Back to Journals » Journal of Inflammation Research » Volume 14
Associations of Single Nucleotide Polymorphisms in IL-18 Gene with Plasmodium falciparum-Associated Malaria
Authors AlRuwaisan AU, Al-Anazi MR, Shafeai MI, Rudiny FH, Motaen AM, Bin Dajem SM, Alothaid H ![]() , Morsy K, Alkahtani S
, Morsy K, Alkahtani S ![]() , Al-Qahtani AA
, Al-Qahtani AA ![]()
Received 7 April 2021
Accepted for publication 23 June 2021
Published 26 July 2021 Volume 2021:14 Pages 3587—3619
DOI https://doi.org/10.2147/JIR.S314638
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Ning Quan
Alaa U AlRuwaisan,1 Mashael R Al-Anazi,2 Mohammed I Shafeai,3 Fuad H Rudiny,3 Ali M Motaen,3 Saad M Bin Dajem,4 Hani Alothaid,5 Kareem Morsy,4,6 Saad Alkahtani,1 Ahmed A Al-Qahtani2,7
1Zoology Department, College of Science, King Saud University, Riyadh, Saudi Arabia; 2Department of Infection and Immunity, Research Centre, King Faisal Specialist Hospital & Research Centre, Riyadh, Saudi Arabia; 3Sabya General Hospital, Sabya, Saudi Arabia; 4Department of Biology, College of Science, King Khalid University, Abha, Saudi Arabia; 5Department of Basic Sciences, Faculty of Applied Medical Sciences, Al-Baha University, Al-Baha, Saudi Arabia; 6Zoology Department, Faculty of Science, Cairo University, Cairo, Egypt; 7Department of Microbiology and Immunology, College of Medicine, Alfaisal University, Riyadh, Saudi Arabia
Correspondence: Ahmed A Al-Qahtani Email [email protected]
Introduction: Interleukin-18 (IL-18) is a pro-inflammatory cytokine, reported to be involved in the initial immune responses against malaria. Genetic variations in the host are an important factor that influences the etiology of malaria at several disease levels. Polymorphisms within the IL-18 gene are associated with susceptibility and clinical outcome of several diseases.
Methods: We genotyped single nucleotide polymorphisms (SNPs) in IL-18 of patients infected with Plasmodium falciparum with varying extent of parasitemia and different age groups.
Results: SNPs rs5744292 (OR = 70.446; 95% CI = 4.318– 1149.323; p< 0.0001) and rs544354 (OR = 1.498; 95% CI = 1.088– 2.063; p=0.013) were found to be significantly associated with parasitemia in P. falciparum-infected patients when compared with healthy control subjects. SNP rs5744292 (OR = 7.597; 95% CI=1.028– 56.156; p=0.019) was associated with increased parasite density in infected patients. SNPs rs544354 (OR 0.407; 95% CI=0.204– 0.812; p = 0.009) and rs360714 (OR of 0.256; 95% CI=0.119– 0.554; p = 0.001) were significantly associated with parasite density in an age-dependent manner, with the risk alleles present more frequently among the younger (1– 9 years) patients. Several haplotypes were found to have a significant association with parasitemia. In-vitro expression analysis using luciferase reporter assay showed that SNPs rs1946518 and rs187238 in the IL-18 gene promoter region and rs360728 and rs5744292 in the 3ʹ-untranslated region of the IL-18 gene were associated with enhanced transcriptional activity.
Conclusion: Our results suggest that polymorphisms within the IL-18 gene are associated with the susceptibility to P. falciparum infection and related parasitemia among groups with different parasite density and across various age groups.
Keywords: Plasmodium falciparum, IL-18 gene, SNPs
Introduction
Malaria is a parasitic disease caused by the unicellular protozoan of the genus Plasmodium and is spread mainly through infected female Anopheles mosquitoes. Despite several control programs, malaria continues to be a major health concern worldwide, particularly in low and middle-income countries. The WHO estimates there were 228 million cases of malaria globally resulting in 405,000 deaths in 2018.1 Malaria is presently widespread along the tropical and sub-tropical belt, in certain areas of the Americas, many parts of Asia, and much of Africa, particularly sub-Saharan Africa.2 Malaria has been known to be endemic in the lowlands of Saudi Arabia, and three species of Anopheles mosquitoes have been identied in this region, namely An. arabiensis, An. Sergentii and An. Gambiae.3,4 Among the different species of Plasmodium infecting humans, Plasmodium falciparum is the most prevalent in African countries and Saudi Arabia and it is linked to most of the malaria-related deaths worldwide.5,6
Large variation has been observed in the clinical presentation of the P. falciparum infection, from asymptomatic individuals to patients with severe cerebral malaria and other fatal forms of the infection. The individual variations in the clinical manifestation and progression to severe forms of P. falciparum infection have been reported to be influenced by several factors such as age, genetic polymorphisms in the host, genotype of the parasite, and the environmental/social conditions.7,8 Numerous studies have explored genetic influence in the pathomechanism and identified several host genes that can significantly influence the diverse clinical outcomes of malaria such as hemoglobin variants,9,10 glucose-6-phosphate dehydrogenase,11 pyruvate kinase,12 and haptoglobin.13
Cytokines orchestrate the initial immune response against invading malarial parasites. Previous studies have reported an increased concentration of pro-inflammatory cytokines like IL-6 and anti-inflammatory cytokines like IL-10 in P. falciparum-infected patients.14 IL-18 is a pro-inflammatory cytokine encoded by the IL-18 gene on chromosome 11q23.1 and belongs to the IL-1 superfamily. IL-18 is expressed by a wide range of cells, including macrophages, Kupffer cells, keratinocytes, osteoblasts, astrocytes, and dendritic cells (DCs).15 IL-18 is synthesized as 24 kD pro-IL-18, a biologically inactive precursor peptide, which is subsequently cleaved by caspase-1 into a biologically active IL-18 cytokine.15 The most salient biological property of IL-18 that distinguishes it from IL-1 is its ability to induce secretion of high levels of interferon (IFN)–γ in the presence of IL-12.16 Also, a substantial pool of intracellular IL-18 is present in the cytoplasm and is regulated mainly by caspase-1-mediated pro-IL-18 processing.15 IL-18 has a wide array of biological functions, many of which are accomplished synergistically with other cytokines, such as IFN-γ, as an early immune response to pathogenic invasions.16 Several investigations into biological role of IL-18 have shown that IL-18 is a potent inducer of (i) tumor necrosis factor alpha (TNFα), Fas-ligand, and IL-2 from T and NK cells; (ii) IL-1b, IL-8, and macrophage inflammatory protein (MIP)-1α from peripheral blood mononuclear cells; and iii) cyclooxygenase (COX2), and inducible nitric oxide synthase.17–19
The initial immune response to P. falciparum infection is mediated by IFN-γ that is induced by the combination of interleukin 12 (IL-12) and 18 (IL-18). This cytokine secretion is crucial for the control of parasitemia and resolution of malarial infection through TNFα induction and enhanced release of reactive nitrogen and oxygen radicals.20 Torre et al (2001) demonstrated that IL-18 may have a pro-inflammatory role in patients with uncomplicated P. falciparum infection.21 In another study, it was found that IL-18 and IL-12 levels were higher in children with mild malaria than in children with a severe form of the disease.22 Also, high quantities of both IL-18 and IFN-γ molecules was found to be associated with severe forms of malarial infections.20 A study in western Kenya investigated the relationship between IL-18 and clinical malaria phenotypes in children, and reported association between higher secretion of IL-18 with severe malarial anemia (SMA).23 Similarly, a study in adults demonstrated a close association between increased IL-18 levels and severe falciparum malaria.24 Taken together, these findings indicate that IL-18 is an important factor in the pathogenesis of P. falciparum induced malaria.
Several polymorphisms within the IL-18 gene have been reported to play an important role in the etiology of several diseases such as atherosclerosis,25 rheumatoid arthritis (RA), systemic lupus erythematosus (SLE),26,27 allergic asthma, and allergic dermatitis,28 mostly by altering the circulating levels of IL-18. However, very few studies have investigated IL-18 gene polymorphisms in relation to its role in malaria pathogenesis.
Previously, we have shown that genetic variations in the IL-22 gene could be correlated with susceptibility to malaria infection.29 In this study, we investigated whether the single nucleotide polymorphisms (SNPs) spanning the IL-18 gene were associated with the susceptibility to P. falciparum infection and related parasitemia. We also analyzed the influence of patient age and genetic polymorphisms in relation to extent of P. falciparum parasitemia.
Materials and Methods
Ethical Considerations
Ethical approval to conduct this study was granted by Ethical Review Board (ERB) of King Fahad Central Hospital (KFCH), Jazan (Registry no. 041). The study followed ethical guidelines set by the Helsinki Declaration of 1975. Prior to study subject enrolment, informed consent was obtained from each subject; legal guardians provided consent on behalf of children. All patient-related research data including human biological samples were de-identified and secured to preserve the privacy of study participants.
Study Design and Population
This case–control study was conducted and a total of 450 subjects were enrolled. Blood samples were collected from 250 patients diagnosed with P. falciparum malaria infection admitted at Malaria Center, Jazan region, situated in the Southwest part of the Saudi Arabia. In addition, 200 healthy controls were recruited as a control cohort. We used WHO manual (“plus system” scale) to identify the parasite density in a thick blood smear from infected individuals.30 The P. falciparum infected malaria subjects were classified into four categories based on the parasite density; 1) Group I: presence of 1–10 malarial parasites per 100 thick blood smear fields; 2) Group II: presence of 11–100 malarial parasites per 100 thick blood smear fields; 3) Group III: presence of 1–10 malarial parasites per single thick blood smear field; and 4) Group IV: presence of over 10 malarial parasites per single thick blood smear field. Patients were excluded from the study if they were co-infected with Dengue virus (DENV), Hepatitis C virus (HCV), Hepatitis B virus (HBV) or Human immunodeficiency virus (HIV).
Selection and Genotyping of IL-18 SNPs
The IL-18 gene is made up of 20.9 kb and consists of a 5ʹ-untranslated region (UTR) promoter, 6 exons and a 3ʹUTR region. The SNPs were genotyped using the polymerase chain reaction (PCR)-based direct DNA sequencing and SNP-specific TaqMan assays. Ten tag SNPs were identified based on the following criteria: minor allele frequency (MAF) more than 5% and linkage disequilibrium (LD) threshold of r2 ≥ 0.8. The 10 identified tag SNPs were: rs11214101, rs544354, rs360729, rs5744260, rs549908, rs360714, rs7106524, rs5744231, rs360718, and rs1293344. In addition, six SNPs, rs187238, rs1946518 and rs1946519 from the promoter region and rs360727, rs5744292 and rs360728 from and 3ʹUTR of the IL-18 gene were selected. The details of the SNPs are summarized in Figure 1 and Supplementary Table 1.
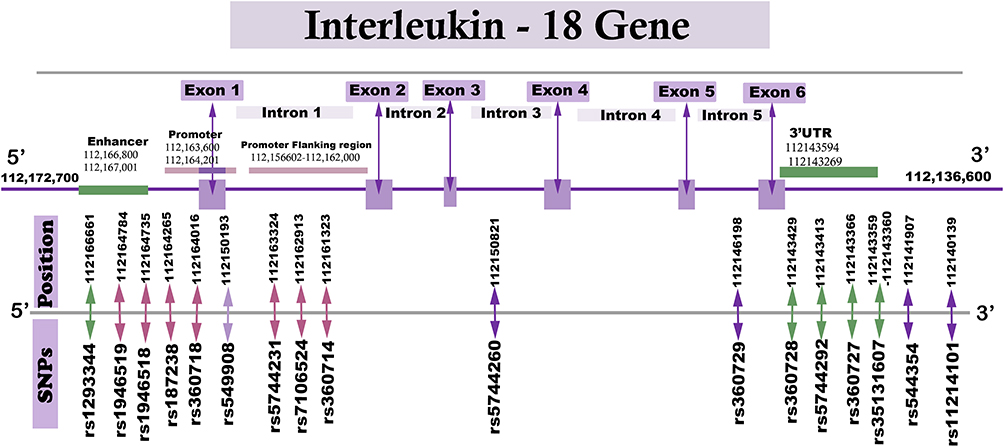 |
Figure 1 Schematic drawing illustrating the positions of IL-18 SNPs investigated in this study. |
DNA Extraction and Polymerase Chain Reaction (PCR)
Using the manufacturer’s guide, DNA was extracted via the DNeasy® Blood & Tissue Kit (Qiagen, Hilden, Germany). The PCR was carried out using GoTaq® Master Mixes (Promega Corporation, Madison, WI, USA). Primers were designed through the Primer3 Input version 0.4.0 available at (http://bioinfo.ut.ee/primer3-0.4.0/) and they are shown in Supplementary Table 2. All primers were M13-tailed to improve sequencing of the PCR products. The resultant PCR products were run on 2% agarose gel electrophoresis to ensure PCR products’ quality. The amplified PCR products underwent direct sequencing by a BigDye™ Terminator v3.1 Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA) as per the recommended protocol. Afterward, each of the PCR products was subjected to both reverse and forward sequencing. The final sequences were evaluated and modified using the SeqMan Pro 15 Lasergene (DNASTAR, Inc., Madison, WI, USA).
Transfection and Luciferase Reporter Assay
Several DNA fragments of the IL-18 promoter and 3ʹ UTR region were cloned and transfected in HuH7 (obtained from the Japanese Collection of Research Bioresources Cell Bank, catalogue number: JCRB0403) cells and the variations in the luciferase gene activity were monitored. The chromosomal positions of the fragments and the primers used for amplification are shown in Supplementary Tables 3 and 4 for the promoter and 3ʹ UTR regions, respectively. All of the fragments were cloned in pGL3 Basic Vector (accession number: U47297) (Promega, Madison, WI, USA). HotStar DNA Polymerase (Qiagen, Hilden, Germany) was used for PCR amplification using the guidelines of the manufacturer. For colony selection on ampicillin plates, Escherichia coli DH5α competent cells (catalogue number: 18265017) (ThermoFisher Scientific, Sunnyvale, CA, USA) were utilized. Replicates were selected and DNA was extracted through QIAamp DNA Mini Kit (Qiagen, Valencia, CA). Extracted DNA fragments were then transfected into HuH7 cells, which were cultivated and evaluated for luciferase activity after 48 hours.
Statistical Analyses
We used HaploView software version 4.0 (Broad Institute of MIT and Harvard, Cambridge, MA, USA) to analyze SNPs and measure linkage disequilibrium (LD), haplotype frequencies and MAFs. To assess the genotypic association for selected genetic variants, we used the De Finetti program available at (http://ihg.gsf.de/cgi-bin/hw/hwa1.pl). In addition, we conducted analyses to identify for the allelic, recessive and dominant modes of inheritance for each genetic variant. The study findings were presented as odds ratios (OR) with their 95% confidence intervals (CI). The Hardy–Weinberg equilibrium (HWE) was performed to test for deviation; P value <0.05 was considered as a statistically significant deviation from HWE. The P value <0.05, obtained in a two-tailed test, was set as a threshold for statistically significant association.
Results
Genotype and Allele Frequency Analysis Between P. falciparum-Infected Patients and Healthy Control Group
Genotypic distributions of SNPs in the healthy control group and the P. falciparum-infected patients are shown in Table 1. We found that SNPs rs5744292 (OR = 70.446; 95% CI= 4.318–1149.323; p<0.0001) and rs544354 (OR = 1.498; 95% CI= 1.088–2.063; p = 0.013) were of significantly higher occurrence in P. falciparum-infected patients when compared to healthy control group. The rs5744292-G allele (8%) and rs544354-A allele (26%) showed increased frequency among infected patients in comparison with the control group. We also observed that the individuals carrying either the homozygous genotype rs5744292-GG or the heterozygous AG were associated with increased risk of malarial infection with an OR of 13.828 and 48.857, respectively, when compared to homozygous AA carriers. Under a dominant model of inheritance, the rs5744292-G allele was found to be associated with an increased risk of infection (OR = 61.763). Similarly, we found that individuals carrying the homozygous AA genotype with SNP rs544354 had an increased risk of parasitemia (OR = 5.253) when compared to homozygous GG carrying individuals. In addition, the rs544354-A allele was significantly associated with risk of P. falciparum infection in both dominant (OR = 1.435) and recessive (OR = 0.209) models. The homozygous SNP rs360714 GG genotype was found to be associated with an increased risk of malaria infection (OR = 0.147) compared to the homozygous AA genotype, whereas the heterozygous rs7106524 AG genotype was associated with an increased risk of the infection (OR = 1.652) compared to the homozygous GG genotype.
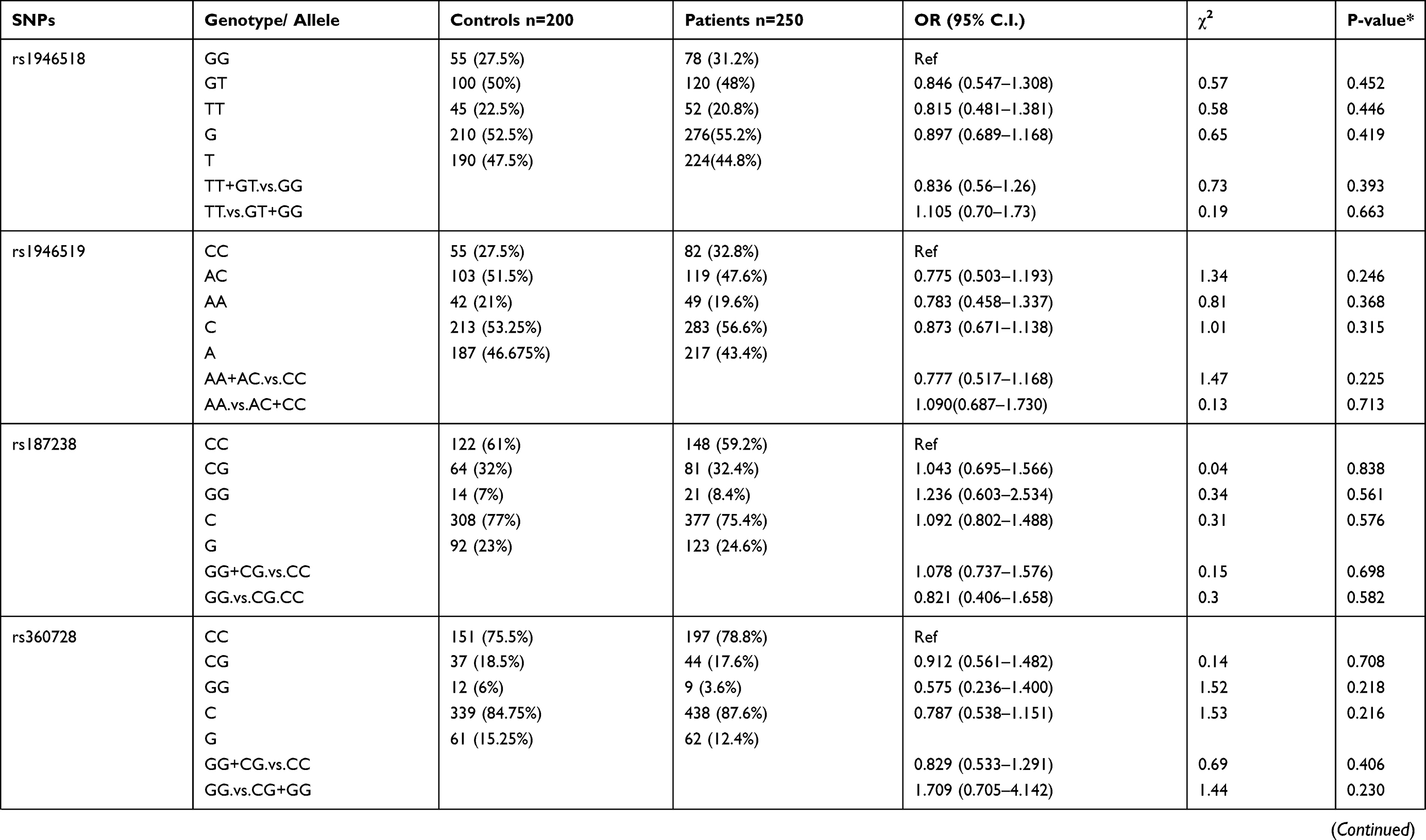 |
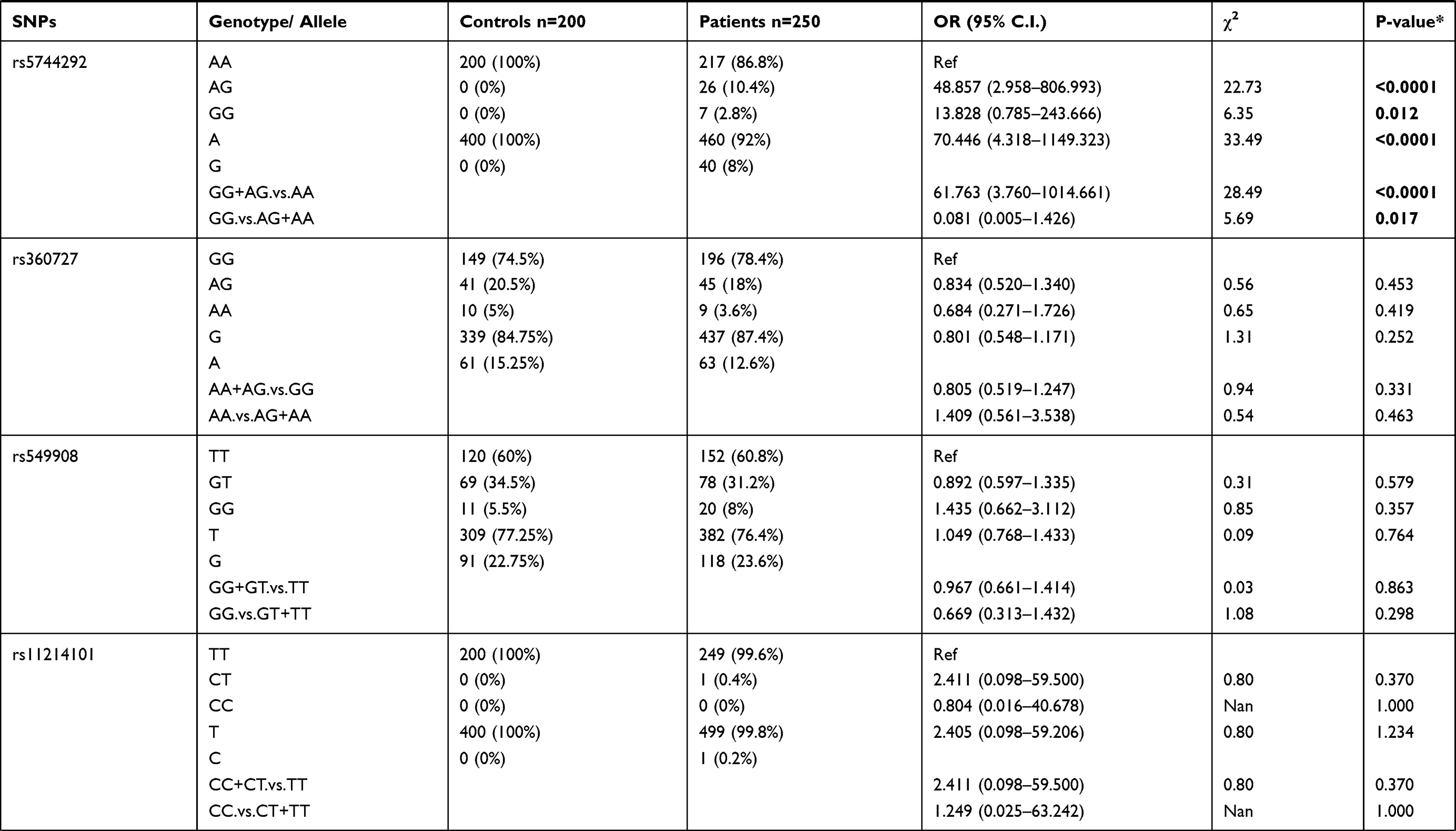 |
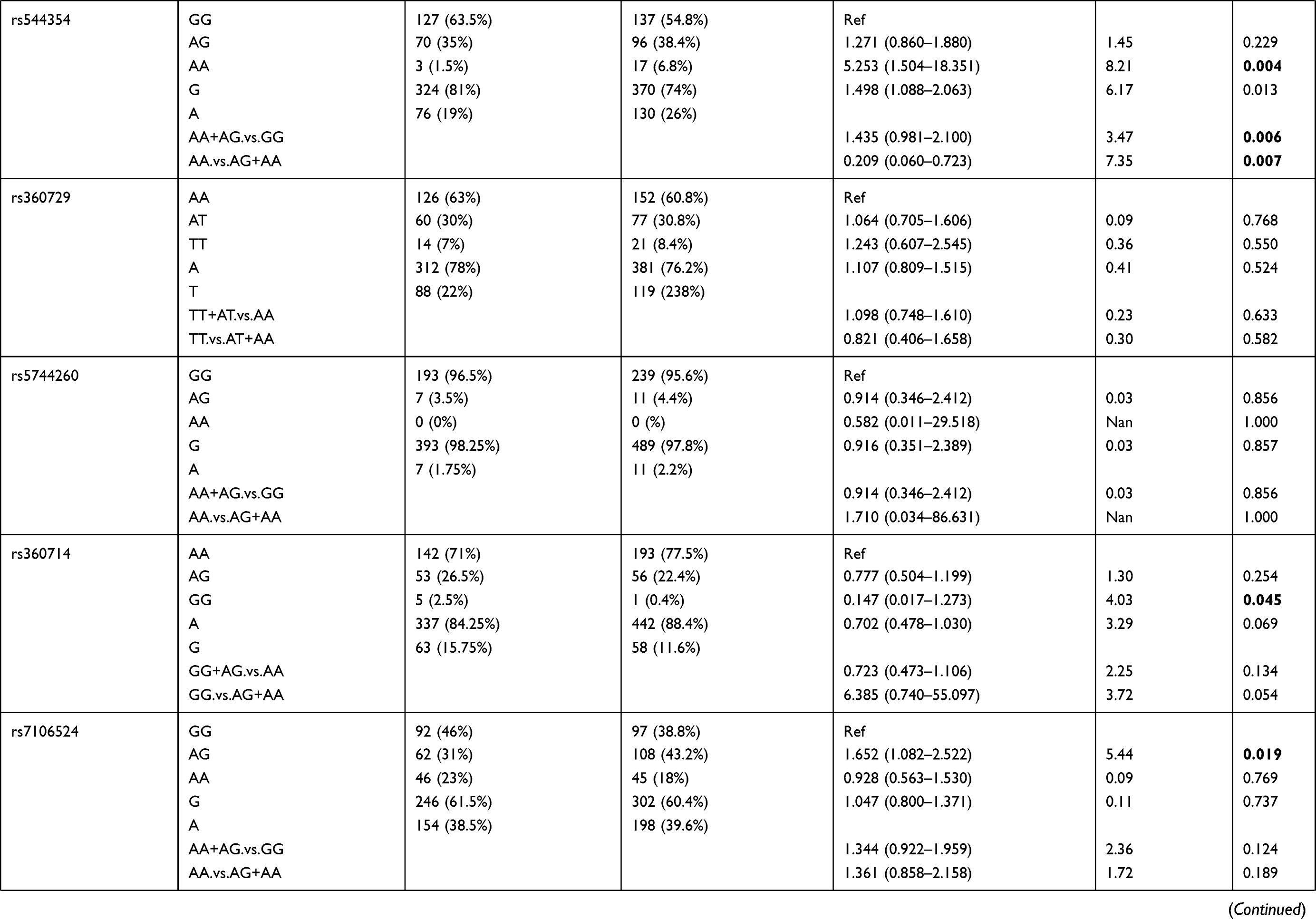 |
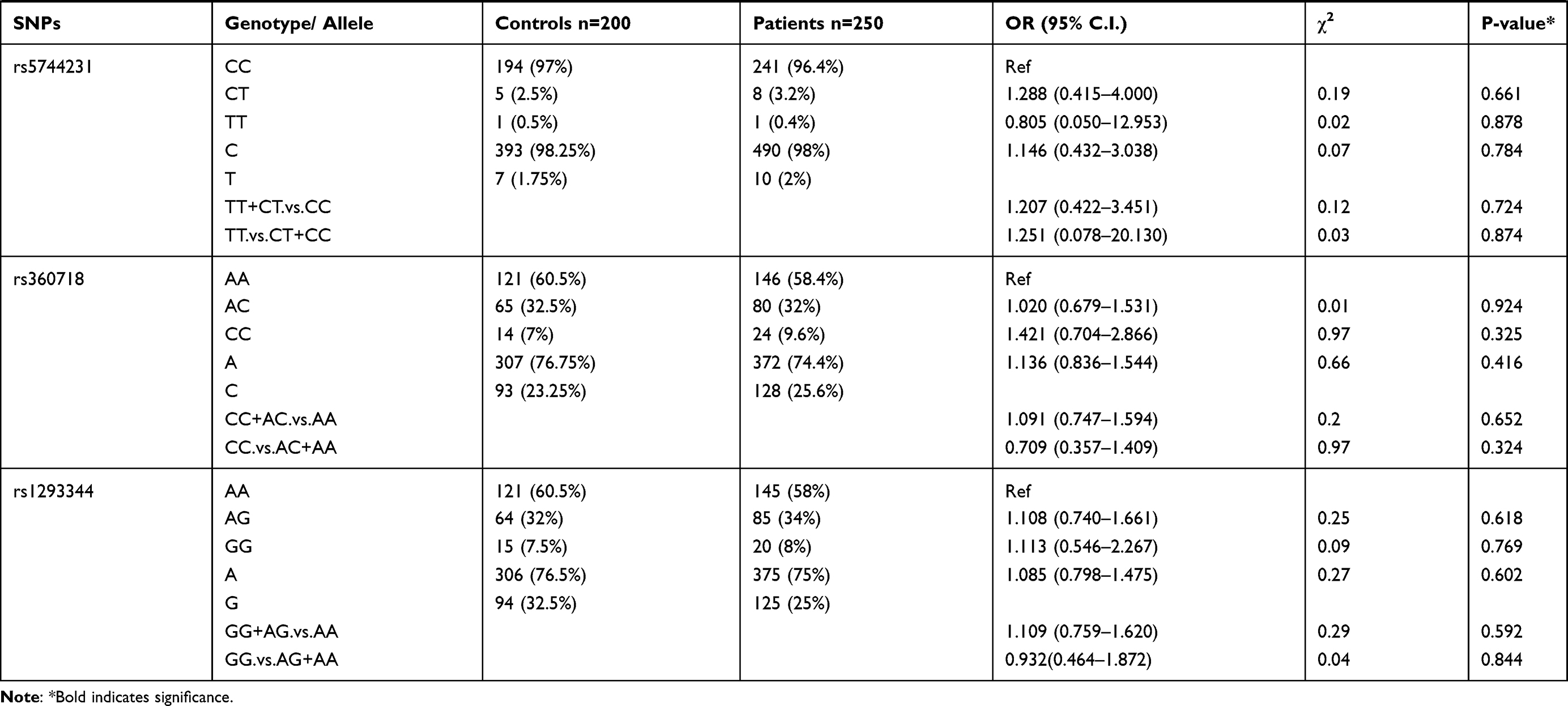 |
Table 1 Genotype Distribution and Allele Frequency of IL-18 Gene SNPs Among P. falciparum-Infected Patients and Healthy Control Subjects |
Genotype and Allele Frequency Analysis Based on Parasite Density
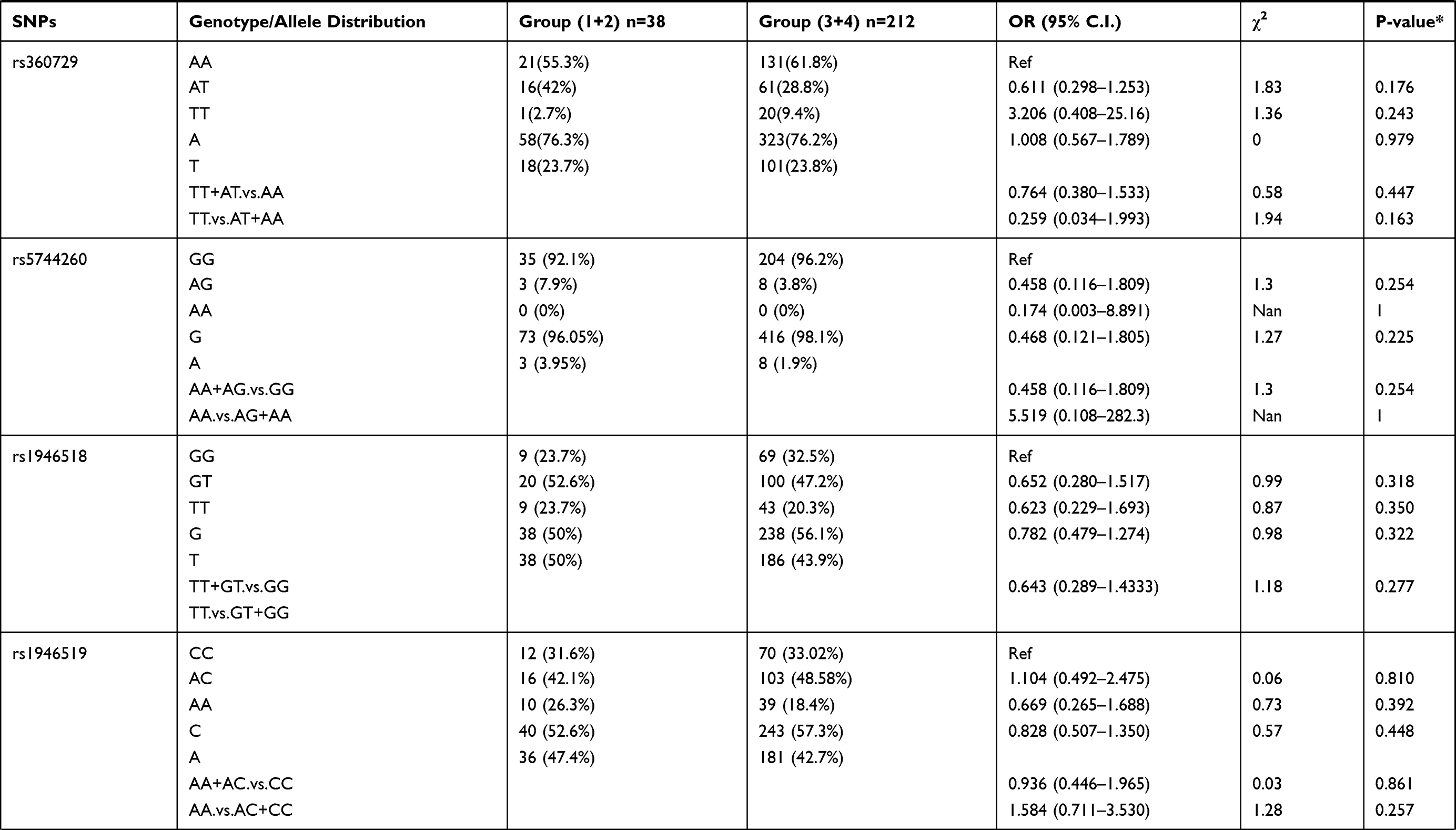
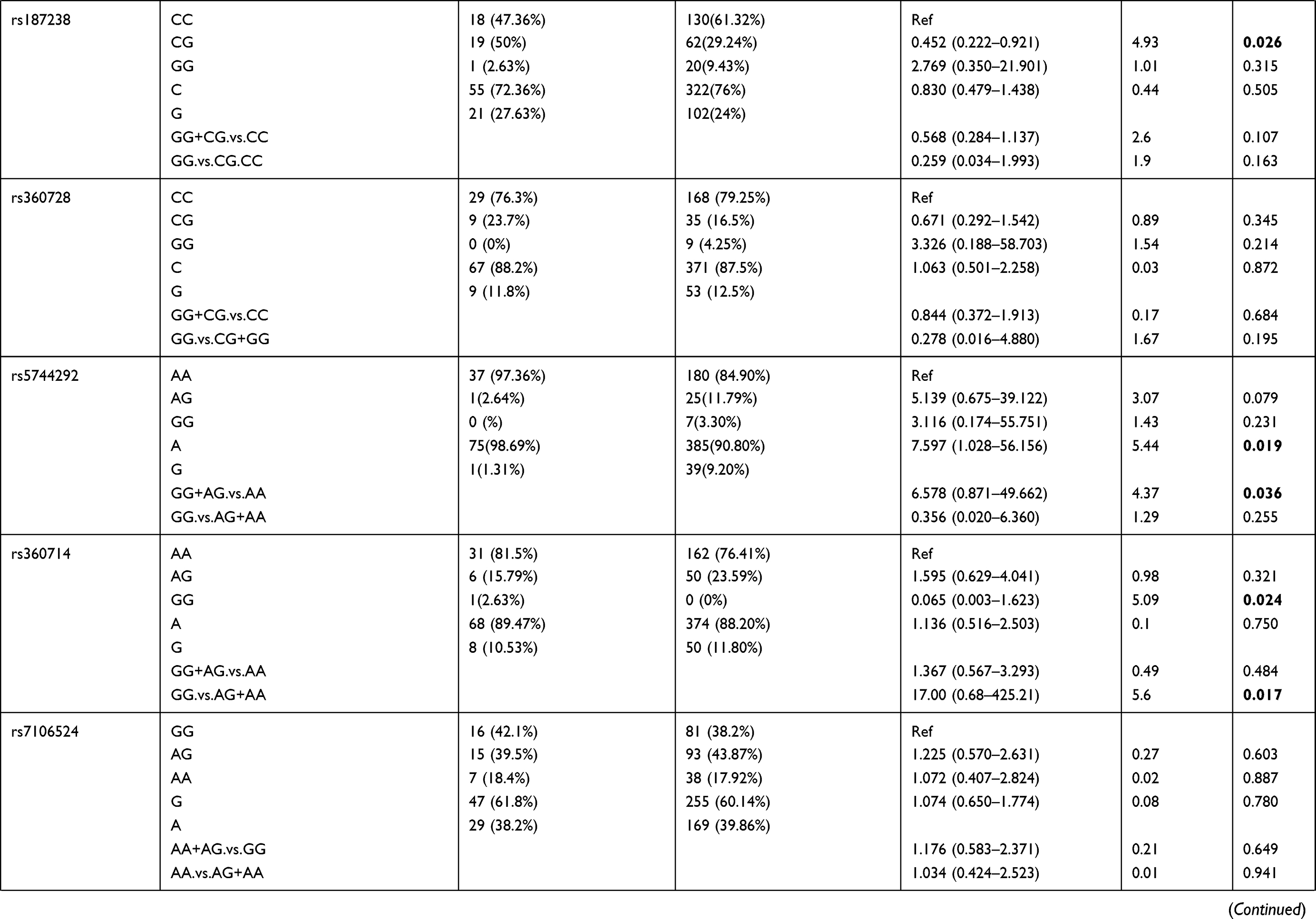
Patients were divided into 4 groups based on increasing parasite density: Group 1 (lowest density; 14 patients), Group 2 (24 patients), Group 3 (52 patients), and Group 4 (highest density; 160 patients). The 16 selected polymorphisms were further investigated to determine their association with varying levels of parasitemia in the P. falciparum-infected individuals. Allelic and genotypic frequency comparisons were done between the patients in group 1+2 with the patients in group 3+4 (Table 2). We found that SNP rs5744292 (OR = 7.597; 95% CI = 1.028–56.156; p = 0.01967) was associated with increased parasite density in P. falciparum-infected patients. Furthermore, the rs5744292-G allele was significantly associated with higher parasitemia levels under the dominant model with an OR of 6.578. We also found heterozygous genotype CG for SNP rs187238 (OR = 0.452) and AG for SNP rs1293344 (OR = 0.492), to be associated with variations in malarial parasitemia, when compared with homozygous genotypes CC and AA, respectively. Under a recessive model of inheritance, rs360714-G allele showed a significant association with increased levels of parasitic density (OR = 17).
 |
 |
 |
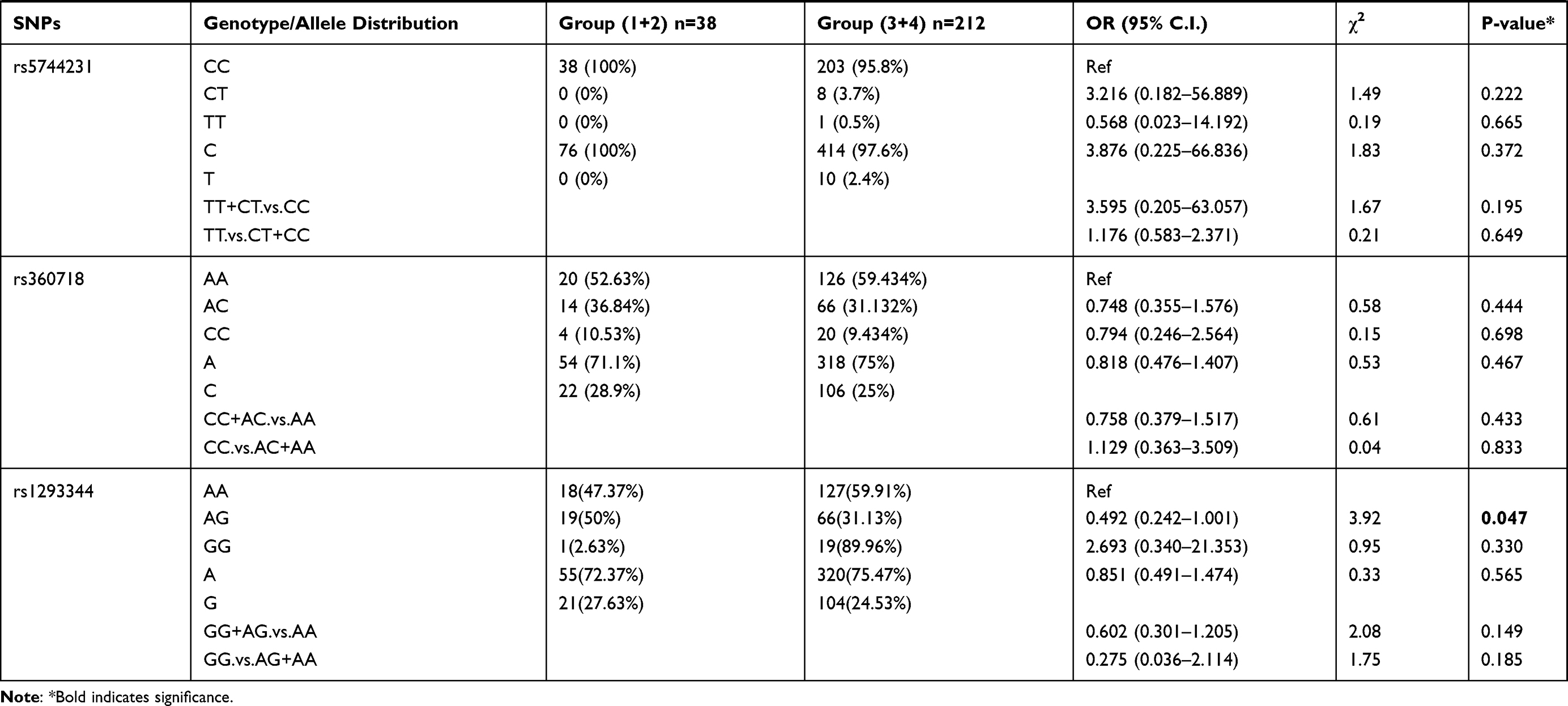 |
Table 2 Genotype Distribution and Allele Frequency of IL-18 Gene SNPs When Group 1+2 of Parasite Density Was Compared with Group 3+ 4 |
We also compared allele frequency for these SNPs in group 1+2+3 against group 4. Under a recessive mode of inheritance, we found that rs549908 G allele (OR = 0.290) and rs360729 T allele (OR = 0.272) were both significantly associated with increased level of parasitemia. No other association was observed between these two groups (Table 3).
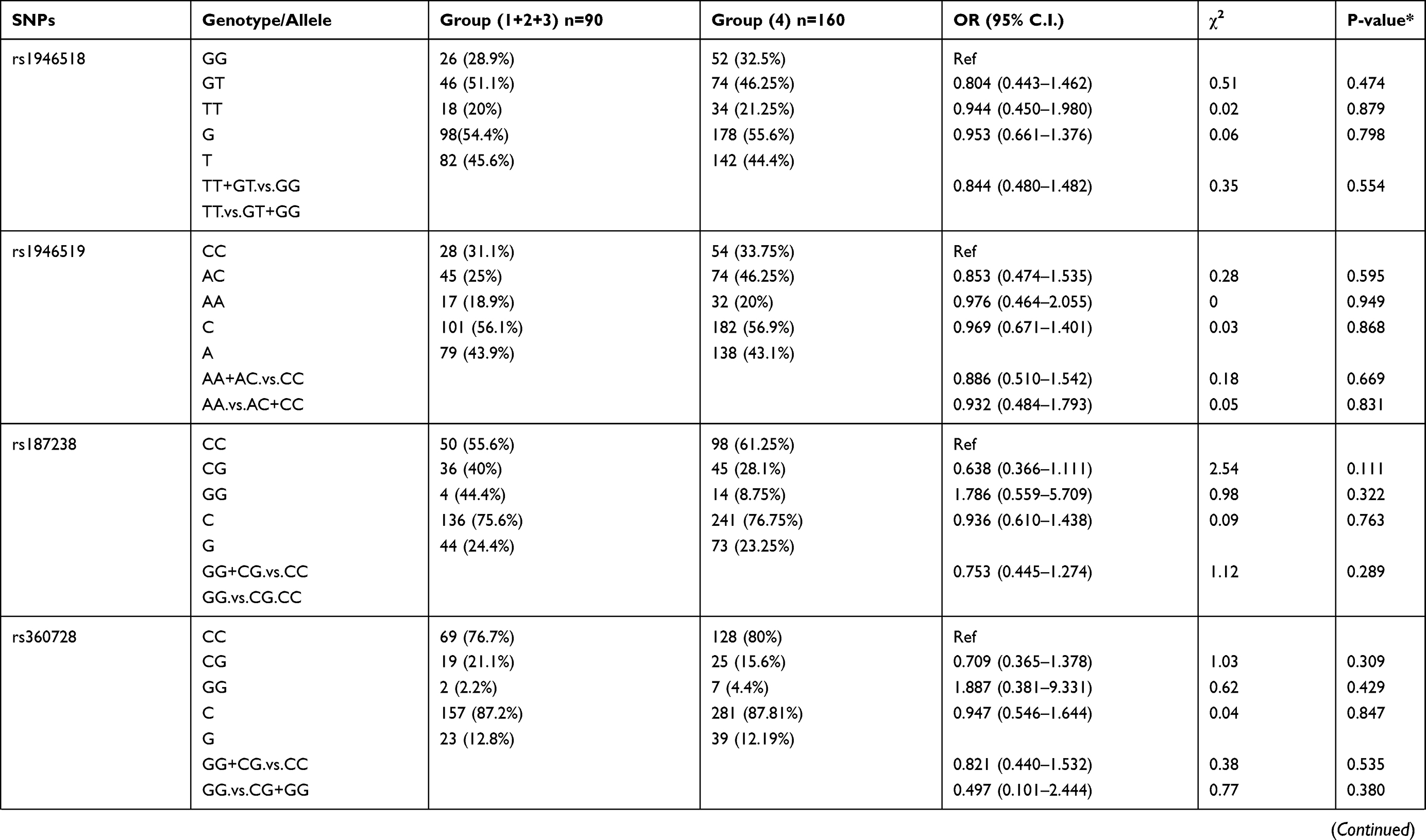 |
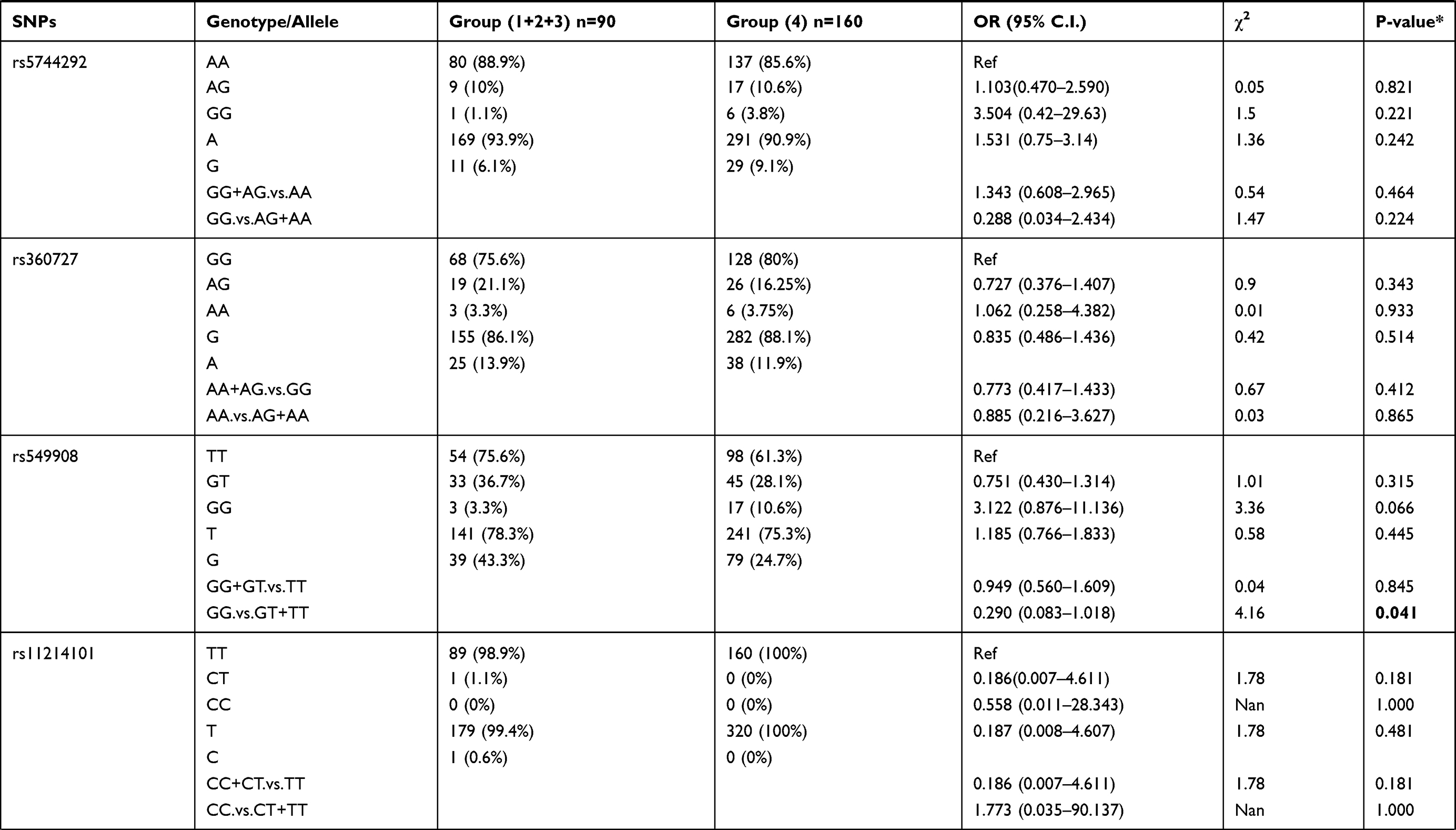 |
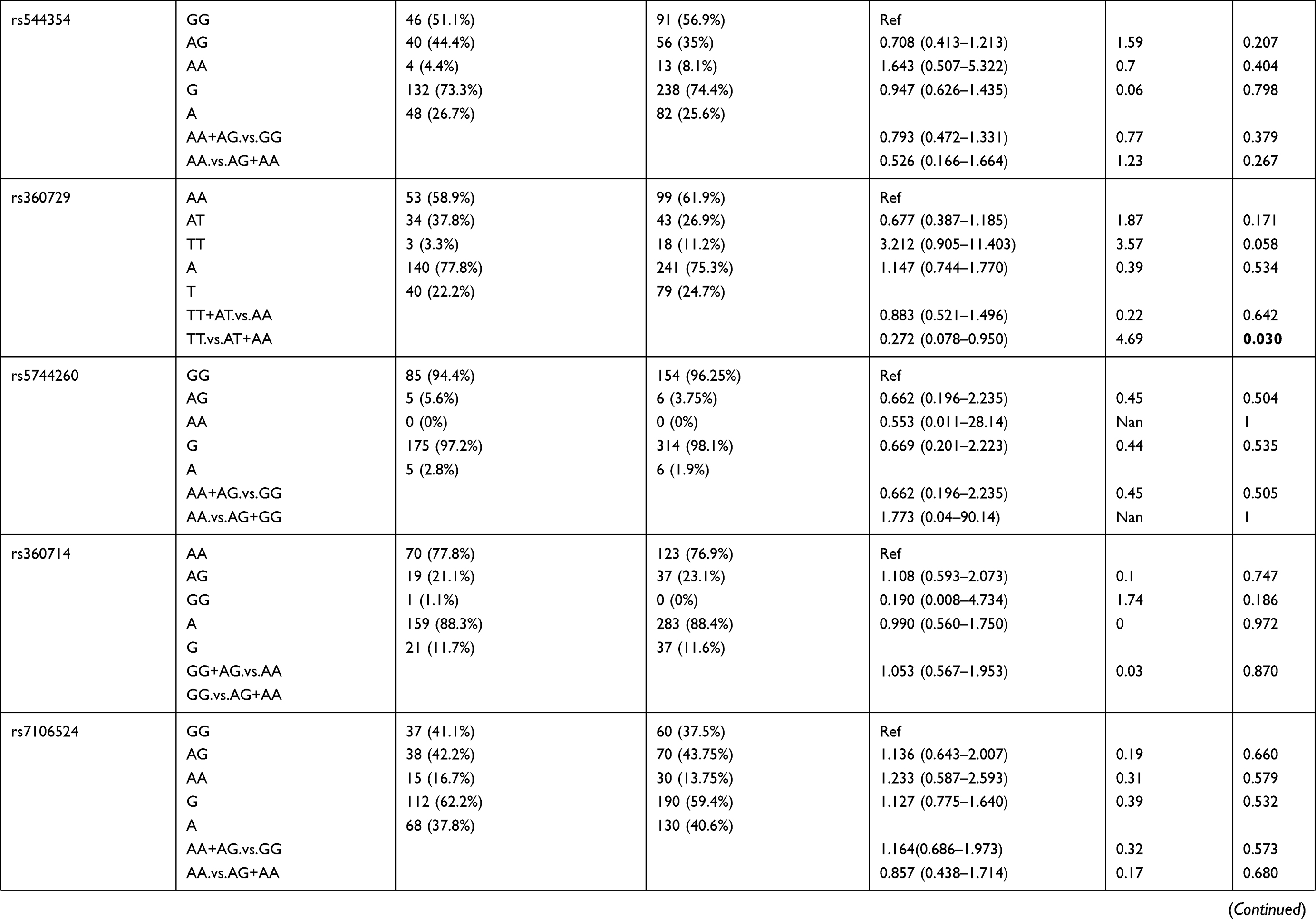 |
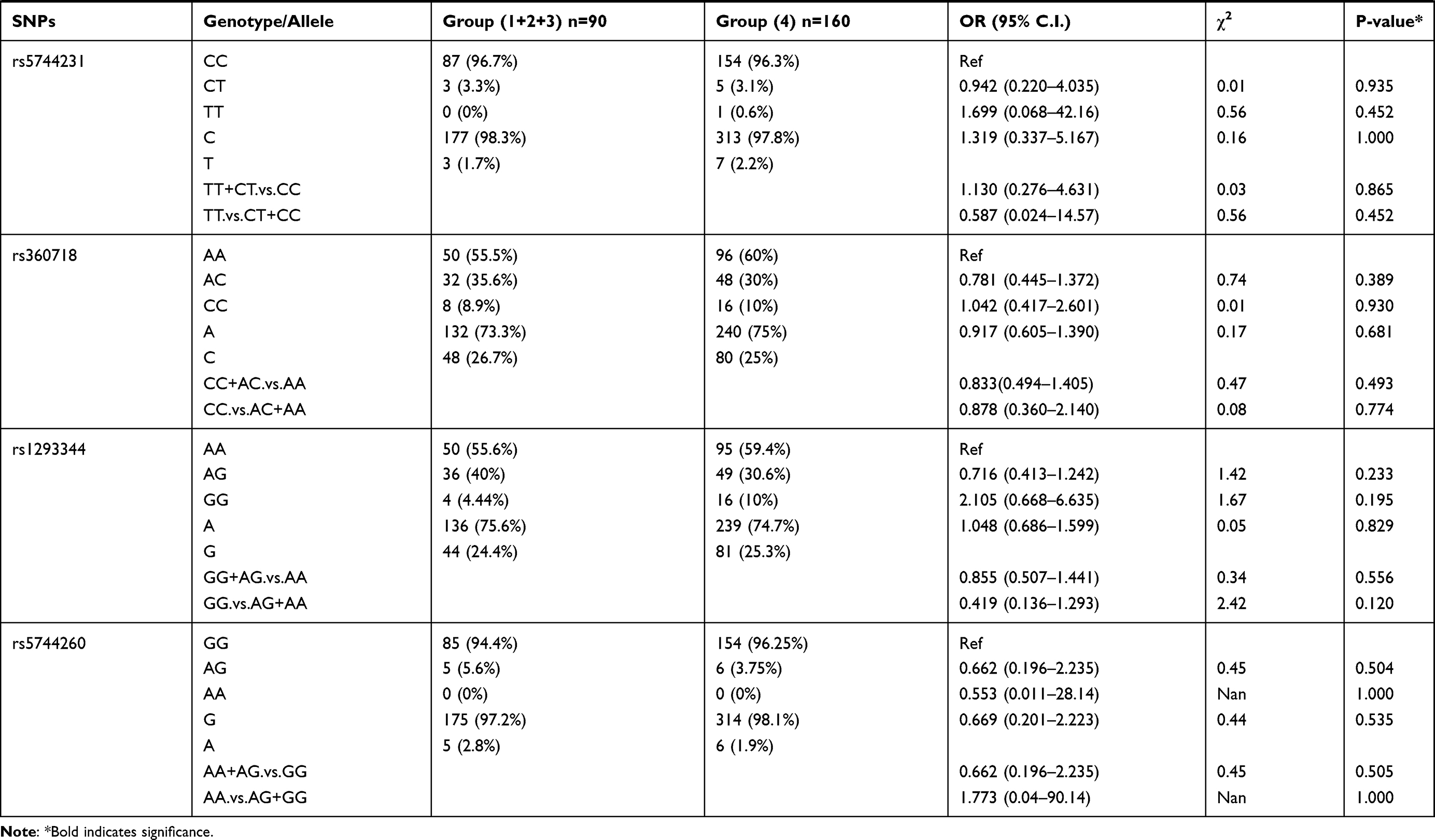 |
Table 3 Genotype Distribution and Allele Frequency of IL-18 Gene SNPs When Group 1+2+3 of Parasite Density Was Compared with Group 4 |
Haplotype Association Analysis
Haplotype analysis was done between the healthy control group and P. falciparum-infected patients and the result is summarized in Table 4. Out of the three haplotypes identified, the occurrence frequency of two haplotypes, GT and AT were significant between these two groups. GT (rs544354 and rs549908) was found to be more frequent within the healthy controls (f = 0.586), compared to P. falciparum-infected patients (f = 0.516), (χ2 = 4.352; p = 0.037). However, AT (rs544354 and rs549908) was more prevalent among the P. falciparum-infected patients (f = 0.248), compared to the healthy controls (f = 0.187), (χ2 = 4.834; p = 0.028).
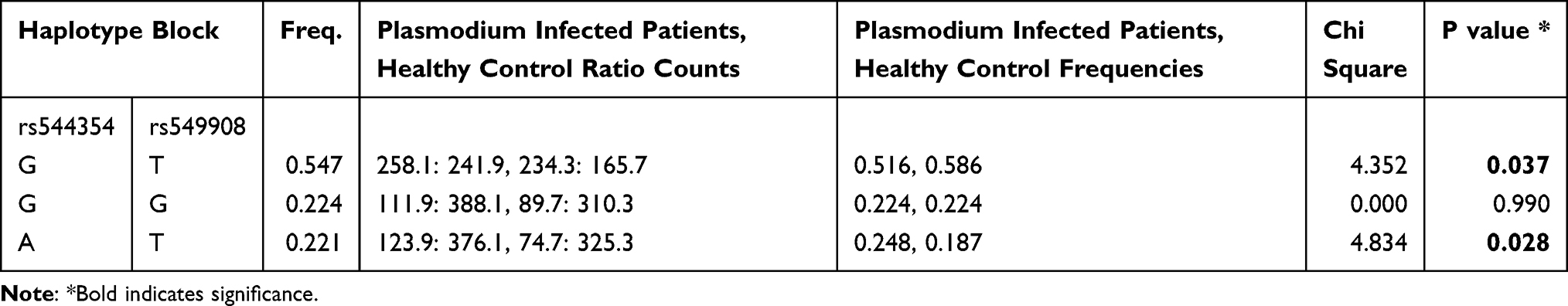 |
Table 4 Haplotype Analysis of IL-18 Gene Polymorphisms Among P. falciparum-Infected Patients versus Healthy Control Subjects |
Similar haplotype analysis was done between P. falciparum-infected patients in group 2+3+4 and group 1 for parasitic density. We observed that the haplotype CAT (rs360728, rs360729 and rs549908) had significantly higher frequency (f=0.821) within the group 1 patients, compared to combined group 2+3+4 patients (f = 0.637), (χ2 = 3.926; p = 0.048) (Table 5).
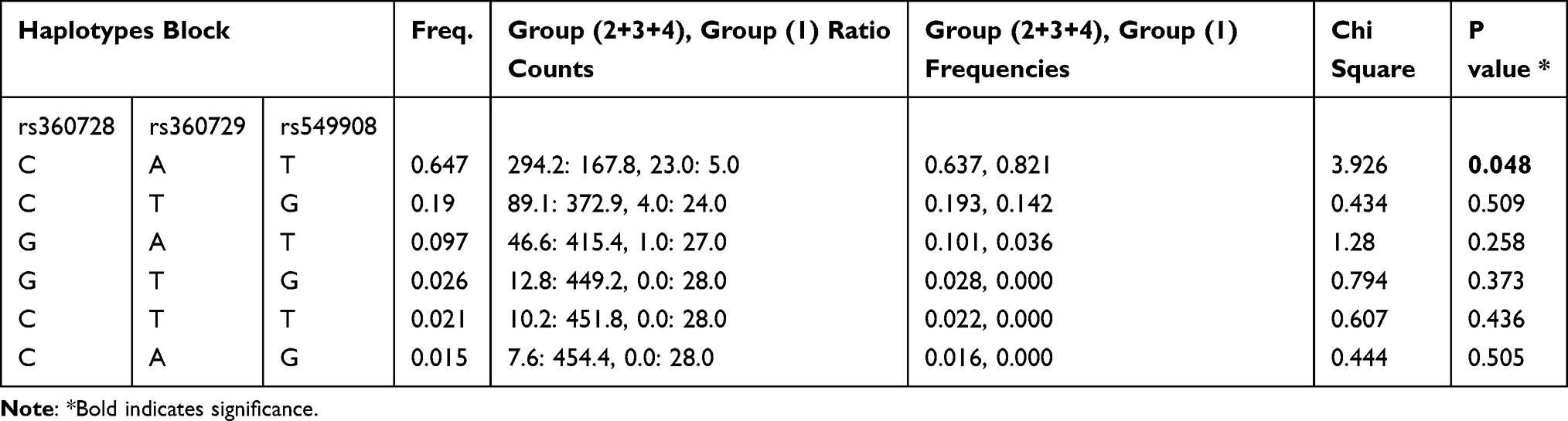 |
Table 5 Haplotype Analysis of IL-18 Gene Polymorphisms in Group 1 versus Group 2+3+4 of P. falciparum-Infected Patients |
Genotype and Allele Frequency Analysis Based on Age
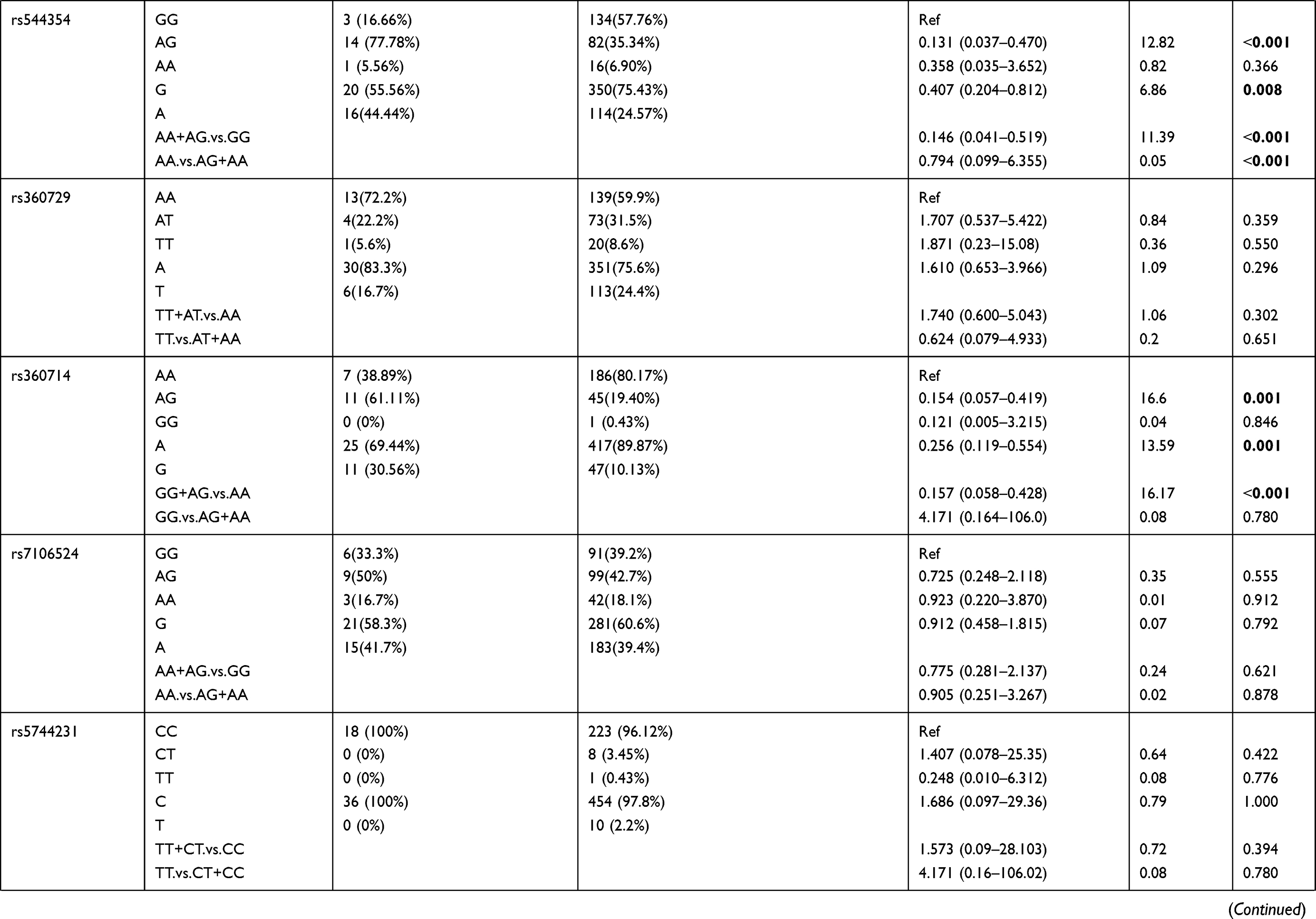
We also examined the relation of the 16 selected SNPs in the IL-18 gene with the age of the P. falciparum-infected patients. For this analysis, the 250 patients with parasitemia were divided into different age groups. The genotypic distribution of patients in 1–9 years age group compared to the patient group ≥10 years of age is represented in Table 6. We observed rs544354 (OR = 0.407; 95% CI = 0.204–0.812; p = 0.008) and rs360714 (OR = 0.256; 95% CI = 0.119–0.554; p=0.001) to be significantly associated with age in P. falciparum-infected patients. Heterozygous rs544354-AG genotype was present in significantly higher frequency (f = 0.77) in age group 1–9 years, compared to patients in ≥10 years age group (f = 0.35). It was observed that both under dominant (OR = 0.146; 95% CI = 0.041–0.519; p <0.001) and recessive (OR = 0.794; 95% CI = 0.099–6.355; p <0.001) models of inheritance, rs544354-A allele occurrence was associated with the age of the infected patients. Similarly, for SNP rs360714, heterozygous-AG genotype showed a higher incidence (f = 0.61) in P. falciparum-infected patients in the 1–9 years age group compared to patients above 10 years of age (f = 0.19). In the dominant model, rs360714-G allele was also significantly associated with patient age (OR = 0.157; 95% CI = 0.058–0.428; p <0.001).
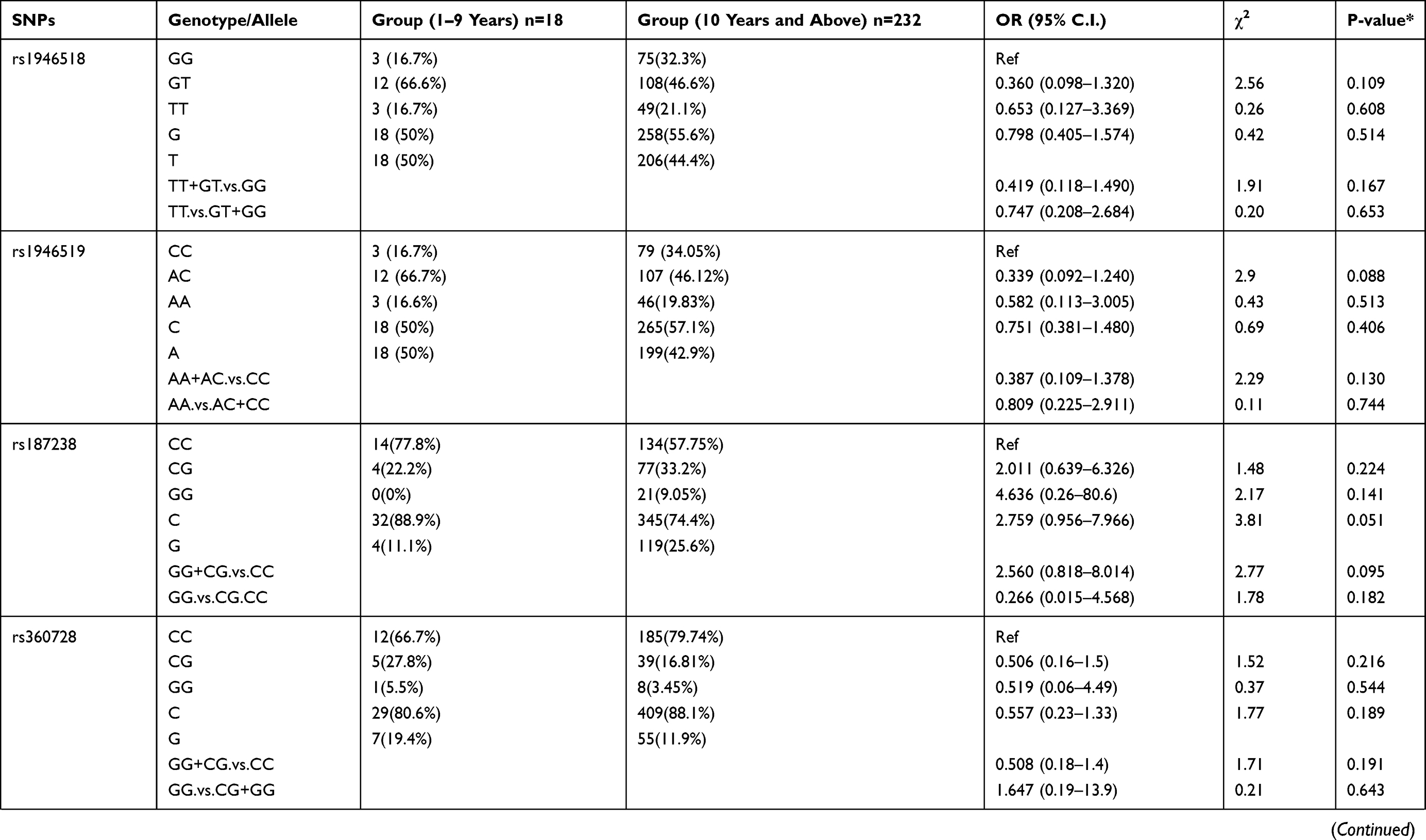 |
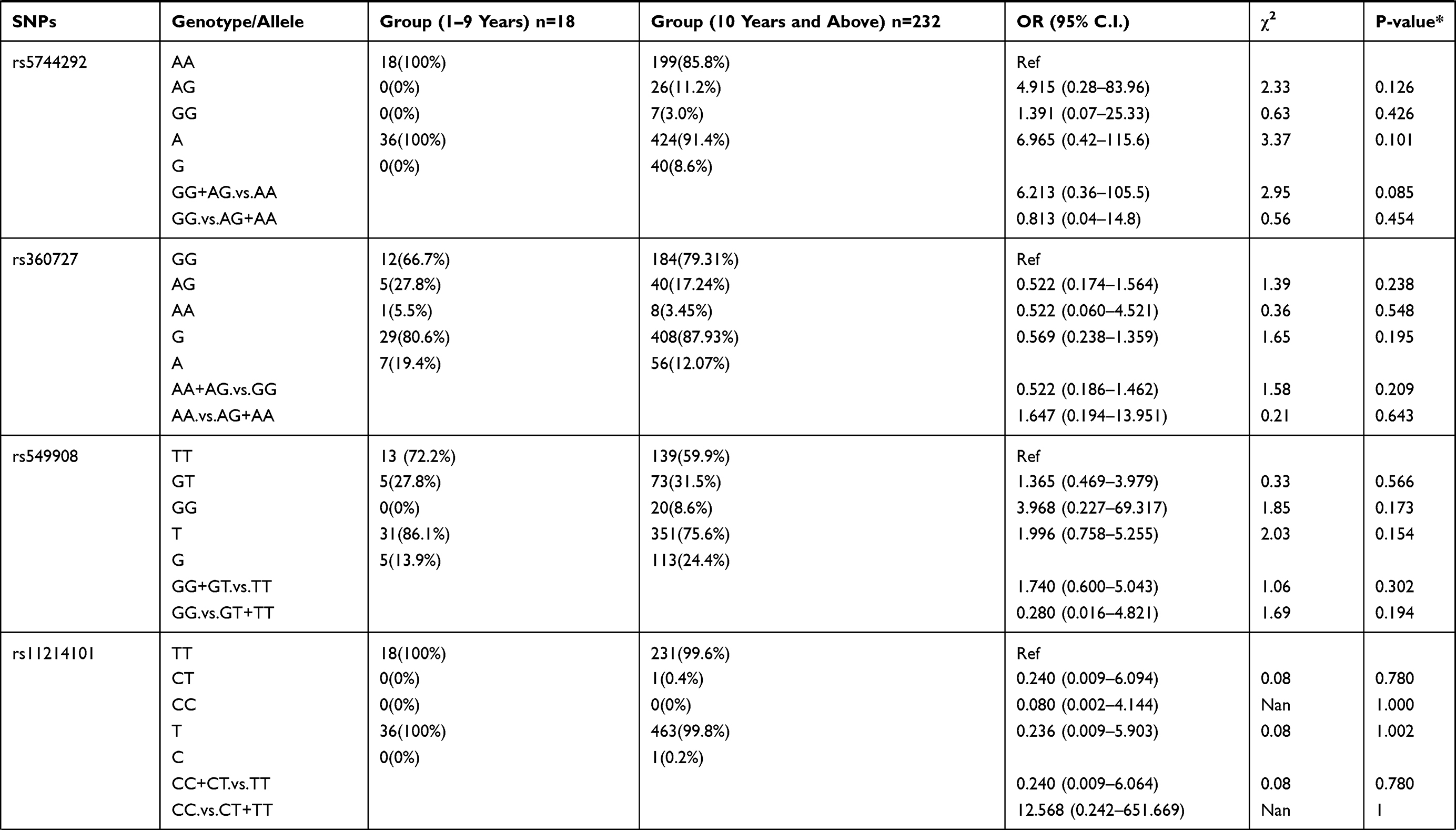 |
 |
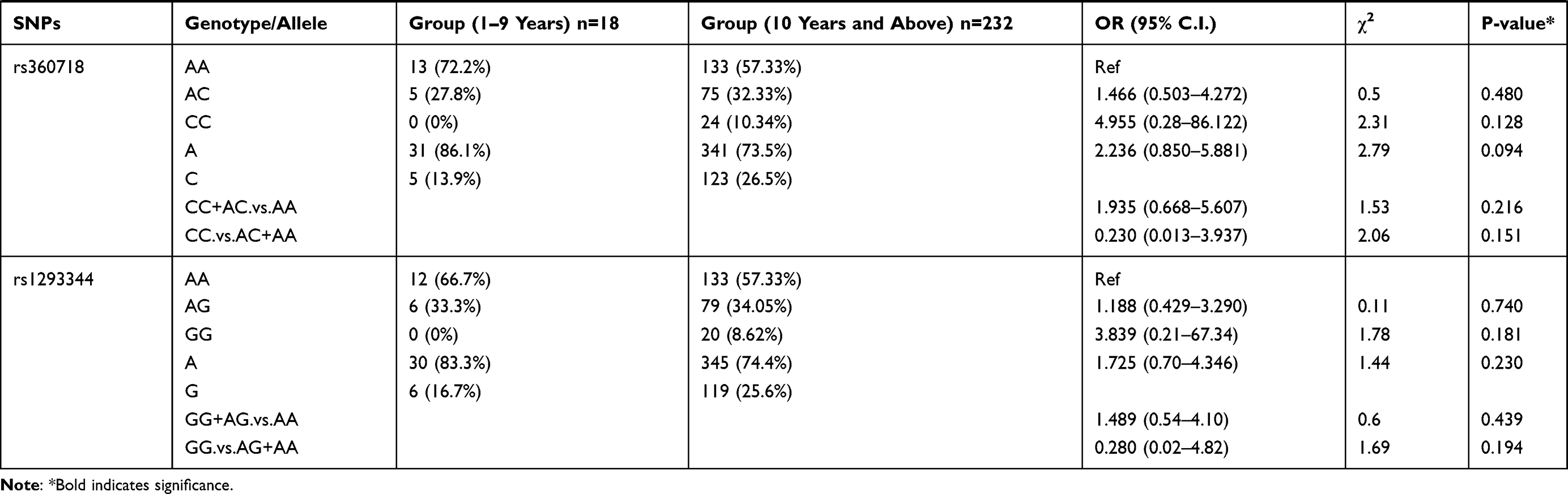 |
Table 6 Genotype Distribution and Allele Frequency of IL-18 Gene SNPs When Age Group 1–9 Years Was Compared with Group 10 Years and Above |
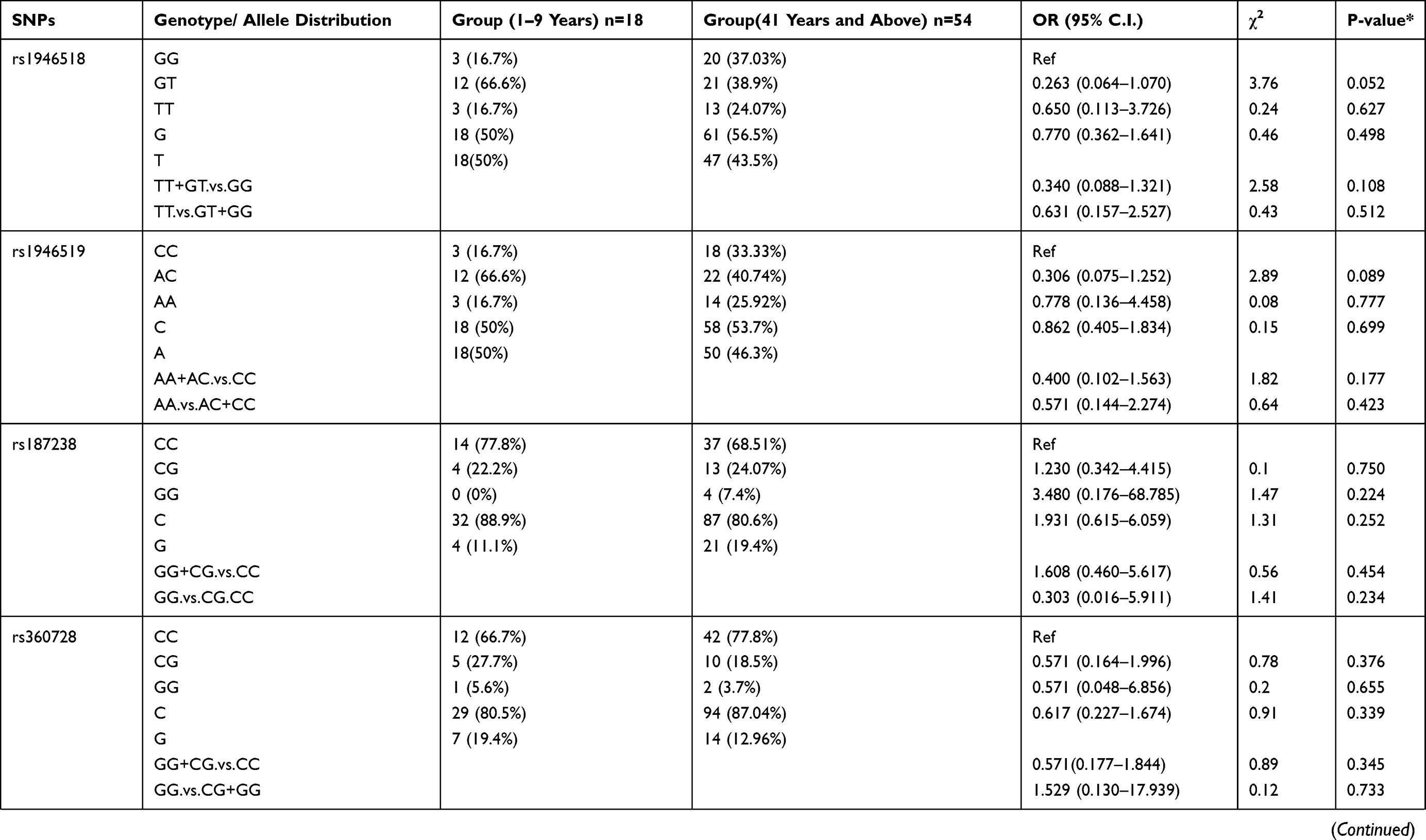
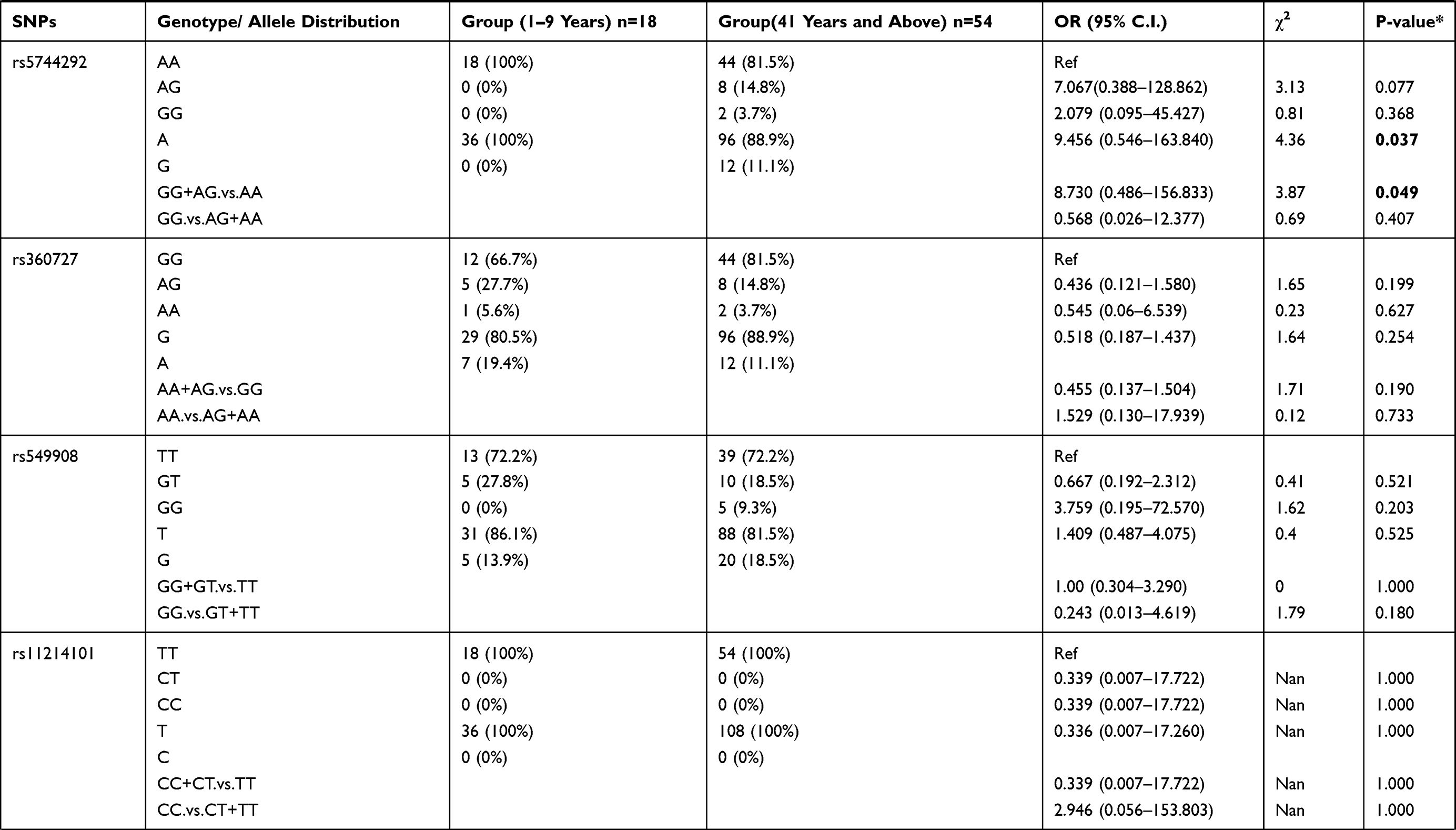
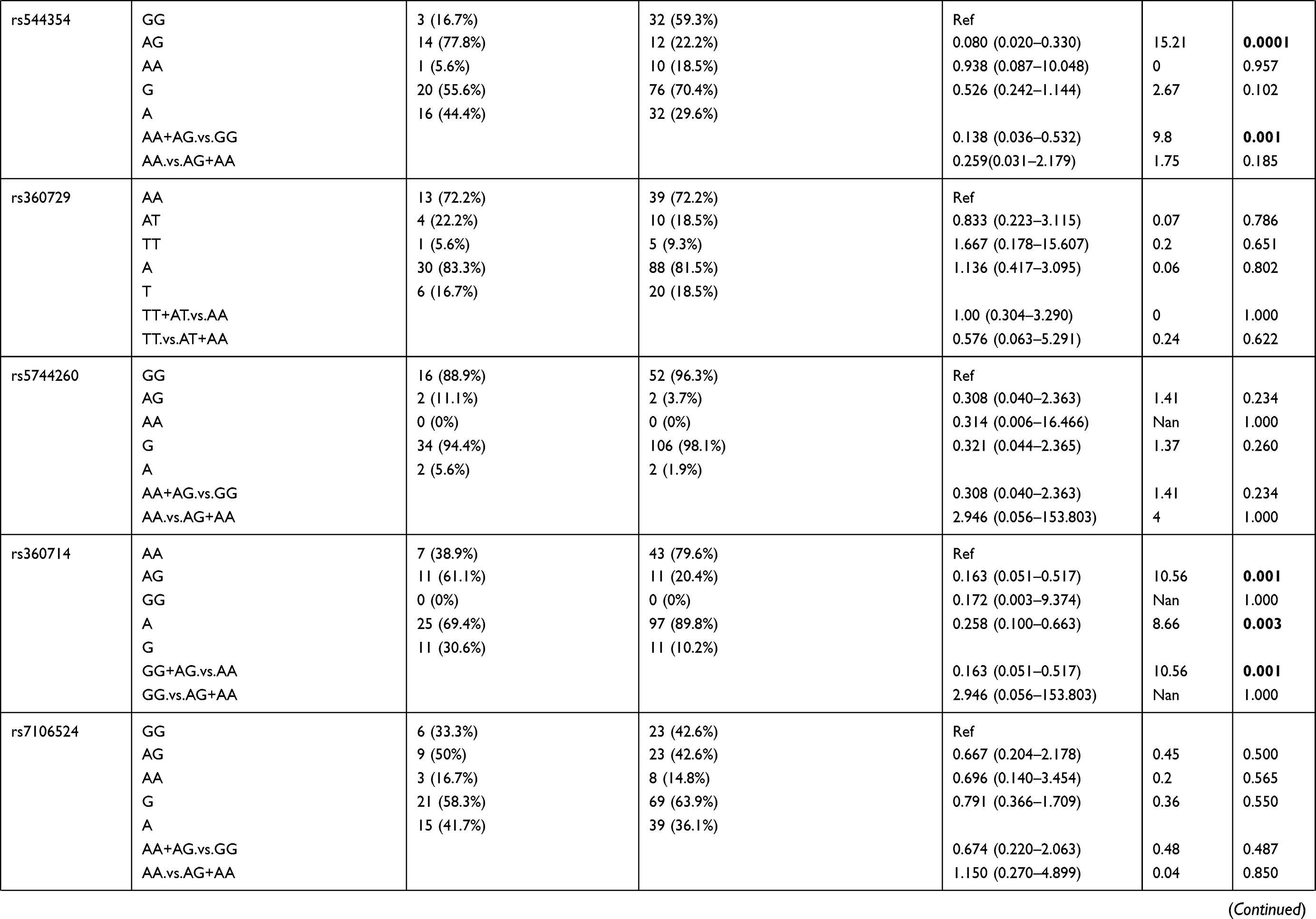
Further analysis was performed between the patient group 1–9 years age group compared to the patient group ≥41 years of age (Table 7). The results showed that rs5744292 A allele (OR = 9.456; 95% CI = 0.546–163.840; p = 0.037), rs544354 AG genotype (OR = 0.080; 95% CI = 0.020–0.330; p=0.0001) and rs360714 AG genotype (OR = 0.163; 95% CI = 0.051–0.517; p=0.001) are significantly associated with age in P. falciparum-infected patients. Heterozygous rs544354-AG genotype was present in significantly higher frequency (f = 0.77) in age group 1–9 years, compared to patients in ≥41 years age group (f = 0.22). Also, heterozygous rs360714-AG was present in a significantly higher frequency (f = 0.61) in age group 1–9 years, compared to patients in ≥41 years age group (f = 0.20)
 |
 |
 |
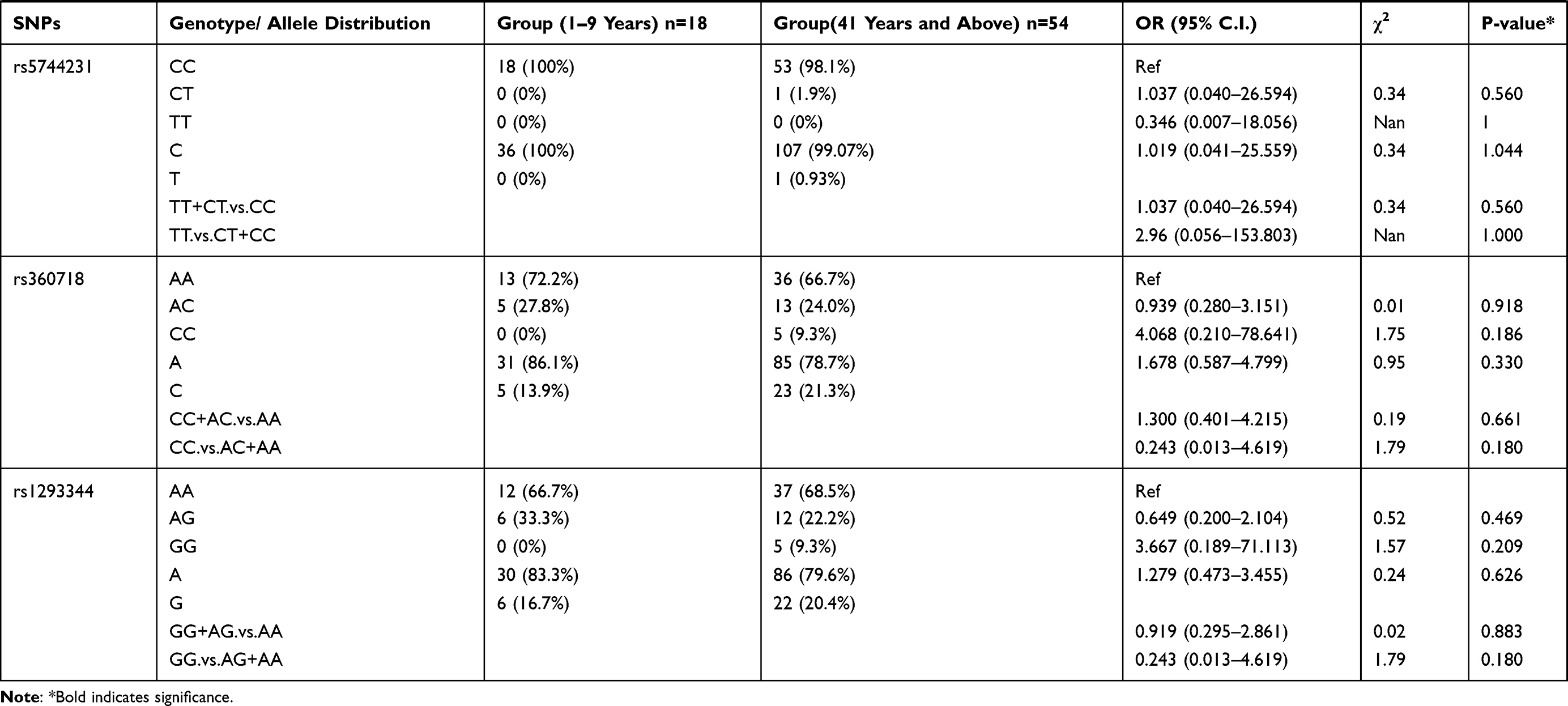 |
Table 7 Genotype Distribution and Allele Frequency of IL-18 Gene SNPs When Age Group 1–9 Years Was Compared with Group 41 Years and Above |
Haplotype Analysis Between Different Age Groups
We performed a haplotype analysis between the different age groups within the P. falciparum-infected patients. Haplotype analysis between the different age groups within the P. falciparum infected patients identified two haplotypes GG (rs544354 and rs360727) with a significantly higher frequency among ≥10 years age group (f = 0.674) compared to 1–9 years age group (f = 0.491), (χ2 = 4.93; P = 0.026), and haplotypes AA (rs544354 and rs360727) with a significantly higher frequency among 1–9 years age group (f = 0.130) compared to in patients above 10 years of age (f = 0.045), (χ2 = 4.984; P = 0.026) (Table 8).
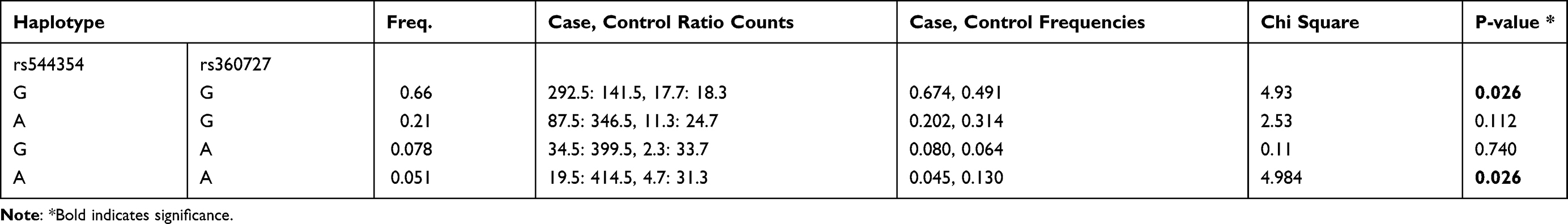 |
Table 8 Haplotype Analysis for IL-18 Gene Polymorphisms Between Group 1 (1–9 Years) Was Compared to 10 Years and Above |
Haplotype analysis was also done for the IL-18 SNPs between the patients within the age groups 1–9 years and ≥41 years. Two significant haplotypes were identified for these age groups (Table 9). This included haplotype AAG (rs360729, rs360714 and rs7106524), present with increased frequency within the age group ≥41 years (f = 0.375) compared to patients within age group 1–9 years (f = 0.168), (χ2 = 4.917; p = 0.027), while haplotype AGG (rs360729, rs360714 andrs7106524) was present with increased frequency among the patients in the age group 1–9 years (f = 0.278), compared to patients above 41 years of age (f = 0.103), (χ2 = 5.687; p = 0.017).
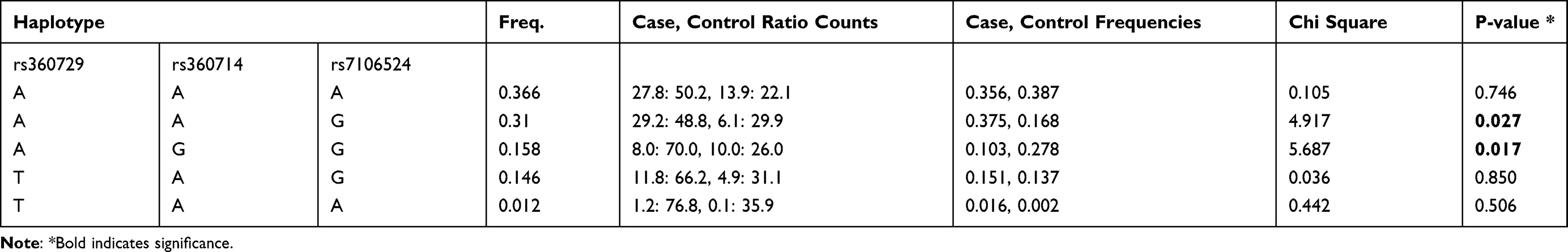 |
Table 9 Haplotype Analysis for IL-18 Gene Polymorphisms Between Groups (1–9 Years) Was Compared to 41 Years and Above |
Effect of IL-18 SNPs on Expression Levels in Luciferase Reporter Assay
Luciferase reporter assay was performed to investigate the effect of the SNPs in the promoter region of IL-18 on its expression levels. Several DNA fragments of the IL-18 promoter were cloned and transfected in HuH7 cells and the variations in the luciferase gene activity were monitored. Figure 2 shows the schematic representation of the influence of IL-18 promoter segments, with the corresponding SNPs, on its transcriptional activity. Three different promoter fragments represented as; fragment 1 [from −795 to +28], fragment 2 [from −795 to −523] and fragment 3 [from −182 to +28] were analyzed. We found that fragment 1 exhibited high luciferase activity compared to fragments 2 and 3 (Figure 2A). Fragment 1 includes the two SNPs, rs1946518 and rs187238 that were studied here. As such, we generated three additional fragments that contain different haplotype combinations. Haplotype TG (rs1946518 and rs187238) showed the highest luciferase activity (Figure 2B).
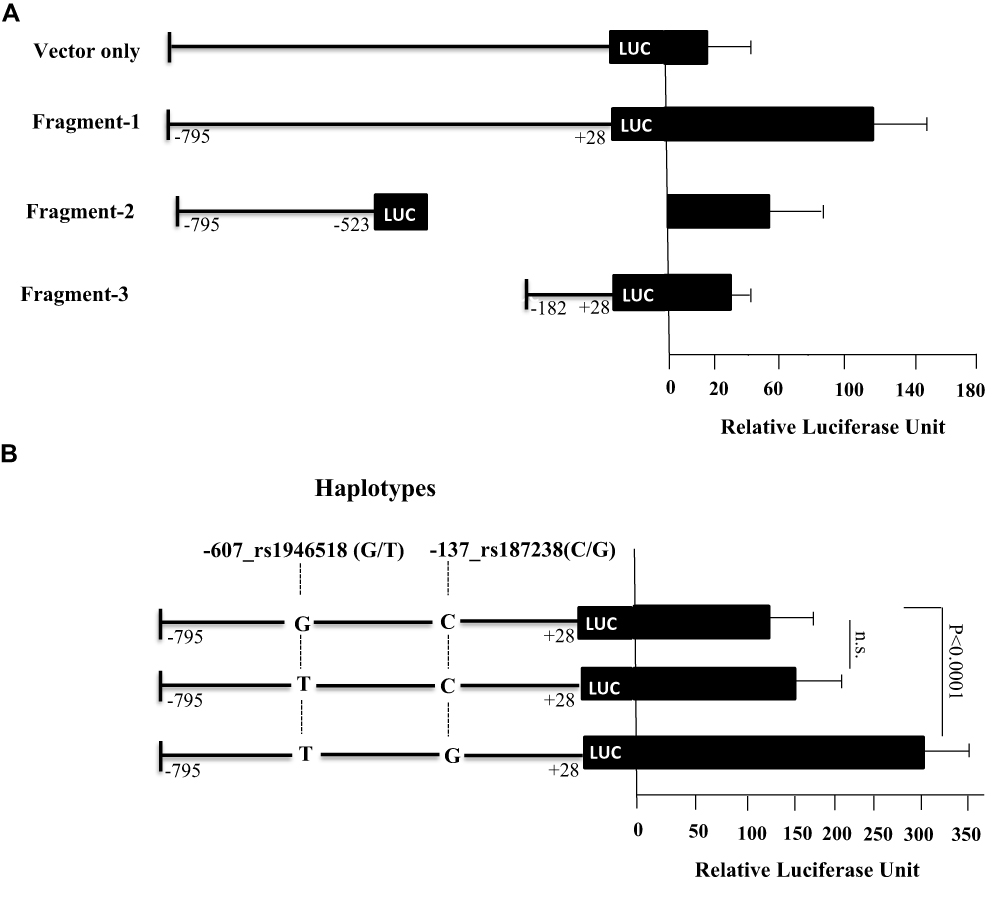 |
Figure 2 Luciferase activity assay for the fragments containing IL-18 promoter. (A) nested deletions of this region were generated, cloned into pGL3 luciferase vector and transfected in HuH7 cell line for 24 h. (B) As fragment 1 showed the highest luciferase activity, it was further tested to evaluate the effect of the SNPs included in this fragment on the expression activity. Cells were harvested and tested for luciferase activity after normalization using renilla luciferase enzyme. Each experiment was performed in triplicates. Values represent mean ±SD. |
The luciferase reporter assay was also conducted for different DNA segments of the 3ʹUTR region of the IL-18 gene. Three different fragments of the 3ʹUTR IL-18 region analyzed were represented as fragment 1 [from +20472 to +20984], fragment 2 [from +20500 to +20984] and fragment 3 [from +20472 to +20733]. Fragment 3 containing 2 SNPs, rs360728 and rs5744292, showed the highest level of luciferase activity (Figure 3A). As it showed the highest luciferase activity, fragment 3 was selected for haplotype expression analysis. Three different allelic combinations for SNPs rs360728 and rs5744292 were tested for their effect on luciferase expression. Haplotype CG (rs360728 and rs5744292) was found to have the highest luciferase activity (Figure 3B).
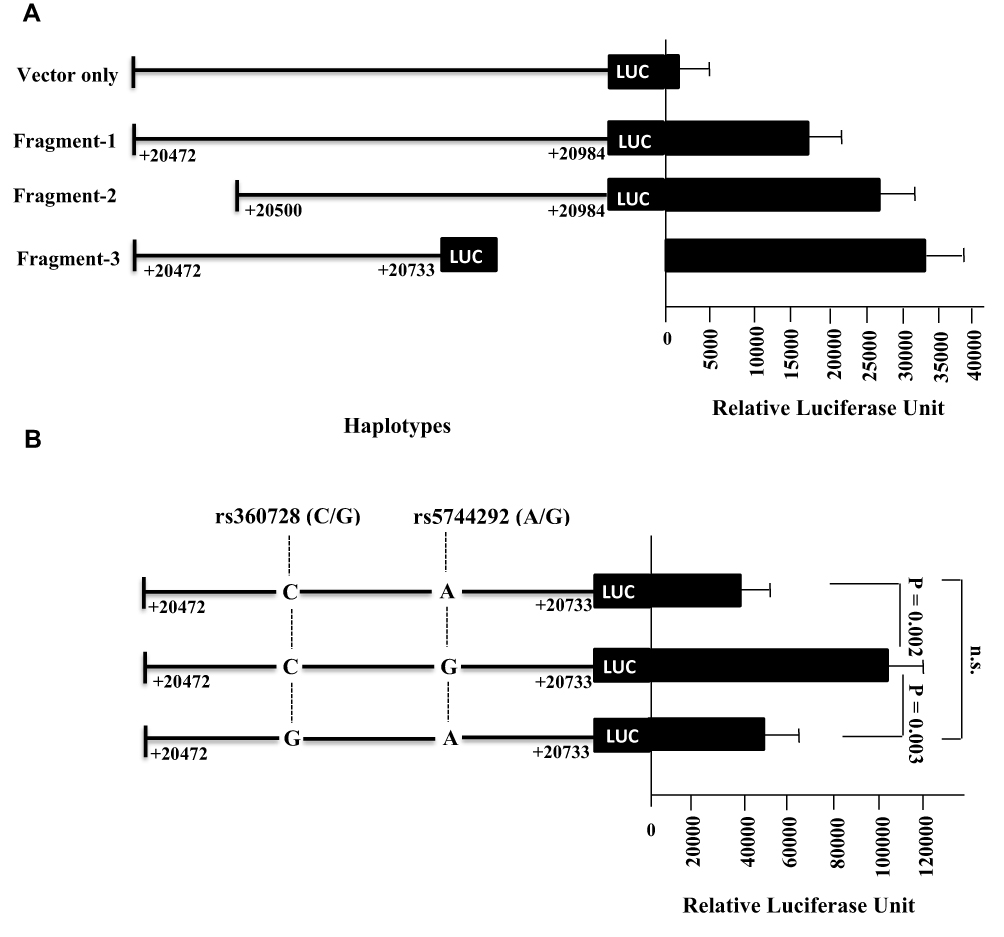 |
Figure 3 Luciferase activity assay for the fragments containing IL-18 3’ UTR. (A) nested deletions of this region were generated, cloned into pGL3 luciferase vector and transfected in HuH7 cell line for 24 h. (B) As fragment 3 showed the highest luciferase activity, it was further tested to evaluate the effect of the SNPs included in this fragment on the expression activity. Cells were harvested and tested for luciferase activity after normalization using renilla luciferase enzyme. Each experiment was performed in triplicates. Values represent mean ±SD. |
Discussion
IL-18, an important immune response mediator that has been reported to be involved in the course of malarial infection.20–24 IL-18 is known as a potent proinflammatory cytokine, and its physiological levels have been reported to increase at different stages of P. falciparum infection.31 Information about the influence of genetic variations within the IL-18 gene on parasitemia caused by P. falciparum is scarce. In this study, we examined the plausible role and influence of polymorphisms within the IL-18 gene on parasitemia levels in P. falciparum-infected patients. We found SNP rs5744292 to be significantly associated with an increased risk of P. falciparum infection when compared to healthy controls. It was observed that the rs5744292-G allele occurred more frequently in infected individuals than in the control group. rs5744292 has been implicated in several diseases such as cardiovascular diseases32,33 and coronary atherosclerosis.34 It has also been reported that a decrease in IL-18 concentration is associated with the G allele of rs5744292.25 We also identified another SNP, rs544354-A allele, to be significantly associated with an increased risk of parasitemia in infected patients. Further, individuals with homozygous AA genotype were found to have an elevated risk of parasitemia caused by P. falciparum. There is very little information available about the clinical impact of the SNP rs544354 which is located in the 3ʹUTR of the IL-18 gene.35 Another polymorphism found to be significantly associated with parasitemia in the infected patients was the rs360714-GG homozygote, present in the intron region of the IL-18 gene. However, this result needs to be interpreted with caution as the number of individuals in study population carrying GG genotype at the rs360714 SNP were very few.
SNP rs7106524, also located in the intron region of the IL-18 gene, has been investigated for its role in several clinical conditions such as those involving liver transplant.36,37 A previous study by He et al (2010) found an association between polymorphism rs7106524 with IL-18 levels, where the minor allele genotype (AA) was found to be associated with the highest concentration of serum IL-18.38 We found rs7106524 heterozygous GA genotype to be associated with an increased risk of P. falciparum parasitemia in comparison to the other two homozygous genotypes. Possibly, moderate levels of IL-18 linked to rs7106524-GA genotype may have a functional consequence in increasing the risk of malarial parasitemia and such a conclusion needs to be investigated further.
Haplotype analysis also revealed 2 haplotypes GT and AT (rs544354 and rs549908) that show significant differences between the controls and the infected patients. Haplotype GT was found more frequently in the controls whereas haplotype AT was found more frequently in the infected patient group. Haplotype AT includes rs544354 risk allele A whereas haplotype GT contains rs544354 allele G at the corresponding position. This haplotype analysis confirms the association of polymorphism rs544354 with an increased risk of P. falciparum infection.
We also analyzed the influence of these SNPs in the IL-18 gene on the extent of parasitemia among the P. falciparum-infected patients and found the rs5744292-G allele to be significantly associated with increased parasite density in patients. Polymorphisms within the cytokine genes have been reported to be associated with various degrees of parasitemia. Sortica et al (2013) identified variations in IL-6 and IL-12B to be associated with the levels of parasitemia in P. vivax-infected Brazilian population.39 Additionally, rs5744292 G-allele has been reported to be associated with downregulation of IL-18 levels.25,32 The cellular/molecular mechanism through which rs5744292-G allele contributes to an increase in parasitemia in P. falciparum-infected patients is unclear and needs further investigation. Three more SNPs, rs187238, rs360714 and rs1293344, were also found to be significantly associated with varying parasitemia levels in our study group. We observed that rs187238-CG, rs360714-GG and rs1293344-AG genotypes were found to be associated with protection from a higher degree of parasitemia. SNP rs187238, located in the promoter region of IL-18 gene, has been previously associated with hepatitis C and type 1 diabetes.40,41 Anyona et al (2011) observed that rs187238-C allele correlated with decreased IL-18 levels.23 The authors of this study suggested that this G to C substitution destroys a histone 4 transcription factor-1 (H4TF-1) nuclear factor-binding site, resulting in lower promoter activity with subsequent downregulation of IL-18 levels. Based on these results, it can be inferred that the protective effect of rs187238 heterozygote variants CG may be due to its regulation of IL-18 levels thereby controlling parasitemia.
Balavarca et al (2014) found that polymorphism rs1293344, located in the 5ʹ flanking region of the IL-18 gene, is associated with improved overall survival of colorectal cancer.42 Also, no significant association was found between polymorphism at rs1293344 and the changes in the serum concentration of IL-18.43 In this study, we found rs1293344-AG genotype to be correlated with improved disease outcomes by protecting against increased parasitemia in P. falciparum-infected patients.
Our analysis also identified several haplotypes with differential frequencies among infected patients with varying degrees of parasitemia. A comparison of patients with the lowest degree of parasitemia with those that had a higher degree of parasitemia revealed one significant haplotype. The haplotype CAT exhibited a higher frequency in group 1 parasitemia patients than the combined group 2+3+4 parasitemia patients. The mechanism through which this haplotype could affect the course of infection and/or expression of IL-18 requires further investigation.
Age is regarded as one of the crucial factors affecting the susceptibility and severity of the malarial infection.44 It has been observed to correlate with protective immunity in malaria-endemic areas.45 When compared to adolescents and adults, young children have been reported to be more susceptible to malarial infection, usually with severe complications that may be fatal if untreated.8,45,46 Due to the development of acquired immunity, adults are protected from severe malaria.46,47 In this study, we investigated the influence of the selected SNPs in the IL-18 gene within the different age groups of the P. falciparum-infected patients. On comparing genotypic distribution among patients in the 1–9 years group with patients in age group ≥10 years, 2 SNPs- rs544354 and rs360714 were found to show a significant association with the latter group. We found that increased frequency of rs544354-A allele was predictive of parasitemia in the P. falciparum-infected patients included in the 1–9 years age group. Additionally, the A allele had a similar association in both dominant and recessive modes of inheritance as well as heterozygotic AG genotype. Similarly, a higher prevalence of the rs360714-G allele was associated with an increased risk of parasitemia in the patients of age group 1–9 years. Our results are consistent with the observation in the Kenyan population, where parasitemia was found to be strongly dependent on the age factor, with children under 5 years exhibiting the highest malarial parasitemia levels.8 Further, another study has reported overall higher levels of parasitemia in children ≤9 years.48 IL-18 levels have also been reported to exhibit an age-related pattern in malarial patients.49,50 The functional mechanism(s) of these IL-18 SNPs must be studied to elucidate the role of these polymorphisms in determining age-related susceptibility to parasitemia.
We also performed a haplotype analysis between different age groups within the P. falciparum-infected patients. Single haplotype CA was found to be differentially distributed between the younger and older age groups. CA occurred more frequently within the 1–5 year age groups than in the older parasitemia patients. SNP rs187238 is positioned in the promoter region of the IL-18 gene and its C allele has been reported to correlate with decreased concentration of IL-18.23 The haplotype analysis among the parasitemia patients within the age groups 1–9 years and ≥ 41 years revealed the presence of two haplotypes AAG and AGG of SNPs rs360729, 360714 and rs7106524, respectively, which showed differential distribution between these groups. AAG occurred more frequently in older patients whereas haplotype AGG was found to occur at a higher frequency among the 1–9 years age group. Although rs360714 is located in the intron region of IL-18 gene, the two haplotypes, AAG and AGG differ only with regards to rs360714 A or G allele. This suggest that rs360714 does have some functional impact in different age groups, a conclusion that needs further testing to confirm its biological/physiological importance.
The functional activity of the SNPs in the promoter and 3ʹUTR of the IL-18 gene was analyzed by luciferase reporter assay. Two SNPs, rs1946518 and rs187238, of the promoter region within the IL-18 gene were found to modulate the transcriptional activity in the vector system. Our results are in agreement with the previous studies that have reported an association of promoter polymorphism with IL-18 expression. A study investigating patients with sarcoidosis analyzed the promoter activity by luciferase assay and found polymorphisms rs1946519 and rs1946518 to be significantly associated with serum levels of IL-18.51 Liang et al (2005) also reported the association of rs187238, among several other SNPs, with the transcriptional activity in the reporter assay system.52 Another study examining the functional activity of IL-18 promoter region found polymorphisms rs1946518 and rs187238 to be associated with the transcriptional activity.53 Similarly, the SNPs in the potential regulatory 3ʹUTR region have been shown to affect the expression levels of IL-18. Previous studies have reported that the 3ʹUTR region of the IL-18 gene contains regulatory elements that may be responsible for mRNA stability and translation rate.41,54 Polymorphism rs5744292 has been previously found to influence the IL-18 expression in various clinical conditions such as coronary artery disease patients.25,32 Although not much is known about the polymorphism rs360728 in relation to IL-18 expression, we found rs360728-C/rs5744292-G haplotype to be associated with enhanced luciferase activity. This was in contrast to the previous report that linked rs5744292-G containing haplotypes with decreased transcriptional activity,54 as well as our observation that rs5744292-G allele is linked to upregulation of IL-18 expression. These variations may be attributed to the fact that in-vitro studies using isolated fragments of the regulating regions may not be able to correctly reflect the functional activity in-vivo. Different elements including both cis- and trans-acting can influence the folding of the regulatory regions and subsequently their transcriptional activity. These results need to be confirmed in-vivo to clarify the role of these SNPs in modulating IL-18 expression.
In conclusion, this study has identified several individual polymorphisms and haplotypes within the pro-inflammatory IL-18 gene and revealed their association with the risk and extent of parasitemia in P. falciparum-infected patients. This is the first time such a detailed analysis of IL-18 gene polymorphism concerning malaria has been reported in Saudi Arabia, which in turn provides important insights into the role of population genetics in malaria.
Acknowledgments
The authors extend their appreciation to the Deputyship for Research and Innovation, Ministry of Education in Saudi Arabia for funding this research work through the project number KKU-420.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors have nothing to declare with respect to the research, authorship, and/or publication of this article.
References
1. World Health Organization. Malaria World Report. World Health Organization; 2019.
2. Hay SI, Guerra CA, Tatem AJ, Noor AM, Snow RW. The global distribution and population at risk of malaria: past, present, and future. Lancet Infect Dis. 2004;4(6):327–336. doi:10.1016/S1473-3099(04)01043-6
3. Coleman M, Al-Zahrani MH, Coleman M, et al. A country on the verge of malaria elimination--the Kingdom of Saudi Arabia. PLoS One. 2014;9(9):e105980. doi:10.1371/journal.pone.0105980
4. Snow RW, Amratia P, Zamani G, et al. The malaria transition on the Arabian Peninsula: progress toward a malaria-free region between 1960–2010. Adv Parasitol. 2013;82:205–251. doi:10.1016/B978-0-12-407706-5.00003-4
5. El Hassan IM, Sahly A, Alzahrani MH, et al. Progress toward malaria elimination in Jazan Province, Kingdom of Saudi Arabia: 2000–2014. Malar J. 2015;14:444. doi:10.1186/s12936-015-0858-1
6. Gething PW, Patil AP, Smith DL, et al. A new world malaria map: plasmodium falciparum endemicity in 2010. Malar J. 2011;10:378. doi:10.1186/1475-2875-10-378
7. Leoratti FM, Farias L, Alves FP, et al. Variants in the toll-like receptor signaling pathway and clinical outcomes of malaria. J Infect Dis. 2008;198(5):772–780. doi:10.1086/590440
8. Idris ZM, Chan CW, Kongere J, et al. High and Heterogeneous Prevalence of Asymptomatic and Sub-microscopic Malaria Infections on Islands in Lake Victoria, Kenya. Sci Rep. 2016;6:36958. doi:10.1038/srep36958
9. de Mendonca VR, Goncalves MS, Barral-Netto M. The host genetic diversity in malaria infection. J Trop Med. 2012;2012:940616. doi:10.1155/2012/940616
10. Kwiatkowski DP. How malaria has affected the human genome and what human genetics can teach us about malaria. Am J Hum Genet. 2005;77(2):171–192. doi:10.1086/432519
11. Gilles HM, Fletcher KA, Hendrickse RG, Lindner R, Reddy S, Allan N. Glucose-6-phosphate-dehydrogenase deficiency, sickling, and malaria in African children in South Western Nigeria. Lancet. 1967;1(7482):138–140. doi:10.1016/S0140-6736(67)91037-9
12. Ayi K, Turrini F, Piga A, Arese P. Enhanced phagocytosis of ring-parasitized mutant erythrocytes: a common mechanism that may explain protection against falciparum malaria in sickle trait and beta-thalassemia trait. Blood. 2004;104(10):3364–3371. doi:10.1182/blood-2003-11-3820
13. Imrie H, Ferguson DJP, Day KP. Human serum haptoglobin is toxic to Plasmodium falciparum in vitro. Mol Biochem Parasitol. 2004;133(1):93–98. doi:10.1016/j.molbiopara.2003.07.007
14. Lyke KE, Burges R, Cissoko Y, et al. Serum levels of the proinflammatory cytokines interleukin-1 beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis factor alpha, and IL-12(p70) in Malian children with severe Plasmodium falciparum malaria and matched uncomplicated malaria or healthy controls. Infect Immun. 2004;72(10):5630–5637. doi:10.1128/IAI.72.10.5630-5637.2004
15. Arend WP, Palmer G, Gabay C. IL-1, IL-18, and IL-33 families of cytokines. Immunol Rev. 2008;223:20–38. doi:10.1111/j.1600-065X.2008.00624.x
16. Dinarello CA, Fantuzzi G. Interleukin-18 and host defense against infection. J Infect Dis. 2003;187(Suppl 2):S370–84. doi:10.1086/374751
17. Dinarello CA. IL-18: a TH1-inducing, proinflammatory cytokine and new member of the IL-1 family. J Allergy Clin Immunol. 1999;103(1 Pt 1):11–24. doi:10.1016/s0091-6749(99)70518-x
18. Dinarello CA. The IL-1 family and inflammatory diseases. Clin Exp Rheumatol. 2002;20(5 Suppl 27):S1–13.
19. Greenlee W, Hagmann WK, Plattner JJ, et al. Annual Reports in Medicinal Chemistry. Elsevier Science; 1999.
20. Wroczynska A, Nahorski W, Bakowska A, Pietkiewicz H. Cytokines and clinical manifestations of malaria in adults with severe and uncomplicated disease. Int Marit Health. 2005;56(1–4):103–114.
21. Torre D, Giola M, Speranza F, Matteelli A, Basilico C, Biondi G. Serum levels of interleukin-18 in patients with uncomplicated Plasmodium falciparum malaria. Eur Cytokine Netw. 2001;12(2):361–364.
22. Malaguarnera L, Pignatelli S, Musumeci M, Simporè J, Musumeci S. Plasma levels of interleukin-18 and interleukin-12 in Plasmodium falciparum malaria. Parasite Immunol. 2002;24(9–10):489–492. doi:10.1046/j.1365-3024.2002.00485.x
23. Anyona SB, Kempaiah P, Raballah E, et al. Functional promoter haplotypes of interleukin-18 condition susceptibility to severe malarial anemia and childhood mortality. Infect Immun. 2011;79(12):4923–4932. doi:10.1128/IAI.05601-11
24. Kojima S, Nagamine Y, Hayano M, Looareesuwan S, Nakanishi K. A potential role of interleukin 18 in severe falciparum malaria. Acta Trop. 2004;89(3):279–284. doi:10.1016/j.actatropica.2003.10.005
25. Tiret L, Godefroy T, Lubos E, et al. Genetic analysis of the interleukin-18 system highlights the role of the interleukin-18 gene in cardiovascular disease. Circulation. 2005;112(5):643–650. doi:10.1161/CIRCULATIONAHA.104.519702
26. Huang XZ, Zhuang JH, Ren YG, Zhou LJ, Zhou Q. Association of interleukin-6 and interleukin-18 gene polymorphism with rheumatoid arthritis in Guangdong Han population. Nan Fang Yi Ke Da Xue Xue Bao. 2007;27(11):1661–1664.
27. Xu Q, Tin SK, Sivalingam SP, Thumboo J, Koh DR, Fong KY. Interleukin-18 promoter gene polymorphisms in Chinese patients with systemic lupus erythematosus: association withCC genotype at position −607. Ann Acad Med Singapore. 2007;36(2):91–95.
28. Cheng D, Hao Y, Zhou W, Ma Y. The Relationship between Interleukin-18 Polymorphisms and Allergic Disease: a Meta-Analysis. Biomed Res Int. 2014;2014:290687. doi:10.1155/2014/290687
29. Aljarba NH, Al-Anazi MR, Shafeai MI, et al. Interleukin-22 Polymorphisms in Plasmodium falciparum-Infected Malaria Patients. Mediators Inflamm. 2020;2020:5193723. doi:10.1155/2020/5193723
30. World Health Organization, Control CfD. Basic Malaria Microscopy: Tutor’s Guide. World Health Organization; 2010.
31. Nagamine Y, Hayano M, Kashiwamura S, et al. Involvement of interleukin-18 in severe Plasmodium falciparum malaria. Trans R Soc Trop Med Hyg. 2003;97(2):236–241. doi:10.1016/S0035-9203(03)90130-1
32. Opstad TB, Pettersen AA, Arnesen H, Seljeflot I. Circulating levels of IL-18 are significantly influenced by the IL-18 +183 A/G polymorphism in coronary artery disease patients with diabetes type 2 and the metabolic syndrome: an observational study. Cardiovasc Diabetol. 2011;10:110. doi:10.1186/1475-2840-10-110
33. Opstad TB, Pettersen AA, Arnesen H, Seljeflot I. The co-existence of the IL-18+183 A/G and MMP-9-1562 C/T polymorphisms is associated with clinical events in coronary artery disease patients. PLoS One. 2013;8(9):e74498. doi:10.1371/journal.pone.0074498
34. Hernesniemi JA, Karhunen PJ, Oksala N, et al. Interleukin 18 gene promoter polymorphism: a link between hypertension and pre-hospital sudden cardiac death: the Helsinki Sudden Death Study. Eur Heart J. 2009;30(23):2939–2946. doi:10.1093/eurheartj/ehp316
35. Sanchez E, Palomino-Morales RJ, Ortego-Centeno N, et al. Identification of a new putative functional IL18 gene variant through an association study in systemic lupus erythematosus. Hum Mol Genet. 2009;18(19):3739–3748. doi:10.1093/hmg/ddp301
36. Shi BJ, Yu XY, Li H, et al. Association between donor and recipient Interleukin-18 gene polymorphisms and the risk of infection after liver transplantation. Clin Invest Med. 2017;40(5):E176–E187. doi:10.25011/cim.v40i5.28623
37. Fan J, Zhang X, Ren L, et al. Donor IL-18 rs5744247 polymorphism as a new biomarker of tacrolimus elimination in Chinese liver transplant patients during the early post-transplantation period: results from two cohort studies. Pharmacogenomics. 2015;16(3):239–250. doi:10.2217/pgs.14.166
38. He M, Cornelis MC, Kraft P, et al. Genome-wide association study identifies variants at the IL18-BCO2 locus associated with interleukin-18 levels. Arterioscler Thromb Vasc Biol. 2010;30(4):885–890. doi:10.1161/ATVBAHA.109.199422
39. Sortica VA, Cunha MG, Ohnishi MD, et al. Role of IL6, IL12B and VDR gene polymorphisms in Plasmodium vivax malaria severity, parasitemia and gametocytemia levels in an Amazonian Brazilian population. Cytokine. 2014;65(1):42–47. doi:10.1016/j.cyto.2013.09.014
40. An P, Thio CL, Kirk GD, Donfield S, Goedert JJ, Winkler CA. Regulatory polymorphisms in the interleukin-18 promoter are associated with hepatitis C virus clearance. J Infect Dis. 2008;198(8):1159–1165. doi:10.1086/592047
41. Thompson SR, Humphries SE. Interleukin-18 genetics and inflammatory disease susceptibility. Genes Immun. 2007;8(2):91–99. doi:10.1038/sj.gene.6364366
42. Balavarca Y, Habermann N, Scherer D, et al. Genetic variants in inflammation‐related genes and interaction with NSAID use on colorectal cancer risk and prognosis. Genes, Chromosomes & Cancer. 2014;53(7):568–578. doi:10.1002/gcc.22167
43. Frayling TM, Rafiq S, Murray A, et al. An interleukin-18 polymorphism is associated with reduced serum concentrations and better physical functioning in older people. J Gerontol a Biol Sci Med Sci. 2007;62(1):73–78. doi:10.1093/gerona/62.1.73
44. Carneiro I, Roca-Feltrer A, Griffin JT, et al. Age-patterns of malaria vary with severity, transmission intensity and seasonality in sub-Saharan Africa: a systematic review and pooled analysis. PLoS One. 2010;5(2):e8988. doi:10.1371/journal.pone.0008988
45. Laishram DD, Sutton PL, Nanda N, et al. The complexities of malaria disease manifestations with a focus on asymptomatic malaria. Malar J. 2012;11:29. doi:10.1186/1475-2875-11-29
46. Doolan DL, Dobano C, Baird JK. Acquired immunity to malaria. Clin Microbiol Rev. 2009;22(1):13–36. doi:10.1128/CMR.00025-08
47. Grobusch MP, Kremsner PG. Uncomplicated malaria. Curr Top Microbiol Immunol. 2005;295:83–104.
48. Bassey SE, Nwakaku IL. Prevalence of Malaria Parasitemia among Children between 1–10 Years Old Attending Federal Medical Centre, Yenagoa, Bayelsa State, Nigeria. EC Pharmacol Toxicol. 2017;3(2):54.
49. Tatfeng YM, Agbonlahor DE. Age-related cytokine profile in uncomplicated Plasmodium malaria infection. Biochem Biophys Res Commun. 2011;41:2011.
50. Sawian CE. Contribution of Innate Responses to Plasmodium Falciparum Infections. Tezpur, Assam, India: Tezpur University; 2013.
51. Zhou Y, Yamaguchi E, Hizawa N, Nishimura M. Roles of functional polymorphisms in the interleukin-18 gene promoter in sarcoidosis. Sarcoidosis Vasc Diffuse Lung Dis. 2005;22(2):105–113.
52. Liang XH, Cheung W, Heng CK, Wang DY. Reduced transcriptional activity in individuals with IL-18 gene variants detected from functional but not association study. Biochem Biophys Res Commun. 2005;338(2):736–741. doi:10.1016/j.bbrc.2005.10.012
53. Giedraitis V, He B, Huang WX, Hillert J. Cloning and mutation analysis of the human IL-18 promoter: a possible role of polymorphisms in expression regulation. J Neuroimmunol. 2001;112(1–2):146–152. doi:10.1016/S0165-5728(00)00407-0
54. Barbaux S, Poirier O, Godefroy T, et al. Differential haplotypic expression of the interleukin-18 gene. Eur J Hum Genet. 2007;15(8):856–863. doi:10.1038/sj.ejhg.5201842
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.