Back to Journals » Cancer Management and Research » Volume 14
Autophagy Related Noncoding RNAs: Emerging Regulatory Factors of Gastric Cancer
Authors Lu L, Liang Q, Zhang X, Xu Y, Meng D, Liang Z ![]()
Received 2 March 2022
Accepted for publication 12 July 2022
Published 19 July 2022 Volume 2022:14 Pages 2215—2224
DOI https://doi.org/10.2147/CMAR.S364761
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Seema Singh
Ling Lu,1,* Qiaoyan Liang,2,* Xinyi Zhang,3 Yumeng Xu,3 Dehua Meng,4 Zhaofeng Liang3
1Child Healthcare Department, the Fourth Affiliated Hospital of Jiangsu University, Zhenjiang, People’s Republic of China; 2Health Care Department, People’s Liberation Army Navy No. 971 Hospital, Qingdao, People’s Republic of China; 3School of Medicine, Jiangsu University, Zhenjiang, People’s Republic of China; 4Department of Allergy, Dongtai People’s Hospital, Yancheng, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Zhaofeng Liang, School of Medicine, Jiangsu University, 301 Xuefu Road, Zhenjiang, Jiangsu, 212013, People’s Republic of China, Email [email protected] Dehua Meng, Department of Allergy, Dongtai People’s Hospital, 02 Fukang West Road, Yancheng, Jiangsu, 224200, People’s Republic of China, Email [email protected]
Abstract: Gastric cancer (GC) is one of the most common malignant cancers that seriously affect human health. Autophagy is a highly conserved self-defense mechanism found to plays an important role in the occurrence, progression, drug resistance, and prognosis of GC. Noncoding RNAs (ncRNAs) play a critical role in the occurrence and development of a variety of diseases including GC. In recent years, increasing attention has been given to research on autophagy-related ncRNAs, such as miRNA, lncRNA, and circRNA in GC. Herein, we briefly summarize the roles, functions, and the research progress of autophagy and autophagy-related ncRNAs in GC with a focus on the potential application in GC tumorigenesis, development, prognosis, and drug resistance. We also discussed prospects of clinical application, future research direction, and challenges in future research of autophagy-related ncRNAs.
Keywords: gastric cancer, autophagy, noncoding RNA, function
Introduction
Gastric cancer (GC) is one of the most common malignant tumors affecting human health. There are more than 1 million new cases and about 769,000 deaths worldwide every year.1,2 GC is usually asymptomatic in the early stage of onset, which may delay diagnosis and miss the opportunity of surgical treatment.3 Although in recent years, great progress has been made in GC diagnosis and treatment, there is still a lack of effective early diagnostic indicators, and the prognosis of advanced GC is still not optimistic. Therefore, it is particularly important to explore new pathogenesis, diagnosis, and treatment targets. Autophagy and its related non-coding RNAs (ncRNAs) have attracted more attention in the diagnosis, progress, and prognosis of GC.
Autophagy is a highly conserved self-defense mechanism in mammals. It can maintain the stability of the intracellular environment by removing and recovering misfolded and damaged proteins, and aging or damaged organelles.4,5 Moderate autophagy is particularly important for cell survival and homeostasis maintenance. Autophagy disorder has been associated with the occurrence and progression of many types of cancer. Evidence shown that autophagy plays a key role in the occurrence, development, treatment, prognosis, and drug resistance of GC.6–8 Autophagy and autophagy-related ncRNAs play a critical role in the different phases of cancer and other diseases.8–11 Recently, the role of autophagy-related miRNA, long noncoding RNAs (lncRNAs), and circular RNA (circRNA) in GC has attracted more attention.8,9,12–14 Emerging evidence shown that autophagy and its related ncRNAs have been applied as novel targets and biomarkers in the occurrence, progress, diagnosis, and prognosis of GC.
Herein, we discussed the relationship between autophagy, autophagy-related ncRNAs, and the occurrence and development of GC, with particular attention to the role, mechanism, and potential clinical application of autophagy-related ncRNA in GC. This review summarizes the role of autophagy related ncRNAs in the occurrence, development, drug resistance and prognosis of GC, which provides providing a new basis for the discovery of new diagnostic and drug resistance markers.
Autophagy and Gastric Cancer
Autophagy is a ubiquitous biological phenomenon in eukaryotic cells, which has been shown to have an important role in pathophysiological reactions and human diseases such as cancer.15 GC is a multi-factor and multi-step malignant tumor. Although the mechanism of GC is not clear, dysregulation of the balance between cell proliferation and death is a central feature, and autophagy is closely related to the carcinogenesis and progression of GC.
Autophagy plays a dual role in cancer. It may either act as a cancer suppressor by preventing the accumulation of damaged proteins and organelles, or serve as a mechanism of cell survival to promote the growth of cancer.16 Autophagy has different promoting and inhibiting effects on different types of cancers at different stages of development. For example, autophagy not only prevent tumorigenesis in the initial stage of tumors but also help cells survive in the case of nutritional deficiency.8,16 Moreover, autophagy participates in the degradation of cancer inhibitors, proapoptotic proteins, anti-proliferative factors, cytotoxic agents, radiation and hypoxia tolerance, and maintenance of Warburg effect. Concerning its protective role, autophagy promotes oncogene degradation, immunogenic apoptosis, maintenance of genomic stability and intracellular protein homeostasis, and forms “autophagic death” through radiotherapy and chemotherapy to inhibit cancer.
Autophagy is usually in a disordered state in GC. Peng et al reported that Circul2 inhibited GC cell autophagy by targeting miR-142-3p to regulate malignant transformation and cisplatin resistance of GC.14 Hu et al showed that lncRNA MALAT1, as a competitive endogenous RNA of miR-23b-3p, weakened the inhibitory effect of miR-23b-3p on ATG12, resulting in enhanced autophagy and reduced drug resistance of GC cells induced by chemotherapy.17 The imbalance of autophagy in GC cells is related to the progress of GC. KLF5 activated lncRNA DANCR and inhibited GC cell autophagy and apoptosis through the miR-194/AKT2 axis, thereby accelerating the progression of GC.18 Autophagy imbalance is significantly associated with drug resistance in many cancers including GC. It is reported that the CAGE/miR-181b-5p/S1PR1 axis promoted drug resistance of GC cells by mediating autophagy.19 In summary, autophagy and autophagy-related molecules play a critical role in the occurrence, progress, recurrence, and resistance of GC, but the exact role and molecular mechanisms still need to be explored.
Autophagy-Related Noncoding RNA and Gastric Cancer
Autophagy, as a highly conserved homeostatic pathway, is regulated by different signaling pathways (PI3K/AKT, P53, MAPK, PTEN, AMPK, etc.) and ncRNAs (miRNA, lncRNA, circRNAs).20 Autophagy-related ncRNAs contribute to the occurrence, progress, and treatment resistance of GC.
Autophagy Related miRNA and Gastric Cancer
MiRNA has recently been shown to regulate multiple steps of the autophagy process occurring in GC.21 We summarized the role of autophagy related miRNA in the occurrence, development, prognosis and drug resistance of GC (Figure 1).
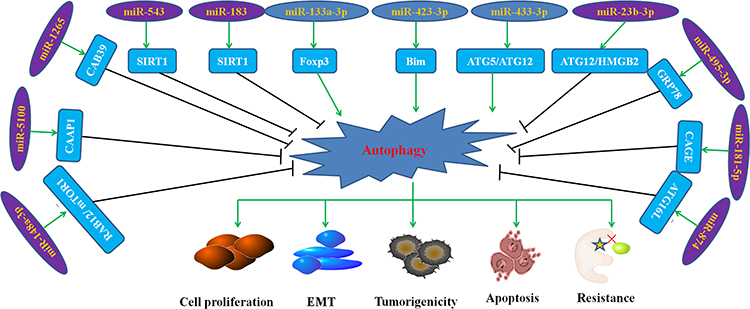 |
Figure 1 Role of representative autophagy-related miRNAs in gastric cancer. |
It was found that miR-423-3p could promote the proliferation and invasion of GC cells, was also found to be associated with abnormal autophagy and low survival of GC patients. After inhibiting autophagy, the GC-promoting effect of miR-423-3p was alleviated.22 MiR-5100 was related to the development and prognosis of GC and plays the role of a cancer suppressor gene in GC cells.23 Zhang et al found that miR-5100 could inhibit autophagy and increase the apoptosis level by targeting CAAP1 in GC cells.23 It was found that the level of miR-1265 was negatively correlated with the progression of GC. Further functional analysis revealed that miR-1265 suppresses the progression of GC by targeting CAB39, thereby impairing oncogenic autophagy in GC.24 MiR-133a-3p reduced Foxp3 expression by targeting its 3’- UTR, thereby increasing GC cell autophagy and proliferation.25 Moreover, MiR-133a-3p inhibited GC cell proliferation and metastasis by blocking autophagy-mediated glutamine decomposition.12 The increased expression of miR-543 induced by CagA is a powerful promoter for the proliferation, migration, and invasion of GC cells.26 CagA induces overexpression of miR-543, which subsequently targets SIRT1 to inhibit autophagy, thus enhancing the proliferation, migration, and invasion of GC cells. The increased expression of miR-543 inhibits GC cell autophagy, resulting in enhanced migration and invasion of GC cells.26 Zhang and his colleagues found that miR-183 affected the development of GC by regulating GC cell autophagy via the SIRT1 and PI3K/AKT/mTOR pathways.27
Chemotherapy is an important clinical treatment modality for GC. However, due to drug resistance of GC cells, especially multidrug resistance occurrence, chemotherapy usually fails. Emerging pieces of evidence indicated that autophagy and its related miRNAs are involved in the drug resistance of GC. Cisplatin and Oxaliplatin resistance are one of the main reasons for the poor prognosis of GC patients. MiR-148a-3p recombination inhibited cytoprotective autophagy and played a key role in cisplatin resistance of GC by suppressing RAB12 expression and mTOR1 activation.28 Oxidative stress-induced DNA damage repair activated autophagy is a key mechanism of oxaliplatin resistance. The findings of Wang et al illustrated that NORAD, activated by oxidative stress can actively regulate ATG5 and ATG12 by adsorbing miR-433-3p, enhancing the autophagy flux to suppress the oxidative stress, and reversing oxaliplatin resistance.29 MiRNAs are key regulators of multidrug resistance by regulating target genes and autophagy. An et al also reported that miRNA plays a critical role in GC multidrug resistance30 by demonstrating that miR-23b-3p targets ATG12/HMGB2 to regulate autophagy.30 The study of Chen et al revealed that miR-495-3p regulated the process of autophagy and inhibit multidrug resistance via the target gene GRP7831. Furthermore, the overexpression of CAGE enhances autophagy flux and invasive of GC cells. Yeon and his colleagues found that miR-181-5p negatively regulated the expression level of CAGE and autophagy flux in GC cells, and then proposed a new role of the CAGE/miR-181b-5p axis in drug-resistant and autophagy in GC.19 In other studies, the high expression of miR-874 inhibited the drug resistance of GC cells in vitro while miR-874 inhibited autophagy and sensitized GC cells to anticancer chemicals by targeting ATG16L1.32
Autophagy-associated MiRNAs have regulatory functions in GC cells and have potential diagnostic value in the early detection of GC. Many other studies have also shown that miRNA regulates the autophagy of GC cells and affect the proliferation, apoptosis, invasion, and drug resistance of GC cells33–43 as presented in Table 1. thus, autophagy-related miRNA has a good clinical application prospect in the early diagnosis, clinical treatment, and prognosis judgment in GC.
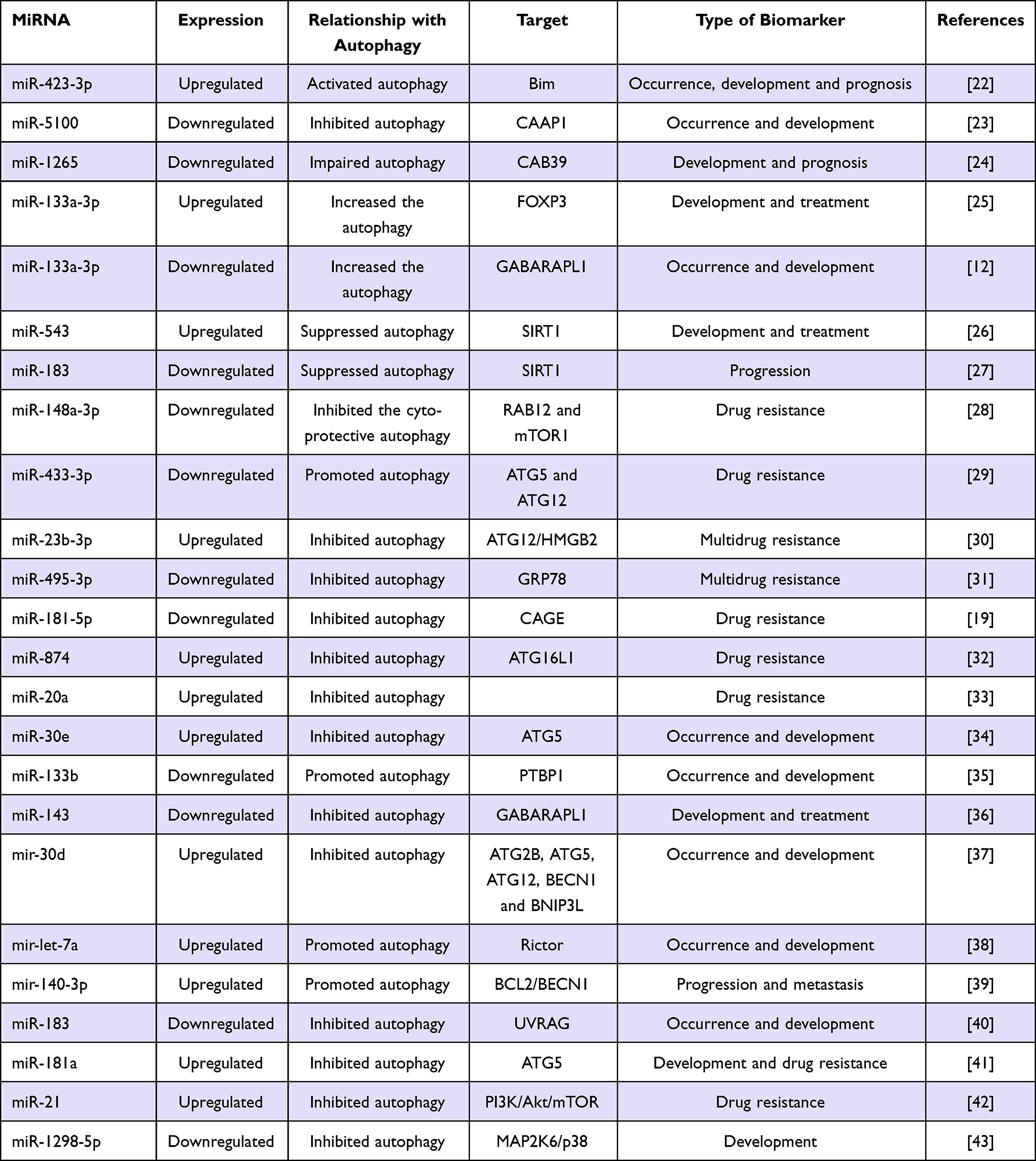 |
Table 1 Overview of the Role of Autophagy Related miRNAs in GC |
Autophagy Related lncRNA and Gastric Cancer
LncRNAs have regulatory functions on GC cells and potential diagnostic values in the early detection of GC. While GC cells autophagy is related to the occurrence and progression of GC, the role and mechanism of lncRNAs are not clear. However, a number of studies have been conducted to date, as summarized in Figure 2.
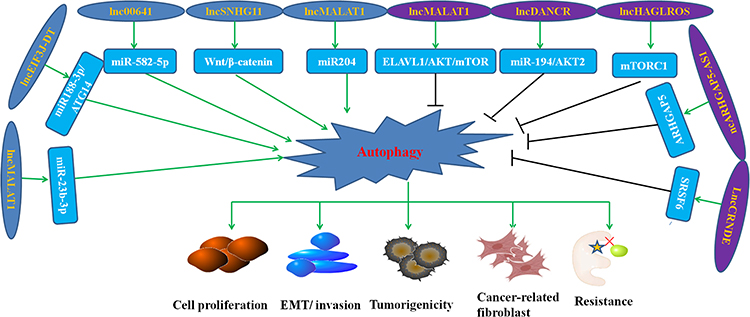 |
Figure 2 Role of representative autophagy-related lncRNAs in the progression, prognosis, and drug resistance of GC. |
It was reported that the down-regulation of lncHAGLROS significantly inhibited the proliferation, migration, and invasion of GC cells.44 Further studies found that activation of the mTORC1 signaling pathway by lncHAGLROS inhibits autophagy, thus promoting excessive proliferation and maintaining the malignant phenotype of GC cells. In another study, the overexpression of LncSNHG11 was closely related to poor prognosis of GC patients.8 The researchers found that LncSNHG11 aggravated oncogenic autophagy to facilitate the stemness, proliferation, migration, invasion, and EMT in GC by activating the Wnt/β-catenin pathway.8 Wang et al found that lncMALAT1 can promote IL-6 secretion by blocking autophagy flux in GC cells, causing the activation of normal to cancer-related fibroblast transformation, which plays an important role in the development of GC.45 In a similar study, Chen et al reported that the expressions of KLF5 and lncDANCR were up-regulated in GC tissues,18 where KLF5 activated lncDANCR, inhibited autophagy of GC cells, and accelerated the progression of GC.18 Other autophagy-related lncRNAs play important roles in the occurrence, development, and treatment of GC.46–50 These studies provide new insights into the early diagnosis and clinical treatment of GC.
Drug resistance has been considered the main obstacle in GC therapy. Elucidating the potential mechanism of drug resistance will lead to new strategies to improve the response of GC cells to chemotherapeutic drugs. It is reported that autophagy and its related lncRNAs play a critical regulatory role in the chemoresistance of GC cells. Hu et al found that drug-resistant GC cells had higher expression of lncMALAT1 and stronger autophagy.17 Further analysis showed that lncMALAT1 regulated autophagy-related drug resistance through miR-23b-3p.17 LncCRNDE was associated with the chemosensitivity of GC patients and PDX models.9 These results revealed that lncCRNDE downregulated and inhibited the autophagy flux of drug-resistant GC cells, showing the important role of lncCRNDE in GC cell autophagy regulation and drug resistance.9 Luo et al found that lncEIF3J-DT induced drug resistance of GC cells via autophagy activation by silencing miR188-3p and targeting ATG14.51 LncARHGAP5-AS1 was up-regulated in chemotherapy-resistant GC cells and correlated with poor prognosis of GC.52 The abundance of lncARHGAP5-AS1 was affected by autophagy, and inhibition of autophagy in chemotherapy-resistant cells resulted in the up-regulation of lncARHGAP5-AS1.52 In another study, lnc00641 was increased and miR-582-5p was decreased in oxaliplatin resistant GC cells. Further experiments indicated that lnc00641/miR-582-5p mediated oxaliplatin-resistance by activating autophagy.53
These observations implicate autophagy imbalance as a potential mechanism of GC cell drug resistance and lncRNA participates in regulating GC cell drug resistance (Table 2). Several other autophagy-related lncRNAs play important roles in GC cell drug resistance.54–60
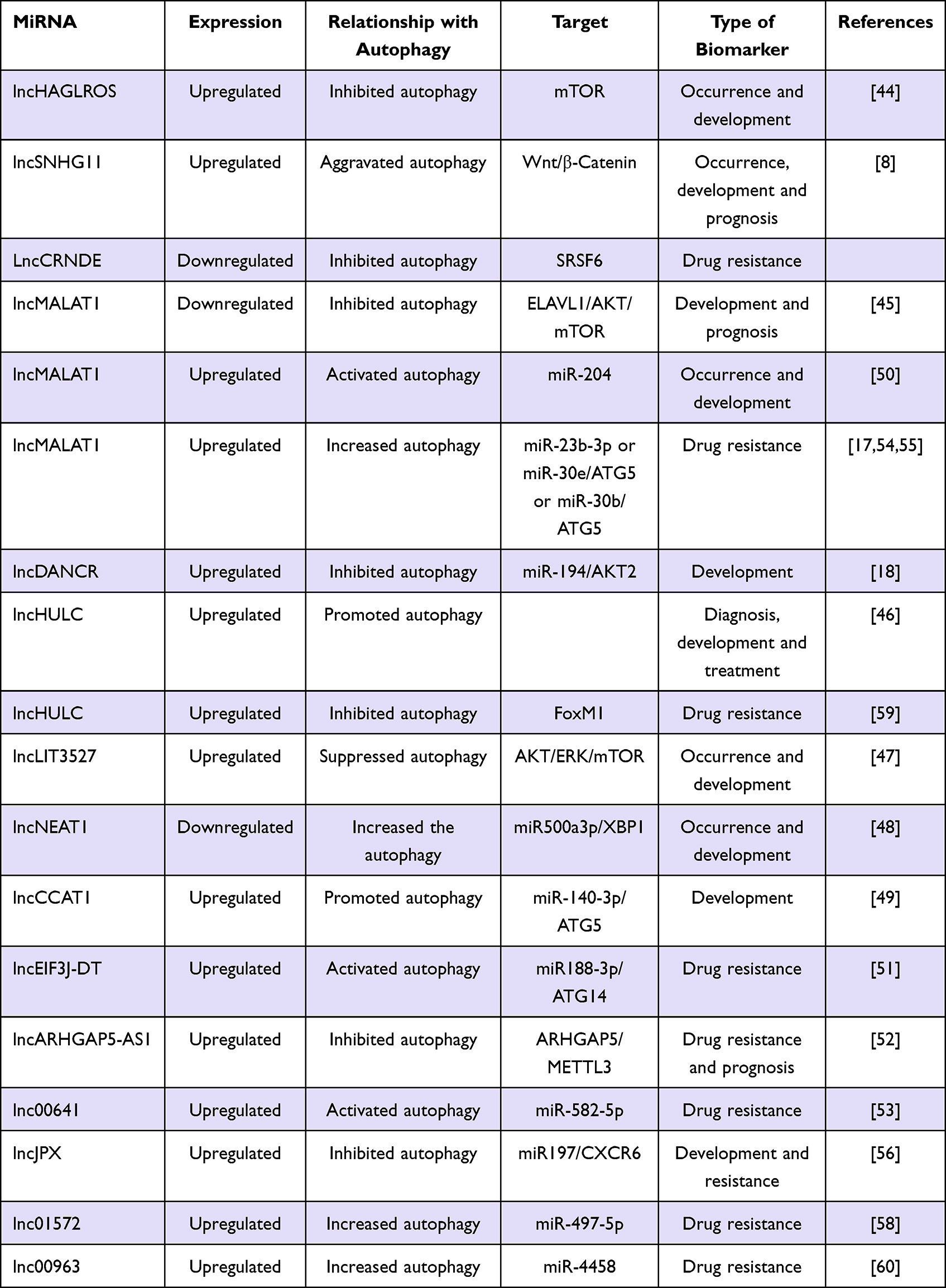 |
Table 2 Overview of the Role of Autophagy Related lncRNAs in GC |
Autophagy-Related circRNA and Gastric Cancer
CircRNA play an essential role in the occurrence and development of many diseases, including GC61,62 and are involved in autophagy in GC. The role and mechanism of autophagy-related circRNA in the occurrence, progress, and drug resistance of GC are summarized in Figure 3.
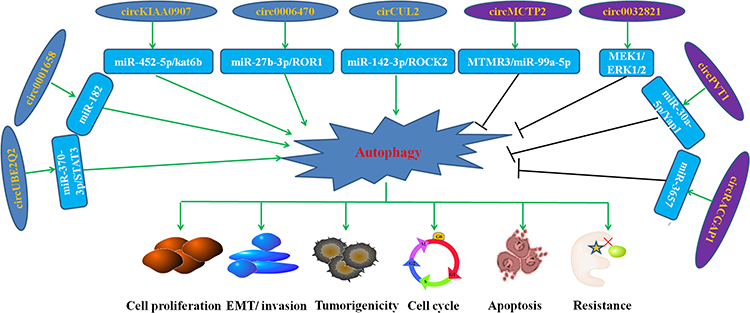 |
Figure 3 Role of representative autophagy-related circRNAs in the progression, prognosis, and drug resistance of GC. |
Knockdown of circUBE2Q2 inhibited GC cell proliferation, invasion, tumorigenicity, and increased autophagy through the circUBE2Q2/miR-370-3p/STAT3 axis.63 It is reported that circ0032821 induced the proliferation, EMT, migration, and invasion and inhibited autophagy of GC cells by activating the MEK1/ERK1/2 signaling pathway.64 CircKIAA0907 is reported to be downregulated in GC. According to Zhu et al, the upregulation of circKIAA0907 inhibits autophagy, proliferation, cell cycle, and tumorigenicity in GC cells through the miR-452-5p/kat6b axis.65 Moreover, circ0006470 promotes the proliferation, migration, and invasion of GC cells, and suppresses autophagy through targeting miR-27b-3p/ROR1.66 The level of cirCUL2 was found to decrease significantly, which otherwise was stable and located in the cytoplasm in GC tissues and cells. Peng et al found that circCUL2 may play a key role in tumor inhibition and cisplatin sensitivity regulation through miR-142-3p/ROCK2-mediated autophagy.14 Their report shown that circ0001658 regulates apoptosis and autophagy of GC cells through the miR-182/Rab-10 signal axis.67
The expression level of circMCTP2 was decreased in cisplatin-resistant GC cells and tissues. Mechanistic studies indicated that circMCTP2 sensitizes GC cells to cisplatin through MTMR3/miR-99a-5p.68 Similarly, studies have shown that exosomal circPVT1 regulates autophagy and invasion of GC cells through the miR-30a-5p/Yap1 axis, thereby promoting cisplatin resistance.69 It was found that apatinib promoted GC cell autophagy by up-regulating ATG7. Ma et al reported that the silencing of circRACGAP1 regulates autophagy by targeting miR-3657, making GC cells sensitive to apatinib.70
Autophagy-related circRNAs are expressed differently in the different stages of GC carcinogenesis, and contribute to the mechanisms involved in the occurrence and development of GC. These results suggested that autophagy-related circRNAs can be used as potential clinical markers for the occurrence, development, and drug resistance of GC (Table 3).
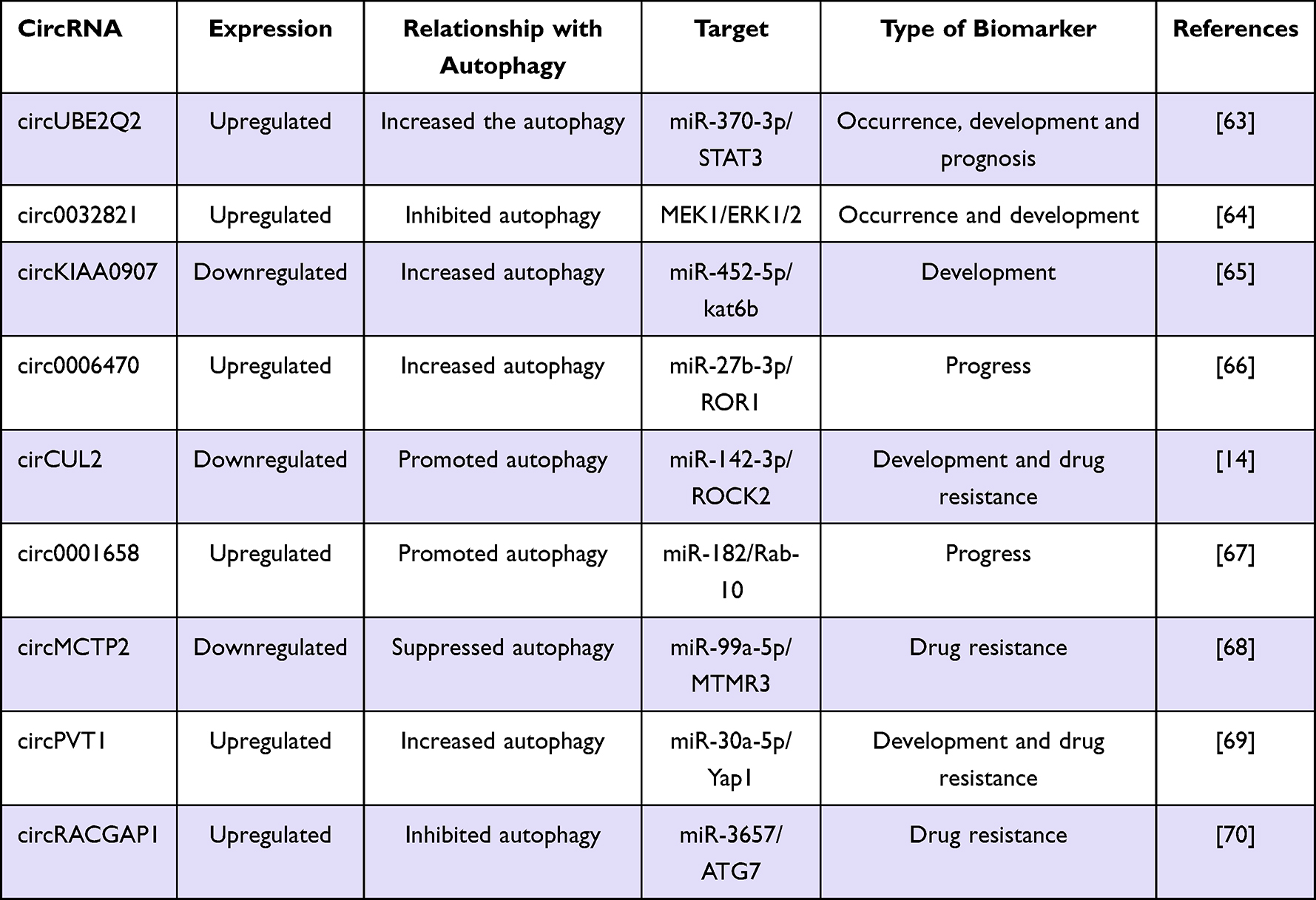 |
Table 3 Overview of the Role of Autophagy Related circRNAs in GC |
Summary and Challenges
Autophagy has been shown to play an important role in the pathogenesis of various cancers, including GC. NcRNAs play crucial roles in the occurrence, development, prevention, treatment, and drug resistance of GC. Literature indicates that miRNA, lncRNA, and circRNA regulate autophagy differently in different stages of GC and different cell states, which may enhance understanding the dual role of autophagy in GC tumorigenesis and development. Autophagy-related ncRNAs are expected to be used as potential clinical markers of GC. However, to achieve clinical application, there are still many aspects of the subject to be improved and many challenges to overcome in future research.
Firstly, the regulatory mechanism between autophagy and autophagy-related ncRNAs expression is not clear. In future studies, we believe that the mechanism of autophagy-related ncRNAs in GC may become one of the key research focuses, especially, the role of autophagy-related ncRNA derived from exosomes in GC. Secondly, clarifying the mechanism of autophagy ncRNA generation, beneficiation, and degradation may be an important link in promoting its clinical application. Besides, the clinical application of autophagy-related ncRNAs in the occurrence, progression, and drug resistance of GC needs more detailed experimental verification. The research and verification of large-scale population tissue samples, detailed safety, stability, and specificity evaluation need to be carried out before clinical application. The repeatability, specificity, and sensitivity of autophagy-related ncRNAs detection and application need to be further evaluated. In addition, it is unclear whether autophagy-related ncRNAs can affect the occurrence, progress, and drug resistance of GC by affecting GC microenvironment cells. Exosomes can carry mRNA, ncRNA, protein, and other components to participate in cell-cell communication. Whether autophagy-related ncRNAs can affect the microenvironment cells of GC through exosomes, and then play key roles in the occurrence, progress and drug resistance of GC needs to be explored.
Based on the above studies, we also speculated on the potential future development direction of autophagy and autophagy-related ncRNAs. Firstly, the development of relevant experimental methods or detection techniques, followed by the detection of autophagy and its related ncRNAs, would allow clinicians to diagnose early GC, monitor the progress of GC, and judge GC drug resistance and prognosis. It is particularly important to detect autophagy-related ncRNAs in blood or exosomes and analyze the relationship between the abnormally expressed ncRNAs and GC as a means of achieving liquid biopsy application in diagnostics and prognosis. Finally, as potential therapeutic targets of GC, autophagy-related ncRNAs may help prolong and improve the quality of life of GC patients through the convenience and repeatability of detection techniques, and the specificity, sensitivity, and effectiveness of the biomarkers.
Acknowledgments
This work was supported by grants from the National Natural Science Foundation of China (No. 81602883), the project of social development in Zhenjiang (No. SH2021045, FZ2019038), “Jinshan Doctor” medical field talent training plan of Zhenjiang, the Foundation for Excellent Young Teachers of Jiangsu University, and Clinical Medical Science and Technology Development Foundation of Jiangsu University (No. JLY2021013).
Disclosure
All authors of this study declare no conflict of interest.
References
1. Arnold M, Abnet CC, Neale RE, et al. Global Burden of 5 Major Types of Gastrointestinal Cancer. Gastroenterology. 2020;159(1):335–349 e315.
2. Sung H, Ferlay J, Siegel RL, et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021;71(3):209–249.
3. Joshi SS, Badgwell BD. Current treatment and recent progress in gastric cancer. CA Cancer J Clin. 2021;71(3):264–279.
4. Saha S, Panigrahi DP, Patil S, Bhutia SK. Autophagy in health and disease: a comprehensive review. Biomed Pharmacother. 2018;104:485–495.
5. Denton D, Kumar S. Autophagy-dependent cell death. Cell Death Differ. 2019;26(4):605–616.
6. Cao Y, Luo Y, Zou J, et al. Autophagy and its role in gastric cancer. Clin Chim Acta. 2019;489:10–20.
7. Li GM, Li L, Li MQ, et al. DAPK3 inhibits gastric cancer progression via activation of ULK1-dependent autophagy. Cell Death Differ. 2021;28(3):952–967.
8. Wu Q, Ma J, Wei J, Meng W, Wang Y, Shi M. lncRNA SNHG11 Promotes Gastric Cancer Progression by Activating the Wnt/beta-Catenin Pathway and Oncogenic Autophagy. Mol Therapy. 2021;29(3):1258–1278.
9. Zhang F, Wang H, Yu J, et al. LncRNA CRNDE attenuates chemoresistance in gastric cancer via SRSF6-regulated alternative splicing of PICALM. Mol Cancer. 2021;20(1):6.
10. Hall DP, Cost NG, Hegde S, et al. TRPM3 and miR-204 establish a regulatory circuit that controls oncogenic autophagy in clear cell renal cell carcinoma. Cancer Cell. 2014;26(5):738–753.
11. Chen L, He M, Zhang M, et al. The Role of non-coding RNAs in colorectal cancer, with a focus on its autophagy. Pharmacol Ther. 2021;226:107868.
12. Zhang X, Li Z, Xuan Z, et al. Novel role of miR-133a-3p in repressing gastric cancer growth and metastasis via blocking autophagy-mediated glutaminolysis. J Exp Clin Cancer Res. 2018;37(1):320.
13. Zhang X, Wang S, Wang H, et al. Circular RNA circNRIP1 acts as a microRNA-149-5p sponge to promote gastric cancer progression via the AKT1/mTOR pathway. Mol Cancer. 2019;18(1):20.
14. Peng L, Sang H, Wei S, et al. circCUL2 regulates gastric cancer malignant transformation and cisplatin resistance by modulating autophagy activation via miR-142-3p/ROCK2. Mol Cancer. 2020;19(1):156.
15. Klionsky DJ, Petroni G, Amaravadi RK, et al. Autophagy in major human diseases. EMBO J. 2021;40(19):e108863.
16. Das J, Chakraborty S, Maiti TK. Mechanical stress-induced autophagic response: a cancer-enabling characteristic? Semin Cancer Biol. 2020;66:101–109.
17. YiRen H, YingCong Y, Sunwu Y, et al. Long noncoding RNA MALAT1 regulates autophagy associated chemoresistance via miR-23b-3p sequestration in gastric cancer. Mol Cancer. 2017;16(1):174.
18. Cheng Z, Liu G, Huang C, Zhao X. KLF5 activates lncRNA DANCR and inhibits cancer cell autophagy accelerating gastric cancer progression. NPJ Genomic Medicine. 2021;6(1):75.
19. Yeon M, Kim Y, Pathak D, et al. The CAGE-MiR-181b-5p-S1PR1 Axis Regulates Anticancer Drug Resistance and Autophagy in Gastric Cancer Cells. Front Cell Dev Biol. 2021;9:666387.
20. Xu JL, Yuan L, Tang YC, et al. The Role of Autophagy in Gastric Cancer Chemoresistance: friend or Foe? Front Cell Dev Biol. 2020;8:621428.
21. Pourhanifeh MH, Vosough M, Mahjoubin-Tehran M, et al. Autophagy-related microRNAs: possible regulatory roles and therapeutic potential in and gastrointestinal cancers. Pharmacol Res. 2020;161:105133.
22. Kong P, Zhu X, Geng Q, et al. The microRNA-423-3p-Bim Axis Promotes Cancer Progression and Activates Oncogenic Autophagy in Gastric Cancer. Mol Therapy. 2017;25(4):1027–1037.
23. Zhang HM, Li H, Wang GX, et al. MKL1/miR-5100/CAAP1 loop regulates autophagy and apoptosis in gastric cancer cells. Neoplasia. 2020;22(5):220–230.
24. Xu Z, Li Z, Wang W, et al. MIR-1265 regulates cellular proliferation and apoptosis by targeting calcium binding protein 39 in gastric cancer and, thereby, impairing oncogenic autophagy. Cancer Lett. 2019;449:226–236.
25. Li JP, Zhang HM, Liu MJ, et al. miR-133a-3p/FOXP3 axis regulates cell proliferation and autophagy in gastric cancer. J Cell Biochem. 2020;121(5–6):3392–3405.
26. Shi Y, Yang Z, Zhang T, Shen L, Li Y, Ding S. SIRT1-targeted miR-543 autophagy inhibition and epithelial-mesenchymal transition promotion in Helicobacter pylori CagA-associated gastric cancer. Cell Death Dis. 2019;10(9):625.
27. Li H, He C, Wang X, Wang H, Nan G, Fang L. MicroRNA-183 affects the development of gastric cancer by regulating autophagy via MALAT1-miR-183-SIRT1 axis and PI3K/AKT/mTOR signals. Artif Cells, Nanomed Biotechnol. 2019;47(1):3163–3171.
28. Li B, Wang W, Li Z, et al. MicroRNA-148a-3p enhances cisplatin cytotoxicity in gastric cancer through mitochondrial fission induction and cyto-protective autophagy suppression. Cancer Lett. 2017;410:212–227.
29. Wang J, Sun Y, Zhang X, et al. Oxidative stress activates NORAD expression by H3K27ac and promotes oxaliplatin resistance in gastric cancer by enhancing autophagy flux via targeting the miR-433-3p. Cell Death Dis. 2021;12(1):90.
30. An Y, Zhang Z, Shang Y, et al. miR-23b-3p regulates the chemoresistance of gastric cancer cells by targeting ATG12 and HMGB2. Cell Death Dis. 2015;6:e1766.
31. Chen S, Wu J, Jiao K, et al. MicroRNA-495-3p inhibits multidrug resistance by modulating autophagy through GRP78/mTOR axis in gastric cancer. Cell Death Dis. 2018;9(11):1070.
32. Huang H, Tang J, Zhang L, Bu Y, Zhang X. miR-874 regulates multiple-drug resistance in gastric cancer by targeting ATG16L1. Int J Oncol. 2018;53(6):2769–2779.
33. Xin L, Zhou LQ, Liu L, Yuan YW, Zhang HT, Zeng F. METase promotes cell autophagy via promoting SNHG5 and suppressing miR-20a in gastric cancer. Int J Biol Macromol. 2019;122:1046–1052.
34. Ye Y, Fang Y, Xu W, Wang Q, Zhou J, Lu R. 3,3’-Diindolylmethane induces anti-human gastric cancer cells by the miR-30e-ATG5 modulating autophagy. Biochem Pharmacol. 2016;115:77–84.
35. Sugiyama T, Taniguchi K, Matsuhashi N, et al. MiR-133b inhibits growth of human gastric cancer cells by silencing pyruvate kinase muscle-splicer polypyrimidine tract-binding protein 1. Cancer Sci. 2016;107(12):1767–1775.
36. Du F, Feng Y, Fang J, Yang M. MicroRNA-143 enhances chemosensitivity of Quercetin through autophagy inhibition via target GABARAPL1 in gastric cancer cells. Biomed Pharmacother. 2015;74:169–177.
37. Yang XJ, Si RH, Liang YH, et al. Mir-30d increases intracellular survival of Helicobacter pylori through inhibition of autophagy pathway. World j Gastroenterol. 2016;22(15):3978–3991.
38. Fan H, Jiang M, Li B, et al. MicroRNA-let-7a regulates cell autophagy by targeting Rictor in gastric cancer cell lines MGC-803 and SGC-7901. Oncol Rep. 2018;39(3):1207–1214.
39. Chen J, Cai S, Gu T, Song F, Xue Y, Sun D. MiR-140-3p Impedes Gastric Cancer Progression and Metastasis by Regulating BCL2/BECN1-Mediated Autophagy. Onco Targets Ther. 2021;14:2879–2892.
40. Yuan Y, Zhang Y, Han L, Sun S, Shu Y. miR-183 inhibits autophagy and apoptosis in gastric cancer cells by targeting ultraviolet radiation resistance-associated gene. Int J Mol Med. 2018;42(6):3562–3570.
41. Zhao J, Nie Y, Wang H, Lin Y. MiR-181a suppresses autophagy and sensitizes gastric cancer cells to cisplatin. Gene. 2016;576(2 Pt 2):828–833.
42. Gu Y, Fei Z, Zhu R. miR-21 modulates cisplatin resistance of gastric cancer cells by inhibiting autophagy via the PI3K/Akt/mTOR pathway. Anticancer Drugs. 2020;31(4):385–393.
43. Li X, Zhu M, Zhao G, et al. MiR-1298-5p level downregulation induced by Helicobacter pylori infection inhibits autophagy and promotes gastric cancer development by targeting MAP2K6. Cell Signal. 2022;93:110286.
44. Chen JF, Wu P, Xia R, et al. STAT3-induced lncRNA HAGLROS overexpression contributes to the malignant progression of gastric cancer cells via mTOR signal-mediated inhibition of autophagy. Mol Cancer. 2018;17(1):6.
45. Wang Z, Wang X, Zhang T, et al. LncRNA MALAT1 promotes gastric cancer progression via inhibiting autophagic flux and inducing fibroblast activation. Cell Death Dis. 2021;12(4):368.
46. Zhao Y, Guo Q, Chen J, Hu J, Wang S, Sun Y. Role of long non-coding RNA HULC in cell proliferation, apoptosis and tumor metastasis of gastric cancer: a clinical and in vitro investigation. Oncol Rep. 2014;31(1):358–364.
47. Wang B, Deng Y, Jin J, Wu Y, Shen L. Long Noncoding RNA LIT3527 Knockdown induces Apoptosis and Autophagy through inhibiting mTOR pathway in Gastric Cancer Cells. J Cancer. 2021;12(16):4901–4911.
48. Zhou Y, Sha Z, Yang Y, Wu S, Chen H. lncRNA NEAT1 regulates gastric carcinoma cell proliferation, invasion and apoptosis via the miR500a3p/XBP1 axis. Mol Med Rep. 2021;24:1.
49. Yang F, Peng ZX, Ji WD, et al. LncRNA CCAT1 Upregulates ATG5 to Enhance Autophagy and Promote Gastric Cancer Development by Absorbing miR-140-3p. Dig Dis Sci. 2021;1:485.
50. Shao G, Zhao Z, Zhao W, et al. Long non-coding RNA MALAT1 activates autophagy and promotes cell proliferation by downregulating microRNA-204 expression in gastric cancer. Oncol Lett. 2020;19(1):805–812.
51. Luo Y, Zheng S, Wu Q, et al. Long noncoding RNA (lncRNA) EIF3J-DT induces chemoresistance of gastric cancer via autophagy activation. Autophagy. 2021;1:1–19.
52. Zhu L, Zhu Y, Han S, et al. Impaired autophagic degradation of lncRNA ARHGAP5-AS1 promotes chemoresistance in gastric cancer. Cell Death Dis. 2019;10(6):383.
53. Hu Y, Su Y, Lei X, et al. LINC00641/miR-582-5p mediate oxaliplatin resistance by activating autophagy in gastric adenocarcinoma. Sci Rep. 2020;10(1):14981.
54. Zhang YF, Li CS, Zhou Y, Lu XH. Propofol facilitates cisplatin sensitivity via lncRNA MALAT1/miR-30e/ATG5 axis through suppressing autophagy in gastric cancer. Life Sci. 2020;244:117280.
55. Xi Z, Si J, Nan J. LncRNA MALAT1 potentiates autophagyassociated cisplatin resistance by regulating the microRNA30b/autophagy related gene 5 axis in gastric cancer. Int J Oncol. 2019;54(1):239–248.
56. Han X, Liu Z. Long noncoding RNA JPX promotes gastric cancer progression by regulating CXCR6 and autophagy via inhibiting miR197. Mol Med Rep. 2021;23(1):548.
57. Li Z, Lu M, Zhou Y, et al. Role of Long Non-Coding RNAs in the Chemoresistance of Gastric Cancer: a Systematic Review. Onco Targets Ther. 2021;14:503–518.
58. Song Z, Jia N, Li W, Zhang XY. LINC01572 Regulates Cisplatin Resistance in Gastric Cancer Cells by Mediating miR-497-5p. Onco Targets Ther. 2020;13:10877–10887.
59. Xin L, Zhou Q, Yuan YW, et al. METase/lncRNA HULC/FoxM1 reduced cisplatin resistance in gastric cancer by suppressing autophagy. J Cancer Res Clin Oncol. 2019;145(10):2507–2517.
60. Hou M, Li C, Dong S. LINC00963/miR-4458 regulates the effect of oxaliplatin in gastric cancer by mediating autophagic flux through targeting of ATG16L1. Sci Rep. 2021;11(1):20951.
61. Chen LL. The expanding regulatory mechanisms and cellular functions of circular RNAs. Nat Rev Mol Cell Biol. 2020;21(8):475–490.
62. Kristensen LS, Andersen MS, Stagsted LVW, Ebbesen KK, Hansen TB, Kjems J. The biogenesis, biology and characterization of circular RNAs. Nat Rev Genet. 2019;20(11):675–691.
63. Yang J, Zhang X, Cao J, et al. Circular RNA UBE2Q2 promotes malignant progression of gastric cancer by regulating signal transducer and activator of transcription 3-mediated autophagy and glycolysis. Cell Death Dis. 2021;12(10):910.
64. Jiang Y, Zhang Y, Chu F, Xu L, Wu H. Circ_0032821 acts as an oncogene in cell proliferation, metastasis and autophagy in human gastric cancer cells in vitro and in vivo through activating MEK1/ERK1/2 signaling pathway. Cancer Cell Int. 2020;20:74.
65. Zhu L, Wang C, Lin S, Zong L. CircKIAA0907 Retards Cell Growth, Cell Cycle, and Autophagy of Gastric Cancer In Vitro and Inhibits Tumorigenesis In Vivo via the miR-452-5p/KAT6B Axis. Med Sci Monitor. 2020;26:e924160.
66. Cui Y, Cao J, Huang S, et al. circRNA_0006470 promotes the proliferation and migration of gastric cancer cells by functioning as a sponge of miR-27b-3p. Neoplasma. 2021.
67. Duan X, Yu X, Li Z. Circular RNA hsa_circ_0001658 regulates apoptosis and autophagy in gastric cancer through microRNA-182/Ras-related protein Rab-10 signaling axis. Bioengineered. 2022;13(2):2387–2397.
68. Sun G, Li Z, He Z, et al. Circular RNA MCTP2 inhibits cisplatin resistance in gastric cancer by miR-99a-5p-mediated induction of MTMR3 expression. J Exp Clin Cancer Res. 2020;39(1):246.
69. Yao W, Guo P, Mu Q, Wang Y. Exosome-Derived Circ-PVT1 Contributes to Cisplatin Resistance by Regulating Autophagy, Invasion, and Apoptosis Via miR-30a-5p/YAP1 Axis in Gastric Cancer Cells. Cancer Biother Radiopharm. 2021;36(4):347–359.
70. Ma L, Wang Z, Xie M, et al. Silencing of circRACGAP1 sensitizes gastric cancer cells to apatinib via modulating autophagy by targeting miR-3657 and ATG7. Cell Death Dis. 2020;11(3):169.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Solanine Represses Gastric Cancer Growth by Mediating Autophagy Through AAMDC/MYC/ATF4/Sesn2 Signaling Pathway
Tang X, Guo Y, Zhang S, Wang X, Teng Y, Jin Q, Jin Q, Shen W, Wang R
Drug Design, Development and Therapy 2023, 17:389-402
Published Date: 8 February 2023