability, strengthening the inferred directionality of causation. Several metabolites, including lysine, kynurenine, and arachidonate (20:4n6), exhibited nominal associations with CH genetic risk. Notably, elevation of kynurenine, a tryptophan pathway metabolite involved in neuroimmune modulation, may reflect secondary inflammatory or metabolic adaptations following repeated nociceptive activation, rather than a primary pathogenic mechanism 25,26.
From a clinical perspective, these findings have important implications. First, they suggest that targeting central oxidative stress, mitochondrial dysfunction, and lipid metabolic pathways could offer new avenues for therapeutic intervention 20,27. Pharmacological agents enhancing antioxidant capacity, improving mitochondrial function, or modulating lipid-mediated inflammatory responses may hold promise in the management of CH 28,29. Moreover, by leveraging genetic instruments and a bidirectional MR framework, our study minimizes confounding and reverse causality, thereby providing a more robust foundation for biomarker and therapeutic target discovery.
Several strengths enhance the credibility of our findings. The use of a bidirectional design allowed a rigorous evaluation of causal directionality. High-quality GWAS datasets for both CSF metabolites and CH improved the precision of the causal estimates. Extensive sensitivity analyses, including MR-Egger, MR-PRESSO, and leave-one-out approaches, consistently demonstrated minimal evidence of heterogeneity or horizontal pleiotropy.
Nevertheless, certain limitations should be acknowledged. The predictive strength of genetic instruments (R2) was modest for some metabolites, potentially limiting the power to detect weaker causal effects. While multiple sensitivity analyses were applied, residual pleiotropy cannot be entirely ruled out, particularly for myo-inositol. Furthermore, the generalizability of our findings to non-European populations remains to be confirmed. Instrument strength was modest for some metabolites, and residual horizontal pleiotropy cannot be fully excluded. All GWAS data were derived from individuals of European ancestry, which may limit generalizability. The small case sample in FinnGen also constrains statistical precision. As the present MR analysis was conducted using summary-level data, clinical heterogeneity and temporal progression could not be evaluated, and causal estimates should be interpreted at the population level rather than the individual clinical scale. Diagnoses of CH were based on ICD-10 coding and could not be individually validated against ICHD-3 criteria, and information on chronic pain history among controls was unavailable. Although our study focused on metabolic mechanisms underlying CH, it is important to note that structural and anatomical factors—such as trigeminal nerve compression, impingement, and osteological variations—may also contribute to headache syndromes and modulate trigeminal-autonomic reflex pathways.24 Future studies with larger and more ethnically diverse populations are warranted to validate these findings and to explore how metabolic alterations interact with structural or neuroanatomical variations in the pathogenesis of CH.
Conclusion
In summary, this two-sample bidirectional Mendelian randomization analysis provides preliminary genetic evidence linking cerebrospinal fluid metabolites involved in oxidative stress, mitochondrial metabolism, and lipid signaling with the risk of cluster headache. While these findings identify potential biomarkers and biological pathways of interest, the translational feasibility of using CSF metabolites in clinical settings remains limited due to the invasive nature of lumbar puncture. Future studies may instead focus on identifying peripheral correlates or metabolic signatures detectable in plasma or serum.
Several limitations should also be acknowledged. The predictive strength of some genetic instruments was modest, and all datasets were derived from European populations, which may limit generalizability. Although sensitivity analyses did not reveal major pleiotropic effects, residual pleiotropy cannot be completely excluded.
Finally, our results are consistent with previous metabolomic studies implicating altered energy metabolism and oxidative imbalance in headache disorders, while extending these findings by demonstrating potential causal links in cluster headache specifically. Validation in larger and ethnically diverse cohorts is needed to substantiate these causal relationships and support translation into clinical biomarkers or therapeutic targets.
Ethics Approval
All relevant ethics approvals are from original GWASs.This study has received an exempted approval from the Ethics Committee of Yiwu Central Hospital.
Consent to Participate
This study only used publicly available summary statistics from published GWASs. No individual-level data were involved, and no additional informed consent is needed in this study.
Consent for Publication
No individual-level data were involved, and no consent for publication is needed for this study.
Acknowledgment
We thank all the consortium studies for making the summary association statistics data publicly available.
Author Contributions
All authors made a significant contribution to the work reported, whether that was in the conception, study design, execution, data acquisition, analysis and interpretation, or in all these areas. All authors took part in drafting, revising, or critically reviewing the article, approved the final version to be published, agreed on the journal to which the article has been submitted, and agree to be accountable for all aspects of the work.
Funding
This study was not supported by any funding.
Disclosure
The authors declare no conflicts of interest in this work.
References
1. Russell MB. Epidemiology and genetics of cluster headache. Lancet Neurol. 2004;3(5):279–11. doi:10.1016/S1474-4422(04)00735-5
2. Hoffmann J, May A. Diagnosis, pathophysiology, and management of cluster headache. Lancet Neurol. 2018;17(1):75–83. doi:10.1016/S1474-4422(17)30405-2
3. May A, Schwedt TJ, Magis D, Pozo-Rosich P, Evers S, Wang S-J. Cluster headache. Nat Rev Dis Primers. 2018;4(1):1–17. doi:10.1038/s41572-018-0001-z
4. Goadsby PJ. Pathophysiology of cluster headache: a trigeminal autonomic cephalgia. Lancet Neurol. 2002;1(4):251–257. doi:10.1016/S1474-4422(02)00104-7
5. Chen S-T, Wu J-W. A new era for migraine: the role of calcitonin gene-related peptide in the trigeminovascular system. Prog Brain Res. 2020;255:123–142.
6. Mohamadi M, Rojhani-Shirazi Z, Asadsangabi R, Rahimi-Jaberi A. Proton magnetic resonance spectroscopy to detect correlations between clinical symptoms and brain metabolite levels in patients with tension-type headache. J Biomed Phy Eng. 2020;10(5):583. doi:10.31661/jbpe.v0i0.1039
7. Sprenger T, Ruether K, Boecker H, et al. Altered metabolism in frontal brain circuits in cluster headache. Cephalalgia. 2007;27(9):1033–1042. doi:10.1111/j.1468-2982.2007.01386.x
8. Rainero I, Martino PD, Pinessi L. Hypocretins and primary headaches: neurobiology and clinical implications. Expert Rev Neurotherap. 2008;8(3):409–416. doi:10.1586/14737175.8.3.409
9. Lynch CJ, Adams SH. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat Rev Endocrinol. 2014;10(12):723–736. doi:10.1038/nrendo.2014.171
10. Zhao Z, Xing N, Hou L. Cerebrospinal fluid metabolites as potential biomarkers for epilepsy: insights from genome‐wide association studies. Epilepsia Open. 2025;10(1):233–242. doi:10.1002/epi4.13101
11. Luykx JJ, Bakker SC, Lentjes E, et al. Genome-wide association study of monoamine metabolite levels in human cerebrospinal fluid. Mol Psychiatry. 2014;19(2):228–234. doi:10.1038/mp.2012.183
12. Wang S, Liu Y, Wu K, Xia D, Dong X. Osteoarthritis and risk of cardiovascular diseases: a Mendelian randomization study. Injury. 2023;54(6):1569–1576. doi:10.1016/j.injury.2023.03.026
13. Wang Z, Wang Y, Xiong J, et al. Causal effects of hypertension on risk of erectile dysfunction: a two-sample Mendelian randomization study. Front Cardiovasc Med. 2023;10:1121340. doi:10.3389/fcvm.2023.1121340
14. Sekula P, Fabiola Del Greco M, Pattaro C, Köttgen A. Mendelian randomization as an approach to assess causality using observational data. J Am Soc Nephrol. 2016;27(11):3253–3265. doi:10.1681/ASN.2016010098
15. Thanassoulis G, O’Donnell CJ. Mendelian randomization: nature’s randomized trial in the post-genome era. JAMA. 2009;301(22):2386–2388. doi:10.1001/jama.2009.812
16. Burgess S, Butterworth A, Thompson SG. Mendelian randomization analysis with multiple genetic variants using summarized data. Genetic Epidemiol. 2013;37(7):658–665. doi:10.1002/gepi.21758
17. Emdin CA, Khera AV, Kathiresan S. Mendelian randomization. JAMA. 2017;318(19):1925–1926. doi:10.1001/jama.2017.17219
18. Panyard DJ, Kim KM, Darst BF, et al. Cerebrospinal fluid metabolomics identifies 19 brain-related phenotype associations. Commun Biol. 2021;4(1):63. doi:10.1038/s42003-020-01583-z
19. Borkum JM. Brain energy deficit as a source of oxidative stress in migraine: a molecular basis for migraine susceptibility. Neurochem Res. 2021;46(8):1913–1932. doi:10.1007/s11064-021-03335-9
20. Xue X, Wang M, Cui J, et al. Glutathione metabolism in ferroptosis and cancer therapy. Cancer Lett. 2025;621:217697. doi:10.1016/j.canlet.2025.217697
21. Wu G, Lupton JR, Turner ND, Fang Y-Z, Yang S. Glutathione metabolism and its implications for health. J Nutr. 2004;134(3):489–492. doi:10.1093/jn/134.3.489
22. Bohra SK, Achar R, Chidambaram S, et al. Current perspectives on mitochondrial dysfunction in migraine. Eur J Neurosci. 2022;56:3738–3754. doi:10.1111/ejn.15676
23. Lodi R, Kemp G, Pierangeli G, et al. Quantitative analysis of skeletal muscle bioenergetics and proton efflux in migraine and cluster headache. J Neurol Sci. 1997;146:73–80. doi:10.1016/S0022-510X(96)00287-0
24. Ogut E, Armagan K, Barut C. Reappraisal of the types of trigeminal porus and importance in surgical applications. Surg Radiol Anat. 2021;43(7):1169–1178. doi:10.1007/s00276-020-02651-z
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Identification of Biomarkers Associated with Oxidative Stress in Aortic Dissection Based on Bulk Transcriptome Analyses
Li Z, Li C, Shao Y, Ran H, Shi H, Zhou R, Liu X, Wu Q, Zhang C
International Journal of General Medicine 2024, 17:5633-5650
Published Date: 28 November 2024
Does Bilirubin Have a Causal Relationship With Vitiligo? A Mendelian Randomization Study and Bioinformatics Analysis
Xu D, Yin Y, Teng Y, Huang Y, Yu Y, Tao X, Fan Y, Ding X
Clinical, Cosmetic and Investigational Dermatology 2025, 18:1107-1119
Published Date: 6 May 2025
Dissecting the Causal Pathway from Herpes Zoster to Postherpetic Neuralgia: A Multi-Stage Mendelian Randomization Study Implicating VZV-Specific IgE and Cerebrospinal Fluid Metabolites
Zhang J, Chen Y, Li X, Feng C, Li J, Hu Z, Fan G, Liao X
Journal of Pain Research 2026, 19:581884
Published Date: 1 May 2026