")
Back to Journals » ImmunoTargets and Therapy » Volume 10
Anti-IL-17 Agents in the Treatment of Axial Spondyloarthritis
Authors Atzeni F, Carriero A , Boccassini L, D'Angelo S
Received 26 February 2021
Accepted for publication 15 April 2021
Published 3 May 2021 Volume 2021:10 Pages 141—153
DOI https://doi.org/10.2147/ITT.S259126
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Shurin
Fabiola Atzeni,1 Antonio Carriero,2,3 Laura Boccassini,4 Salvatore D’Angelo2
1Rheumatology Unit, Department of Clinical and Experimental Medicine, University of Messina, Messina, Italy; 2Rheumatology Institute of Lucania (IReL), Rheumatology Department of Lucania, San Carlo Hospital of Potenza and Madonna delle Grazie Hospital of Matera, Potenza, Italy; 3Translational and Clinical Medicine, Department of Medicine and Health Sciences, University of Molise, Campobasso, Italy; 4Rheumatology Unit, Internal Medicine Department, ASST Fatebenefratelli-Sacco, University School of Medicine, Milan, Italy
Correspondence: Fabiola Atzeni
Rheumatology Unit, Department of Clinical and Experimental Medicine, University of Messina, Via C. Valeria 1, Messina, 98100, Italy
Tel +39 0902009
Fax +39 0902000
Email [email protected]
Abstract: Axial spondyloarthritis (axSpA) describes a group of chronic inflammatory rheumatic diseases primarily involving the axial skeleton. IL-17 is involved in the pathogenesis of numerous inflammatory diseases, including inflammatory arthritis. Until a few years ago, the only biological agents licensed for the treatment of axSpA and nr-axSpA were TNF inhibitors. However, as some patients did not respond to TNF inhibition or experienced secondary failure, the introduction of the first two IL-17 inhibitors (secukinumab [SEC] and ixekizumab [IXE]) has extended the treatment options, and there are now three others (bimekizumab, brodalumab and netakimab) in various stages of clinical development. The last ten years have seen the development of a number of therapeutic recommendations that aimed at improving the management of axSpA patients. The aim of this narrative review of the published literature concerning the role of IL-17 in the pathogenesis of SpA, and the role of IL-17 inhibitors in the treatment of axSpA, is to provide a comprehensive picture of the clinical efficacy and safety of the drugs themselves, and the treatment strategies recommended in the international guidelines.
Keywords: spondyloarthritis, ankylosing spondylitis, non radiographic axial spondyloarthritis, axial spondyloarthritis, anti-TNF drugs, anti-IL17 drugs, interleukin 17
Introduction
The term “axial spondyloarthritis” (axSpA) describes a group of chronic inflammatory rheumatic diseases primarily involving the axial skeleton. These are divided into the two major subtypes of radiographic axSpA (rx-axSpA) or ankylosing spondylitis (AS), and non-radiographic axSpA (nr-axSpA) on the basis of the presence or absence of radiographically detected changes in the sacroiliac joints and/or spine. Both forms are characterised by active inflammation that causes pain, stiffness, and bone formation, and thus leads to severely limited spinal mobility and functional impairment, but they may also involve the entheses and peripheral joints, as well as the eyes, skin and bowel.1,2
Nr-axSpA mainly affects females and AS mainly affects males, but there may also be sex-related differences in disease presentation: females often have a high Bath Ankylosing Spondylitis Disease Activity Index (BASDAI) and high Patient Global Assessment (PGA) scores, and more frequently experience fatigue and peripheral involvement, whereas males often have high C-reactive protein (CRP) levels, are positive for HLA B27, and show magnetic resonance imaging (MRI) evidence of inflammation.3–7
The main aims of axSpA treatment are to decrease inflammation and prevent or slow structural spinal damage in order to reduce pain and stiffness, and preserve spinal mobility, and this has led to the introduction of targeted biological agents against TNF and (more recently) IL-17, which are highly effective in reducing disease signs and symptoms, improving physical function, and increasing the quality of life.
The aim of this narrative review of the published literature concerning the role of IL-17 in the pathogenesis of SpA, and the role of IL-17 inhibitors in the treatment of axSpA, is to provide a comprehensive picture of the clinical efficacy and safety of the drugs themselves, and the treatment strategies recommended in the international guidelines.
The Role of IL-17
IL-17 pro-inflammatory cytokines are produced by CD8+ T cells, γδ T cells, natural killer (NK) T cells, mucosal associated invariant (MAI) T cells, and other cells involved in immune processes.8 The most widely studied is IL-17A, the expression of which is regulated by other inflammatory cytokines, including IL-23 which, together with IL-17 in the so-called IL-23/IL-17 axis, makes a crucial contribution to host protection and inflammation.9,10
IL-17 is involved in the pathogenesis of numerous inflammatory diseases, including inflammatory arthritis,10,11 and its effects on specific cells (eg fibroblasts, epithelial cells and synoviocytes) lead to the transcription of pro-inflammatory genes and the subsequent secretion of pro-inflammatory cytokines (TNF, IL-1, IL-6) and chemokines (CCL20, CCL2, CCL7, CXCL1, CXCL2, CXCL5, CXCL8).12–15 It also increases the production of granulomatosis-stimulating factors (G-CSF, GM-CSF),16 and regulates the production of antimicrobial peptides (defensins and S100 proteins).17 However, despite its protective function, excessive activation of the IL-17 pathway can lead to autoimmune responses or inflammatory diseases:18 it can guide the degradation of the joint extracellular matrix in patients with inflammatory arthritis;19,20 cause osteoclast activation and bone destruction;21 and also promote the angiogenesis that allows inflamed joints to be reached by inflammatory cells.22,23
IL-17F plays a similar role to that of IL-17A, but induces a weaker inflammatory response.24 It increases the production of certain pro-inflammatory mediators and matrix metalloproteinases in fibroblasts and epithelial cells,25,26 and is involved in the neutrophilia associated with severe asthma.27
IL-17A and IL-17F therefore both have pro-inflammatory, osteoclastogenic and angiogenic potential, thus making them critical drivers of inflammation.18
The IL-23/IL-17 axis plays a clear role in the pathogenesis of SpA.18 Taams et al have reviewed its role in animal models of inflammatory arthritis, discussed the presence of IL-17 family members in the blood and tissue of SpA patients, and investigated the genetic variants involving the axis that leading to susceptibility to SpA.18 They have also explored the synergistic effect of IL-17 and other pro-inflammatory cytokines on joint inflammation and altered bone homeostasis.18
A number of animal models of inflammatory arthritides, including rheumatoid arthritis (RA) and SpA, have demonstrated the key pathogenetic role of the IL-17 and IL-23 pathways.18 It has been shown that the inhibition of IL-17A decreases disease activity and joint damage in an adjuvant-induced model of arthritis.28,29 Moreover, IL-17 or IL-17 receptor knockout in an animal model of collagen-induced arthritis (CIA) prevents the development of the disease, which depends on IL-17 in its early phases, is partially suppressed by IL-17 receptor inhibition during the phase of active inflammation, and is exacerbated in terms of severity and joint destruction when IL-17 is overexpressed.30–33
One comprehensive review of these animal models34 has reported that the overexpression of HLA-B27 in rats leads to the proliferation of IL-17+ CD4+T cells.35,36 It has also been shown that the IL-17 produced by IL-23 stimulates arthritis within an SpA syndrome in the SKG mouse model of RA,37–39 and that the introduction of exogenous IL-23 stimulates the production of IL-17 and exacerbates enthesitis in the CIA model.40,41 Furthermore, IL-17 plays a key role in the development of ankylosis and psoriasis-like dermatitis in certain mouse strains.42,43
IL-17A and IL-17F Cytokines in SpA Patients
IL-17A and IL-17F have been investigated in fewer studies of SpA than studies of RA, but the results have shown that their serum levels are significantly higher in SpA patients than in healthy subjects.43,44 Moreover, serum IL-17 levels in AS patients correlate positively with disease activity.45,46 It has also been found that IL-17 levels are higher in the synovial fluid than in the serum of psoriatic arthritis (PsA) patients,47 and higher in the synovial fluid of patients with reactive arthritis or undifferentiated SpA than in those with osteoarthritis (OA) or RA.48 Furthermore, the expression of IL-17 receptor A is higher in the synoviocytes of PsA and RA patients than in the synoviocytes of patients with OA.49
One study has found that the gut (terminal ileum) of AS patients is a significant source of IL-23 but not IL-17.50
The only data available concerning IL-17F in SpA patients show that it is more frequently present and more expressed in the synovial tissue of patients with PsA than in OA patients.51,52
The synergistic effects of IL-17A and other cytokines and mediators lead to an increased pro-inflammatory response and, although these effects have been explored in only a few studies of SpA, they have been widely studied in RA patients.18
The most widely studied is the synergy of IL-17A and TNF.53–55
However, the synergistic role of IL-17A and TNF in bone formation and destruction still needs further and more detailed clarification.18
IL-17A can also have synergistic effects with other pro-inflammatory cytokines:18 together with IL-1β, it increases the production of IL-6 by RA synoviocytes56 and the production of CCL20 by fibroblast-like synoviocytes57 and, together with IFNα, it enhances the expression of intercellular adhesion molecule 1 (ICAM1), thus increasing inflammation in PsA patients.58
The lack of robust evidence means that the role of IL-17F in the pathogenesis of SpA still needs to be definitely confirmed,18,59 but it is known that it acts synergistically with IL-17A in the process of inflammation52,59 and it has been shown that dual IL-17A and IL-17F blockade reduces inflammation better than IL-17A blockade alone.59
The mechanisms allowing the synergistic effects of IL-17A and IL-17F are not clear, but it has been hypothesised that IL-17A may stabilise mRNA transcripts, thus increasing gene expression and protein production,60 and that phospholipase D enzymes may up-regulate cytokine secretion.61 For example, although there is no evidence concerning IL-17F,18 it has been reported that IL-8 mRNA and other mRNA transcripts (including ACT1, MIP2 and CSF2) are involved in the synergy between TNF and IL-17A.15,18
Among all of the immunity cell types capable of producing IL-17, it has been shown that a number of T cell subsets produce IL-17 in patients with SpA.11,62–64
- IL-17+ CD8+ T cells have been identified as potential sources of IL-17 in studies of immune-mediated inflammatory diseases, including psoriasis, multiple sclerosis and PsA (eg in psoriatic skin lesions).58,62,65–70 These T cells can be found in the peripheral blood of patients with AS, and their frequency increases with the severity of the disease;71 furthermore, they are more frequent in the synovial fluid than in the peripheral blood of both PsA and AS patients.65,72 Their immunophenotype shows pro-inflammatory capacity,73 and their presence is associated with markers of disease activity such as CRP levels and Doppler findings.65 However, increased levels of IL-17-producing CD8+ T cells have not been found in RA patients,65 and seem to be related to the development of only HLA class I and not HLA class II-associated SpA.18
- Tissue-resident memory (TRM) T cells. CD8+ TRM cells express IL-17 and other cytokines in the skin of healthy and psoriatic subjects69,74–76 and, although less widely studied, CD4+ TRM cells have also been found in human skin.77 It has been shown that IL-17-producing TRM cells can be considered disease drivers in a mouse model of SpA,78 and they may be present in the synovial fluid of patients with juvenile inflammatory arthritis (JIA)79 or SpA,73 which underlines their importance in the pathogenesis of inflammatory arthritides.18
- MAIT cells. CD8+ MAIT cells (particularly Vα7.2/IL-17+CD8+ T cells) have been identified in the skin and blood of psoriatic patients.80,81 They are also more frequently detected in the peripheral blood of AS patients than in the peripheral blood of healthy subjects.82,83
- Invariant NKT cells. Very few studies have described the presence of IL-17-producing NKT cells in SpA patients, but the findings of a murine study suggest that NKT cells may maintain or activate TH17 cells, and thus contribute to the development of inflammation.84
- γδ T cells producing IL-17 have been very frequently detected in the peripheral blood of patients with PsA, AS, reactive arthritis, and enthesitis-related JIA,78,85–87 as well as in the synovial fluid of patients with PsA, reactive arthritis, and undifferentiated SpA.87,88 It has also been shown that γδ T cells produce IL-17 in mouse models of SpA and psoriatic skin inflammation,89–91 and accumulate in tissues often affected by SpA, such as the entheses, the aortic root, and the eye.92
Other Cells
IL-17 can also be produced by immunity cells other than T cells, including group 3 innate lymphoid cells (ILC3s), which have been described in the peripheral blood and synovial fluid of PsA patients, where their levels correlate with disease activity.93,94 High levels of CD3− CD56+ NK cells have been detected in patients with enthesitis-related arthritis,86 and IL-17-producing NK cells have been found in the synovial fluid of patients with reactive arthritis or undifferentiated SpA.88
Finally, it has been found that mast cells, which were initially hypothesized to be IL-17 producing cells, actually have the function of storing and releasing exogenous IL-17A.95,96
IL-17 Inhibition in axSpA Patients: Randomised Controlled Trials and Real-Life Data
Until a few years ago, the only biological agents licensed for the treatment of AS and nr-axSpA were TNF inhibitors. However, as some patients did not respond to TNF inhibition or experienced secondary failure, the introduction of the first two IL-17 inhibitors (secukinumab [SEC] and ixekizumab [IXE]) has extended the treatment options, and there are now three others (bimekizumab, brodalumab and netakimab) in various stages of clinical development.97
The last ten years have seen the development of a number of therapeutic recommendations aimed at improving the management of axSpA patients. The guidelines of the Assessment of Spondyloarthritis International Society/European League Against Rheumatism (ASAS/EULAR)98 and those of the American College of Rheumatology/Spondylitis Association of America/Spondyloarthritis Research and Treatment Network (ACR/SAA/SPARTAN)99 recommend the use of non-steroidal anti-inflammatory drugs (NSAIDs) and physical therapy in the first-line treatment of pain and stiffness.
As for conventional disease-modifying anti-rheumatic drugs (DMARDs), only sulfasalazine has demonstrated a mild efficacy in treating the axial manifestations of SpA but, as well as methotrexate and leflunomide, it may play a role in the treatment of peripheral SpA solely.
In the case of non-responders to NSAIDs or patients for whom NSAIDs are contraindicated, biological drugs (TNF and IL-17 inhibitors) are strongly recommended. As there is no indication that any one of these is more effective than the others, the choice should be made on the basis of the available safety data, the presence of extra-articular manifestations, and patient preference. Since no predictive factors of good response to IL-17 inhibitors are currently available, treatment response is still far from ideal for many patients, and we lack the biomarkers to predict which medication is most appropriate for an individual patient. The ASAS/EULAR98 and ACR/SAA/SPARTAN99 guidelines recommend TNF inhibitors as first line of biologic agents, mainly because of a broader clinical experience and larger pharmacovigilance data with these agents as compared to IL-17 inhibitors. This approach may change in future when real life data and results of comparative and strategy studies on IL-17 inhibitors will be available.
Clinical trials are of course the gold standard for assessing the efficacy and safety of new biological drugs but, as they are conducted under standardized conditions and exclude certain types of patients and/or situations, their findings may not reflect their real-world prescription and use.
Secukinumab
The efficacy and safety of SEC, a fully human antibody that selectively targets IL-17A and inhibits its interaction with IL-17 receptors,100 has been tested in AS patients in one phase II101 and five Phase III trials (MEASURE 1,102 MEASURE 2,103 MEASURE 2-J,104 MEASURE3105 and MEASURE4106) and their extensions. In the Phase II proof-of-concept study, 59% of the patients in the SEC group showed a week-64 ASAS20 response as against 24% of the patients receiving placebo (PBO).
MEASURE 1 and 2,97 which respectively enrolled 371 and 219 patients, tested two doses of the active drug (75 and 150 mg) against PBO, and had the percentage of patients with a week-16 ASAS20 response as their primary endpoint (Table 1). At the end of the MEASURE 1 study, an ASAS20 response was achieved by 61% of the patients in the 75 mg group, 60% of those in the 150 mg group, and 28% of those in the PBO group; in MEASURE 2, the corresponding figures were respectively 41%, 61% and 28%.102,103 The drug was efficacious regardless of whether it was received by patients as their first biological drug or by patients failing to respond to previous anti-TNF treatment. About 80% of the patients did not experience radiographic progression over a period of two years,107 and the drug was well tolerated.108
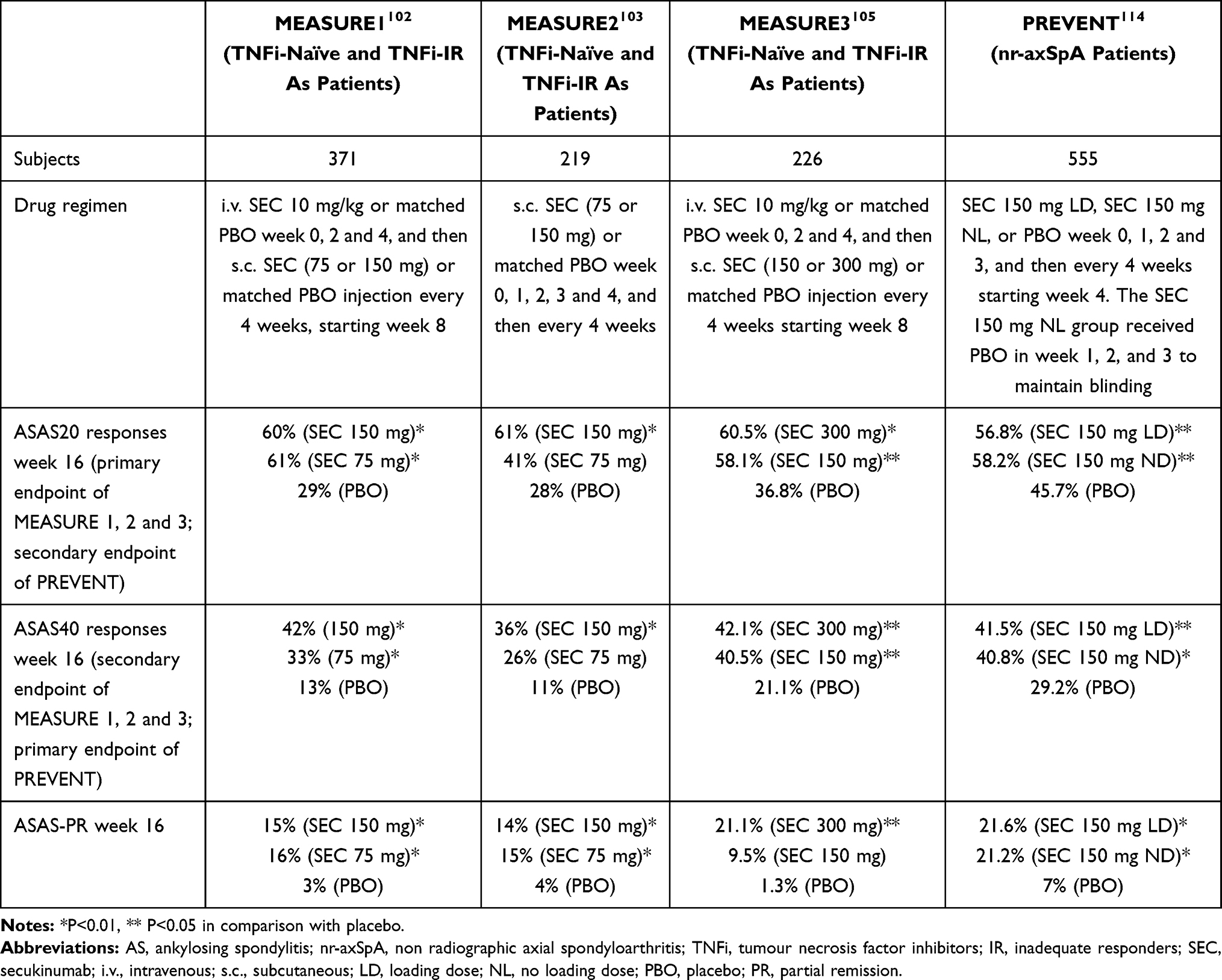 |
Table 1 Summary of Secukinumab Trials |
Sustained responses were observed in the 5-year, long-term extension of MEASURE 1, during which 74% of the patients in the 75 mg group and 79% of those in the 150 mg group achieved an ASAS20 response, and respectively 54% and 65% achieved an ASAS40 response.109,110 Among the 150 patients who completed five years’ treatment with SEC 150 mg in the extension of MEASURE 2,111 ASAS20 responses were recorded in 67% and ASAS40 responses in 50%, and there were sustained improvements in the other efficacy endpoints. The drug’s safety profile remained consistent with that described in previous reports.111
A pooled analysis of MEASURE 1 and 2 showed that the majority of the SEC-treated patients who achieved remission by week 16 remained in remission for up to three years.112
The use of SEC 150 mg in the MEASURE 2-J trial led to sustained improvement in the signs and symptoms of Japanese AS patients for 24 weeks without giving rise to any new or unexpected safety issues.104
In the MEASURE 3 trial, in comparison with 36.8% of the patients receiving PBO, a 16-week ASAS20 response (the primary endpoint) was observed in 60.5% of the patients treated with SEC 300 mg (p<0.01) and 58.1% of those treated with SEC 150 mg (p<0.05).105 There were improvements in the primary and secondary endpoints in both TNF inhibitor-naïve patients and patients inadequately responding to previous TNF inhibition.113 The 52-week ASAS20 and ASAS40 response rates in the SEC 150 mg group were respectively 54% and 41%.105
MEASURE 4 assessed the long-term efficacy, safety and tolerability of SEC. The initial 16-week treatment was completed by 97% of the patients, and the subsequent two-year treatment was completed by 83%.106 The primary endpoint of 16-week ASAS20 responses was not reached because of the very high response rate in the PBO group,109 but the drug’s two-year safety profile matched that described in previous studies.113
PREVENT was the first phase III trial of SEC in active nr-axSpA patients with objective signs of inflammation (MRI with SI joint inflammation and/or high-sensitivity CRP).114 The treatment improved disease signs and symptoms throughout the 52-week study period without leading to any new safety findings. The study met both of its primary endpoints: the 16-week ASAS40 response rate was higher among the patients receiving SEC 150 mg with loading doses than among those receiving PBO (41.5% vs 29.2%; p = 0.0197), and the 52-week ASAS40 response rate was also higher in the patients receiving SEC 150 mg without loading doses (39.8% vs 19.9%; p <0.0021).114
A very recent a systematic review and meta-analysis of ten randomized controlled trials (RCTs) assessing the efficacy and safety of IL-17A inhibitors in a total of 2,613 patients with AS (six trials of SEC, two of IXE, and one each of netakimab and bimekizumab) showed that, in comparison with placebo, the IL-17A inhibitors improved both the ASAS20 response rate (OR = 2.58; p <0.01) and the ASAS40 response rate (OR = 2.80; p <0.01). Although the treatment significantly increased the risk of adverse events (OR = 1.23; p = 0.03) and nasopharyngitis (OR = 1.72; p <0.01), it did not increase the risk of serious adverse events (OR = 0.87; p = 0.60).115
In 2018, Gentileschi et al described the first real-life experience of the short-term efficacy of SEC in 21 axSpA patients enrolled at three Italian referral centers. Between the start of treatment and the 3-month follow-up visit, there was a statistically significant reduction in the BASDAI (p <0.0001) and ASDAS-CRP values (p = 0.0005), with no statistically significant difference between the subgroups receiving SEC 150 or 300 mg, and no significant difference between the biological treatment-naïve patients and the patients previously treated with TNF inhibitors. No adverse events were reported during the observation period.116
The same group also published the findings of a real-life study of the long-term effectiveness of SEC and drug survival in axSpA patients in 2020. There was a statistically significant reduction in the BASDAI and ASDAS-CRP values during the 24-month observation period, with no statistically significant difference between the two doses or between the biological treatment-naïve patients and the patients failing to respond to previous TNF inhibition. The global 24-month drug retention rate was 78.2%, and no adverse event or infectious disease was reported during the study period.117
In 2019, Mann et al published findings taken from the Czech ATTRA that showed rheumatologists considered SEC to be equivalent to a TNF inhibitor in the case of biological DMARD-naïve patients and that, after the failure of ≥3 TNF inhibitors, patients were significantly more likely to be treated with SEC.118
Elliot and Wright described their experience with SEC in a cohort of 45 patients: 36 with PsA (five of whom had predominant axial disease) and nine with AS. SEC proved to be effective in patients inadequately responding to TNF inhibition in their clinical setting.119
Williams et al described the results of their real-world experience of SEC treatment for AS at the Royal National Hospital for Rheumatic Diseases in Bath in 2020, thus providing further evidence that SEC is largely safe and effective.120
Also, in 2020, the Spondyloarthritis Roman Group (STRONG) published the results of a multicentre, prospective observational study121 showing that SEC improved all of the evaluated clinical parameters and patient-reported outcomes after six and 12 months. The treatment was well tolerated, and drug survival was good, particularly among male AS patients.121
Another real-life study of 1,860 axSpA patients in 13 European registries participating in the European Spondyloarthritis Research Collaboration Network was published in 2020.122 SEC retention rates after six and 12 months of treatment were respectively 82% and 72%, and comparable with those observed in studies of TNF inhibition. Response rates were lower than those recorded in the RCTs but consistently better among the biological drug-naïve patients.122
A recent systematic review and meta-analysis of real-life studies of the biological drugs used to treat AS has shown that one-year drug survival rate of SEC was 0.77 (95% confidence interval 0.64–0.90).123
A Canadian cost-effectiveness analysis has shown that SEC 150 mg is a more cost-effective option for biological treatment-naïve AS patients than certolizumab pegol, adalimumab, golimumab, etanercept and an etanercept biosimilar, or infliximab and an infliximab biosimilar,124 and the situation is similar in Finland125 and the UK126 for both biological treatment-naïve and biological treatment-experienced patients with active AS.
The ongoing, longitudinal SERENA study of patients with psoriasis, PsA or AS with an observational period of up to five years is being conducted at 438 centers across Europe and, when its results become available, it will provide valuable information concerning the long-term, real-world effectiveness and safety of SEC.127
Two recent real life cohort studies compared effectiveness of treatment with SEC with that of TNF inhibitors. Both studies showed that axSpA patients with prior TNFi exposure treated with SEC experienced comparable outcomes as patients treated with an alternative TNF inhibitor.128,129
Ixekizumab
The efficacy of IXE, an IgG4 monoclonal antibody that has affinity for the homodimer IL-17A and the heterodimer IL-17A/F, in treating radiographic axSpA has been demonstrated in two phase III RCTs (COAST-V130 and COAST-W131), both of which achieved their primary endpoints and showed significant ASAS40 responses to its administration every two or every four weeks (Table 2).
 |
Table 2 Summary of Ixekizumab Trials |
In the COAST-V trial, which enrolled TNF inhibitor-naїve AS or r-axSpA patients, 16-week ASAS40 responses were observed in 51.8% of those receiving IXE every two weeks, in 48.1% of those receiving IXE every four weeks, and in 18.6% of those receiving PBO.130 The trial also included a fourth study arm of patients receiving adalimumab 40 mg every two weeks that served as an in-study active reference for comparison. Numerically, the ASAS40 response seen with IXE were similar to those observed in the adalimumab group. Week-52 ASAS40 responses were observed in 53.1% of the patients treated with IXE every 4 weeks and in 51% of the patients treated with IXE every 2 weeks.131–134
The COAST-W trial enrolled AS or r-axSpA patients inadequately responding to TNF inhibition. Week-16 and week-52 ASAS40 responses were observed in respectively 25.4 and 34.2% of the patients treated with IXE every four weeks, 30.6 and 30.6% of those treated with IXE every two weeks.131,134
No data are yet available concerning the ability of IXE to inhibit structural progression.
The efficacy and safety of IXE in patients with nr-axSpA was assessed in the COAST-X trial, a phase III RCT that enrolled TNF inhibitor-naïve patients135 (Table 2). IXE was superior to PBO after 16 and 52 weeks: 16-week ASAS40 responses were observed in 35% of the patients receiving IXE every four weeks, 40% of those receiving IXE every two weeks, and 19% of those receiving PBO; 52-week ASAS40 responses were observed in respectively 30%, 31%, and 13%. The adverse events were no different from those found in previous IXE studies, and no new safety issues were identified.136 IXE was approved for the treatment of nr-axSpA by the FDA and the EMA in June 2020.137
To the best of our knowledge, there are no published real-life observational studies of IXE treatment in axSpA patients.
Efficacy of Secukinumab and Ixekizumab on Extra-Axial Manifestations
In addition to axial involvement, axSpA patients may also suffer from peripheral arthritis, enthesitis and dactylitis as well as extra-articular manifestations such as psoriasis, uveitis and inflammatory bowel diseases (IBD). These outcomes have not been specifically assessed in the above mentioned SEC and IXE pivotal trials where only axial outcomes have been studied as primary endpoints.
Nevertheless, some information about peripheral arthritis, enthesitis, dactylitis and psoriasis may be drawn from the development programs of IXE and SEC in PsA showing significant improvements for all these features.113
Treatment emergent anterior uveitis and IBD were reported in long-term clinical trials (MEASURE and COAST programs) and post marketing safety data in psoriasis, PsA and AS with low overall incidences, within the expected ranges in these disorders.110,111,134,138 However, the available data, although limited, would encourage caution in the use of IL-17 inhibition in patients with a history of uveitis or IBD.
Based on the aforementioned data regarding uveitis, IBD and psoriasis, IL-17 inhibitors would not be the first choice in case of associated IBD or uveitis, whereas it would be appropriate in case of psoriasis.
Other IL-17 Inhibitors
Brodalumab (an IL-17A receptor antagonist that also inhibits IL-17F, the IL-17A/F heterodimer and IL-17E) is approved for the treatment of psoriasis.139 However, a phase III trial involving patients with PsA was interrupted because of concerns about major side effects including depression and suicidal behaviour140 (although it is still unclear whether there is a causal relationship between the drug and suicidal ideation and behaviour), and a PBO-controlled phase II trial involving axSpA patients (ClinicalTrials.gov ID NCT02429882) was cancelled and withdrawn in 2015.
The preliminary results of a phase III trial of brodalumab treatment in AS and nr-axSpA patients (ClinicalTrials.gov ID NCT02985983) carried out in Japan were presented at the 2019 EULAR conference.141 The week-16 ASAS40 response rate was significantly higher in the brodalumab group (35/80, 43.8%, p=0.018) than in the PBO group (19/79, 24.1%) and, on the basis of these results, brodalumab may be considered a future therapeutic option for patients with axSpA.
Bimekizumab, an inhibitor of both IL-17A and IL-17F, has been shown to be effective in a phase IIb trial involving AS patients:142 significantly more bimekizumab-treated patients achieved a week-12 ASAS40 response than those receiving PBO (non-responder imputation: 29.5% for bimekizumab 16 mg every four weeks; 42.6% for bimekizumab 64 mg every four weeks; 46.7% for bimekizumab 160 mg every four weeks vs 13.3% for PBO every four weeks; p<0.05).132 Phase II trials involving AS patients (ClinicalTrials.gov ID NCT03355573 and NCT03215277) and phase III trials involving patients with AS (ClinicalTrials.gov ID NCT03928743), nr-axSpA (ClinicalTrials.gov ID: NCT03928704), or AS and nr-axSpA (ClinicalTrials.gov ID NCT04436640) are currently ongoing.
Netakimab is a recombinant humanized IgG1 anti-IL-17 monoclonal antibody with a modified Fc fragment and CDR regions.143 A Phase III, PBO-controlled trial (ClinicalTrials.gov ID NCT03447704) is currently evaluating the safety and efficacy of a 120 mg dose for up to one year in 228 patients with active AS.
Conclusions
The IL-23/IL-17 axis plays a clear role in the pathogenesis of SpA. IL-17A and IL-17F have been investigated in few studies of SpA, but their serum levels are significantly higher in SpA patients and correlate positively with disease activity. TNF and (more recently) IL-17 inhibitors have dramatically changed the scenario of axSpA treatment as they are highly effective in reducing disease signs and symptoms, improving physical function, and increasing the quality of life. The results of clinical trials have shown that IL-17 inhibitors are efficacious and safe, and this has been confirmed by some recent real-world data concerning a range of unselected patients. The guidelines of the ASAS/EULAR and those of the American College of Rheumatology/Spondylitis Association of America/Spondyloarthritis Research and Treatment Network (ACR/SAA/SPARTAN) recommend in the case of non-responders to NSAIDs or patients for whom NSAIDs are contraindicated, biological drugs (TNF and IL-17 inhibitors). Until today, there is no indication that any one of these is more effective than the others, then the choice should be made on the basis of the available safety data, the presence of extra-articular manifestations, and patient preference. However, based on the published data IL-17 inhibitors would not be the first choice in case of associated IBD or uveitis.
Disclosure
Dr Salvatore D’Angelo reports personal fees from Abbvie, Eli Lilly, Novartis, and UCB, outside the submitted work. The authors reported no other potential conflicts of interest for this work.
References
1. Taurog JD, Chhabra A, Colbert RA. Ankylosing spondylitis and axial spondyloarthritis. N Engl J Med. 2016;375:1303. doi:10.1056/NEJMc1609622
2. Rudwaleit M, van der Heijde D, Landewé R, et al. The development of assessment of spondyloarthritis international society classification criteria for axial spondyloarthritis (part II): validation and final selection. Ann Rheum Dis. 2009;68:777–783. doi:10.1136/ard.2009.108233
3. Tournadre A, Pereira B, Lhoste A, et al. Differences between women and men with recent-onset axial spondyloarthritis: results from a prospective multicenter French cohort. Arthritis Care Res. 2013;65:1482–1489. doi:10.1002/acr.22001
4. Akkoc N, Khan MA. ASAS classification criteria for axial spondyloarthritis: time to modify. Clin Rheumatol. 2016;35:1415–1423. doi:10.1007/s10067-016-3261-6
5. Slobodin G, Eshed I. Non-radiographic axial spondyloarthritis. Isr Med Assoc J. 2015;17:770–776.
6. Wallman JK, Kapetanovic MC, Petersson IF, et al. Comparison of non-radiographic axial spondyloarthritis and ankylosing spondylitis patients–baseline characteristics, treatment adherence, and development of clinical variables during three years of anti-TNF therapy in clinical practice. Arthritis Res Ther. 2015;17:378. doi:10.1186/s13075-015-0897-6
7. Deodhar A, Mease PJ, Reveille JD, et al. Frequency of axial spondyloarthritis diagnosis among patients seen by US rheumatologists for evaluation of chronic back pain. Arthritis Rheumatol. 2016;68:1669–1676. doi:10.1002/art.39612
8. Rouvier E, Luciani MF, Mattéi MG, et al. CTLA-8, cloned from an activated T cell, bearing AU-rich messenger RNA instability sequences, and homologous to a herpesvirus saimiri gene. J Immunol. 1993;150:5445–5456.
9. Murphy CA, Langrish CL, Chen Y, et al. Divergent pro- and antiinflammatory roles for IL-23 and IL-12 in joint autoimmune inflammation. J Exp Med. 2003;198:1951–1957. doi:10.1084/jem.20030896
10. Lubberts E. The IL-23-IL-17 axis in inflammatory arthritis. Nat Rev Rheumatol. 2015;11:562. doi:10.1038/nrrheum.2015.128
11. Korn T, Bettelli E, Oukka M, et al. IL-17 and Th17 cells. Annu Rev Immunol. 2009;27:485–517. doi:10.1146/annurev.immunol.021908.132710
12. Yao Z, Painter SL, Fanslow WC, et al. Human IL-17: a novel cytokine derived from T cells. J Immunol. 1995;155:5483–5486.
13. Kao CY, Huang F, Chen Y, et al. Up-regulation of CC chemokine ligand 20 expression in human airway epithelium by IL-17 through a JAK-independent but MEK/NF-kappaB-dependent signaling pathway. J Immunol. 2005;175:6676–6685. doi:10.4049/jimmunol.175.10.6676
14. Shahrara S, Pickens SR, Mandelin AM, et al. IL-17-mediated monocyte migration occurs partially through CC chemokine ligand 2/monocyte chemoattractant protein-1 induction. J Immunol. 2010;184:4479–4487. doi:10.4049/jimmunol.0901942
15. Hartupee J, Liu C, Novotny M, et al. IL-17 enhances chemokine gene expression through mRNA stabilization. J Immunol. 2007;179:4135–4141. doi:10.4049/jimmunol.179.6.4135
16. Schwarzenberger P, La Russa V, Miller A, et al. IL-17 stimulates granulopoiesis in mice: use of an alternate, novel gene therapy-derived method for in vivo evaluation of cytokines. J Immunol. 1998;161:6383–6389.
17. Liang SC, Tan XY, Luxenberg DP, et al. Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides. J Exp Med. 2006;203:2271–2279. doi:10.1084/jem.20061308
18. Taams LS, Steel KJA, Srenathan U, et al. IL-17 in the immunopathogenesis of spondyloarthritis. Nat Rev Rheumatol. 2018;14:453–466. doi:10.1038/s41584-018-0044-2
19. Chabaud M, Garnero P, Dayer JM, et al. Contribution of interleukin 17 to synovium matrix destruction in rheumatoid arthritis. Cytokine. 2000;12:1092–1099. doi:10.1006/cyto.2000.0681
20. Koenders MI, Kolls JK, Oppers-Walgreen B, et al. Interleukin-17 receptor deficiency results in impaired synovial expression of interleukin-1 and matrix metalloproteinases 3, 9, and 13 and prevents cartilage destruction during chronic reactivated streptococcal cell wall-induced arthritis. Arthritis Rheum. 2005;52:3239–3247. doi:10.1002/art.21342
21. Kotake S, Udagawa N, Takahashi N, et al. IL-17 in synovial fluids from patients with rheumatoid arthritis is a potent stimulator of osteoclastogenesis. J Clin Invest. 1999;103:1345–1352. doi:10.1172/JCI5703
22. Pickens SR, Volin MV, Mandelin AM, et al. IL-17 contributes to angiogenesis in rheumatoid arthritis. J Immunol. 2010;184:3233–3241. doi:10.4049/jimmunol.0903271
23. Gullick NJ, Evans HG, Church LD, et al. Linking power Doppler ultrasound to the presence of th17 cells in the rheumatoid arthritis joint. PLoS One. 2010;5:e12516. doi:10.1371/journal.pone.0012516
24. Chang SH, Dong C. A novel heterodimeric cytokine consisting of IL-17 and IL-17F regulates inflammatory responses. Cell Res. 2007;17:435–440. doi:10.1038/cr.2007.35
25. Yang XO, Chang SH, Park H, et al. Regulation of inflammatory responses by IL-17F. J Exp Med. 2008;205:1063–1075. doi:10.1084/jem.20071978
26. Kawaguchi M, Adachi M, Oda N, et al. IL-17 cytokine family. J Allergy Clin Immunol. 2004;114:1265–1273. doi:10.1016/j.jaci.2004.10.019
27. Sorbello V, Ciprandi G, Di Stefano A, et al. Nasal IL-17F is related to bronchial IL-17F/neutrophilia and exacerbations in stable atopic severe asthma. Allergy. 2015;70:236–240. doi:10.1111/all.12547
28. Billingham ME. Models of arthritis and the search for anti-arthritic drugs. Pharmacol Ther. 1983;21:389–428. doi:10.1016/0163-7258(83)90062-1
29. Bush KA, Farmer KM, Walker JS, et al. Reduction of joint inflammation and bone erosion in rat adjuvant arthritis by treatment with interleukin-17 receptor IgG1 Fc fusion protein. Arthritis Rheum. 2002;46:802–805. doi:10.1002/art.10173
30. Nakae S, Nambu A, Sudo K, et al. Suppression of immune induction of collagen-induced arthritis in IL-17-deficient mice. J Immunol. 2003;171:6173–6177. doi:10.4049/jimmunol.171.11.6173
31. Corneth OB, Mus AM, Asmawidjaja PS, et al. Absence of interleukin-17 receptor a signaling prevents autoimmune inflammation of the joint and leads to a Th2-like phenotype in collagen-induced arthritis. Arthritis Rheumatol. 2014;66:340–349. doi:10.1002/art.38229
32. Lubberts E, Koenders MI, van den Berg WB. The role of T-cell interleukin-17 in conducting destructive arthritis: lessons from animal models. Arthritis Res Ther. 2005;7:29–37. doi:10.1186/ar1478
33. Lubberts E, Joosten LA, van de Loo FA, et al. Overexpression of IL-17 in the knee joint of collagen type II immunized mice promotes collagen arthritis and aggravates joint destruction. Inflamm Res. 2002;51:102–104. doi:10.1007/BF02684010
34. Vieira-Sousa E, van Duivenvoorde LM, Fonseca JE, et al. Review: animal models as a tool to dissect pivotal pathways driving spondyloarthritis. Arthritis Rheumatol. 2015;67:2813–2827. doi:10.1002/art.39282
35. DeLay ML, Turner MJ, Klenk EI, et al. HLA-B27 misfolding and the unfolded protein response augment interleukin-23 production and are associated with Th17 activation in transgenic rats. Arthritis Rheum. 2009;60:2633–2643. doi:10.1002/art.24763
36. Glatigny S, Fert I, Blaton MA, et al. Proinflammatory Th17 cells are expanded and induced by dendritic cells in spondylarthritis-prone HLA-B27-transgenic rats. Arthritis Rheum. 2012;64:110–120. doi:10.1002/art.33321
37. Sakaguchi N, Takahashi T, Hata H, et al. Altered thymic T-cell selection due to a mutation of the ZAP-70 gene causes autoimmune arthritis in mice. Nature. 2003;426:454–460. doi:10.1038/nature02119
38. Ruutu M, Thomas G, Steck R, et al. β-glucan triggers spondylarthritis and Crohn’s disease-like ileitis in SKG mice. Arthritis Rheum. 2012;64:2211–2222. doi:10.1002/art.34423
39. Benham H, Rehaume LM, Hasnain SZ, et al. Interleukin-23 mediates the intestinal response to microbial β-1,3-glucan and the development of spondyloarthritis pathology in SKG mice. Arthritis Rheumatol. 2014;66:1755–1767. doi:10.1002/art.38638
40. Gillet P, Bannwarth B, Charrière G, et al. Studies on type II collagen induced arthritis in rats: an experimental model of peripheral and axial ossifying enthesopathy. J Rheumatol. 1989;16:721–728.
41. Sherlock JP, Joyce-Shaikh B, Turner SP, et al. IL-23 induces spondyloarthropathy by acting on ROR-γt+ CD3+CD4-CD8- entheseal resident T cells. Nat Med. 2012;18:1069–1076. doi:10.1038/nm.2817
42. Ebihara S, Date F, Dong Y, et al. Interleukin-17 is a critical target for the treatment of ankylosing enthesitis and psoriasis-like dermatitis in mice. Autoimmunity. 2015;48:259–266. doi:10.3109/08916934.2014.976630
43. Wendling D, Cedoz JP, Racadot E, et al. Serum IL-17, BMP-7, and bone turnover markers in patients with ankylosing spondylitis. Joint Bone Spine. 2007;74:304–305. doi:10.1016/j.jbspin.2006.11.005
44. Romero-Sanchez C, Jaimes DA, Londoño J, et al. Association between Th-17 cytokine profile and clinical features in patients with spondyloarthritis. Clin Exp Rheumatol. 2011;29:828–834.
45. Chen WS, Chang YS, Lin KC, et al. Association of serum interleukin-17 and interleukin-23 levels with disease activity in Chinese patients with ankylosing spondylitis. J Chin Med Assoc. 2012;75:303–308. doi:10.1016/j.jcma.2012.05.006
46. Xueyi L, Lina C, Zhenbiao W, et al. Levels of circulating Th17 cells and regulatory T cells in ankylosing spondylitis patients with an inadequate response to anti-TNF-α therapy. J Clin Immunol. 2013;33:151–161. doi:10.1007/s10875-012-9774-0
47. Leipe J, Grunke M, Dechant C, et al. Role of Th17 cells in human autoimmune arthritis. Arthritis Rheum. 2010;62:2876–2885. doi:10.1002/art.27622
48. Singh R, Aggarwal A, Misra R. Th1/Th17 cytokine profiles in patients with reactive arthritis/undifferentiated spondyloarthropathy. J Rheumatol. 2007;34:2285–2290.
49. Raychaudhuri SP, Raychaudhuri SK, Genovese MC. IL-17 receptor and its functional significance in psoriatic arthritis. Mol Cell Biochem. 2012;359:419–429. doi:10.1007/s11010-011-1036-6
50. Ciccia F, Bombardieri M, Principato A, et al. Overexpression of interleukin-23, but not interleukin-17, as an immunologic signature of subclinical intestinal inflammation in ankylosing spondylitis. Arthritis Rheum. 2009;60:955–965. doi:10.1002/art.24389
51. van Baarsen LG, Lebre MC, van der Coelen D, et al. Heterogeneous expression pattern of interleukin 17A (IL-17A), IL-17F and their receptors in synovium of rheumatoid arthritis, psoriatic arthritis and osteoarthritis: possible explanation for nonresponse to anti-IL-17 therapy? Arthritis Res Ther. 2014;16:426. doi:10.1186/s13075-014-0426-z
52. Glatt S, Baeten D, Baker T, et al. Dual IL-17A and IL-17F neutralisation by bimekizumab in psoriatic arthritis: evidence from preclinical experiments and a randomised placebo-controlled clinical trial that IL-17F contributes to human chronic tissue inflammation. Ann Rheum Dis. 2018;77:523–532. doi:10.1136/annrheumdis-2017-212127
53. Zrioual S, Ecochard R, Tournadre A, et al. Genome-wide comparison between IL-17A- and IL-17F-induced effects in human rheumatoid arthritis synoviocytes. J Immunol. 2009;182:3112–3120. doi:10.4049/jimmunol.0801967
54. Fossiez F, Djossou O, Chomarat P, et al. T cell interleukin-17 induces stromal cells to produce proinflammatory and hematopoietic cytokines. J Exp Med. 1996;183:2593–2603. doi:10.1084/jem.183.6.2593
55. Zrioual S, Toh ML, Tournadre A, et al. IL-17RA and IL-17RC receptors are essential for IL-17A-induced ELR+ CXC chemokine expression in synoviocytes and are overexpressed in rheumatoid blood. J Immunol. 2008;180:655–663. doi:10.4049/jimmunol.180.1.655
56. Chabaud M, Fossiez F, Taupin JL, et al. Enhancing effect of IL-17 on IL-1-induced IL-6 and leukemia inhibitory factor production by rheumatoid arthritis synoviocytes and its regulation by Th2 cytokines. J Immunol. 1998;161:409–414.
57. Kawashiri SY, Kawakami A, Iwamoto N, et al. Proinflammatory cytokines synergistically enhance the production of chemokine ligand 20 (CCL20) from rheumatoid fibroblast-like synovial cells in vitro and serum CCL20 is reduced in vivo by biologic disease-modifying antirheumatic drugs. J Rheumatol. 2009;36:2397–2402. doi:10.3899/jrheum.090132
58. Teunissen MB, Koomen CW, de Waal Malefyt R, et al. Interleukin-17 and interferon-gamma synergize in the enhancement of proinflammatory cytokine production by human keratinocytes. J Invest Dermatol. 1998;111:645–649. doi:10.1046/j.1523-1747.1998.00347.x
59. Wedebye Schmidt EG, Larsen HL, Kristensen NN, et al. TH17 cell induction and effects of IL-17A and IL-17F blockade in experimental colitis. Inflamm Bowel Dis. 2013;19:1567–1576. doi:10.1097/MIB.0b013e318286fa1c
60. Henness S, van Thoor E, Ge Q, et al. IL-17A acts via p38 MAPK to increase stability of TNF-alpha-induced IL-8 mRNA in human ASM. Am J Physiol Lung Cell Mol Physiol. 2006;290:L1283–1290. doi:10.1152/ajplung.00367.2005
61. Friday SC, Fox DA. Phospholipase D enzymes facilitate IL-17- and TNFα-induced expression of proinflammatory genes in rheumatoid arthritis synovial fibroblasts (RASF). Immunol Lett. 2016;174:9–18. doi:10.1016/j.imlet.2016.04.001
62. Srenathan U, Steel K, Taams LS. IL-17+ CD8+ T cells: differentiation, phenotype and role in inflammatory disease. Immunol Lett. 2016;178:20–26. doi:10.1016/j.imlet.2016.05.001
63. Papotto PH, Ribot JC, Silva-Santos B. IL-17+ γδ T cells as kick-starters of inflammation. Nat Immunol. 2017;18:604–611. doi:10.1038/ni.3726
64. Hazenberg MD, Spits H. Human innate lymphoid cells. Blood. 2014;124:700–709. doi:10.1182/blood-2013-11-427781
65. Menon B, Gullick NJ, Walter GJ, et al. Interleukin-17+CD8+ T cells are enriched in the joints of patients with psoriatic arthritis and correlate with disease activity and joint damage progression. Arthritis Rheumatol. 2014;66:1272–1281. doi:10.1002/art.38376
66. Tzartos JS, Friese MA, Craner MJ, et al. Interleukin-17 production in central nervous system-infiltrating T cells and glial cells is associated with active disease in multiple sclerosis. Am J Pathol. 2008;172:146–155. doi:10.2353/ajpath.2008.070690
67. Ortega C, Fernández-A S, Carrillo JM, et al. IL-17-producing CD8+ T lymphocytes from psoriasis skin plaques are cytotoxic effector cells that secrete Th17-related cytokines. J Leukoc Biol. 2009;86:435–443. doi:10.1189/JLB.0109046
68. Res PC, Piskin G, de Boer OJ, et al. Overrepresentation of IL-17A and IL-22 producing CD8 T cells in lesional skin suggests their involvement in the pathogenesis of psoriasis. PLoS One. 2010;5:e14108. doi:10.1371/journal.pone.0014108
69. Hijnen D, Knol EF, Gent YY, et al. CD8(+) T cells in the lesional skin of atopic dermatitis and psoriasis patients are an important source of IFN-γ, IL-13, IL-17, and IL-22. J Invest Dermatol. 2013;133:973–979. doi:10.1038/jid.2012.456
70. Di Meglio P, Villanova F, Navarini AA, et al. Targeting CD8(+) T cells prevents psoriasis development. J Allergy Clin Immunol. 2016;138:274–276.e276. doi:10.1016/j.jaci.2015.10.046
71. Wang C, Liao Q, Hu Y, et al. T lymphocyte subset imbalances in patients contribute to ankylosing spondylitis. Exp Ther Med. 2015;9:250–256. doi:10.3892/etm.2014.2046
72. Al-Mossawi MH, Chen L, Fang H, et al. Unique transcriptome signatures and GM-CSF expression in lymphocytes from patients with spondyloarthritis. Nat Commun. 2017;8:1510. doi:10.1038/s41467-017-01771-2
73. Steel KWS, Srenathan U, Chan E, Kirkham BW, Taams LS. 0016 Synovial IL-17+CD8+ T cells are a pro-inflammatory tissue resident population enriched in spondyloarthritis. Ann Rheum Dis. 2018;77:A8–A9.
74. Cheuk S, Schlums H, Gallais Sérézal I, et al. CD49a expression defines tissue-resident CD8 + T cells poised for cytotoxic function in human skin. Immunity. 2017;46:287–300. doi:10.1016/j.immuni.2017.01.009
75. Cheuk S, Wikén M, Blomqvist L, et al. Epidermal Th22 and Tc17 cells form a localized disease memory in clinically healed psoriasis. J Immunol. 2014;192:3111–3120. doi:10.4049/jimmunol.1302313
76. Clark RA, Chong B, Mirchandani N, et al. The vast majority of CLA+ T cells are resident in normal skin. J Immunol. 2006;176:4431–4439. doi:10.4049/jimmunol.176.7.4431
77. Watanabe R, Gehad A, Yang C, et al. Human skin is protected by four functionally and phenotypically discrete populations of resident and recirculating memory T cells. Sci Transl Med. 2015;7:279ra239. doi:10.1126/scitranslmed.3010302
78. Laggner U, Di Meglio P, Perera GK, et al. Identification of a novel proinflammatory human skin-homing Vγ9Vδ2 T cell subset with a potential role in psoriasis. J Immunol. 2011;187:2783–2793. doi:10.4049/jimmunol.1100804
79. Petrelli A, van Wijk F. CD8(+) T cells in human autoimmune arthritis: the unusual suspects. Nat Rev Rheumatol. 2016;12:421–428. doi:10.1038/nrrheum.2016.74
80. Dusseaux M, Martin E, Serriari N, et al. Human MAIT cells are xenobiotic-resistant, tissue-targeted, CD161hi IL-17-secreting T cells. Blood. 2011;117:1250–1259. doi:10.1182/blood-2010-08-303339
81. Teunissen MBM, Yeremenko NG, Baeten DLP, et al. The IL-17A-producing CD8+ T-cell population in psoriatic lesional skin comprises mucosa-associated invariant T cells and conventional T cells. J Invest Dermatol. 2014;134:2898–2907. doi:10.1038/jid.2014.261
82. Hayashi E, Chiba A, Tada K, et al. Involvement of mucosal-associated invariant T cells in ankylosing spondylitis. J Rheumatol. 2016;43:1695–1703. doi:10.3899/jrheum.151133
83. Gracey E, Qaiyum Z, Almaghlouth I, et al. IL-7 primes IL-17 in mucosal-associated invariant T (MAIT) cells, which contribute to the Th17-axis in ankylosing spondylitis. Ann Rheum Dis. 2016;75:2124–2132. doi:10.1136/annrheumdis-2015-208902
84. Yoshiga Y, Goto D, Segawa S, et al. Invariant NKT cells produce IL-17 through IL-23-dependent and -independent pathways with potential modulation of Th17 response in collagen-induced arthritis. Int J Mol Med. 2008;22:369–374.
85. Kenna TJ, Davidson SI, Duan R, et al. Enrichment of circulating interleukin-17-secreting interleukin-23 receptor-positive γ/δ T cells in patients with active ankylosing spondylitis. Arthritis Rheum. 2012;64:1420–1429. doi:10.1002/art.33507
86. Gaur P, Misra R, Aggarwal A. Natural killer cell and gamma delta T cell alterations in enthesitis related arthritis category of juvenile idiopathic arthritis. Clin Immunol. 2015;161:163–169. doi:10.1016/j.clim.2015.07.012
87. Guggino G, Ciccia F, Di Liberto D, et al. Interleukin (IL)-9/IL-9R axis drives γδ T cells activation in psoriatic arthritis patients. Clin Exp Immunol. 2016;186:277–283. doi:10.1111/cei.12853
88. Chowdhury AC, Chaurasia S, Mishra SK, et al. IL-17 and IFN-γ producing NK and γδ-T cells are preferentially expanded in synovial fluid of patients with reactive arthritis and undifferentiated spondyloarthritis. Clin Immunol. 2017;183:207–212. doi:10.1016/j.clim.2017.03.016
89. Cai Y, Shen X, Ding C, et al. Pivotal role of dermal IL-17-producing γδ T cells in skin inflammation. Immunity. 2011;35:596–610. doi:10.1016/j.immuni.2011.08.001
90. Gray EE, Suzuki K, Cyster JG. Cutting edge: identification of a motile IL-17-producing gammadelta T cell population in the dermis. J Immunol. 2011;186:6091–6095. doi:10.4049/jimmunol.1100427
91. Campbell JJ, Ebsworth K, Ertl LS, et al. IL-17-secreting γδ T cells are completely dependent upon CCR6 for homing to inflamed skin. J Immunol. 2017;199:3129–3136. doi:10.4049/jimmunol.1700826
92. Reinhardt A, Yevsa T, Worbs T, et al. Interleukin-23-dependent γ/δ T cells produce interleukin-17 and accumulate in the enthesis, aortic valve, and ciliary body in mice. Arthritis Rheumatol. 2016;68:2476–2486. doi:10.1002/art.39732
93. Soare A, Weber S, Maul L, et al. Cutting edge: homeostasis of innate lymphoid cells is imbalanced in psoriatic arthritis. J Immunol. 2018;200:1249–1254. doi:10.4049/jimmunol.1700596
94. Leijten EFA, van Kempen TS, Boes M, et al. Brief report: enrichment of activated group 3 innate lymphoid cells in psoriatic arthritis synovial fluid. Arthritis Rheumatol. 2015;67(10):2673–2678. doi:10.1002/art.39261
95. Noordenbos T, Yeremenko N, Gofita I, et al. Interleukin-17-positive mast cells contribute to synovial inflammation in spondylarthritis. Arthritis Rheum. 2012;64:99–109. doi:10.1002/art.33396
96. Noordenbos T, Blijdorp I, Chen S, et al. Human mast cells capture, store, and release bioactive, exogenous IL-17A. J Leukoc Biol. 2016;100:453–462. doi:10.1189/jlb.3HI1215-542R
97. Wendling D, Verhoeven F, Prati C. Anti-IL-17 monoclonal antibodies for the treatment of ankylosing spondylitis. Expert Opin Biol Ther. 2019;19:55–64. doi:10.1080/14712598.2019.1554053
98. van der Heijde D, Ramiro S, Landewé R, et al. 2016 update of the ASAS-EULAR management recommendations for axial spondyloarthritis. Ann Rheum Dis. 2017;76:978–991. doi:10.1136/annrheumdis-2016-210770
99. Ward MM, Deodhar A, Gensler LS, et al. 2019 Update of the American college of rheumatology/spondylitis association of America/spondyloarthritis research and treatment network recommendations for the treatment of ankylosing spondylitis and nonradiographic axial spondyloarthritis. Arthritis Rheumatol. 2019;71:1599–1613. doi:10.1002/art.41042
100. Braun J, Baraliakos X, Kiltz U. Secukinumab (AIN457) in the treatment of ankylosing spondylitis. Expert Opin Biol Ther. 2016;16:711–722. doi:10.1517/14712598.2016.1167183
101. Baeten D, Baraliakos X, Braun J, et al. Anti-interleukin-17A monoclonal antibody secukinumab in treatment of ankylosing spondylitis: a randomised, double-blind, placebo-controlled trial. Lancet. 2013;382:1705–1713. doi:10.1016/S0140-6736(13)61134-4
102. Baeten D, Sieper J, Braun J, et al. Secukinumab, an interleukin-17A inhibitor, in ankylosing spondylitis. N Engl J Med. 2015;373:2534–2548. doi:10.1056/NEJMoa1505066
103. Sieper J, Deodhar A, Marzo-Ortega H, et al. Secukinumab efficacy in anti-TNF-naive and anti-TNF-experienced subjects with active ankylosing spondylitis: results from the MEASURE 2 study. Ann Rheum Dis. 2017;76:571–592. doi:10.1136/annrheumdis-2016-210023
104. Kishimoto M, Taniguchi A, Fujishige A, et al. Efficacy and safety of secukinumab in Japanese patients with active ankylosing spondylitis: 24-week results from an open-label Phase 3 study (MEASURE 2-J). Mod Rheumatol. 2020;30:132–140. doi:10.1080/14397595.2018.1538004
105. Pavelka K, Kivitz A, Dokoupilova E, et al. Efficacy, safety, and tolerability of secukinumab in patients with active ankylosing spondylitis: a randomized, double-blind phase 3 study, MEASURE 3. Arthritis Res Ther. 2017;19:285. doi:10.1186/s13075-017-1490-y
106. Kivitz AJ, Wagner U, Dokoupilova E, et al. Efficacy and safety of secukinumab 150 mg with and without loading regimen in ankylosing spondylitis: 104-week results from MEASURE 4 study. Rheumatol Ther. 2018;5:447–462. doi:10.1007/s40744-018-0123-5
107. Braun J, Baraliakos X, Deodhar A, et al. Effect of secukinumab on clinical and radiographic outcomes in ankylosing spondylitis: 2-year results from the randomised phase III MEASURE 1 study. Ann Rheum Dis. 2017;76:1070–1077. doi:10.1136/annrheumdis-2016-209730
108. D’Angelo S, Carriero A, Gilio M, et al. Safety of treatment options for spondyloarthritis: a narrative review. Expert Opin Drug Saf. 2018;17:475–486. doi:10.1080/14740338.2018.1448785
109. Torgutalp M, Poddubnyy D. IL-17 inhibition in axial spondyloarthritis: current and future perspectives. Expert Opin Biol Ther. 2019;19:631–641. doi:10.1080/14712598.2019.1605352
110. Baraliakos X, Braun J, Deodhar A, et al. Long-term efficacy and safety of secukinumab 150 mg in ankylosing spondylitis: 5-year results from the phase III MEASURE 1 extension study. RMD Open. 2019;5:e001005. doi:10.1136/rmdopen-2019-001005
111. Marzo-Ortega H, Sieper J, Kivitz AJ, et al. 5-year efficacy and safety of secukinumab in patients with ankylosing spondylitis: end-of-study results from the phase 3 MEASURE 2 trial. Lancet Rheumatol. 2020;2:e339–e346. doi:10.1016/S2665-9913(20)30066-7
112. Baraliakos X, Van den Bosch F, Machado PM, et al. Achievement of remission endpoints with secukinumab over 3 years in active ankylosing spondylitis: pooled analysis of two phase 3 studies. Rheumatol Ther. 2020. doi:10.1007/s40744-020-00269-6
113. Dubash S, Bridgewood C, McGonagle D, et al. The advent of IL-17A blockade in ankylosing spondylitis: secukinumab, ixekizumab and beyond. Expert Rev Clin Immunol. 2019;15:123–134. doi:10.1080/1744666X.2019.1561281
114. Deodhar A, Blanco R, Dokoupilová E, et al. Improvement of signs and symptoms of nonradiographic axial spondyloarthritis in patients treated with secukinumab: primary results of a randomized, placebo-controlled phase III study. Arthritis Rheumatol. 2021;73(1):110–120. doi:10.1002/art.41477
115. Wang P, Zhang S, Hu B, et al. Efficacy and safety of interleukin-17A inhibitors in patients with ankylosing spondylitis: a systematic review and meta-analysis of randomized controlled trials. Clin Rheumatol. 2021. doi:10.1007/s10067-020-05545-y
116. Gentileschi S, Vitale A, Rigante D, et al. Prompt clinical response to secukinumab in patients with axial spondyloarthritis: real life observational data from three italian referral centers. Isr Med Assoc J. 2018;20:438–441.
117. Gentileschi S, Rigante D, Sota J, et al. Long-term effectiveness of secukinumab in patients with axial spondyloarthritis. Mediators Inflamm. 2020;2020:6983272. doi:10.1155/2020/6983272
118. Mann HF, Závada J, Šenolt L, et al. Real world use of secukinumab for treatment of axial spondyloarthritis and psoriatic arthritis: nationwide results from the ATTRA registry. Clin Exp Rheumatol. 2019;37:342–343.
119. Elliott A, Wright G. Real-world data on secukinumab use for psoriatic arthritis and ankylosing spondylitis. Ther Adv Musculoskelet Dis. 2019;11:1759720X19858510. doi:10.1177/1759720X19858510
120. Williams T, Wadeley A, Bond D, et al. Real-world experience of secukinumab treatment for ankylosing spondylitis at the Royal National Hospital for Rheumatic Diseases, Bath. Clin Rheumatol. 2020;39:1501–1504. doi:10.1007/s10067-020-04944-5
121. Chimenti MS, Fonti GL, Conigliaro P, et al. One-year effectiveness, retention rate, and safety of secukinumab in ankylosing spondylitis and psoriatic arthritis: a real-life multicenter study. Expert Opin Biol Ther. 2020;20:813–821. doi:10.1080/14712598.2020.1761957
122. Michelsen B, Lindström U, Codreanu C, et al. Drug retention, inactive disease and response rates in 1860 patients with axial spondyloarthritis initiating secukinumab treatment: routine care data from 13 registries in the EuroSpA collaboration. RMD Open. 2020;6(3):e001280. doi:10.1136/rmdopen-2020-001280
123. Yu CL, Yang CH, Chi CC. Drug survival of biologics in treating ankylosing spondylitis: a systematic review and meta-analysis of real-world evidence. BioDrugs. 2020;34:669–679. doi:10.1007/s40259-020-00442-x
124. Goeree R, Chiva-Razavi S, Gunda P, et al. Cost-effectiveness analysis of secukinumab in ankylosing spondylitis from the Canadian perspective. J Med Econ. 2019;22:45–52. doi:10.1080/13696998.2018.1539400
125. Purmonen T, Puolakka K, Mishra D, et al. Cost-effectiveness of secukinumab compared to other biologics in the treatment of ankylosing spondylitis in Finland. Clinicoecon Outcomes Res. 2019;11:159–168. doi:10.2147/CEOR.S192235
126. Emery P, Van Keep M, Beard S, et al. Cost effectiveness of secukinumab for the treatment of active ankylosing spondylitis in the UK. Pharmacoeconomics. 2018;36:1015–1027. doi:10.1007/s40273-018-0675-9
127. Kiltz U, Sfikakis PP, Gaffney K, et al. Secukinumab use in patients with moderate to severe psoriasis, psoriatic arthritis and ankylosing spondylitis in real-world setting in Europe: baseline data from SERENA study. Adv Ther. 2020;37:2865–2883. doi:10.1007/s12325-020-01352-8
128. Micheroli R, Tellenbach C, Scherer A, et al. Effectiveness of secukinumab versus an alternative TNF inhibitor in patients with axial spondyloarthritis previously exposed to TNF inhibitors in the Swiss Clinical Quality Management cohort. Ann Rheum Dis. 2020;79:1203–1209. doi:10.1136/annrheumdis-2019-215934
129. Glintborg B, Lindstrom U, Di Giuseppe D, et al. One-year treatment outcomes of secukinumab versus tumor necrosis factor inhibitors in Spondyloarthritis. Arthritis Care Res. 2020. doi:10.1002/acr.24523
130. van der Heijde D, Cheng-Chung Wei J, Dougados M, et al. Ixekizumab, an interleukin-17A antagonist in the treatment of ankylosing spondylitis or radiographic axial spondyloarthritis in patients previously untreated with biological disease-modifying anti-rheumatic drugs (COAST-V): 16 week results of a phase 3 randomised, double-blind, active-controlled and placebo-controlled trial. Lancet. 2018;392:2441–2451. doi:10.1016/S0140-6736(18)31946-9
131. Deodhar A, Poddubnyy D, Pacheco-Tena C, et al. Efficacy and safety of ixekizumab in the treatment of radiographic axial spondyloarthritis: sixteen-week results from a phase III randomized, double-blind, placebo-controlled trial in patients with prior inadequate response to or intolerance of tumor necrosis factor inhibitors. Arthritis Rheumatol. 2019;71:599–611. doi:10.1002/art.40753
132. Ritchlin C, Adamopoulos IE. Axial spondyloarthritis: new advances in diagnosis and management. BMJ. 2021;372:m4447. doi:10.1136/bmj.m4447
133. Rademacher J, Poddubnyy D. Emerging drugs for the treatment of axial spondyloarthritis. Expert Opin Emerg Drugs. 2018;23:83–96. doi:10.1080/14728214.2018.1445719
134. Dougados M, Wei JC, Landewé R, et al. Efficacy and safety of ixekizumab through 52 weeks in two phase 3, randomised, controlled clinical trials in patients with active radiographic axial spondyloarthritis (COAST-V and COAST-W). Ann Rheum Dis. 2020;79:176–185. doi:10.1136/annrheumdis-2019-216118
135. Deodhar A, van der Heijde D, Gensler LS, et al. Ixekizumab for patients with non-radiographic axial spondyloarthritis (COAST-X): a randomised, placebo-controlled trial. Lancet. 2020;395:53–64. doi:10.1016/S0140-6736(19)32971-X
136. Kiwalkar S, Beier S, Deodhar A. Ixekizumab for treating ankylosing spondylitis. Immunotherapy. 2019;11:1273–1282. doi:10.2217/imt-2019-0094
137. San Koo B, Kim TH. The role of ixekizumab in non-radiographic axial spondyloarthritis. Ther Adv Musculoskelet Dis. 2021;13:1759720X20986734. doi:10.1177/1759720X20986734
138. Schreiber S, Colombel JF, Feagan BG, et al. Incidence rates of inflammatory bowel disease in patients with psoriasis, psoriatic arthritis and ankylosing spondylitis treated with secukinumab: a retrospective analysis of pooled data from 21 clinical trials. Ann Rheum Dis. 2019;78:473–479. doi:10.1136/annrheumdis-2018-214273
139. Papp KA, Leonardi C, Menter A, et al. Brodalumab, an anti-interleukin-17-receptor antibody for psoriasis. N Engl J Med. 2012;366:1181–1189. doi:10.1056/NEJMoa1109017
140. Lebwohl MG, Papp KA, Marangell LB, et al. Psychiatric adverse events during treatment with brodalumab: analysis of psoriasis clinical trials. J Am Acad Dermatol. 2018;78:81–89.e85. doi:10.1016/j.jaad.2017.08.024
141. Wei JCC, Kim TH, Kishimoto M, et al. OP0234 efficacy and safety of brodalumab, an anti-interleukin-17 receptor a monoclonal antibody, in patients with axial spondyloarthritis: a 16 week results of a phase 3, multicenter, randomized, double-blind, placebo-controlled study. Ann Rheum Dis. 2019;78:195. doi:10.1136/annrheumdis-2019-eular.6888
142. van der Heijde D, Gensler LS, Deodhar A, et al. Dual neutralisation of interleukin-17A and interleukin-17F with bimekizumab in patients with active ankylosing spondylitis: results from a 48-week phase IIb, randomised, double-blind, placebo-controlled, dose-ranging study. Ann Rheum Dis. 2020;79:595–604. doi:10.1136/annrheumdis-2020-216980
143. Erdes S, Nasonov E, Kunder E, et al. Primary efficacy of netakimab, a novel interleukin-17 inhibitor, in the treatment of active ankylosing spondylitis in adults. Clin Exp Rheumatol. 2020;38:27–34.
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms.php
and incorporate the Creative Commons Attribution
- Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.