Back to Journals » Journal of Inflammation Research » Volume 19
Analysis of the Immunoinflammatory Microenvironment Characteristics of Epilepsy Based on Single-Cell Transcriptomics
Authors Chen Y, Zhang W, Xin M, Liu M, Zhang T, Xiao J, Liang J, Wu X
Received 11 October 2025
Accepted for publication 7 January 2026
Published 12 January 2026 Volume 2026:19 573666
DOI https://doi.org/10.2147/JIR.S573666
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Dr Adam Bachstetter
Yutong Chen,1,2 Wei Zhang,1 Meiying Xin,1 Ming Liu,1 Tongye Zhang,1 Jinyu Xiao,1,2 Jianmin Liang,1,2 Xuemei Wu1,2
1Department of Pediatric Neurology, Children’s Medical Center, The First Hospital of Jilin University, Changchun, People’s Republic of China; 2Jilin Provincial Key Laboratory of Pediatric Neurology, Changchun, People’s Republic of China
Correspondence: Xuemei Wu, Department of Pediatric Neurology, Children’s Medical Center, The First Hospital of Jilin University, 1 Xinmin Street, Changchun, Jilin, 130000, People’s Republic of China, Email [email protected]
Abstract: Epilepsy is a common neurological disease that affects a wide range of people and seriously impairs patients’ quality of life and social adaptability. Neuroinflammation plays an important role in the pathogenesis of epilepsy. Glial cells, particularly astrocytes and their differentiated forms, as well as their interactions with other cell types, are critical in the process of neuroinflammation. A better understanding of the inflammatory mechanisms of epilepsy might facilitate the development of new therapeutic strategies. Single-cell RNA sequencing has provided new insights into the inflammatory microenvironment of the nervous system in epilepsy by enabling high-resolution characterization of cell type-specific gene expression and signaling pathways. In this review, we summarized the differentiation, interactions, and transcriptional dynamics of microglia, astrocytes, and neurons in epilepsy using single-cell sequencing data from patients and animal models, providing a basis for the development of new strategies for epilepsy treatment.
Keywords: epilepsy, inflammation, microglia, astrocytes, single-cell RNA sequencing
Introduction
Epilepsy is a common neurological disorder in which inflammation plays an important role in disease progression.1 The activation of microglia, astrocytes, the blood–brain barrier (BBB) endothelial cells, and peripheral immune cells, along with the concomitant production of neuroinflammatory mediators, was initially studied in neuroinflammation. As neuroinflammation has been recognized as a trigger of epilepsy in various types of epilepsies. The association between epilepsy and neuroinflammation has gradually emerged. Over the past decade, studies using both acute and chronic epilepsy models have identified neuroinflammation as a key factor for epileptogenesis. Glial cells, particularly astrocytes and their differentiated forms play important roles in epilepsy by interacting with T cells and monocytes in the nervous system.2 In epilepsy, glial cells interact with T cells and monocytes in the nervous system through multiple key factors, such as interleukin-1 beta (IL-1β) and tumor necrosis factor-alpha (TNF-α).3 Complex inflammatory responses compromise the integrity and regulatory function of the BBB, thereby disrupting central nervous system homeostasis.4 Collectively, these changes further exacerbate neuroinflammation. Despite the availability of a wide range of antiepileptic drugs, approximately one-third of patients suffer from uncontrollable seizures.5 Preventing epilepsy development or improving long-term prognosis remains a difficult and unresolved problem. The regulation of neuroinflammation might relieve seizures and ictal discharge during epilepsy. To date, many studies have focused on the effect and mechanism of neuroinflammation during the pathogenesis of epilepsy. Single-cell RNA sequencing (scRNA-seq) is a technique that could reveal cell specificities, differentiation, gene expression, and interactions between different cells by RNA analyzing in individual cells.6 This genetic analysis has deepened our understanding of the mechanisms underlying epilepsy. However, despite its superior resolution, findings derived from scRNA-seq require further validation through functional experiments and integration with spatial information.7 In this review, we summarized the interactions between different cell types in epilepsy, focusing on microglia, astrocytes, neurons involved in the inflammatory response and revealed the differentiation, gene expression, and interactions between different cells during epilepsy.
Developmental Differentiation of Glial Cells and Differentially Expressed Genes (DEGs) in Epilepsy
Recent studies have shown that localized neuroinflammation can trigger seizures in epilepsy. In particular, activated microglia and astrocytes may play a key role in this process. For instance, Liu et al reported elevated expression of pro-inflammatory genes (eg Spp1, Lyz2, and Cd14) in all glial cell types in an epileptic mouse model. Conversely, genes such as Tle4 and Sipa113 are downregulated in glial cells and their reduced expression are mainly related to axon genesis, dendrite development, neuronal spine formation, and microtubule protein binding. Spatial transcriptomics analysis has revealed activation of pathways involving astrocytic synaptic projections in the hippocampal region of temporal lobe epilepsy, consistent with reactive changes in microglia and astrocytes.8
Microglia are innate immune cells in the nervous system that account for approximately 5%–10% of brain cells. The morphology and function of microglia change throughout the progression of neurological diseases.9 Several transcriptional characteristics of microglia have been reported in different diseases, such as the amyotrophic lateral sclerosis-specific phenotype,10 microglia type associated with neurodegenerative diseases type,11 and lipopolysaccharide (LPS) subgroup.12 RNA sequencing of single-cell suspensions obtained from epileptic brain tissues revealed that the majority of cells were microglia.13 Different clusters of microglia widely express cytokines, such as IL-1β, TNF-α, and C-C chemokine ligand 4 (CCL4), which interactively affect neuronal excitability. For instance, IL-1β inhibits the transport of glutamate by astrocytes, thereby increasing extracellular glutamate concentrations. It also abolishes the inhibitory effect of gamma-aminobutyric acid (GABA) and increases susceptibility to epileptic seizures.14 TNF-α promotes the apoptosis of hippocampal neural progenitor cells and inhibits neurogenesis.15 Cao et al constructed protein–protein interactions networks of DEGs and grouped them into distinct functional modules. They found that the top 10 DEGs in the inflammatory response module of patients with epilepsy, compared with the findings in healthy controls, were associated with microglial migration.16 Westfall et al performed single-nucleus RNA sequencing on surgically resected brain tissues from patients with Rasmussen’s encephalitis and identified 11 microglia subclusters. Eight of these subclusters exhibited high expression of inflammation-related genes. Clusters 7 and 10 showed abundant expression of GRID2, a gene implicated in microglia-neuron crosstalk and the organization of synaptic and postsynaptic tissues. These microglia also highly expressed genes that regulate glutamate receptor signaling pathways and glutamatergic synaptic transmission. Dysregulation of the these genes may contribute to inflammation and seizures.17 Microglia are commonly classified into two polarization states, namely M1 and M2, each categorized by distinct markers and mediators. M1 microglia exert pro-inflammatory effects and potentially disrupt axonal regeneration, whereas M2 microglia exhibit anti-inflammatory properties and potentially promote oligodendrocyte production.18 The expression of both pro-inflammatory and anti-inflammatory cytokines increases in microglia following the occurrence of epilepsy. Inflammatory microglia clusters are particularly prominent at epileptogenic foci. It has been demonstrated that epileptic foci can be identified by localizing microglia exhibiting a pro-inflammatory phenotype.19 In the initial stage of the disease, microglia engulf dead cell debris and release cytokines, mitigating tissue damage. However, as the inflammatory response persists, this initial protective effect gradually diminishes, leading to an increase in harmful effects.20
Astrocytes represent the most abundant cell type in the brain, supporting neurons, participating in the formation of the BBB, and playing an important role in immune regulation. Wheeler et al identified the broadest astrocyte subpopulation in experimental autoimmune encephalomyelitis (EAE) using the single-cell sequencing technique. These cells are characterized by the activation of the nuclear factor kappa B and inducible nitric oxide synthase pathway associated with pro-inflammatory and neurotoxic cells, confirming the pro-inflammatory and neurotoxic characteristics of astrocytes.21 Astrocytes become reactive during epilepsy and contribute to neuroinflammation.22 Similar to microglia, activated astrocytes can be classified into two types: A1 (relatively protective) and A2 (relatively destructive).23 Qian et al classified astrocytes into eight subpopulations and identified seven functional expression modules by integrating transcriptional signatures from large-scale scRNA-seq datasets covering a wide range of human central nervous system diseases. In epilepsy, clusters 1, 2, 3, and 5 are involved in the onset and development of epilepsy. The SNHG14 gene expressed in cluster 1 can inhibit inflammatory responses, whereas the FOS gene expressed in cluster 2 is involved in oxidative stress. Cluster 5 is characterized by high expression of immune response genes. In addition, the expression of the immune function M5 module is significantly increased during the occurrence of epilepsy. The SLC8A1 and LRMDA genes, which are abundant in this module, possess immune activation properties.24 However, since these astrocyte clusters share similar transcriptional profile in multiple sclerosis, this evidence does not definitively link specific gene expression patterns to seizure generation but rather suggests a general direction for future research.
In epilepsy, the expression of immune-related genes is increased in glial cells, allowing them to differentiate into different phenotypes that can either protect or damage neurons. These responses have markedly different effects on the regression of inflammation. Promoting the early differentiation of glial cells toward an anti-inflammatory and neuroprotective phenotype, along with the upregulation of immunoprotective genes, might help delay or even reverse the pathological process of epilepsy.
Interaction Between Microglia and Astrocytes in Epilepsy
Astrocytes and microglia, as glial cells in the brain parenchyma, play important roles in the inflammatory response associated with epilepsy. Complement component 3 (C3) is an important marker of A1 astrocytes that is predominately produced during neuroinflammatory processes.25 The activation of A1 astrocytes is induced by microglia that are activated during inflammatory response.26 Activated microglia secrete various molecules, such as IL-1α, TNF-α, and complement C1q, which work together to activate astrocytes and promote C3 expression.27 Previous research suggested that the activation of astrocytes requires the coordinated action of these molecules, whereas individual molecules alone cannot induce this activation.28 C3 released by astrocytes is cleaved into C3a and C3b. C3a is recognized by the C3a receptor (C3aR) on the surface of microglia.29 When C3aR is activated, it further promotes the inflammatory response in microglia, stimulates the secretion of complement C1q and pro-inflammatory cytokines, enhances phagocytic ability, and thus promotes the activation of astrocytes.30 The positive feedback between C3aR and C3 in astrocytes promotes the activation of the A1 phenotype in rats, which can affect the activation and function of microglia. In LPS-induced rats, C3aR knockout in astrocytes resulted in the upregulation of the following genes in different clusters of microglia compared with the findings in wild-type controls, as identified by scRNA-seq:31 cluster 0, Atf3, Ac134224.1, and Il1b; cluster 1, Rt1ba, Ac134224.1, and Il1b; cluster 2, Atf3, Nrp1, and Fos; and cluster 3, Hba-a2.1, Nrp1, and AC134224.1. The Atf3 gene inhibits the release of inflammatory mediators, suppresses the expression of pro-inflammatory genes, and alleviates inflammation.32 These data indicate that the activation of A1 astrocytes and M1 microglia can mutually promote each other. Microglia represent the main source of TNF-α in the early stage of epilepsy. Henning et al found that TNF-α disrupted the gap junction coupling of astrocytes through tumor necrosis factor receptor-1 on the surface of astrocytes, leading to astrocyte dysfunction and acute epileptic seizures.33 In addition, the signaling pathway mediated by semaphorin 4D (Sema4D) between microglia and astrocytes enhances the inflammatory response in the central nervous system. Clark et al, using barcode virus tracking and scRNA-seq, found that the expression of Sema4D in microglia and Plexin-B2 (PLXNB2) in astrocytes drives the activation of the Sema4D/PLXNB2 pathway at the peak of EAE. They interconnected PLXNB2+/− astrocytes with Sema4D+/− microglia, confirming an increase in signal transduction between PLXNB2 astrocytes and Sema4D microglia. Additionally, they identified a signaling pathway mediated by Ephrin-B receptors during EAE. Ephrin-B receptors are activated through interactions with Ephrin-B1, Ephrin-B2, and Ephrin-B3, which are encoded by Efnb1, Efnb2, and Efnb3, respectively. The sequencing results revealed that Efnb3 expression increased in microglia, whereas no significant changes in Efnb1 and Efnb2 expression were observed. Furthermore, in multiple sclerosis neuropathy, the number of Ephrin-B3–expressing microglia and astrocytes was elevated.34 The role of Ephrin-B3 in promoting the development of refractory epilepsy has been demonstrated.35 Therefore, the Ephrin-B3/Ephrin-B3 receptor signaling pathway in astrocytes and microglia might promote inflammation in the central nervous system.
In summary, microglia and astrocytes can mutually promote inflammation via the C3 system, pro-inflammatory factors such as TNF-α, and the Ephrin-B3/Ephrin-B3 receptor pathway, thereby amplifying neuroinflammatory responses. Interventions targeting these signaling pathways might attenuate the progression of inflammation in the nervous system.
Interactions Between Glial Cells and Neurons in Epilepsy
Transcriptomic analysis of pentylenetetrazol-induced epileptic rat models identified highly expressed genes such as Bad, Ccl2, and Kcnip3, which are associated with neuronal death or neurotransmitter regulation. Beyond direct neuronal death, neuroinflammation also significantly affects neuronal function.36 Fractalkine, also known as chemokine ligand 1 (CX3CL1), is a chemokine expressed by neuronal cells. Its receptor, CX3CR1, is primarily expressed on microglia. This unique distribution makes the CX3CL1/CX3CR1 pathway a crucial mechanism mediating microglia–neuron interactions.37 Previous studies demonstrated that during epileptic seizures, CX3CL1 released by neurons activates microglia, promoting inflammatory responses. The CX3CL1/CX3CR1 signaling cascade in epileptic rats can activate microglia, leading to the release of pro-inflammatory cytokines, exacerbating inflammation and seizures and exerting neurodegenerative effects.38 Zhou et al similarly demonstrated in an epileptic rat model that epilepsy induces microglial activation via the XC3CL1/CX3CR1 axis.39 In a study on Alzheimer’s disease, CX3CL1 was also found to contribute to neuroinflammation.40 However, other studies identified a physical interaction between microglia and neurons in epilepsy termed microglial process convergence (MPC), which exhibits neuroprotective effects. Ssh2 and Fgd3 are predominantly expressed in microglia and are significantly upregulated in epileptic mice. Both of them are key players in the regulation of actin cytoskeleton, suggesting a central role of microglia and immune-related functions in epileptogenesis.41 Among the inflammatory mediators released upon microglial CX3CL1 receptor activation, IL-1β can act on neurons to induce ATP release, thereby promoting the local recruitment of microglia via P2Y12 receptors. CX3CR1 activation is a critical step in the progression of MPC.42 However, previous evidence suggested that CX3CR1 is downregulated in the microglia of patients with epilepsy. Pathological brain tissue cells from patients with drug-resistant epilepsy (DRE) can be divided into 26 clusters. Among them, 13 clusters (0–7, 9–12, and 14) were identified as microglia based on their CD45 expression, with lower CX3CR1 expression noted in clusters 5, 7, 9, and 11. In clusters with higher IL-1β expression, pro-inflammatory cytokines and chemokines were also highly expressed, whereas CX3CR1 expression was decreased compared with that in other clusters.13 Low CX3CR1 expression suppressed MPC, diminished the protective effect of MPC, and exacerbated the inflammatory response.
Astrocytes promote the onset and progression of epilepsy through four key mechanisms: imbalance between excitatory and inhibitory neural signaling, disruption of metabolic homeostasis in the neuronal microenvironment, neuroinflammation, and the formation of abnormal neural circuits.43 A large number of inflammatory mediators are released during inflammation. An increase in TNF-α levels can upregulate aquaporin-4 in astrocytes, resulting in disrupted water transport across the cell membrane, which in turn induces astrocyte edema and reduces glutamate uptake.44 TNF-α can also impair the coupling of gap junctions in astrocytes, leading to their dysfunction. In addition, glutamate transporters are replaced by ionotropic glutamate receptors, especially Glu1R flip, resulting in prolonged depolarization of astrocytes.45 All of these changes reduce the sensitivity of astrocytes to glutamate and their ability to regulate extracellular glutamate concentrations, leading to a higher glutamate concentration in the extracellular environment. Additionally, brain tissues from epilepsy patients shows complex dysregulation of multiple glutamate receptor subunits and neuronal activity-related genes. Most genes encoding glutamate receptor subunits, such as Gria1, Gria3, Gria4, and Grik3, are highly expressed in epilepsy, which has been linked to aberrant hyperexcitability in epileptic neural circuits.46 Meanwhile, increased levels of inflammatory mediators decelerate signal transduction by GABA receptors, leading to a decline in the function of GABAergic inhibitory neurons. Albumin enters the brain parenchyma through the damaged BBB, stimulating astrocytes and impairing their primary buffering function, particularly their ability to buffer K+ released through neurotransmission.47 Under the combined influence of these factors, neuronal excitability increases, facilitating the generation of abnormal discharge activity. Although global neuronal loss and damage are present in the hippocampus of mice with temporal lobe epilepsy, DEG-based analyses indicated that reactive neurogenesis still occurs in this region.8 Synaptic transmission, a fundamental process for nerve impulse propagation, is closely related to astrocyte formation and function.48 During seizures, the signal transduction from astrocytes to neurons increases the likelihood of neurons reaching the seizure threshold. Elevations in Ca2+ levels and glutamate release from astrocytes are associated with the occurrence and maintenance of focal epileptic discharge.49 Astrocytes release gliotransmitters, such as glutamate and serine, to regulate neuronal activity. In the early stages of epileptic seizures, elevated Ca2+ levels in astrocytes promote vesicular transport and neurotransmitter release, thereby activating nearby neurons.50 Adenosine triphosphate (ATP) is an important neurotransmitter released by astrocytes that activates P2 receptors and promotes epileptic seizures. Conversely, adenosine, as a hydrolysis product of ATP, has the opposite effect on the occurrence and development of epilepsy. It exerts anti-convulsant effects by inhibiting neuronal activity.51 Following a seizure, epilepsy triggers a series of tissue inflammatory responses. The NLR family pyrin domain containing 3 inflammasome in astrocytes increases the expression of adenosine kinase,52 accelerating the conversion of adenosine to adenosine monophosphate. The reduction in adenosine levels diminishes its inhibitory effect on neuronal activity, further contributing to epileptic seizures.
Activation of microglial CX3CR1 receptors can exert an opposite effect on neurons through different signaling pathways. Astrocytes influence neuronal excitability by elevating Ca2+ levels, creating a glutamate-rich environment, and reducing adenosine levels. The direct interaction between glial cells and neurons is highly complex and incompletely understood. Different findings from previous studies can be attributed to the complex feedback regulatory mechanisms, which warrant further investigation.
Effects of Immune Cells on BBB in Epilepsy
The central nervous system is often considered an immune-privileged organ, largely because of the absence of a lymphatic system, limited major histocompatibility complex expression, and the restrictive effect of the BBB.53 Fang et al reported a unique type of smooth muscle cells migrating into the brain parenchyma in 15 patients with drug-resistant epilepsy. The migration leads to vascular malformations, disrupts endothelial cell function, and creates a hypoxic microenvironment. Consequently, the HIF-1α/mTOR/S6 pathway is upregulated, impairing neuronal and astrocytic activity and resulting in neuronal loss.54 Damage-associated molecular patterns released following neuronal damage in epilepsy can lead to the substantial release of purinergic substances. P2 purinergic receptors, which are classified into P2X and P2Y subtypes (P2XR and P2YR), are distributed on the surface of microglia. P2XR acts as an ATP-gated ionotropic receptor, and its ligands include P2X4 and P2X7. Meanwhile, P2YR functions as a G protein-coupled metabolic receptor, and its ligands include P2Y6, P2Y12, and P2Y13. These receptors jointly regulate phenotypic changes in microglia.55 P2X7-mediated neuroinflammation plays a crucial role in neurodegenerative diseases, such as multiple sclerosis and status epilepticus.56 Following brain tissue injury, the expression of ATP/adenosine diphosphate-sensitive P2YR, such as P2ry1, P2ry12, and P2ry13 decreases, while that of uridine triphosphate/uridine diphosphate-sensitive receptors such as P2ry2, P2ry4, and P2ry6, increases.57 Elevated levels of purines can activate the P2Y13 receptor, resulting in morphological alternations in microglia, characterized by contraction and increased membrane folding.58 Another important P2Y receptor, P2Y12R, is exclusively expressed in microglia in brain tissues, distinguishing microglia from other peripheral immune cells. Upon activation, P2Y12R promotes microglia remodeling, enhancing their chemotaxis and phagocytic ability, which enables them to migrate to the sites of tissue damage. The migration of microglia facilitates BBB repair, reduces substance leakage from blood vessels into the brain parenchyma, and alleviates inflammation and seizure severity.59 Kumar et al identified a notable characteristic of DRE microglial clusters 7, 5, 9, and 11 using single-cell sequencing of epileptic surgical resection tissues, namely P2Y12R downregulation. Differential gene expression analysis revealed the downregulation of 81 genes and upregulation of 12 genes in P2RY12-expressing microglia, along with higher CX3CR1 expression, compared with the findings in IL-1β–expressing pro-inflammatory microglia.13 The functional and structural changes in P2RY12-expressing microglia play a protective role in epilepsy. Along with the aforementioned evidence, these findings confirm that the absence of P2Y12R exacerbates epileptic seizures.
The expression of CDC25B, DNMT1, GZMA, MTX1, and SSH2 is associated with a reduced risk of epilepsy, whereas FGD3, RAF1, and SH3BP5L are correlated with an increased risk.41 Epilepsy patients exhibited an altered peripheral immune profile, notably with increased activation of immune cells.8 Infiltration of monocytes is often observed in epileptic brain tissues. As a chemokine, chemokine C-C motif ligand 2 (CCL2) specifically activates and attracts monocytes through the C-C motif chemokine receptor 2 (CCR2) signaling. Upregulation of IL-8 and CCL2 in the hippocampus of patients with mesial temporal lobe epilepsy is associated with hippocampal sclerosis.60 This pattern differs from the significant upregulation of interferon-mediated pathway-related genes in the microglia of patients with Rasmussen’s encephalitis.17 Additionally, CCL2 levels are increased during LPS-induced seizure enhancement. Antagonism of CCL2/CCR2 signaling effectively reduces inflammation-induced seizures.61 Using scRNA-seq of microglia, Kumar et al detected the expression of pro-inflammatory cytokines (eg, IL-1β, TNF, CCL2, CCL4) and chemokine genes in patients with DRE, whereas microglia from control groups did not express these factors.13 CCL2 is significantly upregulated in the brains of patients with epilepsy,62 facilitating monocyte infiltration into the brain. In addition to directly inducing monocyte chemotaxis, CCL2 also disrupts the BBB by increasing its permeability. The binding of CCL2 to CCR2 on endothelial cells weakens intercellular connections between endothelial cells, compromising the integrity of the BBB.63 Infiltrating monocytes also contribute to BBB damage, forming a feedback loop. Specifically, monocytes facilitate CCL2-mediated BBB disruption, which in turn can increase CCL2 release from the BBB, attracting more monocytes and further increasing BBB permeability. This evidence supports that abnormal P2Y12R expression and CCL release from microglia in epilepsy affect BBB integrity and function. As an important component that regulates the communication between brain parenchyma cells and peripheral immunoreactive cells, the BBB plays a critical role in the pathological progression of epilepsy. Therefore, targeting BBB dysfunction might represent a new antiepileptic strategy for epilepsy.
Direct Interaction Between T Cells and Microglia in Epilepsy
Autoreactive T cells and autoantibodies are contributors to neuroinflammation. Previous studies indicated the presence of T cell infiltration in the brain tissue of patients with epilepsy.64 In patients with Rasmussen’s encephalitis and focal cortical dysplasia, genes highly expressed in microglia have been found to mediate T cell activation.17 Kumar et al identified infiltrating CD4+ and CD8+ T cells as the most abundant immune cell subtypes in brain samples from patients with epilepsy. They distinguished 16 immune cell clusters, with most CD4+ and CD8+ T cells expressing pro-inflammatory genes, such as CCL4, CCL5, TNF, IL1B, and IFNG. However, there were also some immature cell clusters. Genes expressed in cluster 6 (ie, CCR7, LEF1, SOX4, RPL9, RPS6) are characteristic of immature and proliferative CD4+ T cells, whereas immature CD8+ T cells express CD2, THEMIS, IKZF1, KLRC1, ATM, GBP5, and RAC2. They also identified a mixed bimodal cluster (cluster 3) with simultaneous expression of the surface markers CD3, CD11b, and HLA-DR, indicating a bimodal pattern involving T cells, microglia, and/or macrophages. Further analysis of this bimodal population enabled the isolation of key immune cell subtypes that directly interact with microglia. CD8+ and CD4+ T cell clusters are the two primary clusters in bimodal cells, highlighting a direct physical interaction between microglia and T cells that might amplify their pro-inflammatory and cytotoxic effects.13 Westfall et al observed increased expression of microglial T cell activation markers (ie CD40, CD80), which were physically localized near distinct populations of CD4+ and CD8+ lymphocytes.17 These data illustrate that T cells are active in inflammatory responses in epilepsy, and they can enhance these responses by interacting with microglia.
Conclusion
Analyzing cell development and differentiation in epilepsy-related inflammation provides new insights into potential therapeutic approaches for this disease. Using scRNA-seq, we can identify various types of cells in epilepsy, exploring unique developmental profiles, gene expression patterns, and cell interaction pathways. Investigating the differentiation of microglia, astrocytes, and neurons through gene expression profiling can clarify their developmental trajectories in epilepsy. DEG analysis can improve our understanding of transcriptional dynamics in epilepsy. As cell interactions are fundamental to both physiological and pathological processes, mapping these interactions at the single-cell level offers a comprehensive view of cellular interplay in epileptic inflammation (Figure 1). Exploring the mechanisms of epilepsy at the single-cell level holds promise for identifying novel therapeutic strategies. This methodology is rapidly advancing toward greater spatial resolution, omics integration, and dynamic analysis. However, this approach has multiple limitations, including reduced sensitivity to low-abundance transcripts, limited availability and heterogeneity of human brain samples, and gaps between data interpretation and functional validation. Additional research is needed to address these areas.
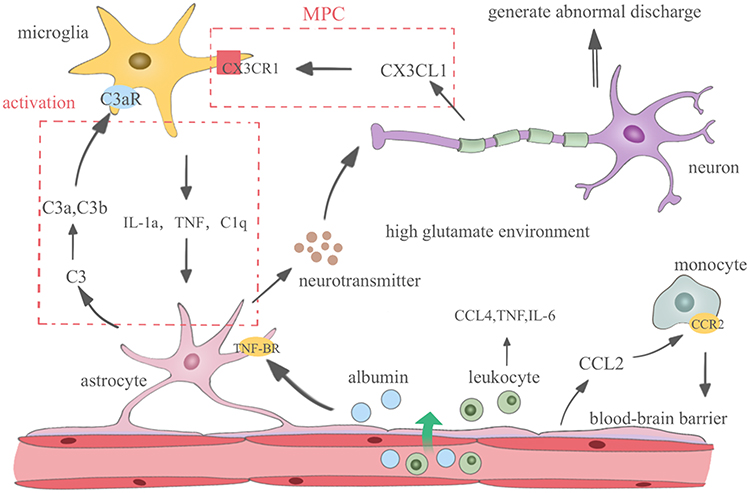 |
Figure 1 The inflammatory response in epilepsy involves various cell types in the central nervous system. Activated microglia release inflammatory mediators, such as IL-1α, TNF, and component C1q, whereas leukocytes can release CCL4, TNF, and IL-6, triggering a cascade of inflammatory responses. Microglia and astrocytes mutually activate each other via the C3 complement system, whereas microglia interact with neurons through MPC mediated by the CX3CL1/CX3CR1 pathway. Astrocytes accumulate albumin through TGF-β receptors and subsequently release additional IL-6. The damaged endothelium releases CCL2, attracting monocytes and further disrupting the blood–brain barrier. Astrocytes release large amounts of neurotransmitters, such as glutamate and serine, creating a high extracellular glutamate environment. Inflammatory mediators, such as IL-1α, function with neurons to induce neuronal ictal discharge. Inflammation in epilepsy leads to seizures and neuronal damage, which in turn promotes inflammation. Abbreviations: IL-1α, interleukin-1 alpha; TNF, tumor necrosis factor; CCL4, C-C chemokine ligand 4; IL-6, interleukin-6; MPC, microglial process convergence; CX3CL1, chemokine ligand 1; CX3CR1, chemokine ligand 1 receptor; TNF-β, tumor necrosis factor beta; CCL2, C-C chemokine ligand 2; C3, component 3; C3a, component 3a; C3b, component 3b; C3aR, component 3a receptor. |
Data Sharing Statement
Data availability is not applicable as this study did not generate or analyze new data.
Acknowledgments
We thank Medjaden Inc. for scientific editing of this paper.
Author Contributions
Yutong Chen: Conceptualization, Writing - original draft, Writing - review & editing; Wei Zhang, Meiying Xin and Jianmin Liang: Data curation, Investigation, Funding acquisition, Writing - review & editing; Ming Liu, Tongye Zhang and Jinyu Xiao: Data curation, Investigation, Writing - review & editing; Xuemei Wu: Conceptualization, Funding acquisition, Project administration, Supervision, Writing - review & editing. All authors gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This study was supported by the National Natural Science Foundation of China (Nos. 81801284, 82101599 and 82201606) and the National Natural Science Foundation of Jilin province (YDZJ202201ZYTS091).
Disclosure
The authors have no conflicts of interest to declare.
References
1. Ali A. Global health: epilepsy. Semin Neurol. 2018;38(02):191–10. doi:10.1055/s-0038-1646947
2. Zhang LM, Zeng T, Zhang BR, et al. Mendelian randomization combined with single-cell sequencing data analysis of chemokines and chemokine receptors and key genes and molecular mechanisms associated with epilepsy. Neuroreport. 2025;36(9):467–486. doi:10.1097/WNR.0000000000002168
3. Bartolini L, Libbey JE, Ravizza T, Fujinami RS, Jacobson S, Gaillard WD. Viral triggers and inflammatory mechanisms in pediatric epilepsy. Mol Neurobiol. 2018;56(3):1897–1907. doi:10.1007/s12035-018-1215-5
4. Candelario-Jalil E, Dijkhuizen RM, Magnus T. Neuroinflammation, stroke, blood-brain barrier dysfunction, and imaging modalities. Stroke. 2022;53(5):1473–1486. doi:10.1161/STROKEAHA.122.036946
5. Mesraoua B, Brigo F, Lattanzi S, Abou-Khalil B, Al Hail H, Asadi-Pooya AA. Drug-resistant epilepsy: definition, pathophysiology, and management. J Neurol Sci. 2023;452:120766. doi:10.1016/j.jns.2023.120766
6. Khoshkhoo S, Lal D, Walsh CA. Application of single cell genomics to focal epilepsies: a call to action. Brain Pathol. 2021;31(4):e12958. doi:10.1111/bpa.12958
7. Wang M, Gu M, Liu L, Liu Y, Tian L. Single-cell RNA sequencing (scRNA-seq) in cardiac tissue: applications and limitations. Vasc Health Risk Manag. 2021;17:641–657. doi:10.2147/VHRM.S288090
8. Liu Q, Shen C, Dai Y, et al. Single-cell, single-nucleus and xenium-based spatial transcriptomics analyses reveal inflammatory activation and altered cell interactions in the hippocampus in mice with temporal lobe epilepsy. Biomarker Res. 2024;12(1). doi:10.1186/s40364-024-00636-3
9. Borst K, Dumas AA, Prinz M. Microglia: immune and non-immune functions. Immunity. 2021;54(10):2194–2208. doi:10.1016/j.immuni.2021.09.014
10. Chiu IM, Morimoto ET, Goodarzi H, et al. A neurodegeneration-specific gene-expression signature of acutely isolated microglia from an amyotrophic lateral sclerosis mouse model. Cell Rep. 2013;4(2):385–401. doi:10.1016/j.celrep.2013.06.018
11. Keren-Shaul H, Spinrad A, Weiner A, et al. A unique microglia type associated with restricting development of Alzheimer’s disease. Cell. 2017;169(7):1276–1290e1217. doi:10.1016/j.cell.2017.05.018
12. Sousa C, Golebiewska A, Poovathingal SK, et al. Single-cell transcriptomics reveals distinct inflammation-induced microglia signatures. EMBO Rep. 2018;19(11). doi:10.15252/embr.201846171
13. Kumar P, Lim A, Hazirah SN, et al. Single-cell transcriptomics and surface epitope detection in human brain epileptic lesions identifies pro-inflammatory signaling. Nat Neurosci. 2022;25(7):956–966. doi:10.1038/s41593-022-01095-5
14. Terrone G, Balosso S, Pauletti A, Ravizza T, Vezzani A. Inflammation and reactive oxygen species as disease modifiers in epilepsy. Neuropharmacology. 2020;167:107742. doi:10.1016/j.neuropharm.2019.107742
15. Araki T, Ikegaya Y, Koyama R. The effects of microglia- and astrocyte-derived factors on neurogenesis in health and disease. Eur J Neurosci. 2021;54(5):5880–5901. doi:10.1111/ejn.14969
16. Cao J, Gan H, Xiao H, et al. Key protein-coding genes related to microglia in immune regulation and inflammatory response induced by epilepsy. Math Biosci Eng. 2021;18(6):9563–9578. doi:10.3934/mbe.2021469
17. Westfall JJ, Schwind WN, Sran S, et al. Molecular and spatial heterogeneity of microglia in Rasmussen encephalitis. Acta Neuropathologica Commun. 2022;10(1). doi:10.1186/s40478-022-01472-y
18. Zhou Q, Lin L, Li H, et al. Melatonin reduces neuroinflammation and improves axonal hypomyelination by modulating M1/M2 microglia polarization via JAK2-STAT3-telomerase pathway in postnatal rats exposed to lipopolysaccharide. Mol Neurobiol. 2021;58(12):6552–6576. doi:10.1007/s12035-021-02568-7
19. Wang C, Li Z, Zhu X, et al. Ultrabright ratiometric Raman-guided epilepsy surgery by intraoperatively visualizing proinflammatory microglia. Cell Rep Med. 2025;6.
20. Yu C, Deng XJ, Xu D. Microglia in epilepsy. Neurobiol Dis. 2023;185:106249. doi:10.1016/j.nbd.2023.106249
21. Wheeler MA, Clark IC, Tjon EC, et al. MAFG-driven astrocytes promote CNS inflammation. Nature. 2020;578(7796):593–599. doi:10.1038/s41586-020-1999-0
22. Sanz P, Garcia-Gimeno MA. Reactive glia inflammatory signaling pathways and epilepsy. Int J Mol Sci. 2020;21(11):4096. doi:10.3390/ijms21114096
23. Fan YY, Huo J. A1/A2 astrocytes in central nervous system injuries and diseases: angels or devils? Neurochem Int. 2021;148:105080. doi:10.1016/j.neuint.2021.105080
24. Qian Z, Qin J, Lai Y, Zhang C, Zhang X. Large-scale integration of single-cell RNA-seq data reveals astrocyte diversity and transcriptomic modules across six central nervous system disorders. Biomolecules. 2023;13(4):692. doi:10.3390/biom13040692
25. Cameron EG, Nahmou M, Toth AB, et al. A molecular switch for neuroprotective astrocyte reactivity. Nature. 2023;626(7999):574–582. doi:10.1038/s41586-023-06935-3
26. Liu LR, Liu JC, Bao JS, Bai QQ, Wang GQ. Interaction of microglia and astrocytes in the neurovascular unit. Front Immunol. 2020;11:1024. doi:10.3389/fimmu.2020.01024
27. Li X, Zhou R, Peng H, Peng J, Li Q, Mei M. Microglia PKM2 mediates neuroinflammation and neuron loss in mice epilepsy through the astrocyte C3-neuron C3R signaling pathway. Brain Sci. 2023;13:262.
28. Liddelow SA, Guttenplan KA, Clarke LE, et al. Neurotoxic reactive astrocytes are induced by activated microglia. Nature. 2017;541(7638):481–487. doi:10.1038/nature21029
29. Gnanaguru G, Tabor SJ, Bonilla GM, et al. Microglia refine developing retinal astrocytic and vascular networks through the complement C3/C3aR axis. Development. 2023;150(5). doi:10.1242/dev.201047
30. Zhang W, Ding L, Chen H, et al. Cntnap4 partial deficiency exacerbates alpha-synuclein pathology through astrocyte-microglia C3-C3aR pathway. Cell Death Dis. 2023;14(4):285. doi:10.1038/s41419-023-05807-y
31. Zhu A, Cui H, Su W, Liu C, Yu X, Huang Y. C3aR in astrocytes mediates post-thoracotomy pain by inducing A1 astrocytes in male rats. Biochim Biophys Acta Mol Basis Dis. 2023;1869(5):166672. doi:10.1016/j.bbadis.2023.166672
32. Wieczorek-Szukala K, Markiewicz M, Walczewska A, Zgorzynska E. Docosahexaenoic acid (DHA) reduces LPS-induced inflammatory response via ATF3 transcription factor and stimulates Src/Syk signaling-dependent phagocytosis in microglia. Cell Physiol Biochem. 2023;57:411–425.
33. Henning L, Antony H, Breuer A, et al. Reactive microglia are the major source of tumor necrosis factor alpha and contribute to astrocyte dysfunction and acute seizures in experimental temporal lobe epilepsy. Glia. 2023;71(2):168–186. doi:10.1002/glia.24265
34. Clark IC, Gutierrez-Vazquez C, Wheeler MA, et al. Barcoded viral tracing of single-cell interactions in central nervous system inflammation. Science. 2021;372(6540). doi:10.1126/science.abf1230
35. Huang H, Li R, Yuan J, et al. Up-regulated ephrinB3/EphB3 expression in intractable temporal lobe epilepsy patients and pilocarpine induced experimental epilepsy rat model. Brain Res. 2016;1639:1–12. doi:10.1016/j.brainres.2016.02.035
36. Li X, Wang Q, Zhang D-W, et al. Hippocampus RNA sequencing of pentylenetetrazole-kindled rats and upon treatment of novel chemical Q808. Front Pharmacol. 2022;13:820508.
37. Mecca C, Giambanco I, Donato R, Arcuri C. Microglia and aging: the role of the TREM2–DAP12 and CX3CL1-CX3CR1 axes. Int J Mol Sci. 2018;19(1):318. doi:10.3390/ijms19010318
38. Yeo SI, Kim JE, Ryu HJ, et al. The roles of fractalkine/CX3CR1 system in neuronal death following pilocarpine-induced status epilepticus. J Neuroimmunol. 2011;234(1–2):93–102. doi:10.1016/j.jneuroim.2011.03.005
39. Zhou Y, Zhang L, Hao Y, Yang L, Fan S, Xiao Z. FKN/CX3CR1 axis facilitates migraine-Like behaviour by activating thalamic-cortical network microglia in status epilepticus model rats. J Headache Pain. 2022;23(1):42. doi:10.1186/s10194-022-01416-w
40. Liu PP, Liu XH, Ren MJ, et al. Neuronal cathepsin S increases neuroinflammation and causes cognitive decline via CX3CL1-CX3CR1 axis and JAK2-STAT3 pathway in aging and Alzheimer’s disease. Aging Cell. 2025;24(2):e14393. doi:10.1111/acel.14393
41. Shi J, Xie J, Yang Y, et al. Integrated Mendelian randomization and single-cell transcriptomics analysis identifies critical blood biomarkers and potential mechanisms in epilepsy. CNS Neurosci Ther. 2025;31(1):e70172. doi:10.1111/cns.70172
42. Eyo UB, Peng J, Murugan M, et al. Regulation of physical microglia-neuron interactions by fractalkine signaling after status epilepticus. eNeuro. 2016;3(6):
43. Chen Y, Hu J, Zhang Y, et al. Epilepsy therapy beyond neurons: unveiling astrocytes as cellular targets. Neural Regeneration Res. 2026;21(1):23–38. doi:10.4103/NRR.NRR-D-24-01035
44. Lu H, Ai L, Zhang B. TNF-alpha induces AQP4 overexpression in astrocytes through the NF-kappaB pathway causing cellular edema and apoptosis. Biosci Rep. 2022;42(3). doi:10.1042/BSR20212224
45. Seifert G, Huttmann K, Schramm J, Steinhauser C. Enhanced relative expression of glutamate receptor 1 flip AMPA receptor subunits in hippocampal astrocytes of epilepsy patients with Ammon’s horn sclerosis. J Neurosci. 2004;24(8):1996–2003. doi:10.1523/JNEUROSCI.3904-03.2004
46. Pfisterer U, Petukhov V, Demharter S, et al. Identification of epilepsy-associated neuronal subtypes and gene expression underlying epileptogenesis. Nat Commun. 2020;11(1):5038. doi:10.1038/s41467-020-18752-7
47. Vezzani A, Ravizza T, Bedner P, Aronica E, Steinhauser C, Boison D. Astrocytes in the initiation and progression of epilepsy. Nat Rev Neurol. 2022;18(12):707–722. doi:10.1038/s41582-022-00727-5
48. Allen NJ, Eroglu C. Cell biology of astrocyte-synapse interactions. Neuron. 2017;96(3):697–708. doi:10.1016/j.neuron.2017.09.056
49. Gomez-Gonzalo M, Losi G, Chiavegato A, et al. An excitatory loop with astrocytes contributes to drive neurons to seizure threshold. PLoS Biol. 2010;8(4):e1000352. doi:10.1371/journal.pbio.1000352
50. Bohmbach K, Schwarz MK, Schoch S, Henneberger C. The structural and functional evidence for vesicular release from astrocytes in situ. Brain Res Bull. 2018;136:65–75. doi:10.1016/j.brainresbull.2017.01.015
51. Beamer E, Kuchukulla M, Boison D, Engel T. ATP and adenosine-two players in the control of seizures and epilepsy development. Prog Neurobiol. 2021;204:102105. doi:10.1016/j.pneurobio.2021.102105
52. Zhang H, Yu S, Xia L, Peng X, Wang S, Yao B. NLRP3 inflammasome activation enhances ADK expression to accelerate epilepsy in mice. Neurochem Res. 2022;47(3):713–722. doi:10.1007/s11064-021-03479-8
53. Huang X, Hussain B, Chang J. Peripheral inflammation and blood–brain barrier disruption: effects and mechanisms. CNS Neurosci Ther. 2020;27(1):36–47. doi:10.1111/cns.13569
54. Fang C, Zhang X, Yang L, et al. Transcriptomic and morphologic vascular aberrations underlying FCDIIb etiology. Nat Commun. 2025;16(1):3320. doi:10.1038/s41467-025-58535-6
55. Calovi S, Mut-Arbona P, Sperlagh B. Microglia and the purinergic signaling system. Neuroscience. 2019;405:137–147. doi:10.1016/j.neuroscience.2018.12.021
56. Ahn YH, Tang Y, Illes P. The neuroinflammatory astrocytic P2X7 receptor: alzheimer’s disease, ischemic brain injury, and epileptic state. Expert Opin Ther Targets. 2023;27(9):763–778. doi:10.1080/14728222.2023.2258281
57. Alves M, Gomez-Villafuertes R, Delanty N, et al. Expression and function of the metabotropic purinergic P2Y receptor family in experimental seizure models and patients with drug-refractory epilepsy. Epilepsia. 2017;58(9):1603–1614. doi:10.1111/epi.13850
58. Milior G, Morin-Brureau M, Chali F, et al. Distinct P2Y receptors mediate extension and retraction of microglial processes in epileptic and peritumoral human tissue. J Neurosci. 2020;40(7):1373–1388. doi:10.1523/JNEUROSCI.0218-19.2019
59. Das R, Chinnathambi S. Microglial remodeling of actin network by Tau oligomers, via G protein-coupled purinergic receptor, P2Y12R -driven chemotaxis. Traffic. 2021;22(5):153–170. doi:10.1111/tra.12784
60. Aulicka S, Ceska K, Sana J, et al. Cytokine-chemokine profiles in the hippocampus of patients with mesial temporal lobe epilepsy and hippocampal sclerosis. Epilepsy Res. 2022;180:106858. doi:10.1016/j.eplepsyres.2022.106858
61. Cerri C, Genovesi S, Allegra M, et al. The chemokine CCL2 mediates the seizure-enhancing effects of systemic inflammation. J Neurosci. 2016;36(13):3777–3788. doi:10.1523/JNEUROSCI.0451-15.2016
62. Broekaart DWM, Anink JJ, Baayen JC, et al. Activation of the innate immune system is evident throughout epileptogenesis and is associated with blood-brain barrier dysfunction and seizure progression. Epilepsia. 2018;59(10):1931–1944. doi:10.1111/epi.14550
63. Stamatovic SM, Dimitrijevic OB, Keep RF, Andjelkovic AV. Protein kinase Calpha-RhoA cross-talk in CCL2-induced alterations in brain endothelial permeability. J Biol Chem. 2006;281(13):8379–8388. doi:10.1074/jbc.M513122200
64. Fornari Caprara AL, Rissardo JP, Nagele EP. Rasmussen encephalitis: clinical features, pathophysiology, and management strategies—a comprehensive literature review. Medicina. 2024;60(11):1858. doi:10.3390/medicina60111858
 © 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2026 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
Recommended articles
Complex Neuroimmune Involvement in Neurodevelopment: A Mini-Review
Monet MC, Quan N
Journal of Inflammation Research 2023, 16:2979-2991
Published Date: 19 July 2023
Ligusticum chuanxiong Hort. Ameliorates Neuropathic Pain by Regulating Microglial M1 Polarization: A Study Based on Network Pharmacology
Cui S, Feng X, Xia Z
Journal of Pain Research 2024, 17:1881-1901
Published Date: 23 May 2024
Single-Cell RNA Sequencing Reveals That C5AR1 in Follicle Monocyte Cells Could Predict the Development of POI

Han Y, Diao J, Wang X, Zhang S, Yuan L, Ping Y, Zhang Y, Luo H
Journal of Inflammation Research 2024, 17:11221-11234
Published Date: 18 December 2024
Integrating Bulk and Single-Cell Transcriptomic Data to Identify Ferroptosis-Associated Inflammatory Gene in Alzheimer’s Disease
Zhou H, Peng Y, Huo X, Li B, Liu H, Wang J, Zhang G
Journal of Inflammation Research 2025, 18:2105-2122
Published Date: 10 February 2025
Therapeutic Potential of TPT-260 in Ischemic Stroke: An Investigation Into Its Anti-Inflammatory Effects and Impact on Microglial Activation
Qian J, Guo X, Xu Q, Huang Z
Journal of Inflammation Research 2025, 18:3055-3066
Published Date: 1 March 2025