Back to Journals » Drug Design, Development and Therapy » Volume 17
Antimicrobial Peptide Cec4 Eradicates Multidrug-Resistant Acinetobacter baumannii in vitro and in vivo
Authors Peng J, Wang Y, Wu Z, Mao C, Li L, Cao H, Qiu Z, Guo G, Liang G, Shen F
Received 20 January 2023
Accepted for publication 22 March 2023
Published 30 March 2023 Volume 2023:17 Pages 977—992
DOI https://doi.org/10.2147/DDDT.S405579
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Anastasios Lymperopoulos
Jian Peng,1– 3,* Yue Wang,3,* Zhaoyin Wu,3 Chengju Mao,3 Lu Li,1,2 Huijun Cao,4 Zhilang Qiu,3 Guo Guo,3,5 Guiyou Liang,4,5 Feng Shen1,2
1Department of Intensive Care Unit, Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, 550009, People’s Republic of China; 2Guizhou Institute of Precision Medicine, Affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, 550009, People’s Republic of China; 3Key Laboratory of Infectious Immune and Antibody Engineering of Guizhou Province, Cellular Immunotherapy Engineering Research Center of Guizhou Province, School of Biology and Engineering/School of Basic Medical Sciences, Guizhou Medical University, Guiyang, Guizhou, 550025, People’s Republic of China; 4Department of Cardiac Surgery, the affiliated Hospital of Guizhou Medical University, Guiyang, Guizhou, 550009, People’s Republic of China; 5Translational Medicine Research Center, Guizhou Medical University, Guiyang, Guizhou, 550025, People’s Republic of China
*These authors contributed equally to this work
Correspondence: Feng Shen; Guiyou Liang, Email [email protected]; [email protected]
Introduction: Acinetobacter baumannii has become a major difficulty in the treatment of bacteria-associated infection. The previously reported antimicrobial peptide Cec4 exhibited good and stable activity against A. baumannii in vitro, but the mechanisms and effects in vivo are elusive.
Methods: The effects of Cec4 on bacterial membrane permeability, membrane potential and bacterial reactive oxygen species were measured. The cell membrane localization of antimicrobial peptides was studied by fluorescence labelling. The ability of bacteria to develop resistance to antimicrobial peptides was studied by continuous induction, and transcriptome difference was analysed. The in vivo toxicity of Cec4 against nematodes and mice was studied, and the in vivo therapeutic potential of Cec4 against A. baumannii was assessed.
Results: Cec4 effectively cleared multidrug-resistant A. baumannii by altering bacterial cell membrane permeability, changing bacterial cell membrane polarity, and increasing bacterial intracellular reactive oxygen species. Cec4 affected the expression of the secretion system, outer membrane, and efflux pump genes of A. baumannii. In addition, the bacteria did not acquire stable drug-resistant ability. Cec4 at 1.024 mg/mL did not affect the proliferation of HeLa and HepG2 cells, and Cec4 at 45 mg/kg had little effect on the mortality of Caenorhabditis elegans, even the liver and kidney tissues of mouse. Most importantly, Cec4 could effectively improve the survival rates and reduce the bacterial load of various tissues in the mouse model of infection.
Conclusion: In conclusion, Cec4 can damage the cell membrane of bacteria, and the bacteria is not easy to produce resistance to Cec4. Besides, Cec4 has good potential for the treatment of multidrug-resistant A. baumannii infections.
Keywords: antimicrobial peptide, A. baumannii, sepsis, skin infection, drug safety
A Letter to the Editor has been published for this article.
Introduction
Acinetobacter baumannii, as a Gram-negative aerobic bacterium, is commonly found in the clinical intensive care unit, haematology department, infectious department, urology department, etc. The bacteria are mainly transmitted indirectly through the surface of objects, and with the spread of COVID-19, respiratory and associated A. baumannii infections are also on the rise.1 The emergence of drug-resistant A. baumannii has led to increasingly serious problems in the treatment of clinical infectious diseases. It has been reported that patients infected with carbapenem-resistant pathogens have higher morbidity and mortality compared to patients infected with susceptible pathogens.2,3 For the moment, carbapenem antibiotics are the most-used first-line antibiotics for the treatment of Gram-negative infections, and the accelerated emergence of carbapenem-resistant A. baumannii (CRAB) is a serious threat to global public health.4 Since 2017, when the World Health Organisation focused on CRAB, various potential drugs and therapies against CRAB have been proposed. For example, New β-lactam and β-lactamase inhibitor combination therapies have played a role in the mitigation of CRAB infections.5 Tigecycline and polymyxin have become limited drugs for the treatment of carbapenem-resistant bacteria, but the efficacy and toxicity of tigecycline and polymyxin in the treatment of infection are problems.6 On the other hand, colistin-resistant A. baumannii has emerged, and there is no other way for the treatment of colistin-resistant bacteria except combination therapy7 and the development of new drugs.
Antimicrobial peptides (AMPs) protect the host from pathogens as part of innate immunity.8 AMPs can eliminate multiple and pan-resistant bacteria,9 and new peptide biotechnologies and targeted transport increase the antimicrobial activity of AMPs.10 Most AMPs have less than 50 amino acids, and the current AMPs are against drug-resistant A. baumannii include cathepsins, defensins,11 frog antimicrobial peptides, and insect antimicrobial peptides,12 modified peptides based on tilapia antimicrobial 2,13 and other natural or engineered small molecular proteins.14,15 According to the summary,16 most of the above AMPs against A. baumannii have α-helical structures. More surprisingly, some of the AMPs are even more effective than conventional antibiotics in killing drug-resistant microbial pathogens.16 Studies have shown that the mechanisms of direct attack of these peptides on bacteria are usually divided into two categories: bacterial membrane attack and intracellular targeting, and some peptides can possess both mechanisms.17,18 In the development and use of AMPs, many researchers have used them in combination with one,2 or multiple antibiotics19 to improve the efficiency of treatment of microbial infections.
In the previous study, our group found that the minimum inhibitory concentration (MIC) of Cec4 was 4 μg/mL against the standard strain of A. baumannii (ATCC19606) in the planktonic state, the inhibitory effect was better than that of the reported similar AMP Cec1 and cecropin A.20,21 Furthermore, Cec4 was shown to inhibit CRAB biofilms,22 but the specific mechanism of Cec4 antimicrobial activity and its toxicity and antimicrobial activity in vivo are elusive. Therefore, this study focused on exploring the specific mechanism of Cec4 antibacterial activity and assessing the toxicity of the antimicrobial peptide Cec4 by its effects on the proliferation of human cervical cell lines, hepatocyte lines, and on C. elegans or mice. Finally, the potential of Cec4 as a drug for the treatment of A. baumannii-associated infections was evaluated using C. elegans and mice-infected model with multidrug-resistant A. baumannii.
Materials and Methods
Bacterial Strains, Culture Conditions, and Mice Source
The standard strain of A. baumannii (ATCC19606) was kept by the Key Laboratory of Pathogen Biology, Guizhou Medical University. The multidrug-resistant A. baumannii (AB4367992) and the clinical multidrug-resistant A. baumannii were isolated from a tertiary hospital in Guizhou Province. Uracil-deficient E. coli OP50 and N2 wild-type C. elegans were gifted by Professor Yu-Cong Xie, Southern University of Science and Technology. Both E. coli OP50 and A. baumannii were grown in Luria–Bertani (LB) medium and incubated overnight at 37°C. SPF-grade female BALB/c mice (6–8 weeks, 17–21 g) were purchased from SPF (Beijing) Biotechnology Co., LTD. The experiments were approved by the Institutional Animal Care and Use Committee of Guizhou Medical University.
Peptide Synthesis
The antimicrobial peptide Cec4 with 41 amino acids (GWLKKIGKKIERVGQNTRDATIQAIGVAQQAANVAATLKGK) in this study and fluorescein isothiocyanate (FITC)-labelled Cec4 were synthesised by solid-phase chemical synthesis, and purified by HPLC method with purity >95% from Shanghai Gill Biochemical Co.
Antimicrobial Susceptibility Assays
MICs were determined by the microdilution method,23 and the concentration of the bacterial solution was adjusted to 1×106 CFU (Colony Forming Unit)/mL. And 100 μL of MH medium and twofold gradient dilution of 1–128 μg/mL of Cec4 were added in 96-well polypropylene plates. Automated antimicrobial susceptibility test (AST) determinations were made using the VITEK 2 XL (bioMérieux) according to the manufacturer’s instructions.
Membrane Permeability
The effect of the antimicrobial peptide Cec4 on the permeability of the inner membrane of ATCC 19606 cells was reflected by detecting changes in the fluorescence intensity of propidium iodide (PI).24 The bacteria were cultured to the logarithmic growth stage and adjusted to a concentration of 1×108 CFU/mL with phosphate buffer saline (PBS). The final concentrations of Cec4 were 8, 16, 32, and 128 μg/mL.
Membrane Potential
The effect of Cec4 on the cell membrane potential of ATCC 19606 was detected using a cell membrane potential-sensitive probe 3,3’dipropylthiocarbocyanine iodide (DiSC3-5), as described by Liu et al.25 The bacterial concentration is the same as 2.4. The final concentrations of Cec4 were 4 μg/mL, 8 μg/mL, 16 μg/mL, 32 μg/mL, and 128 μg/mL.
Intracellular Reactive Oxygen Species (ROS)
ROS levels in Cec4-treated ATCC 19606 were detected by 2’,7’-dichlorofluorescein diacetate (DCFH-DA).26 The bacterial concentration is the same as 2.4. Fifty microlitres of bacteria were mixed with the dye, and equal volumes of different concentrations of Cec4 solutions (final concentrations with 8, 16, 32, and 64 μg/mL) were added to the mixtures and incubated for 1 h.
Laser Confocal Experiment
The targeting of Cec4 in ATCC 19606 was observed using laser confocal microscopy.27 Because fluorescein isothiocyanate (FITC) glows green after laser excitation, it can be used to locate the intracellular distribution of FITC-labelled antimicrobial peptide. The bacterial concentration is the same as 2.4. Cec4 was labelled by fluorescein isothiocyanate (FITC) with green fluorescence, cell membranes were labelled with 1,1’-dioctadecyl-3,3,3’,3’-tetramethylindole carbocyanine perchlorate (DiI) with red fluorescence, and DNA was labelled with 4’,6-diamidino-2-phenylindole (DAPI), showing blue fluorescence. The above mixture was incubated with Cec4 at a final concentration of 16 μg/mL for 1 h. After that, DiI and DAPI were added to the mixture, both at a final concentration of 1 µM, and incubated for 10 min.
Bacterial RNA Extraction and Transcriptome Analysis
The RNA extraction was performed according to the instruction manual of the RNA extraction kit (Axygen, Sigma-Aldrich, St. Louis, MO, USA).28 Total RNA was detected by 1% agarose gel electrophoresis, and the concentration and quality of total RNA were determined by Nanodrop Spectrophotometer (ND-200, Thermo Scientific, Waltham, MA, USA). The remaining RNA was stored at −20 °C. The library construction and sequencing analysis were entrusted to Beijing Novogene Technology Co.
Induction of Drug Resistance in ATCC19606
The bacterial suspension exposed to 0.5 × MIC was resuspended and grown to the logarithmic phase to determine the MIC with corresponding drugs.28 The above steps were repeated to 30 generations, and then the 30th-generation strain was subcultured without Cec4 for 10 generations. C-7 is the Cec4-resistant isolate at seventh passage from A. baumannii ATCC19606 cultured in the presence of Cec4 sub-inhibitory concentrations.
Cell Culture and Cytotoxicity Assay
HeLa cells (ATCC, Rockville, USA) and HepG2 (ATCC, Rockville, USA) cells are frequently used to evaluate the cytotoxicity of various compounds, and they were cultured in DMEM medium, and all cells were supplemented with 10% fetal bovine serum at 37 °C and 5% CO2.29 After the cells were attached to the wall, carboplatin injection (Qilu Pharmaceutical Co., LTD) at a final concentration of 1.024 mg/mL was added to the positive control group, antibacterial peptide Cec4 at a final concentration of 1.024 mg/mL was added to the experimental group, and an equal volume of PBS was added to the negative control group, with five parallel wells for each group. After the above mixture was incubated for 24 h, the inhibition rate of Cec4 was operated and calculated according to the instructions of the CCK-8 kit (CCK-8, APE-BIO, Houston, Texas, USA). Three replicates of each experiment were performed.
In vivo Toxicity Experiments
Following the method30, different concentrations of Cec4 (final concentrations with 2048, 1024, 512, 256 and 128 μg/mL) were added to each well of the experimental group, and erythromycin dissolved in 5% methanol (final concentration of 2048 μg/mL) was added as the positive control,31,32 mice in the experimental group were injected intraperitoneally with 45 mg/kg of Cec4. After 24 h, six mice in each group were selected and executed after anaesthesia, and blood was collected. The retained liver and kidney tissues were fixed in 10% formalin neutral buffer and paraffin embedded. The tissues were cut into 4–6 mm thick sections, stained with haematoxylin-eosin. Then, the sections were scanned for histopathological observation using a 3D HISTECH Pannoramic250 scanner, and the major lesion sites in the sections were intercepted and pathologically described.
C. elegans Infection Model
Following the method33,34 with slight modifications, there were 20–35 nematodes in each hole of the 96-well plate, and the nematodes were infected with a final concentration of 5×106 CFU/mL of AB4367992. Cec4 at a final concentration of 16 μg/mL and Polymyxin B (PB) (Sigma-Aldrich, St. Louis, MO, USA) at a final concentration of 2.5 μg/mL were added to different wells. The survival rate was determined by sinusoidal movement of living nematodes, while dead nematodes were rod-shaped or rod-shaped and did not respond to external stimuli. The survival of nematodes in each group within 7 days was counted and observed every 24 hours. Nine hours after infection with C. elegans, they were treated with drugs for 24 h to detect the number of bacteria in each C. elegans according to the method in the reference.
Mouse Skin Infection Model
Following the method,35,36 36 female BALB/c mice (6–8 weeks) were randomly divided into 3 groups (n = 12), and 5×107 CFU of AB4367992 was applied to 1 cm2 of the marked depilatory area and allowed to colonise for 16–18 h. Previous studies have found that Cec4 and polymyxin B with 1×MIC concentration have bacteriostatic effects on multi-drug resistance A. baumannii AB4367992 for nearly 24 hours and have obvious bactericidal effects in 20 minutes.21 Infected mice were treated with 2.5 μg/mL of PB or 16 μg/mL of Cec4 for 2 h, respectively, and the negative control group was treated with saline. The drugs were dissolved in 100 μL PBS, then soaked in a single layer of gauze for wet compress treatment. After 2 h of treatment, the mice were anaesthetised and executed, and the traumatised skin was peeled off and homogenised in 500 μL saline for 1–2 min.
Peritonitis Model in Mice
Following the method,37 80 female BALB/C mice were divided into 5 groups (n = 16), and the control mice were injected intraperitoneally with sterile saline and an equal volume of 10% porcine mucin. Other BALB/c mice were injected intraperitoneally with 5×106 CFU of AB4367992 (200 μL/each) and an equal volume of 10% porcine mucin to establish a mouse model of peritonitis. 2 h after infection, the mice were treated with PB (2.5 mg/kg), Cec4 (16 mg/kg), or PB combined with Cec4 (2.5 mg/kg PB + 16 mg/kg Cec4), respectively. After 4 h of treatment, six mice in each group were taken, anaesthetised, and executed, and blood, liver, spleen, lung, and kidney samples were obtained by aseptic manipulation for bacterial enumeration. The remaining mice were observed for 72h, every 12h, and the survival rate was calculated.
Statistical Analysis
All curves were fitted using GraphPad Prism 5.0 (GraphPad Prism Software, San Diego, CA) software. The Kaplan–Meier method was used for survival analysis, and the Log rank test was used to analyse the significance of differences between groups. The differences were considered statistically significant at P < 0.05.
Results
Antimicrobial Effect of Cec4 on Clinically Resistant Isolates
The antimicrobial effect of the antimicrobial peptide Cec4 was determined in vitro by measuring the MICs of CRAB (n = 21) sampled from patients with bloodstream infections in a hospital in Guizhou, China. There were 19 strains of multidrug-resistant A. baumannii and two strains of multidrug-resistant A. baumannii (polymyxin B resistant). These clinical strains were resistant to ertapenem, cefepime, levofloxacin, meropenem, etc (Supplementary Table S1). The MIC of Cec4 against the 19 extracted CRAB was 4–8 μg/mL. In addition, Cec4 was also effective against polymyxin B-resistant A. baumannii with an MIC of 4 μg/mL.
Cec4 Disrupts the Cell Membrane of A. baumannii and Promotes the Production of ROS
To determine the site of action of Cec4, laser confocal experiments were performed to observe the targeting position of Cec4 (Figure 1A). The results showed that FITC-Cec4 had good colocalization with DiI in A. baumannii, indicating that Cec4 exerted bactericidal effects by binding to the bacterial cell membrane. Besides, the changes of bacterial cell membrane fluidity indicated that the fluorescence intensity of PI gradually became stronger and showed dose and time dependence, and a significant increase in fluorescence intensity could be observed starting from 8 µg/mL (Figure 1B). In addition, the results of membrane potential showed that different concentrations of the antimicrobial peptide Cec4 caused an increase in fluorescence intensity in a dose-dependent manner (Figure 1C). The production of ROS is thought to be a key factor in antibiotic-mediated bactericidal activity. The results showed that the ROS level of A. baumannii increased significantly starting at 8 µg/mL and stabilised after 16 µg/mL (Figure 1D).
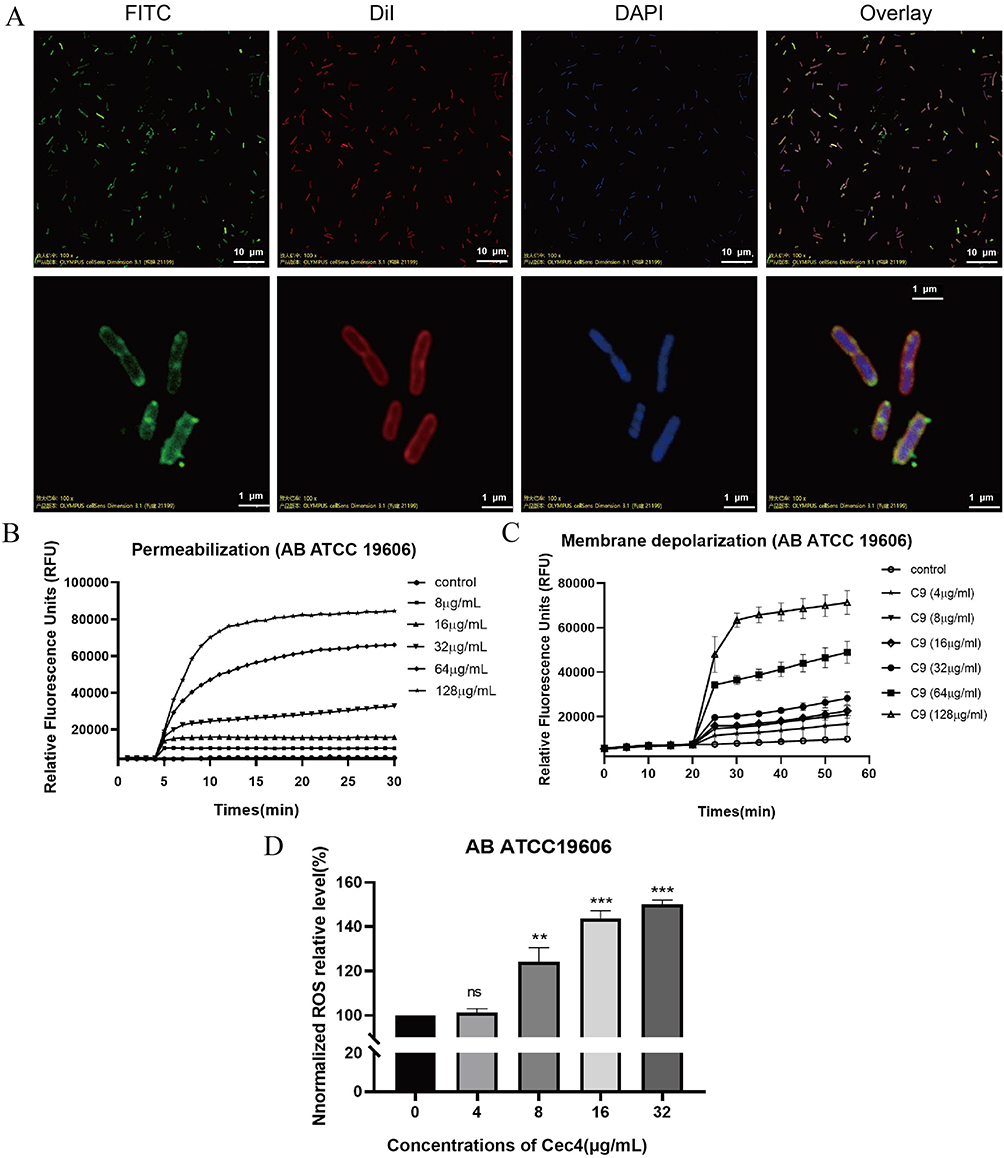 |
Figure 1 The inhibition mechanism of Cec4 against A. baumannii in vitro. (A) FITC-labelled Cec4 colocalized with the cell membrane of A. baumannii. Cec4 labelled by FITC (green) colocalizes cell membranes labelled by DiI (red) and bacterial DNA labelled by DAPI (blue). (B) The effect of Cec4 in enhanced bacterial cell permeability. With the increase of Cec4 concentration, the increase of PI fluorescence intensity indicated the increase of bacterial cell membrane permeability. (C) After Cec4 treatment, the cell membrane of A. baumannii was depolarized. The probe was aggregated and quenched in the normal cell membrane. With the increase of Cec4 treatment concentration, the fluorescence of the probe was enhanced, indicating that the bacterial cell membrane potential was depolarized. (D) The intracellular reactive oxygen species of A. baumannii increased after Cec4 treatment. When the dye enters the cell, it is oxidized and then fluoresces. With the increase of Cec4 concentration, fluorescence enhancement indicated the increase of intracellular ROS. Results were presented as mean ± SD. **P < 0.01, ***P < 0.001. |
Differential Analysis of the Transcriptome of A. baumannii After Cec4 Treatment
To investigate the transcriptional changes of genes in A. baumannii after Cec4 treatment, transcriptome sequencing analysis was performed after treatment of ATCC19606 with sub-repressive concentrations of Cec4 for 24 h. The results indicated that 111 differential genes (DEGs) were obtained after Cec4 treatment, including 88 upregulated genes and 23 downregulated genes (Figure 2A). After clustering, the downregulated genes in the treated group related to pilus (A4U85_RS01480), ABC transport-related genes (A4U85_RS10590), haemolysin secretion/activation protein (A4U85_RS08295), endosomal components of the binding protein-dependent transport system (A4U85_ RS10600), etc (Figure 2B, Table 1). Upregulated genes were significantly enriched on the outer membrane in GO enrichment (P < 0.05), including two genes and one sRNA (sRNA00016/A4U85_RS14860/A4U85_RS17265) (Figure 2C). A total of 19 pathways were enriched in the Kyoto Encyclopaedia of Genes and Genome (KEGG) database, including the significantly enriched ABC transport system (P < 0.05). MacB gene was significantly upregulated in this pathway. At the same time, the enriched pathways included ribosomal and amino acid biosynthesis processes (Figure 2D), which were involved in the process of macrolide antibiotics and bacterial efflux. Therefore, A. baumannii may antagonize Cec4 mainly in a macrolide-like manner upon Cec4 stimulation.
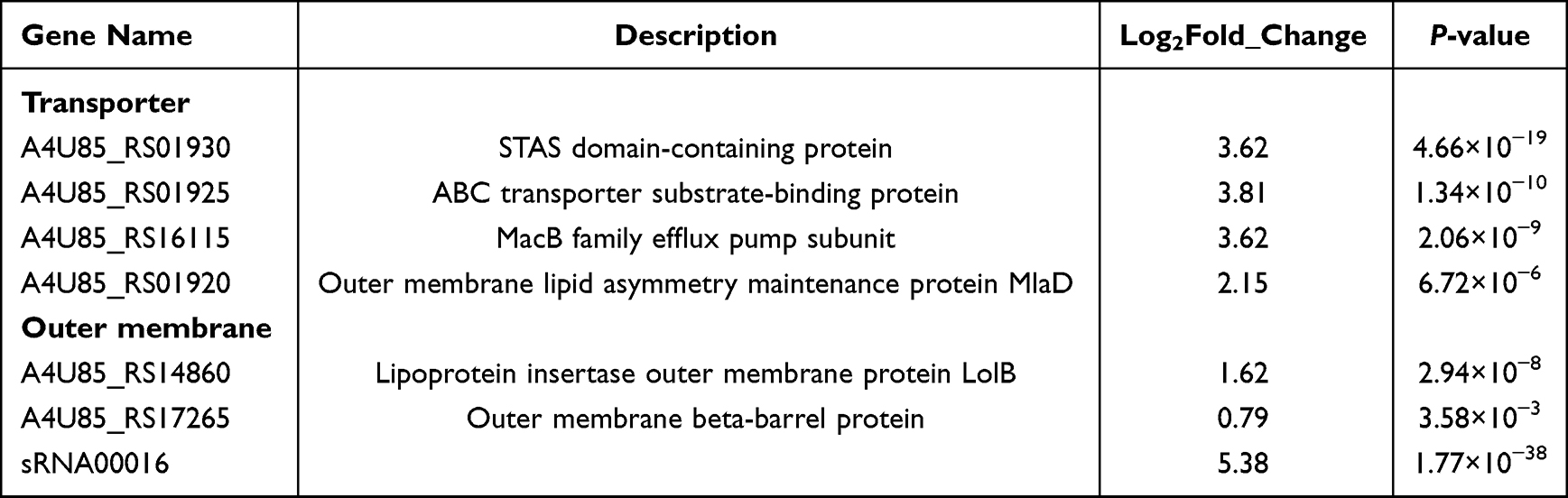 |
Table 1 Some Differential Genes Before and After Cec4 Treatment |
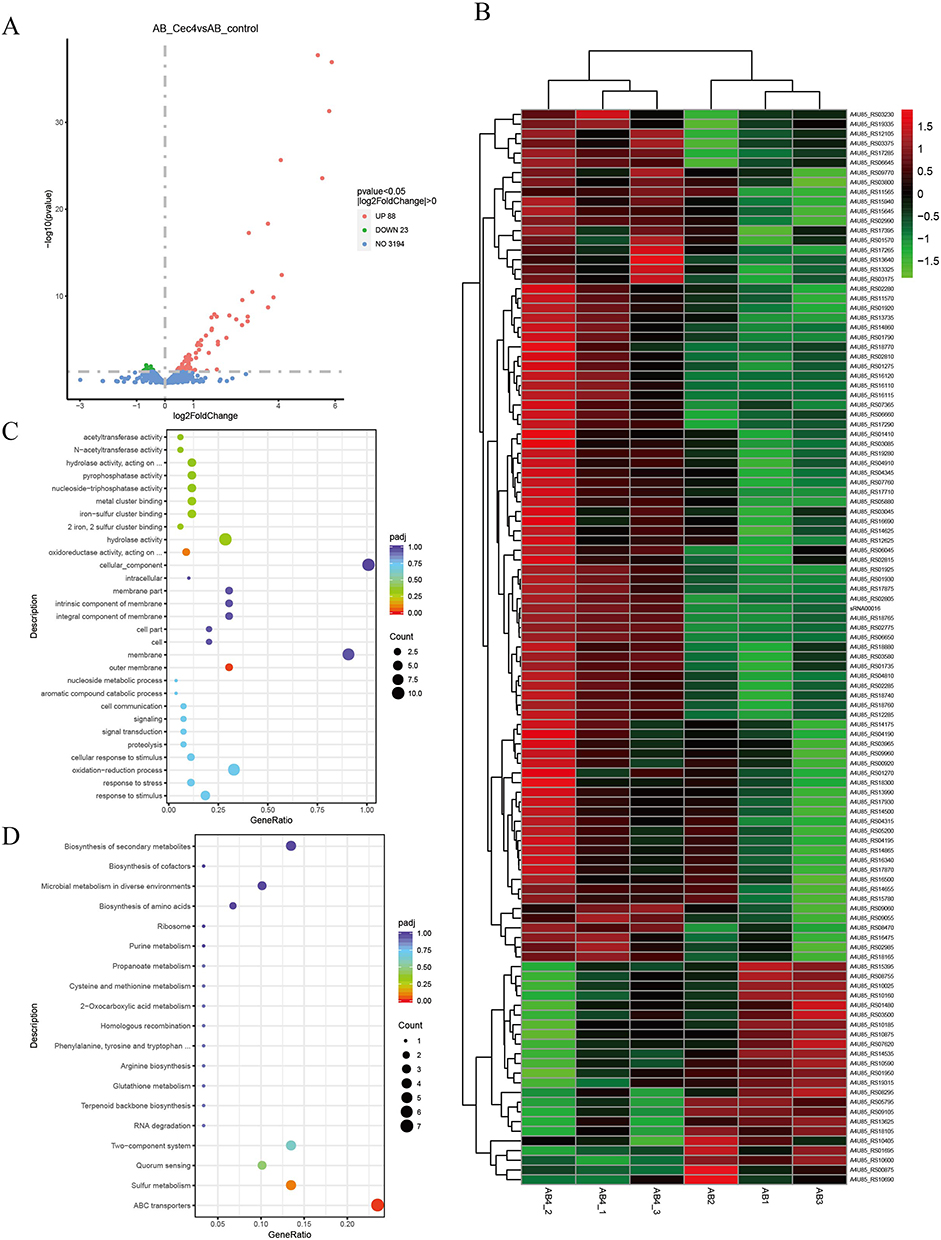 |
Figure 2 The transcriptome analysis of A. baumannii inhibited by Cec4. (A) The volcanic map of A. baumannii differential gene before and after treatment with subinhibitory concentration Cec4. (B) The differential gene expression heat map in different groups. After standardization, green indicated down-regulated gene, while red indicated up-regulated gene. (C) The results of DEGs’ GO enrichment analysis. The size of bubble represents the number of genes enriched in this Term, and the color of bubble represents the enriched padj value. (D) The results of DEGs’ KEGG enrichment analysis. The size of bubble represents the number of genes enriched in this pathway, and the color of bubble represents the enriched padj value. |
Cec4 Exhibits a Low Propensity to Induce Drug Resistance
To investigate whether long-term induction of Cec4 leads to bacterial resistance, ATCC19606 was induced in vitro. The results of passage induction showed that the resistance of A. baumannii to Cec4 was twice that of the primary generation. The resistance increased at the 7th generation (C-7), but the resistance did not increase at the 7th to 30th generations, indicating that Cec4-induced resistance was not stable (Figure 3A). To explore the potential mechanism of Cec4 resistance, transcriptome sequencing was performed on C-7 and ATCC19606. The results showed that a total of 212 DEGs were obtained for drug-resistant strain C-7 compared with wild strain ATCC19606, of which 48 were upregulated and 164 were downregulated (Figure 3B). It is worth noting that most of the exocytosis-related genes were upregulated by more than 1.9-fold. For example, the multidrug exocytosis RND transporter protein permease subunit was up-regulated by 2.35-fold, and the CusA/CzcA series heavy metal exocytosis RND transporter protein was up-regulated by 1.91-fold. The adeC/adeK/oprM class multidrug exocytosis complex outer membrane factor was up-regulated by 1.90-fold, and the MacB class exocytosis pump unit was up-regulated by 2.16-fold (Figure 3C, Table 2).
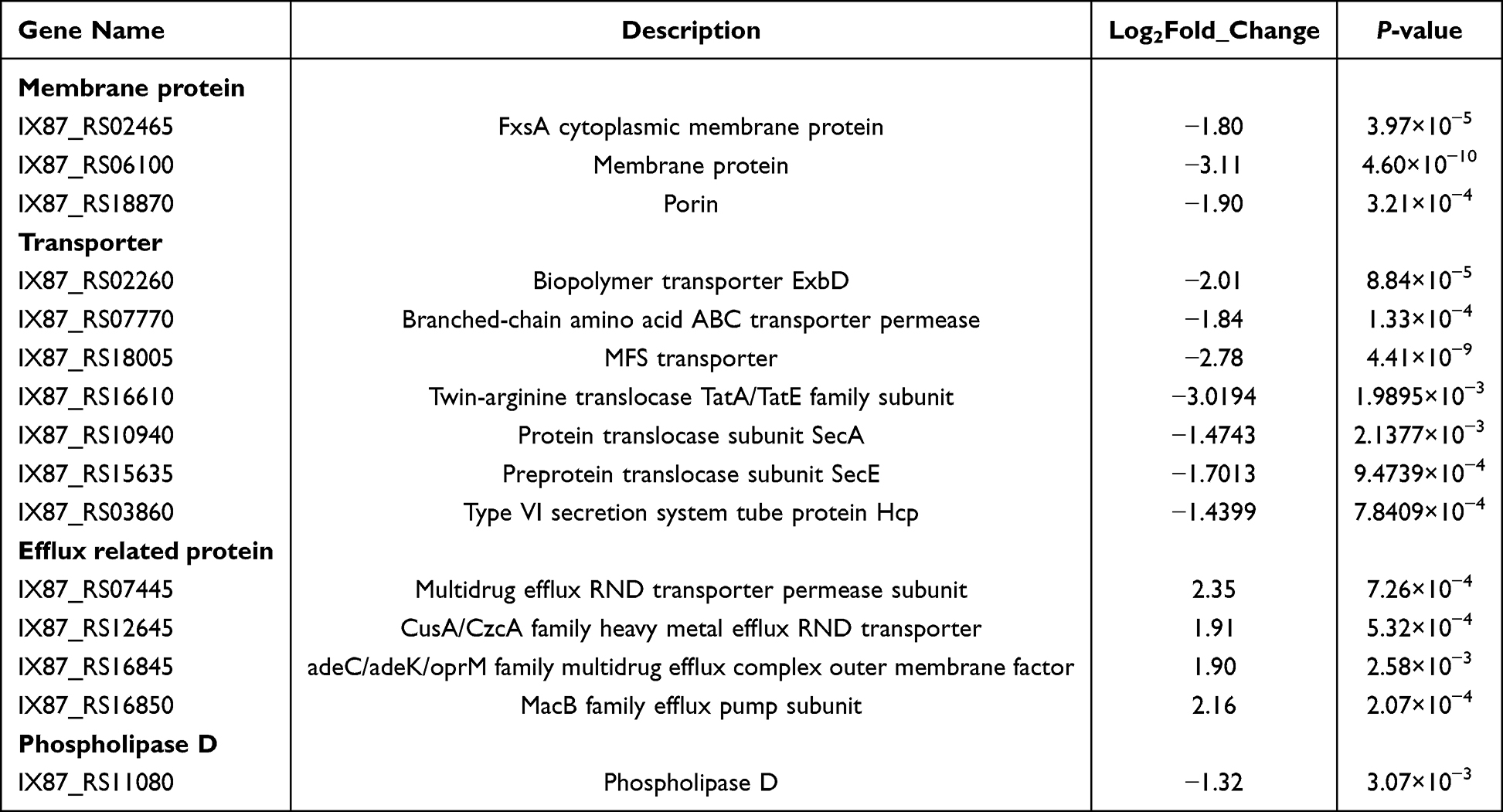 |
Table 2 Some Differential Genes Between C-7 and ATCC19606 |
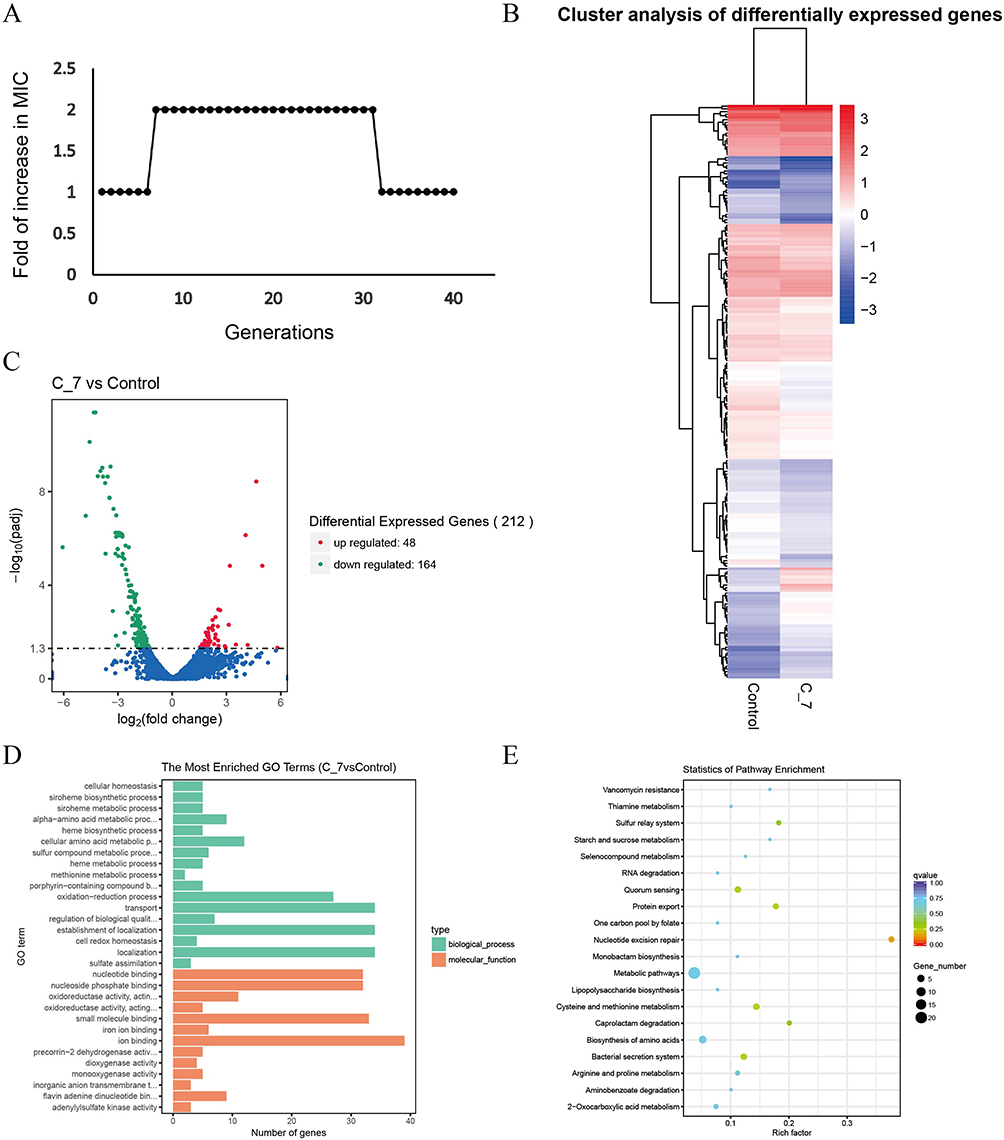 |
Figure 3 Transcriptome sequencing analysis of C-7 resistant strain and ATCC19606. (A) The passaged MIC of ATCC19606 under Cec4 induction. X axis is the number of passages, and Y axis is the corresponding MIC value. (B) The expression pattern of differential genes in C-7 (7th generation induced strain) and ATCC19606. Red is the up-regulated gene and blue is the down-regulated gene. (C) The DEGs volcano map of C-7 and ATCC19606. (D) The DEGs’ GO enrichment analysis of C-7 and ATCC19606. (E) The DEGs’ KEGG enrichment analysis of C-7 and ATCC19606. The size of bubble represents the number of genes enriched in this pathway, and the color of bubble represents the q-value of enrichment. |
Six pathways were significantly enriched in the KEGG database (P < 0.05), in which, three genes were enriched in the protein exocytosis pathway, including the double arginine-targeted protein translocase TatA/E family gene, the preproprotein translocase SecA subunit and the preprotein translocase SecE subunit, and all them were significantly downregulated (Figure 3D and E). The quorum sensing pathway includes the preprotein translocase SecY subunit and the peptide/nickel transport-related ATP-binding protein genes, which also showed a down-regulation trend. These results suggest that Cec4 may be involved in the process of type II secretion system and may target the epialleles of type II secretion system. In addition, the Hcp1 family genes of bacterial type VI secretion system (T6SS) effector genes are also enriched in this pathway, and this gene is significantly down-regulated in C-7, suggesting that the virulence of C-7-resistant strains may be decreased.
The Cec4 with Low Toxicity in vitro and in vivo
To investigate the toxicity of Cec4, an in vitro cytotoxicity assay was performed on Cec4 using HeLa cells and HepG2 cells (Figure 4A). At a high concentration of Cec4 (1.024 mg/mL) treating for 24 h, the Cec4 did not inhibit HeLa cells, and the inhibition rate of HepG2 cells was (9.565 ± 1.265) %. This indicates that the effect on HeLa and HepG2 cells was minimal when the concentration of Cec4 was as high as 1.024 mg/mL. C. elegans was also used to determine the acute toxicity of Cec4 in vivo. The survival rate of nematodes in erythromycin and Cec4 at the same concentration (2048 μg/mL) was 8.8% and 88.9% (Figure 4B). Besides, mice were intraperitoneally injected with Cec4 (45mg/kg) and saline, and their liver and kidneys were compared to further evaluate the toxicity of Cec4 in vivo (Figure 5A). The kidney histological samples from both the control and Cec4 groups showed no significant damage. In the liver samples, the mice in the Cec4 group showed slight damage to liver tissue, including cytoplasmic laxity of hepatocytes and a small amount of focal infiltration of lymphocytes, but the above liver damage did not affect the survival rate and normal physiological activities of the mice (Figure 5B). Blood biochemical tests in mice were used to further confirm the acute toxicity of Cec4. After intraperitoneal injection of mice at the same concentration of Cec4 (45 mg/kg) for 24 h, lactate dehydrogenase (LDH), alanine aminotransferase (ALT), aspartate aminotransferase (AST), and albumin (ALB) levels were measured to assess liver function, and blood creatinine (Cr) was measured to assess renal function. The results indicated that there was no significant difference between serum LDH, ALT, ALB, AST, and Cr values in the experimental mice compared with the untreated group (P > 0.05) (Figure 5C).
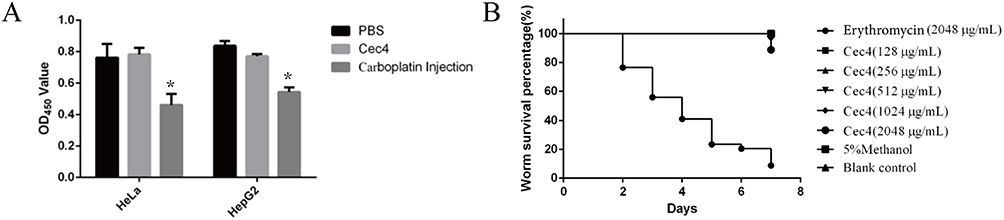 |
Figure 4 Acute toxicity of Cec4 to mammalian cells and C. elegans. (A) Effect of Cec4 on proliferation of HeLa cells and HepG2 cells. X axis shows different cell lines, Y axis indicates CCK-8 colorimetric results. (B) Effects of Cec4 and erythromycin on survival rate of C. elegans. The X-axis is observation days after treatment with erythromycin and Cec4, and the Y-axis is survival rate. Compared with erythromycin, the survival rate of C. elegans treated with Cec4 was much higher. Results were presented as mean ± SD. *P < 0.05. |
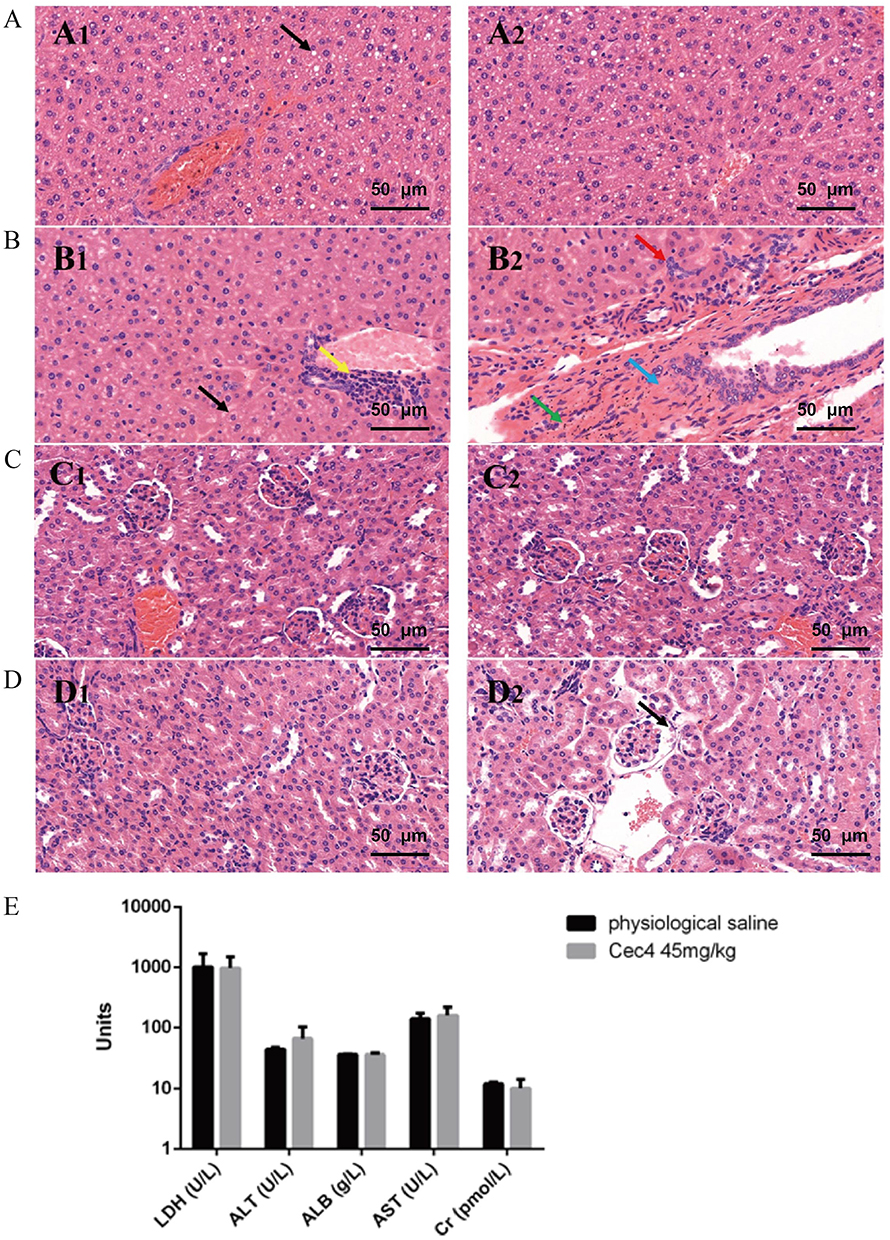 |
Figure 5 Acute toxicity assessment of Cec4 in mice. (A) Histopathological observation of liver tissues of mice injected with normal saline. Black arrows indicate steatosis of hepatocytes (A1 and A2 are different fields of view in the same slice, the same applies below). (B) Histopathological observation of liver tissue of mice injected with Cec4. Black arrows indicate loose hepatocyte cytoplasm, yellow arrows indicate focal infiltration of a few lymphocytes, red arrows indicate bile duct hyperplasia, blue arrows indicate connective tissue hyperplasia, and green arrows indicate hemosiderin exudation. (C) Histopathological observation of renal tissue of mice injected with normal saline. (D) Histopathological observation of renal tissue in mice injected with Cec4, the black arrow indicates eosinophilic material in the renal vesicles. (E) Blood biochemical assays of mouse for C9 (45 mg/kg) treated 24 h. Serum levels of lactate dehydrogenase (LDH), alanine aminotransferase (ALT), aspartate aminotransferase (AST), albumin (ALB), and serum creatinine (Cr) were determined. Data are presented as mean ± SD (n = 6) and the statistical analysis was assayed by the independent Student’s t test (P > 0.05). |
Cec4 is Effective in Treating A. baumannii Infections
To further evaluate the in vivo efficacy of Cec4, C. elegans infection model, skin and intraperitoneal infection model of BALB/c mice were used. C. elegans was infected with the AB4367992 (Supplementary Table S2) and treated with drugs. The results showed that the survival rate of the PB (2.5µg/mL) treatment group was 78.1% and that of Cec4 (16µg/mL) treatment group was 60.7% (Figure 6A). The bacterial load in the three groups of C. elegans was examined, and the results showed that the bacterial count in the nematodes was significantly lower compared with the control group (Figure 6B). Cec4 failed to completely eradicate bacteria from nematodes within 24 hours, possibly because small peptides are less permeable in nematodes than small-molecule compounds such as conventional antibiotics. The mouse skin infection assay was used to observe the therapeutic effect of Cec4 after infection of body surface burns or abrasions, and it was found that both Cec4 (16µg/mL) and PB (2.5µg/mL) treated mice had a significantly lower traumatic skin bacterial load than the untreated group, which indicates that the therapeutic effect of Cec4 in treating clinical body surface burns or abrasions with AB4367992 infection could be close to that of the clinical drug PB (Figure 6C and D). In the mouse model of peritonitis, the 72-h survival rate was 50% in the untreated group. The survival rate of the control group was 100%, and that of the PB group and Cec4 group was 70%. However, the final survival rate of the combined treatment group of Cec4 and PB was 80%, which was significantly higher than that of the treatment group alone (Figure 6E). The colony counts results showed that Cec4 significantly reduced the bacterial load in the liver, spleen, kidney, lung, and blood of the mice (P < 0.05) (Figure 6F). In conclusion, Cec4 was able to treat and reduce the in vivo infection of multidrug-resistant bacteria AB4367992.
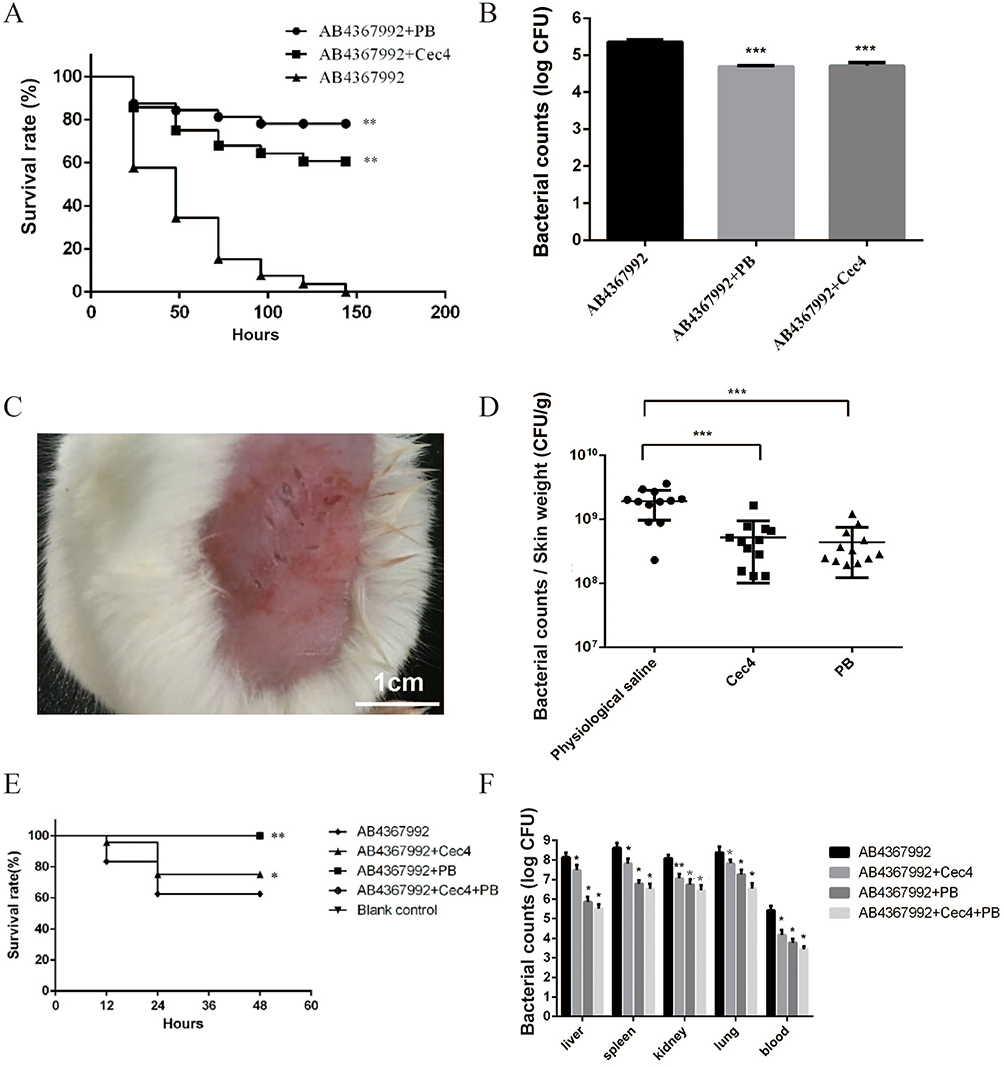 |
Figure 6 Cec4 is effective in the treatment of clinical multidrug-resistant A. baumannii infections. (A) Survival curves after Cec4 treatment of C. elegans infected with A. baumannii. (B) Bacterial load count of C. elegans infected with A. baumannii after Cec4 treatment. (C) Infection model of skin wound on mouse back. (D) Bacterial load count of A. baumannii after Cec4 treatment of mice with back skin infection (bacterial load per gram of skin). (E) Survival curves of mice infected with A. baumannii after Cec4 treatment. (F) Bacterial load count of mice infected with A. baumannii after Cec4 treatment. Results were presented as mean ± SD. *P < 0.05, **P < 0.01, ***P < 0.001. |
Discussion
Clinical treatment of CRAB mainly depends on tigecycline and polymyxin, etc, but the increase of drug resistance of tigecycline and cumulative nephrotoxicity of polymyxin, etc. According to the traditional antimicrobial mechanism of AMP, most AMP hold bactericidal effects by causing damage to the bacterial cell membrane or increasing the permeability of the cell membrane, leading to the leakage of bacterial contents.16,38 On the other hand, it has also been reported that some AMP, such as Tachyplesin III, can affect or inhibit the biosynthesis of unsaturated fatty acids by binding to DNA or entering microbial cells, leading to membrane instability.18 In the present study, genes associated with the efflux of macrolides were found to be upregulated in both transcriptome analysis of Cec4-treated A. baumannii. The MacB gene can form a three-way efflux pump with the MacA adaptor protein and TolC outer membrane exit tube in A. baumannii to expel antibiotics and export virulence factors.39 Previous studies have also shown that the deletion of the MacB gene reduces the resistance of Streptococcus pneumoniae to the human antimicrobial peptide LL-37.40 Macrolide antibiotics have been reported to inhibit bacterial protein synthesis by binding to the 50S large subunit of the ribosome and disrupt protein elongation by causing the dissociation of peptidyl-tRNA.41 However, bacteria can achieve resistance to macrolide antibiotics through ribosome modification42 and macrolide antibiotic efflux pump.43 The above findings show that the antimicrobial peptide Cec4 is also able to alter membrane potential and membrane fluidity to cause increased membrane permeability in bacterial cells, leading to the efflux of contents, which is consistent with the mainstream view. Therefore, it is reasonable to speculate that the bactericidal principle of Cec4 may have a similarity with macrolide antibiotics and that MacB may play a role in this process for Cec4 efflux (Figure 7).
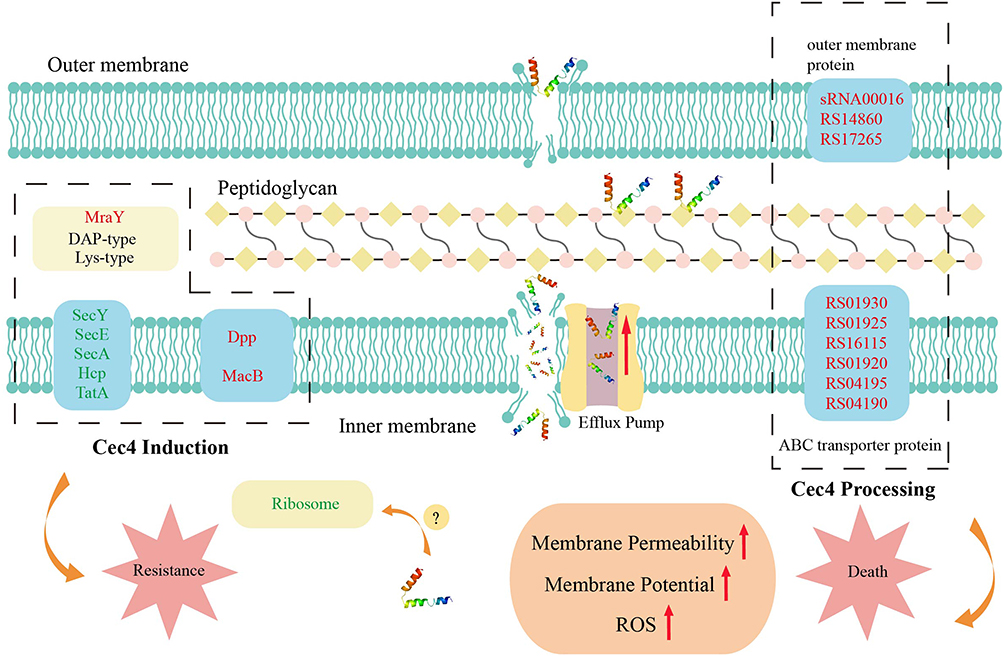 |
Figure 7 Mechanism of the death and resistance of A. baumannii after Cec4 treatment. Red font indicates up-regulated genes, green font indicates down-regulated genes, and red arrow indicates enhanced effects. The dashed boxes on the left are the differentially expressed genes of induced strain C-7 and ATCC19606. The dashed box on the right shows the differentially related genes after Cec4 treatment of ATCC19606. |
Previous studies have reported that A. baumannii will develop resistance to melittin-like AMP.44 Long-term Cec4 treatment resulted in stable downregulation of key genes in the Sec secretion system, suggesting that Cec4 may interact with proteins secreted by the Sec secretion pathway and exert negative effects on bacterial cells (Figure 7). SecA/E/Y gene in the general secretion pathway and TatA gene in the double arginine translocation Tat transporter both showed significant down-regulation. The transporter proteins were for unfolded and folded proteins, both of which are downregulated to affect protein efflux and have an impact on the type II secretion system.45 Dpp, a peptide/positive free transporter protein,46 was upregulated together with the macrolide antibiotic efflux pump MacB gene, suggesting that Cec4 may be able to enter the bacterial cell interior and cause bacterial cellular efflux in A. baumannii during the fight against Cec4. The significant upregulation of MraY, an upstream gene associated with lysine peptidoglycan, DAP-type peptidoglycan synthesis, suggests that Cec4 treatment may cause peptidoglycan damage or indirect damage, which in turn causes upregulation of the corresponding genes in bacterial cell regulation. The increased resistance of C-7 to Cec4 may be the result of reduced efflux of the corresponding target protein of Cec4 or upregulation of the peptide transporter protein gene.47 In addition, increased expression of efflux pump proteins associated with antibiotic resistance may further restrict Cec4 entry into bacterial cells, leading to further bacterial resistance.
In recent studies on AMPs, some peptides with good antibacterial effects, but slight haemolytic or toxic effects are common.48 In the in vitro toxicity test of our experiment, Cec4 (1024 μg/mL) showed little damage to HeLa and HepG2 cells. In addition, the higher concentration of Cec4 in mice may cause a certain burden on liver tissue, but blood biochemical analysis showed that it did not affect liver and kidney function. Therefore, Cec4 derived from the housefly has no haemolytic activity and eukaryotic toxicity at its effective concentration compared with melittins and scorpion venom derivatives. In addition, many AMPs have been shown to treat skin abrasions or burns infected with A. baumannii to a certain extent.49 In this study, Cec4 (16µg/mL) had a good therapeutic effect and could effectively reduce the number of bacteria in the infected area. For the in vivo treatment, Cec4 (16µg/mL) can improve the survival rate of mice and reduce the number of bacteria in the organs of the constructed sepsis model. However, the drug metabolism and distribution of Cec4 remain unclear. Further pharmacokinetic studies will help to clarify the metabolic process and distribution of the Cec4 in mice. Due to the poor stability of peptides in serum or blood, the computer design of Cec4-derived peptides can be further improved to solve these problems.13 For example, the modification of cathelicidin-BF-15 to peptide ZY4 successfully reduced the haemolytic activity of the original peptide, prolonged its half-life in serum, and increased its bactericidal capacity.50 Furthermore, some AMP can be used in combination with antibiotics to improve sterilisation.51 In the present study, we tried to combine Cec4 with the antibiotic polymyxin B, and investigated whether both have synergistic or additive effects in the treatment process in vivo.
In conclusion, Cec4 peptide can increase membrane permeability, cell membrane polarity, and intracellular reactive oxygen species in A. baumannii. The interaction of Cec4 with ribosomal subunits can cause bacteria to activate the efflux pathway to macrolide antibiotics, but the Cec4 is not easy to make bacteria resistant. Most importantly, Cec4 can effectively eliminate clinical drug-resistant bacteria and has a certain therapeutic effect on skin infections and sepsis infections, which possess the potential to treat clinical infection of multidrug-resistant A. baumannii.
Abbreviations
AMP, antimicrobial peptides; AST, Aspartate aminotransferase; Cec4, cecropin-4; CFU, colony-forming units; Cr, creatinine; CRAB, carbapenem-resistant A. baumannii; DAPI, 4’,6-diamidino-2-phenylindole; DiI, 1,1’-dioctadecyl-3,3,3’,3’-tetramethylindole carbocyanine perchlorate; FITC, fluorescein isothiocyanate; LDH, lactate dehydrogenase; MHB, Mueller–Hinton Broth; MIC, minimum inhibitory concentration; PB, polymyxin B; ROS, reactive oxygen species; TSB, Tryptic soy broth.
Data Sharing Statement
The sequencing data in the article have been deposited at the National Center for Biotechnology Information under BioProject PRJNA867035 and PRJNA867115 (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA867035, https://www.ncbi.nlm.nih.gov/bioproject/PRJNA867115). The data used to support the findings of this study are available from the corresponding author upon request.
Ethics Approval and Consent to Participate
All materials used in this study were approved for use by the Institutional Review Board, and all methods/experiments were conducted in accordance with the guidelines approved by the Ethics Committee of Guizhou Medical University, China (No.1603051).
Consent for Publication
All authors have approved the manuscript and given their consent for submission and publication.
Acknowledgment
We would like to thank Professor Yu-Cong Xie for the gift of Uracil-deficient E. coli OP50 and N2 wild-type C. elegans. We would also like to thank Emma Taylor for proofreading the paper (http://www.proof-reading-service.com).
Funding
This research is funded by the Changjiang scholars program of the Ministry of Education (TG2019110), the National Natural Science Foundation of China (No. 82002180, 81660347), the Guizhou Provincial Natural Science Foundation (ZK[2022] Key Program 039), China Postdoctoral Science Foundation (2022MD723770), National Foundation Cultivation Project of Guizhou Medical University(21NSFCP39), the Postdoctoral Foundation of Affiliated Hospital of Guizhou Medical University (BSH-Q-2021-07), the Science and Technology Fund Project of Guizhou Provincial Health Commission (gzwkj2023-567) and the High-Level Talent Initiation Project of Guizhou Medical University (J[2022] 023). Funders had no role in study design, data collection or analysis, preparation of the manuscript or the decision to publish it.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Perez S, Innes GK, Walters MS, et al. Increase in hospital-acquired carbapenem-resistant Acinetobacter baumannii infection and colonization in an acute care hospital during a surge in COVID-19 admissions — New Jersey, February–July 2020. MMWR Morb Mortal Wkly Rep. 2020;69(48):1827–1831. doi:10.15585/mmwr.mm6948e1
2. Choi J, Jang A, Yoon YK, et al. Development of novel peptides for the antimicrobial combination therapy against carbapenem-resistant Acinetobacter baumannii infection. Pharmaceutics. 2021;13(11):11. doi:10.3390/pharmaceutics13111800
3. van Duin D, Kaye KS, Neuner EA, et al. Carbapenem-resistant Enterobacteriaceae: a review of treatment and outcomes. Diagn Microbiol Infect Dis. 2013;75(2):115–120. doi:10.1016/j.diagmicrobio.2012.11.009
4. Sleiman A, Fayad AGA, Banna H, et al. Prevalence and molecular epidemiology of carbapenem-resistant Gram-negative bacilli and their resistance determinants in the Eastern Mediterranean Region over the last decade. J Glob Antimicrob Resist. 2021;25:209–221. doi:10.1016/j.jgar.2021.02.033
5. Bonomo RA, Burd EM, Conly J, et al. Carbapenemase-producing organisms: a global scourge. Clin Infect Dis. 2018;66(8):1290–1297. doi:10.1093/cid/cix893
6. Poirel L, Jayol A, Nordmann P. Polymyxins: antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin Microbiol Rev. 2017;30(2):557–596. doi:10.1128/CMR.00064-16
7. Srisakul S, Wannigama DL, Higgins PG, et al. Overcoming addition of phosphoethanolamine to lipid A mediated colistin resistance in Acinetobacter baumannii clinical isolates with colistin-sulbactam combination therapy. Sci Rep. 2022;12(1):11390. doi:10.1038/s41598-022-15386-1
8. Neshani A, Zare H, Akbari Eidgahi MR, et al. Review of antimicrobial peptides with anti- Helicobacter pylori activity. Helicobacter. 2019;24(1):e12555. doi:10.1111/hel.12555
9. da Costa de Souza G, Roque-Borda CA, Pavan FR. Beta-lactam resistance and the effectiveness of antimicrobial peptides against KPC-producing bacteria. Drug Dev Res. 2022;83(7):1534–1554. doi:10.1002/ddr.21990
10. Roque-Borda CA, Bento da Silva P, Rodrigues MC, et al. Pharmaceutical nanotechnology: antimicrobial peptides as potential new drugs against WHO list of critical, high, and medium priority bacteria. Eur J Med Chem. 2022;241:114640. doi:10.1016/j.ejmech.2022.114640
11. Wang C, Zhao G, Wang S, et al. A simplified derivative of human defensin 5 with potent and efficient activity against multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother. 2018;62(2). doi:10.1128/AAC.01504-17
12. Vila-Farres X, Garcia de la Maria C, López-Rojas R, et al. In vitro activity of several antimicrobial peptides against colistin-susceptible and colistin-resistant Acinetobacter baumannii. Clin Microbiol Infect. 2012;18(4):383–387. doi:10.1111/j.1469-0691.2011.03581.x
13. Hazam PK, Cheng -C-C, Hsieh C-Y, et al. Development of bactericidal peptides against multidrug-resistant Acinetobacter baumannii with enhanced stability and low toxicity. Int J Mol Sci. 2022;23(4):2191. doi:10.3390/ijms23042191
14. Park HJ, Kang HK, Park E, et al. Bactericidal activities and action mechanism of the novel antimicrobial peptide Hylin a1 and its analog peptides against Acinetobacter baumannii infection. Eur J Pharm Sci. 2022;175:106205. doi:10.1016/j.ejps.2022.106205
15. Jariyarattanarach P, Klubthawee N, Wongchai M, et al. Novel D-form of hybrid peptide (D-AP19) rapidly kills Acinetobacter baumannii while tolerating proteolytic enzymes. Sci Rep. 2022;12(1):15852. doi:10.1038/s41598-022-20236-1
16. Neshani A, Sedighian H, Mirhosseini SA, et al. Antimicrobial peptides as a promising treatment option against Acinetobacter baumannii infections. Microb Pathog. 2020;146:104238. doi:10.1016/j.micpath.2020.104238
17. Mwangi J, Yin Y, Wang G, et al. The antimicrobial peptide ZY4 combats multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii infection. Proc Natl Acad Sci U S A. 2019;116(52):26516–26522. doi:10.1073/pnas.1909585117
18. Liu C, Qi J, Shan B, et al. Tachyplesin causes membrane instability that kills multidrug-resistant bacteria by inhibiting the 3-ketoacyl carrier protein reductase FabG. Front Microbiol. 2018;9:825. doi:10.3389/fmicb.2018.00825
19. Caraway HE, et al. Antimicrobial random peptide mixtures eradicate Acinetobacter baumannii biofilms and inhibit mouse models of infection. Antibiotics. 2022;11:3. doi:10.3390/antibiotics11030413
20. Peng J, Long H, Liu W, et al. Antibacterial mechanism of peptide Cec4 against Acinetobacter baumannii. Infect Drug Resist. 2019;12:2417–2428. doi:10.2147/IDR.S214057
21. Peng J, Wu Z, Liu W, et al. Antimicrobial functional divergence of the cecropin antibacterial peptide gene family in Musca domestica. Parasit Vectors. 2019;12(1):537. doi:10.1186/s13071-019-3793-0
22. Liu W, Wu Z, Mao C, et al. Antimicrobial peptide Cec4 eradicates the bacteria of clinical carbapenem-resistant Acinetobacter baumannii biofilm. Front Microbiol. 2020;11:1532. doi:10.3389/fmicb.2020.01532
23. Wiegand I, Hilpert K, Hancock RE. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat Protoc. 2008;3(2):163–175. doi:10.1038/nprot.2007.521
24. Barman S, Konai MM, Samaddar S, et al. Amino acid conjugated polymers: antibacterial agents effective against drug-resistant Acinetobacter baumannii with no detectable resistance. ACS Appl Mater Interfaces. 2019;11(37):33559–33572. doi:10.1021/acsami.9b09016
25. Liu Y, Jia Y, Yang K, et al. Metformin restores tetracyclines susceptibility against multidrug resistant bacteria. Adv Sci. 2020;7(12):1902227. doi:10.1002/advs.201902227
26. Shi J, Chen C, Wang D, et al. Amphipathic peptide antibiotics with potent activity against multidrug-resistant pathogens. Pharmaceutics. 2021;13(4):438. doi:10.3390/pharmaceutics13040438
27. Nagarajan D, Roy N, Kulkarni O, et al. Ω76: a designed antimicrobial peptide to combat carbapenem- and tigecycline-resistant Acinetobacter baumannii. Sci Adv. 2019;5(7):eaax1946. doi:10.1126/sciadv.aax1946
28. Mao C, Wang Y, Yang Y, et al. Cec4-derived peptide inhibits planktonic and biofilm-associated methicillin resistant Staphylococcus epidermidis. Microbiol Spectr. 2022;10(6):e0240922. doi:10.1128/spectrum.02409-22
29. Alkahtani J, Soliman Elshikh M, Almaary KS, et al. Anti-bacterial, anti-scavenging and cytotoxic activity of garden cress polysaccharides. Saudi J Biol Sci. 2020;27(11):2929–2935. doi:10.1016/j.sjbs.2020.08.014
30. Uccelletti D, Zanni E, Marcellini L, et al. Anti- Pseudomonas activity of frog skin antimicrobial peptides in a Caenorhabditis elegans infection model: a plausible mode of action in vitro and in vivo. Antimicrob Agents Chemother. 2010;54(9):3853–3860. doi:10.1128/aac.00154-10
31. Liu Y, Leung SSY, Guo Y, et al. The capsule depolymerase Dpo48 rescues Galleria mellonella and mice From Acinetobacter baumannii systemic infections. Front Microbiol. 2019;10:545. doi:10.3389/fmicb.2019.00545
32. Sahoo TK, Jena PK, Prajapati B, et al. In vivo assessment of immunogenicity and toxicity of the bacteriocin TSU4 in BALB/c mice. Probiotics Antimicrob Proteins. 2017;9(3):345–354. doi:10.1007/s12602-016-9249-3
33. Jayamani E, Rajamuthiah R, Larkins-Ford J, et al. Insect-derived cecropins display activity against Acinetobacter baumannii in a whole-animal high-throughput Caenorhabditis elegans model. Antimicrob Agents Chemother. 2015;59(3):1728–1737. doi:10.1128/aac.04198-14
34. Luca V, Olivi M, Di Grazia A, et al. Anti-Candida activity of 1-18 fragment of the frog skin peptide esculentin-1b: in vitro and in vivo studies in a Caenorhabditis elegans infection model. Cell Mol Life Sci. 2014;71(13):2535–2546. doi:10.1007/s00018-013-1500-4
35. Pastagia M, Euler C, Chahales P, et al. A novel chimeric lysin shows superiority to mupirocin for skin decolonization of methicillin-resistant and -sensitive Staphylococcus aureus strains. Antimicrob Agents Chemother. 2011;55(2):738–744. doi:10.1128/aac.00890-10
36. Thandar M, Lood R, Winer BY, et al. Novel engineered peptides of a phage lysin as effective antimicrobials against multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother. 2016;60(5):2671–2679. doi:10.1128/AAC.02972-15
37. Harris G, KuoLee R, Xu HH, et al. Mouse models of Acinetobacter baumannii infection. Curr Protoc Microbiol. 2017;46:6G 3 1–6G 3 23. doi:10.1002/cpmc.36
38. Mookherjee N, Anderson MA, Haagsman HP, et al. Antimicrobial host defence peptides: functions and clinical potential. Nat Rev Drug Discov. 2020;19(5):311–332. doi:10.1038/s41573-019-0058-8
39. Greene NP, Kaplan E, Crow A, et al. Antibiotic resistance mediated by the MacB ABC Transporter family: a structural and functional perspective. Front Microbiol. 2018;9:950. doi:10.3389/fmicb.2018.00950
40. Majchrzykiewicz JA, Kuipers OP, Bijlsma JJ. Generic and specific adaptive responses of Streptococcus pneumoniae to challenge with three distinct antimicrobial peptides, bacitracin, LL-37, and nisin. Antimicrob Agents Chemother. 2010;54(1):440–451. doi:10.1128/aac.00769-09
41. Schroeder MR, Stephens DS. Macrolide resistance in Streptococcus pneumoniae. Front Cell Infect Microbiol. 2016;6:98. doi:10.3389/fcimb.2016.00098
42. Gupta P, Sothiselvam S, Vázquez-Laslop N, et al. Deregulation of translation due to post-transcriptional modification of rRNA explains why erm genes are inducible. Nat Commun. 2013;4(1):1984. doi:10.1038/ncomms2984
43. Zhang Y, Tatsuno I, Okada R, et al. Predominant role of msr(D) over mef(A) in macrolide resistance in Streptococcus pyogenes. Microbiology. 2016;162(1):46–52. doi:10.1099/mic.0.000206
44. Karyne R, Curty Lechuga G, Almeida Souza AL, et al. Pan-drug resistant Acinetobacter baumannii, but not other strains, are resistant to the bee venom peptide mellitin. Antibiotics. 2020;9(4). doi:10.3390/antibiotics9040178
45. Palmer T, Stansfeld PJ. Targeting of proteins to the twin-arginine translocation pathway. Mol Microbiol. 2020;113(5):861–871. doi:10.1111/mmi.14461
46. Xu X, Chen J, Huang X, et al. The role of a dipeptide transporter in the virulence of human pathogen, Helicobacter pylori. Front Microbiol. 2021;12:633166. doi:10.3389/fmicb.2021.633166
47. Singkham-In U, Higgins PG, Wannigama DL, et al. Rescued chlorhexidine activity by resveratrol against carbapenem-resistant Acinetobacter baumannii via down-regulation of AdeB efflux pump. PLoS One. 2020;15(12):e0243082. doi:10.1371/journal.pone.0243082
48. Feng X, Sambanthamoorthy K, Palys T, et al. The human antimicrobial peptide LL-37 and its fragments possess both antimicrobial and antibiofilm activities against multidrug-resistant Acinetobacter baumannii. Peptides. 2013;49:131–137. doi:10.1016/j.peptides.2013.09.007
49. de Breij A, Riool M, Cordfunke RA, et al. The antimicrobial peptide SAAP-148 combats drug-resistant bacteria and biofilms. Sci Transl Med. 2018;10(423):423. doi:10.1126/scitranslmed.aan4044
50. Mwangi J, Yin Y, Wang G, et al. The antimicrobial peptide ZY4 combats multidrug-resistant Pseudomonas aeruginosa and Acinetobacter baumannii infection. Proc Natl Acad Sci U S A. 2019;116(52):26516–26522. doi:10.1073/pnas.1909585117
51. Jangra M, Raka V, Nandanwar H. In vitro evaluation of antimicrobial peptide Tridecaptin M in combination with other antibiotics against multidrug resistant Acinetobacter baumannii. Molecules. 2020;25(14):3255. doi:10.3390/molecules25143255
 © 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2023 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Thoughtful Discussion on Article “Antimicrobial Peptide Cec4 Eradicates Multidrug-Resistant Acinetobacter baumannii in vitro and in vivo” [Letter]
Hidayat S, Isnawati R, Panjaitan NSD
Drug Design, Development and Therapy 2023, 17:1433-1434
Published Date: 15 May 2023