")
Back to Journals » Infection and Drug Resistance » Volume 15
An Excretory Protein of Echinococcus multilocularis Inhibits Complement Classical Pathway Activation
Authors Qiu Y , Shen S, Yang Y, Wang W
Received 15 November 2021
Accepted for publication 7 February 2022
Published 22 February 2022 Volume 2022:15 Pages 555—568
DOI https://doi.org/10.2147/IDR.S344075
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 3
Editor who approved publication: Professor Suresh Antony
Yiwen Qiu, Shu Shen, Yi Yang, Wentao Wang
Department of Liver Surgery and Liver Transplantation Center, West China Hospital of Sichuan University, Chengdu, People’s Republic of China
Correspondence: Wentao Wang, Department of Liver Surgery and Liver Transplantation Center, West China Hospital of Sichuan University, 37 Guoxue Road, Chengdu, 610041, People’s Republic of China, Tel +86 18980601895, Fax +86-028-85422871, Email [email protected]
Introduction: Alveolar echinococcosis is a lethal zoonosis caused by Echinococcus multilocularis (E.m) larvae. The mechanism by which E.m evades host immune attacks and ensures long-term survival remains unexplained. The complement system is a cascade of sequentially activated complement proteins that results in opsonization-related phagocytosis or membrane lysis of invading organisms. Excretory/secretory proteins (ESPs) of parasites are the main antigens that induce the immune response and play important roles in the long-term survival.
Methods: We investigated the possibility that E.m inhibits complement activation through ESPs and examined the potential related mechanism. A haemolysis assay was used to determine if and how in vitro culture medium of E.m containing ESPs can inhibit complement activation. Potential ESPs were annotated using bioinformatics methods, and one ESP was subsequently expressed as a recombinant protein with a eukaryotic expression system. The ability of this protein to inhibit complement activation was also tested by haemolysis assay.
Results: These assays showed that in vitro culture medium of E.m inhibited activation of the complement classical pathway. EmuJ_000439500 encodes a protein containing seven Sushi domains, which was the only potential E.m-derived complement inhibitor (Em-CI, UniProt: A0A068Y4F2) annotated among the 653 ESPs. Recombinant Em-CI also displayed the ability to inhibit activation of the complement classical pathway.
Discussion: The discovery of Em-CI sheds light on the mechanism by which E.m escapes killing by the complement system and provides potential targets for immunotherapy for parasitic diseases.
Keywords: Echinococcus multilocularis, alveolar echinococcosis, complement inhibition, excretory/secretory protein
Introduction
Alveolar echinococcosis (AE) is a lethal parasitic disease primarily characterized by tumor-like hepatic infiltration caused by the continuous proliferation of Echinococcus multilocularis (E.m) metacestodes.1 Symptoms including jaundice and abdominal pain are mostly the results of lesions invading both the biliary and vascular structures. Frequent infiltration of adjacent or metastases into distant tissues/organs exacerbates the disease.2
The parasitic tissue comprises plentiful tiny, irregular cavities, which are parasitic vesicles containing the actively proliferating parasites. The border of the invasive parasitic tissue is not clearly outlined from the adjacent normal liver parenchyma according to macroscopic examinations because the proliferative behavior of the parasitic tissue is partly contained by the host periparasitic fibro-inflammatory response. The central part of the AE lesion that consists mainly of fibrous tissue is commonly liquefied or calcified through necrosis due to the deficiency of vascularization and occasional bacterial infection.3
The natural life cycle of E.m can occasionally involve humans as intermediate hosts, which become infected through ingestion of tapeworm eggs and subsequently develop into metacestodes (primary infection). Specifically, the E.m oncosphere is released into the intestine and then penetrates the intestinal wall. Before migrating into the liver via portal vein blood, the oncosphere must avoid destruction by the complement system. Moreover, E.m must prevent itself from being attacked by the complement system during the development of the oncosphere into the metacestode in the liver and subsequently chronically infects the host.
The complement system, which is the first line of defence against infestor, involves a cascade of sequentially activated complement proteins that results in opsonization-related phagocytosis or membrane lysis of the invading pathogen.4 Activation of the complement system is triggered via the classical pathway (CP), the lectin pathway (LP), or the alternative pathway (AP). The CP is prompted by antibody-antigen complexes.5 The LP is initiated by the recognition of non-self carbohydrates.6 The AP is continuously active motivated by low-level background enzyme activation.7,8 Complement activation defects and dysfunction are closely related to the development of various infections including Taenia taeniaeformis, Necator americanus, Schistosoma mansoni, etc.9,10
Amplification of complement activation is prevented by a family of proteins called complement regulators. C4b-binding protein (C4BP) is one of the significant complement regulators. This 500 kDa protein can specifically inhibit the action of the CP and LP by interacting with C4 and C3b to act as a key soluble regulator. The function of C4BP is accelerating the decay of C3-convertase and cofactor ability of the serine protease factor I, which cleaves C4b and C3b.11 The complement regulators have recently been found to be characterized as Sushi domain-containing (SUSD) protein family and Human CUB and Sushi multiple domains (CSMD) family, which share homologous structures with known complement inhibitors and modulate complement activity.12
Previous research on schistosomiasis13 and malaria14 has demonstrated that the corresponding parasites interfere with the complement system through a variety of mechanisms, leading to diminished local inflammatory responses and progression into chronic infectious diseases. During Echinococcus granulosus (E.g) infection, the complement system is activated via the AP, and the protoscolex is thereby lysed to control parasitic infection.15,16 However, parasites can evade immune attacks by secreting molecules that regulate the complement system.16 Recently, Ferreira et al found that mice deficient in C5 are more susceptible to E.g infection and exhibit more severe lesions than wild-type mice.17
Excretory/secretory proteins (ESPs) are antigenic substances excreted by parasites. ESPs are directly exposed to the host’s immune system and are the main antigens that induce host immune responses. Moreover, ESPs play vital roles in the long-term survival of parasites within hosts. Recognized as virulence factors, ESPs also function as immunomodulators/inhibitors to regulate local and systemic immune responses, inhibit inflammation, and mediate immune escape, thus creating a microenvironment where parasites can survive for a long time.9,18 Indeed, previous studies on parasites, such as nematodes, trematodes, and tapeworms (including E.m and E.g), have elucidated that these parasites can impair the host immune system and promote their own long-term survival by producing ESPs.10
Despite the significant roles of ESPs produced by other helminths in evasive mechanisms, the functions of ESPs in E.m infection remain undefined. Thus, we aimed to decode the underlying mechanisms of ESPs in E.m-related disease. In the present study, we discovered, for the first time, that in vitro culture medium (CM) from E.m protoscoleces is able to inhibit complement activation. Through bioinformatics analysis, we predicted that E.m ESP molecules have structures similar to those of known complement regulators. Intriguingly, a recombinant ESP produced in a yeast expression system could inhibit complement activation. To our knowledge, this is the first evidence that complement inhibitors are present among E.m-secreted proteins.
Methods
Ethics Statement
All protocols concerning animal experiments were approved by the laboratory animal welfare and ethics committee, West China Hospital, Sichuan University (permit number: 2021275A), in accordance with the ethical guidelines for the care and use of laboratory animals of the National Institute of Parasitic Diseases, Chinese Center for Disease Control and Prevention. Human serum was obtained from the author and performed in accordance with the 1975 Declaration of Helsinki. The study was approved by the ethics committee of West China Hospital of Sichuan University (permit number: 201738), and informed consent was provided.
Isolation and Culture of E.m Protoscoleces
E.m was propagated in Mongolian jirds (Meriones unguiculatus) as described previously.33 To isolate protoscoleces, parasite-containing tissues were procured from infected jirds and homogenized as described by Brehm.34 The homogenate was subsequently filtered through a nylon mesh with a 100 µm pore size to remove larger debris. Sequentially, the filtrate was passed through a nylon mesh with a 40 µm pore size to isolate the protoscoleces from the cellular components. The protoscoleces were washed off from the meshes with sterile PBS containing 100 U/mL penicillin and 100 μg/mL streptomycin (PBS-PS). The activity of protoscoleces was determined by 0.1% eosin staining, with dead protoscoleces staining red.
In vitro culture of E.m protoscoleces was described previously.19 Protoscoleces with ≥95% activity were cultivated in RPMI 1640 (Gibco, Auckland, New Zealand) containing 0.45% (w/v) yeast extract, 0.4% (w/v) glucose, 25% (v/v) foetal bovine serum (Gibco, heated under 56°C for 30 min to inactivate complement), 100 U/mL penicillin and 100 μg/mL streptomycin in a flask at 37°C in 5% CO2. The medium was changed every 3 days. A volume of 2 mL of in vitro CM of E.m was obtained every day for subsequent haemolytic assay, and the culture flask was replenished with an equal volume of fresh medium.
Preparation of Sera and Erythrocytes
NHS was prepared from the peripheral blood of healthy donors. The blood was kept at room temperature for 30 min to coagulate, placed in ice for 60 min, and then centrifuged for 7 min at 700 × g. The serum fraction was collected and stored at −80°C until use.
REs were purchased from Solarbio (Beijing, China). Using Mg-ethylene glycol tetraacetic acid (EGTA) buffer (2075 mM veronal buffer pH 7.3, 10 mM EGTA, 7 mM MgCl2, 0.083% gelatin, 116 mM glucose, 60 mM NaCl), REs (1.6 × 109, corresponding to 2 mL of stock solution) were washed until there was no visible haemoglobin in the supernatant. Then, 1.6 × 109 REs were suspended in 50 μL of Mg-EGTA buffer, and the suspension was used as the stock for the assays.
ShEs were purchased from Solarbio. Using a dextrose gelatin veronal buffer (DGVB) buffer (2.5 mM veronal buffer, pH 7.3, 72 mM NaCl, 140 mM glucose, 0.1% gelatin, 1 mM MgCl2, and 0.15 mM CaCl2), ShEs (4 × 108, corresponding to 1 mL of stock solution) were washed, pelleted and mixed with 2 mL of DGVB containing amboceptor (Solarbio, Beijing) and diluted 1:200. The mixture was then incubated for 10 min at 37°C with shaking. Subsequently, we washed the sensitized ShEs twice and resuspended in 500 μL of DGVB, and the suspension was used as the stock for the assays.
CM Haemolytic Assay
To determine whether CM can inhibit activation of the complement AP or CP, REs or ShEs at different concentrations (corresponding to 50 µL) were incubated in 50 μL of buffers permissive for a given pathway (DGVB or Mg-EGTA) for 30 min in the presence of 50 μL of HI-CM (96°C, 30 min) and 50 μL of NHS at different concentrations. Aliquots of 150 µL of 0.5% Triton X-100 and buffer were used as positive and negative controls, respectively. Thereafter, the erythrocytes were centrifuged (2000 × g, 10 min), and the released haemoglobin was measured at 541 nm using a microplate reader (Thermo Scientific). The haemolysis rate was determined by the average absorbance value. The absorbance value of each sample was calculated as follows: (sample value – negative control group value)/(positive control value – negative control group value). All samples were tested in quintuplicate.
Bioinformatics Analysis of E.m ESPs
The protein sequences of E.m were extracted from the GeneDB website. We first used TMHMM 2.0 to predict the transmembrane domains of all the E.m protein sequences and removed proteins with multiple transmembrane domains.34,35 Phobius was further utilized to exclude proteins with single transmembrane domains.35 SignalP 5.0 analysis (selected organism group: “Eukarya”) was conducted to filter classical ESPs containing signal peptides at their N-termini from proteins that did not cover any transmembrane regions.36 For proteins without signal peptides, SecretomeP (with the “Mammalian” setting) was used to screen out non-classical ESPs containing no N-terminal signal peptides. Target (with the “Non-plant” setting) and ScanProsite (Prosite: PS00014) were employed to filter out classical and non-classical ESPs with subcellular localizations, excluding mitochondrial proteins and endoplasmic reticulum proteins.25 Finally, PredGPI (omega-site prediction mode, “General model” setting, specificity >99%) was used to exclude glycosylphosphatidylinositol-anchored proteins.
The sequences of E.m ESPs were uploaded to the KEGG Automatic Annotation Server (KAAS) and searched by BLAST using the single-directional best hit (SBH) assignment method. Schistosoma mansoni was selected as the GENES data set. OmicsBox (Blast2GO) was used to identify the protein domains, families, and functional sites of the ESPs. Finally, a protein–protein interaction (PPI) network of the ESPs was created with STRING. EmuJ_000439500, which encodes an ESP (UniProt: A0A068Y4F2) that contains multiple Sushi domains, was the Em-CI predicted with the highest confidence.
Quantitative Real-Time PCR
RNA was extracted and purified from metacestode tissue freshly harvested from 6 infected and three healthy jirds using an RNeasy Mini Kit (Qiagen, Hombrechtikon, Switzerland) according to the standard protocol. cDNA synthesis was performed using an Omniscript Reverse Transcription Kit (Qiagen). EmuJ_000439500 mRNA expression was analysed. Quantitative real-time PCR was performed using FastStart SYBR Green Master Mix (Roche) and an Eppendorf Realplex Mastercycler. The primers were designed using the program Primer 5.0 according to EmuJ_000439500 sequence data and included 5’-ATACAAAATGCGATGGTCATAGAGG-3’, 5’-AATGAGGTGGTAACACACACAGC-3’, 5’-GCAGTTTGTTGTCACTCGGGT-3’, and 5’-GGACACTTTTACCTTTCACCTC-3’.
Recombinant Em-CI Expression
The Em-CI predicted by bioinformatics analysis was expressed as a recombinant protein in Pichia pastoris. A codon-optimized synthetic gene encoding the ESP was obtained from Takara Biomedical Technology (Beijing, China) and subsequently cloned into pPICZαA. The EmuJ_000439500 gene was cloned into the EcoRI and XbaI sites of DH5a bacteria. The plasmids were extracted using Qiagen plasmid purification systems. The constructed plasmids were verified by PCR using the forward primer 5’-CCAGTCAACACTACAACAGAAGATG-3’ and the reverse primer 5’-CTTCTTGTACTTCCACTGGTTGC-3’.
The recombinant plasmid containing the EmuJ_000439500 sequence was then linearized with the SacI restriction enzyme and transfected into P. pastoris X-33 by electroporation using a Gene Pulser Xcell™ Electroporation System (Bio-Rad Laboratories, USA) at 1500 V for 10 ms (voltage, 1500 V; resistance, 200 Ω; capacitance, 25 μF). The transformants were initially selected on YPD (10 g/l yeast extract, 20 g/l peptone, 20 g/l glucose and 20 g/l agar) plates containing 150 μg/mL zeocin and subsequently reselected on YPD plates containing 500 μg/mL zeocin. Finally, the transformants that survived at the higher zeocin concentration were used for the production of Em-CI. The P. pastoris transformants were grown in a liquid medium containing 0.5% methanol for induction of Em-CI expression. Em-CI secreted into the medium was purified by cation-exchange (HiTrap Heparin HP) chromatography. An AKTA Prime fast protein liquid chromatography system (GE Healthcare) was used for the purification steps. After separating the proteins by SDS-PAGE, Western blot analysis was carried out with an anti-His antibody (BD Biosciences) for identification of the recombinant protein.
Recombinant Em-CI Haemolytic Assay
To determine whether Em-CI can inhibit activation of the complement CP, 1 × 10^7 ShEs (corresponding to 50 µL) were incubated in 50 μL of DGVB for 30 min in the presence of 50 μL of Em-CI or heat-inactivated Em-CI (HI-Em-CI) (96°C, 30 min) in different concentrations and 50 μL of NHS (1:50 diluted). Aliquots of 150 µL of 0.5% Triton X-100 and buffer were used as positive and negative controls, respectively. Thereafter, the erythrocytes were centrifuged (2000 × g, 10 min), and the released haemoglobin was measured at 541 nm using a microplate reader (Thermo Scientific). The haemolysis rate was determined by the average absorbance value. The absorbance value of each sample was calculated as follows: (sample value – negative control group value)/(positive control value – negative control group value).
Summary
This study explored the interaction between Echinococcus multilocularis (E.m) and the host complement system. Through bioinformatics methods, for the first time, we discovered an excretory protein of E.m playing as a complement classic pathway inhibitor, which was further confirmed by the haemolysis assay. This study provides research strategies and insights regarding the interactions between E.m and the innate immune system.
Statistical Analysis
The experimental results are reported as the mean ± SD. Two-way ANOVA analysis was used for multiple comparisons (GraphPad Prism version 7 for Windows, San Diego, CA, USA). The Bonferroni correction was applied for multiple comparison. Differences at the * p < 0.0332, ** p < 0.0021, *** p < 0.0002 and **** p < 0.0001 levels were considered statistically significant.
Results
Identification of the Effects of CM of E.m Protoscoleces on Complement Activity
We first investigated whether CM of E.m could inhibit complement activity in the AP under different RE concentrations. The serum concentration was initially set at 20%. Rabbit erythrocytes (REs) were incubated with E.m CM obtained on day 3. The haemolysis rate increased rapidly with decreasing concentrations of REs until the RE concentration reached 2 × 108/200µL (Figure 1A and B). After that, we tested the haemolysis rates when 2 × 108 REs were incubated with E.m CM obtained on day 3 with different concentrations of normal human serum (NHS) (Figure 1C and D). Finally, we tested the haemolysis rates of REs (2 × 108) incubated with NHS (1:5 dilution) and CM obtained on different days. There was little change in haemolysis rates with increasing culture time (Figure 1E and F). All three tests showed no significant differences in haemolysis rates between CM and CM or heat-inactivated CM (HI-CM) (p > 0.05), suggesting that CM of E.m might have no inhibitory effects on complement activation through the AP.
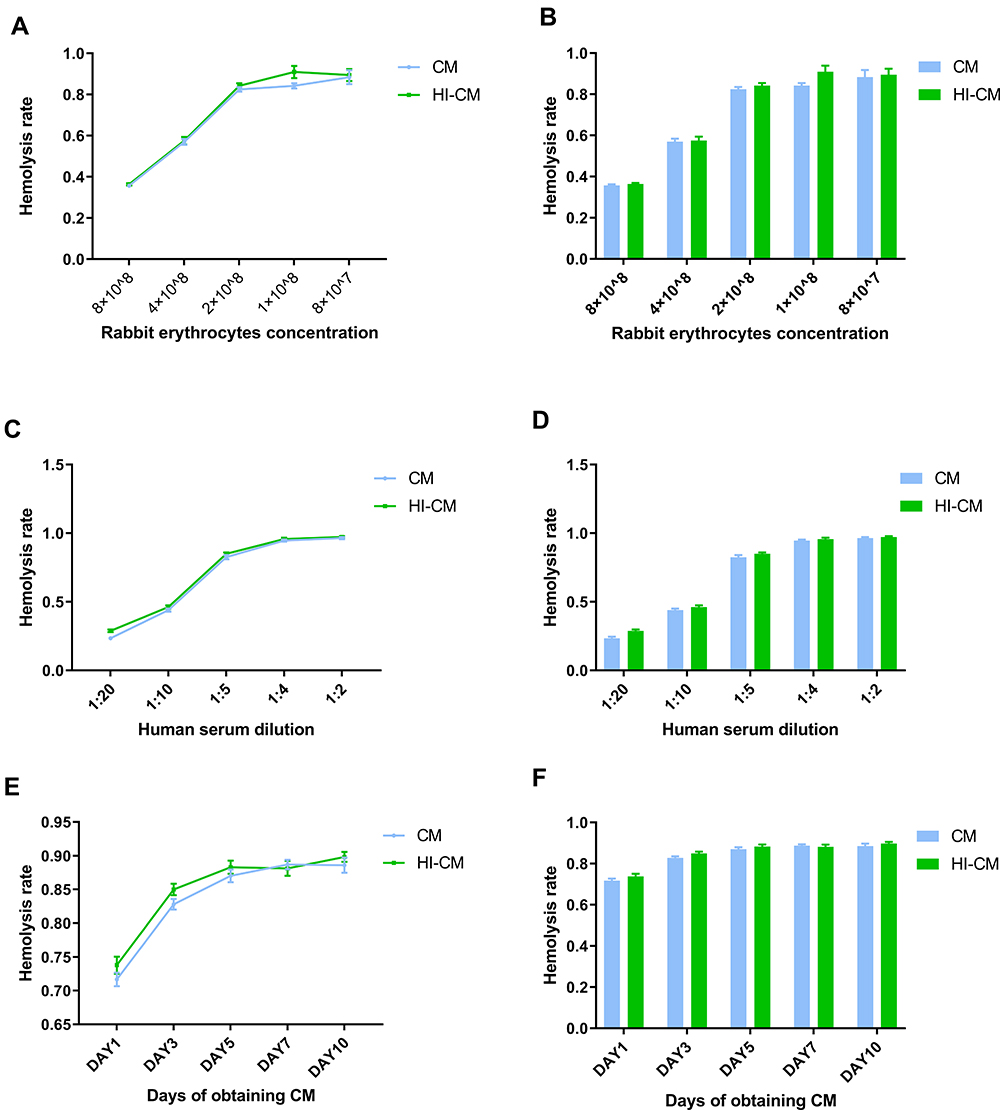 |
Figure 1 Determining complement alternative pathway inhibition ability of in vitro culture medium (CM) of Echinococcus multilocularis (E.m) protoscoleces through haemolytic assay. (A and B) Assay examining CM-mediated haemolysis in REs at different concentrations. NHS was diluted 1:5, and the CM was obtained on day 3. As the RE concentration decreased, the haemolysis rate increased rapidly until the RE concentration reached 2×108. (C and D) Assay examining CM-mediated haemolysis under different NHS concentrations. The RE concentration was 2 × 108, and the CM was obtained on day 3. The haemolysis rate increased rapidly as the NHS concentration increased until it reached 20%. (E and F) Assay examining CM-mediated haemolysis by CM obtained on different days. The RE concentration was 2 × 108, and the NHS was 1:5 diluted. There was little change in haemolysis rate with increasing culture time. After all, there was no significant differences in haemolysis rates between CM and HI-CM (p>0.05). |
We subsequently tested the complement inhibition caused by CM through the CP under different ShE concentrations. The serum concentration was initially set at 2%. Sheep erythrocytes (ShEs) were incubated with E.m CM obtained on day 3. The haemolysis rate increased rapidly with decreasing concentrations of REs until the ShE concentration reached 1×107/200µL (Figure 2A and B). The haemolysis rates when 1 × 107 ShEs were incubated with E.m CM obtained on day 3 and different concentrations of NHS were subsequently tested (Figure 2C and D). Finally, we tested the haemolysis rates of ShEs (1 × 107) incubated with NHS (1:20 dilution) and CM obtained on different days. There was little change in haemolysis rates with increasing culture time (Figure 2E and F). Intriguingly, the haemolysis rates of CM group are significantly lower than that of HI-CM group in most tests, suggesting that CM of E.m might have inhibitory effects on complement activation through the CP.
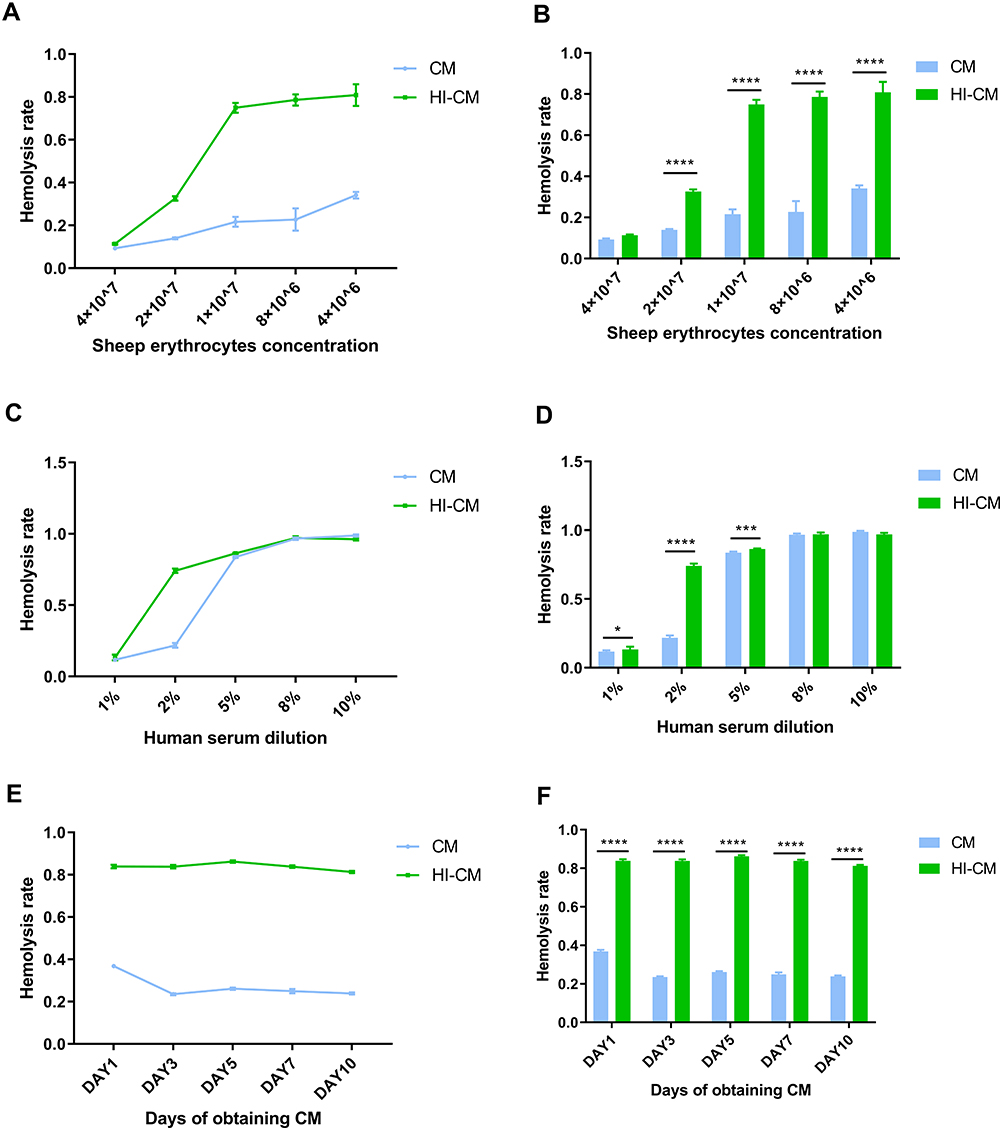 |
Figure 2 Determining complement classic pathway inhibition ability of in vitro culture medium (CM) of Echinococcus multilocularis (E.m) protoscoleces through haemolytic assay. (A and B) Assay examining CM-mediated haemolysis in ShEs at different concentrations. NHS was 1:50 diluted, and the CM was obtained on day 3. As the ShE concentration decreased, the haemolysis rate increased rapidly until the RE concentration reached 1×107. (C and D) Assay examining CM-mediated haemolysis under different NHS concentrations. The RE concentration was 1×107, and the CM was obtained on day 3. The haemolysis rate increased rapidly as the NHS concentration increased until it reached 5%. (E and F) Assay examining CM-mediated haemolysis by CM obtained on different days. The RE concentration was 1×107, and the NHS was 1:20 diluted. There was little change in haemolysis rate with increasing culture time. After all, the haemolysis rates of CM group are significantly lower than that of HI-CM group. *p < 0.0332, ***p < 0.0002 and ****p < 0.0001. |
Analysis and Prediction of the Function of the ESP-Encoding Gene EmuJ_000439500
Based on the above results, we preliminarily speculated that a specific component secreted by E.m in the CM might have suppressive effects on complement activation. ESPs, which are well-studied factors mediating evasion of the host immune system, have become popular research topics. Through protein functional annotation, pathway annotation, Clusters of Orthologous Groups (COGs) functional annotation and Gene Ontology (GO) annotation, we identified protein sequences for 653 ESPs, including 438 classical ESPs and 215 non-classical ESPs; these sequences accounted for 7.5% of the total proteome. Moreover, 360 ESPs were annotated and were enriched mainly for the integral component of membrane and extracellular region terms. These ESPs were associated with various molecular functions, such as protein binding, ATP binding, and serine-type endopeptidase inhibitor activity. In addition, some ESPs were found to be involved in several biological processes, including proteolysis, negative regulation of endopeptidase activity, and regulation of DNA-templated transcription. To better understand the potential functions of the ESPs, we conducted a protein domain enrichment analysis by searching the InterPro database. Of the 653 ESPs, 434 (66.5%) were successfully annotated in at least one database. A total of 490 ESPs were annotated in 300 InterPro protein families (Supplementary Table S1). The most enriched protein families include the pancreatic trypsin inhibitor Kunitz domain superfamily, the immunoglobulin-like fold superfamily, and the Taeniidae antigen family. A total of 332 ESPs were annotated with 206 domains (Supplementary Tables S2 and S3). The most enriched domains included the pancreatic trypsin inhibitor Kunitz domain, the peptidase C1A, papain C-terminal domain, and the papain-like cysteine endopeptidase domain. The ESP (UniProt: A0A068Y4F2)-encoding gene EmuJ_000439500 was annotated with the IPR000436 domain (Sushi domain/short consensus repeat/complement control protein), which suggests that A0A068Y4F2 is a potential complement inhibitor. See supplementary files for more information about the GO annotation, KEGG pathway analysis (Supplementary Figure S1), InterPro analysis, and PPI network analysis (Supplementary Figure S2).
EmuJ_000439500 encodes an ESP (UniProt: A0A068Y4F2, registration name: “C4b binding protein beta chain”) that is composed of 682 amino acids (Supplementary Table S4). The inferred molecular formula is C3265H5052N924O994S51, and the entire amino acid sequence contains 7 Sushi domains located at amino acids 111–168, 171–241, 242–313, 314–371, 372–434, 508–595 and 596–667. The amino acid sequence includes 68 negatively charged amino acid residues (Asp + Glu) and 72 positively charged amino acid residues (Arg + Lys). The relative molecular weight of E.m-derived complement inhibitor (Em-CI) was determined to be 74788.69, the theoretical isoelectric point was 7.85, and the instability coefficient was 39.55, suggesting that this ESP is a stable protein. The average hydrophilicity index, or grand average of hydropathicity (GRAVY), was −0.374, suggesting that Em-CI is hydrophilic. The estimated half-life of Em-CI was determined to be 30 hours in mammalian reticulocytes (in vitro), while it was predicted to be >20 hours in yeast (in vivo) and >10 hours in Escherichia coli (in vivo).
To further evaluate the hydrophilicity/hydrophobicity of Em-CI, we used ProtScale online software to analyse the protein. The approximate region of amino acids 5–20 was the most hydrophobic, while amino acid sites 100–115 and 445–455 exhibited strong hydrophobicity. In contrast, the most hydrophilic region encompassed amino acids 195–205, while amino acid sites 120–130 and 235–245 exhibited strong hydrophilicity (Figure 3A). According to the TMHMM 2.0 and SignalP 4.0 data, Em-CI was further confirmed to feature a signal peptide located at amino acids 1–18 (Figure 3B) and to lack a transmembrane structure (Figure 3C). Em-CI was predicted to have PPIs with 11 other ESPs (Figure 3D). The antigenic peptide prediction analysis (Supplementary Table S5) revealed that Em-CI contained 30 antigenic determinants with an average antigenic propensity of 1.0390. The region including amino acids 79–115 had the highest antigenic propensity, followed by the regions including amino acids 459–485 and 589–613 (Figure 3E). According to data from NetPhos, Em-CI contained 120 phosphorylation sites. Sites 46, 48, 658, 409, and 578 had somewhat higher phosphorylation potential than the other sites (Figure 3F). Self-Optimized Prediction Method with Alignment (SOPMA) analysis indicated that Em-CI contained α-helices, β-sheets, disordered coils, and β-turns with proportions of 4.11%, 22.43%, 68.04%, and 5.43%, respectively. The tertiary structure of Em-CI was visualized with Visual Molecular Dynamics (VMD) (Figure 4).
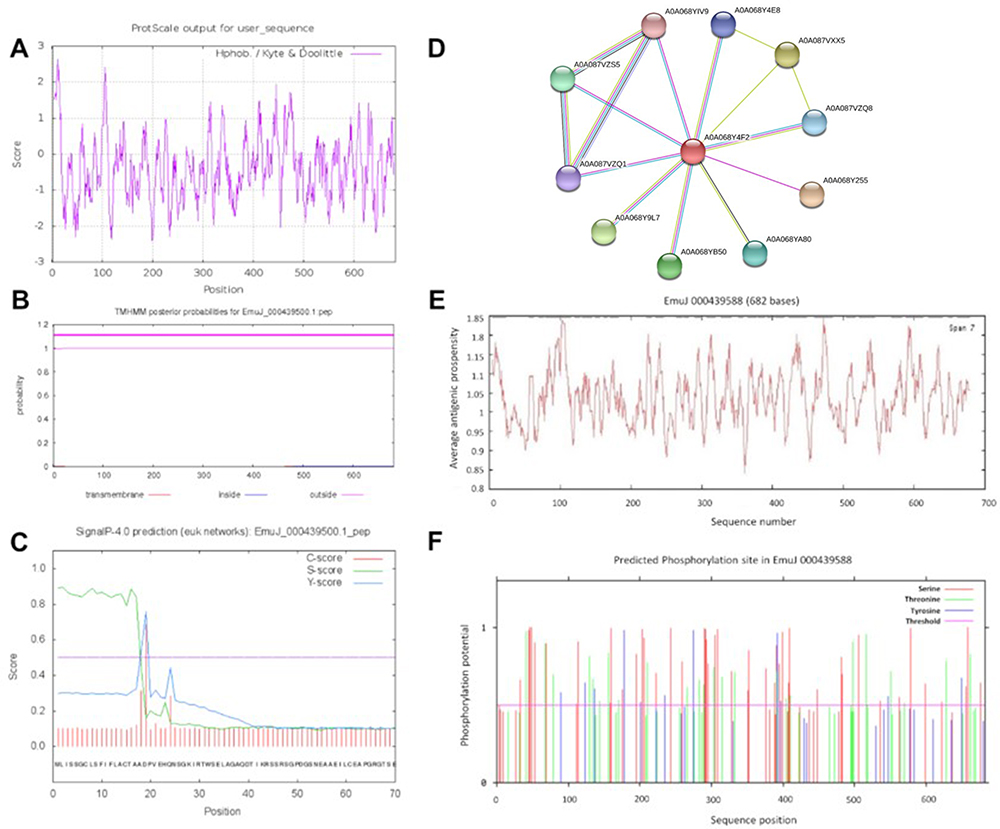 |
Figure 3 Prediction of the physical and chemical features of Echinococcus multilocularis derived complement inhibitor (Em-CI). (A) Hydrophobicity/hydrophilicity of Em-CI. Positive scores indicated hydrophobicity, and negative scores indicated hydrophilicity. A higher absolute value indicates a higher degree of hydrophobicity/hydrophilicity. (B) Em-CI transmembrane structure prediction. (C) Em-CI signal peptide prediction.fgh. (D) Protein-protein interaction network of Em-CI. (E) Antigenic epitope prediction of Em-CI. (F) Phosphorylation site predictions of Em-CI. |
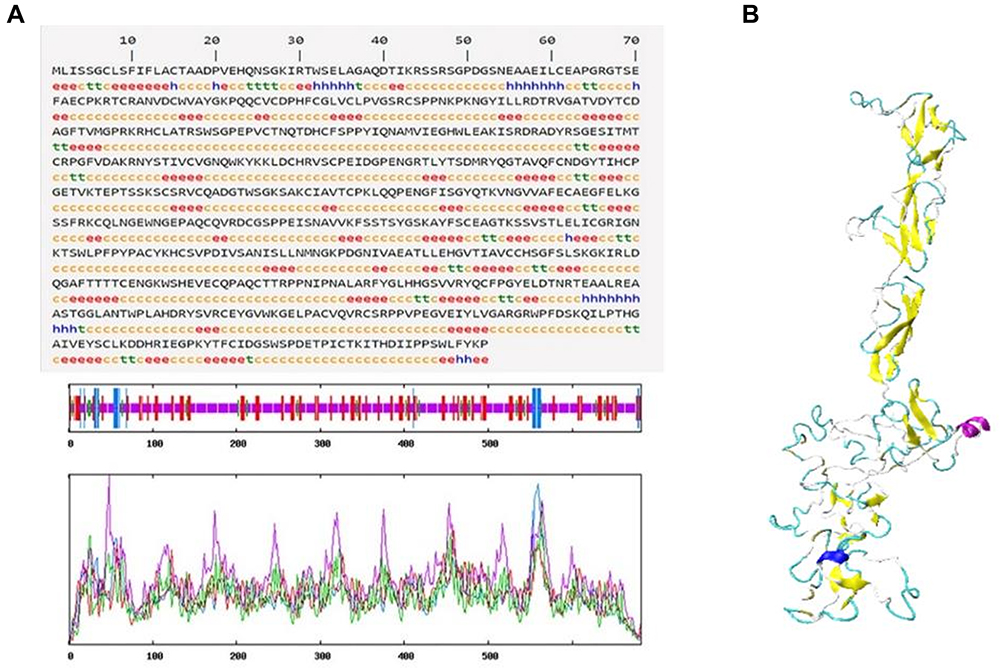 |
Figure 4 (A) Secondary structure of Echinococcus multilocularis derived complement inhibitor (Em-CI). hh denotes an α-helix, ee denotes an extended strand, tt denotes a β-turn, and cc denotes a random coil. (B) Tertiary structure of Em-CI. |
To verify whether EmuJ_000439500 mRNA was expressed in mouse lesions, liver samples were obtained from 6 infected Mongolian jirds and 3 noninfected jirds. As expected, the results of quantitative real-time PCR suggested that EmuJ_000439500 mRNA was expressed in the liver lesions of E.m-infected mice but was not expressed in normal liver specimens. Therefore, EmuJ_000439500, which encodes Em-CI, is markedly expressed in mouse livers during E.m infection (Supplementary Figure S3).
Recombinant EmuJ_000439500-Encoded Em-CI Displays Complement Inhibition Ability
To confirm that EmuJ_000439500 encodes an Em-CI, we constructed a recombinant plasmid containing the EmuJ_000439500 gene. After the pPICZαA vector was amplified with E. coli, the pPICZαA plasmid and the EmuJ_000439500 gene were digested with EcoRI and XbaI, ligated with ligase and transferred into E. coli for amplification (Figure 5A). The verified plasmid was then linearized and transferred into yeast (Supplementary Figures S4–S7). In the presence of 500 µg/mL bleomycin, a small amount of protein was expressed. Protein samples were collected after 24 h, 48 h and 72 h of bleomycin treatment and then verified by SDS-PAGE and Western blot analysis. Compared with empty vector transfection, recombinant vector transfection induced ESP production (Figure 5B and C).
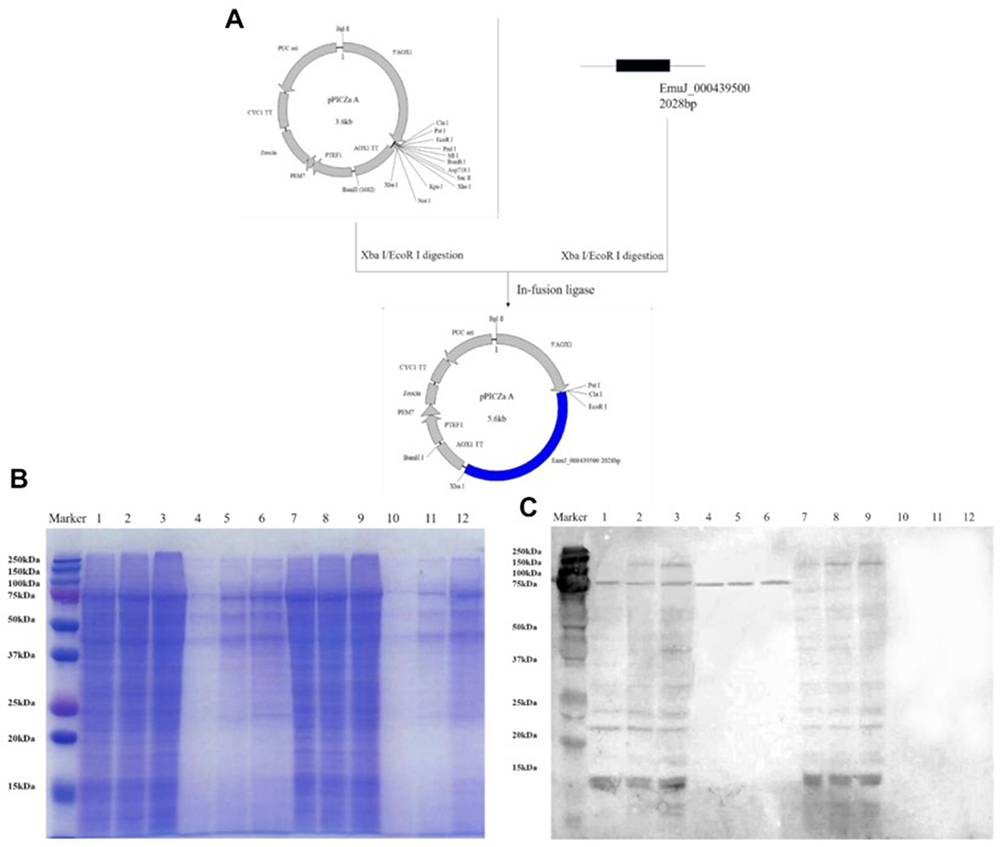 |
Figure 5 (A) Construction process of the pPICZαA recombinant vector. (B and C) Low levels of recombinant protein expression were validated by SDS-PAGE and Western blot analysis. Lanes 1–3: intracellular protein expression in yeast transfected with recombinant pPICZαA at 24 h, 48 h and 72 h. Lanes 4–6: extracellular protein expression by yeast transfected with recombinant pPICZαA at 24 h, 48 h and 72 h. Lanes 7–9: intracellular protein expression in yeast transfected with the empty vector at 24 h, 48 h and 72 h. Lanes 10–12: extracellular protein expression by yeast transfected with the empty vector at 24 h, 48 h and 72 h. |
To evaluate the complement-inhibiting ability of the recombinant Em-CI, we tested the haemolysis rates of 2 × 108 REs incubated with NHS at a 1:5 dilution and with the recombinant Em-CI at different concentrations (5 µg/mL, 10 µg/mL, 20 µg/mL, 50 µg/mL and 100 µg/mL). There was no difference in the haemolysis rate between the Em-CI group and the heat-inactivated Em-CI (HI-Em-CI), which suggested that Em-CI could not inhibit complement activation through the AP (Figure 6A and B). The effect of Em-CI on the CP was further tested in ShEs (concentration: 1 × 107) incubated with NHS at a 1:50 dilution and with recombinant Em-CI at the same concentrations listed above. The haemolysis rate of Em-CI group was significantly lower than that of the HI-Em-CI group, and the rate decreased with increasing concentrations of Em-CI (Figure 6C and D). Taken together, these results suggest that recombinant Em-CI inhibits complement activation through the CP in a dose-dependent manner.
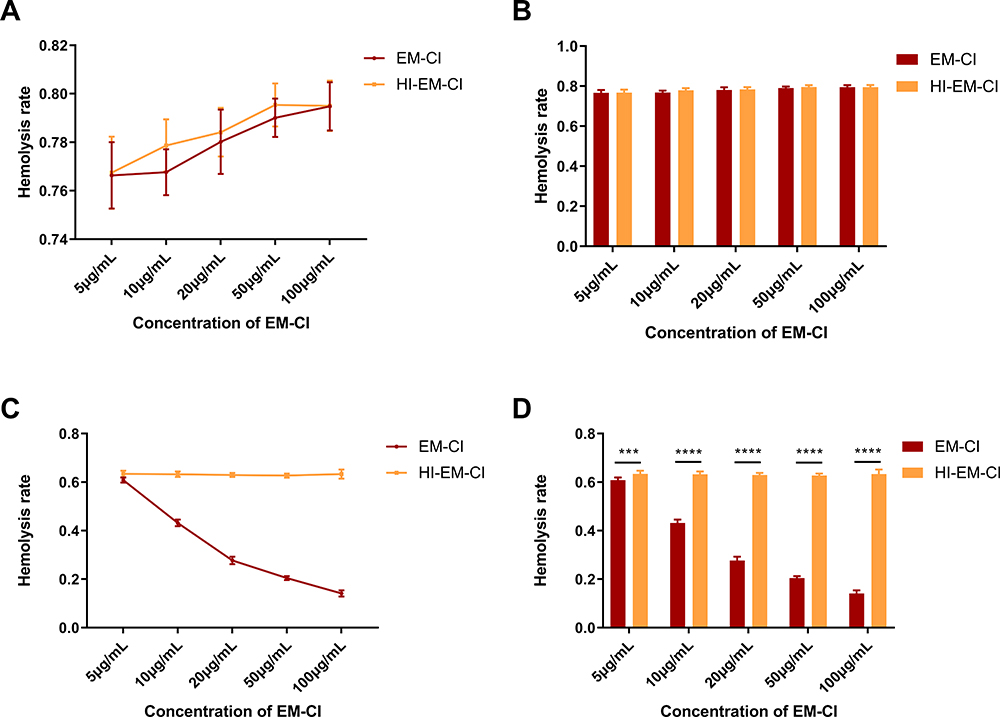 |
Figure 6 Determining complement inhibition ability of recombinant EmuJ_000439500-encoded Echinococcus multilocularis derived complement inhibitor (Em-CI) through haemolytic assay. (A and B) The AP-related haemolysis rates of the recombinant Em-CI and negative control groups were not significantly different when the RE concentration was 2×108, the NHS dilution was 1:5, and different concentrations of recombinant Em-CI were used. (C and D) When the ShE concentration was 1×107 and the NHS dilution was 1:50, the CP-related haemolysis rate of Em-CI group was significantly lower than that of heat-inactivated Em-CI (HI-Em-CI), and the haemolysis rate decreased with increasing concentrations of recombinant Em-CI. ***p < 0.0002 and ****p < 0.0001. |
Discussion
Oncospheres of E.m are able to avoid attack by the host complement system, while they penetrate the intestinal wall to enter the portal vein and eventually colonize the liver. However, the mechanism has remained unclear. Our findings reveal that E.m may selectively inhibit CP activation of the complement system by ESPs to evade complement killing. In addition, this study found that the medium of in vitro-cultured protoscoleces of E.m inhibits complement activation via the CP, suggesting that particular components secreted by protoscoleces into the CM can help inhibit complement activation. Bioinformatics methods were used to annotate and analyse the E.m proteome. The protein A0A068Y4F2 encoded by EmuJ_000439500 was ultimately selected from among 653 ESPs. A0A068Y4F2 was the only ESP containing 7 Sushi domains and was also the only potential inhibitor of complement activation. After predicting the basic physical and chemical properties of this protein, we used a yeast recombination system to produce recombinant Em-CI and found that it could inhibit activation of the complement system through the CP. These results were similar to the results obtained with E.m CM, suggesting that Em-CI may be a component of the CM that inhibits complement activation. The discovery of Em-CI sheds light on the mechanism by which E.m escapes killing by the complement system and provides potential targets for immunotherapy for parasitic diseases.
Following an improved method for in vitro culture of E.m protoscoleces reported by Wang et al,19 we obtained CM containing E.m protoscolex excretory proteins. The complement haemolysis assay verified that the CM inhibited complement CP activation. We thus hypothesized that ESPs contained in the medium inhibited complement activation. In previous studies, ESPs have been able to be discovered and verified only through conventional experimental methods, which are undoubtedly extremely costly in terms of time and financial and material resources. However, it is now possible to discover proteins and predict their functions through bioinformatics.20 The genome of E.m was published by Tsai et al in 2013.21 Based on the whole genome, this study utilized a variety of analysis tools to exclude proteins containing transmembrane regions and then distinguished between classical and non-classical pathway-secreted proteins according to whether the proteins contained N-terminal signal peptide sequences. Mitochondrial proteins, endoplasmic reticulum proteins, and glycosylphosphatidylinositol-anchored proteins were excluded from the list of possible secreted proteins. On this basis, we analysed the theoretical structures, functions, pathways, and interactions of the ESPs. Our findings not only lay a foundation for further discovery of potential complement inhibitors but also provide insights into the mechanisms of action of other E.m-derived molecules during the infection process.
Due to the presence of the typical complement regulatory protein domain, bioinformatics analysis revealed only one potential complement inhibitor among the E.m-derived ESPs. Thus far, only a few studies have used bioinformatics to predict E.m-derived ESPs and their functions. Multiple antigens have been confirmed to have good immunogenicity or immunoregulatory effects.22,23 However, few ESPs have been discovered so far, and there is very little information available on ESP structures and functions. The Em-CI found in this study was annotated as the only complement inhibitor because it contained 7 Sushi domains. The Sushi domain is a highly conserved functional domain. Many studies have demonstrated that multiple continuous Sushi domains are common features of complement inhibitors, such as H factor and CD46.12,24 In this study, Em-CI was annotated as the C4BP β chain in UniProt, suggesting that its function and mechanism may be similar to those of C4BP. However, in the human body, the C4BP α chain, rather than the β chain, binds to the C4-activated fragment C4b and thereby inhibits the formation of C3 convertase to inhibit complement activation. Moreover, C4BP does not even contain a β chain in mice and rabbits.11 Further analysis has shown that the β chain of C4BP in humans has a mass of 45 kDa and contains only two Sushi domains;25 thus, it is quite different from Em-CI. These results show that bioinformatics analyses can provide important clues for proteomics research but still cannot replace conventional proteomics research techniques. The structures and functions of proteins will eventually be able to be verified through experiments with recombinant proteins.
After detecting only one potential Em-CI through bioinformatics analysis, we produced recombinant Em-CI using a yeast recombinant expression system. Further experiments verified that Em-CI inhibited complement activation via the CP in a dose-dependent manner, confirming the prediction results of the bioinformatics analysis. Em-CI may inhibit the activation of CP through a pattern similar to that of C4BP, binding to and limiting the function of the activated complement fragment C4b. Em-CI may also act as a cofactor to the serine protease factor I in the proteolytic inactivation of both soluble and surface-bound C4b fragments, thus preventing the formation of the C4bC2a complex. Moreover, Em-CI may be capable of accelerating the natural decay of C3-convertase, which further inhibits the activation of CP. Unlike other pathogens, including gram-positive26 and negative bacteria27, E.m protoscolex directly excretes C4BP-like protein to inhibit activation of CP rather than recruit host-originated C4BP from serum. This novel mechanism may also enlighten a new avenue of study on pathogen-host complement interaction.
This study still had certain flaws. E.m is a parasite with a very complicated life history; it must develop from an egg into an oncosphere, colonize the liver, and further develop into a metacestode. However, E.m cannot fully mature into the adult stage in intermediate hosts, such as humans and mice and can only reproduce asexually via budding cysts in the liver of the intermediate host.28 E.m expresses different proteins at different stages of its life history.29 Some studies have investigated the use of specific proteins as markers for the identification of specific stages of the E.m life cycle.30–32 The medium collected in this study was used mainly for culturing protoscoleces. Whether the ESPs contained in CM are the same for E.m at other life stages requires further study. In addition, there is still a lack of Em-CI monoclonal antibodies; such antibodies would be useful tools for further research on the protective effect of Em-CI on E.m protoscoleces and even the immunotherapeutic efficacy of Em-CI targeting.
In this study, we discovered that an excretory protein of E.m can inhibit complement activation through the CP. Using bioinformatics analysis, we predicted the identity of a potential complement inhibitor among the excretory proteins of E.m. We confirmed that recombinant EmuJ_000439500, which encodes this excretory protein, is a dose-dependent CP complement inhibitor. This study provides research strategies and insights regarding the mechanisms of the interactions between E.m and the innate immune system. The complement system may be useful as a target for parasite immunotherapy in the future, and complement inhibition may be the final “secret” mechanism by which E.m escapes host immune system-mediated death.
Conclusion
In this research, we revealed that in vitro CM of E.m can inhibit activation of the CP of the complement system. Bioinformatics analysis identified an excretory protein encoded by EmuJ_000439500 that may be an effective component in the CM. Moreover, a recombinant form of this excretory protein expressed in a yeast system could also inhibit activation of the complement system CP. Therefore, we conclude that E.m may inhibit activation of the complement system via excretory proteins and thus evade complement killing.
Abbreviations
AE, Alveolar echinococcosis; E.m, Echinococcus multilocularis; CP, classical pathway; LP, the lectin pathway; AP, alternative pathway; C4BP, C4b-binding protein; E.g, Echinococcus granulosus; CM, in vitro culture medium; ESPs, Excretory/secretory proteins; Res, Rabbit erythrocytes; ShEs, Sheep erythrocytes; Em-CI, E.m-derived complement inhibitor.
Funding
This research was supported by the Science and Technology Program of the Sichuan Science and Technology Department (Nos. 2019YFS0029, 2019YFS0529), the National Natural Science Foundation of China (No. 81770566) and NHC Key Laboratory of Echinococcosis Prevention and Control (No. 2021WZK1004). The corresponding author, Wentao Wang, is the guarantor. The funding body financed the costs of the study and contributed to the design of the study, interpretation of the data, and revision of the manuscript.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Torgerson PR, Keller K, Magnotta M, et al. The global burden of alveolar echinococcosis. PLoS Negl Trop Dis. 2010;4(6):e722. doi:10.1371/journal.pntd.0000722
2. Vuitton DA. The ambiguous role of immunity in echinococcosis: protection of the host or of the parasite? Acta Trop. 2003;85(2):119–132. doi:10.1016/S0001-706X(02)00230-9
3. Kern P, Menezes da Silva A, Akhan O, et al. The echinococcoses: diagnosis, clinical management and burden of disease. Adv Parasitol. 2017;96:259–369. doi:10.1016/bs.apar.2016.09.006
4. Frank MM, Fries LF. The role of complement in inflammation and phagocytosis. Immunol Today. 1991;12(9):322–326. doi:10.1016/0167-5699(91)90009-I
5. Noris M, Remuzzi G. Overview of complement activation and regulation. Semin Nephrol. 2013;33(6):479–492. doi:10.1016/j.semnephrol.2013.08.001
6. Walport MJ, Mackay IR, Rosen FS. Complement. First of two parts. N Engl J Med. 2001;344(14):1058–1066. doi:10.1056/NEJM200104053441406
7. Ricklin D, Hajishengallis G, Yang K, et al. Complement: a key system for immune surveillance and homeostasis. Nat Immunol. 2010;11(9):785–797. doi:10.1038/ni.1923
8. Zipfel PF, Mihlan M, Skerka C. The alternative pathway of complement: a pattern recognition system. Adv Exp Med Biol. 2007;598:80–92.
9. Gottstein B, Hemphill A. Echinococcus multilocularis: the parasite-host interplay. Exp Parasitol. 2008;119(4):447–452. doi:10.1016/j.exppara.2008.03.002
10. Johnston MJ, MacDonald JA, McKay DM. Parasitic helminths: a pharmacopeia of anti-inflammatory molecules. Parasitology. 2009;136(2):125–147. doi:10.1017/S0031182008005210
11. Ermert D, Blom AM. C4b-binding protein: the good, the bad and the deadly. Novel functions of an old friend. Immunol Lett. 2016;169:82–92. doi:10.1016/j.imlet.2015.11.014
12. Gialeli C, Gungor B, Blom AM. Novel potential inhibitors of complement system and their roles in complement regulation and beyond. Mol Immunol. 2018;102:73–83. doi:10.1016/j.molimm.2018.05.023
13. Ma S, Zai J, Han Y, et al. Characterization of Schistosoma japonicum tetraspanning orphan receptor and its role in binding to complement C2 and immunoprotection against murine schistosomiasis. Parasit Vectors. 2017;10(1):288. doi:10.1186/s13071-017-2229-y
14. Reiss T, Rosa TF, Blaesius K, et al. Cutting edge: FHR-1 binding impairs factor H–mediated complement evasion by the malaria parasite Plasmodium falciparum. J Immunol. 2018;201(12):3497–3502. doi:10.4049/jimmunol.1800662
15. Ferreira AM, Würzner R, Hobart MJ, et al. Study of the in vitro activation of the complement alternative pathway by Echinococcus granulosus hydatid cyst fluid. Parasite Immunol. 1995;17(5):245–251. doi:10.1111/j.1365-3024.1995.tb01022.x
16. Díaz A, Ferreira AM, Nieto A. Echinococcus granulosus: interactions with host complement in secondary infection in mice. Exp Parasitol. 1995;80(3):473–482. doi:10.1006/expr.1995.1059
17. Ferreira AM, Breijo M, Sim RB, et al. Contribution of C5-mediated mechanisms to host defence against Echinococcus granulosus hydatid infection. Parasite Immunol. 2000;22(9):445–453. doi:10.1046/j.1365-3024.2000.00323.x
18. Hewitson JP, Grainger JR, Maizels RM. Helminth immunoregulation: the role of parasite secreted proteins in modulating host immunity. Mol Biochem Parasitol. 2009;167(1):1–11. doi:10.1016/j.molbiopara.2009.04.008
19. Wang H, Li J, Guo B, et al. In vitro culture of Echinococcus multilocularis producing protoscoleces and mouse infection with the cultured vesicles. Parasit Vectors. 2016;9(1):411. doi:10.1186/s13071-016-1687-y
20. Cortázar AR, Aransay AM, Alfaro M, et al. SECRETOOL: integrated secretome analysis tool for fungi. Amino Acids. 2014;46(2):471–473. doi:10.1007/s00726-013-1649-z
21. Tsai IJ, Zarowiecki M, Holroyd N, et al. The genomes of four tapeworm species reveal adaptations to parasitism. Nature. 2013;496(7443):57–63. doi:10.1038/nature12031
22. Dang Z, Watanabe J, Kajino K, et al. Molecular cloning and characterization of a T24-like protein in Echinococcus multilocularis. Mol Biochem Parasitol. 2009;168(1):117–119. doi:10.1016/j.molbiopara.2009.06.009
23. Deininger S, Wellinghausen N. Evaluation of a new combined Western and line blot assay (EUROLINE-WB) for diagnosis and species identification of Echinococcus infection in humans. GMS Infect Dis. 2019;7:Doc01. doi:10.3205/id000041
24. Holmquist E, Okroj M, Nodin B, et al. Sushi domain-containing protein 4 (SUSD4) inhibits complement by disrupting the formation of the classical C3 convertase. FASEB J. 2013;27(6):2355–2366. doi:10.1096/fj.12-222042
25. Hillarp A, Hessing M, Dahlbäck B. Protein S binding in relation to the subunit composition of human C4b-binding protein. FEBS Lett. 1989;259(1):53–56. doi:10.1016/0014-5793(89)81492-9
26. Jenkins HT, Mark L, Ball G, et al. Human C4b-binding protein, structural basis for interaction with streptococcal M protein, a major bacterial virulence factor. J Biol Chem. 2006;281(6):3690–3697. doi:10.1074/jbc.M511563200
27. Ngampasutadol J, Ram S, Blom AM, et al. Human C4b-binding protein selectively interacts with Neisseria gonorrhoeae and results in species-specific infection. Proc Natl Acad Sci U S A. 2005;102(47):17142–17147. doi:10.1073/pnas.0506471102
28. Brunetti E, Kern P, Vuitton DA. Expert consensus for the diagnosis and treatment of cystic and alveolar echinococcosis in humans. Acta Trop. 2010;114(1):1–16. doi:10.1016/j.actatropica.2009.11.001
29. Wang J, Gottstein B. Immunoregulation in larval Echinococcus multilocularis infection. Parasite Immunol. 2016;38(3):182–192. doi:10.1111/pim.12292
30. Gottstein B, Eckert J, Woodtli W. Determination of parasite-specific immunoglobulins using the ELISA in patients with echinococcosis treated with mebendazole[J]. Zeitschrift für Parasitenkunde. 1984;70(3): 385-389.
31. Ito A, Nakao M, Kutsumi H, et al. Serodiagnosis of alveolar hydatid disease by Western blotting. Trans R Soc Trop Med Hyg. 1993;87(2):170–172. doi:10.1016/0035-9203(93)90476-7
32. Gauci C, Merli M, Muller V, et al. Molecular cloning of a vaccine antigen against infection with the larval stage of Echinococcus multilocularis. Infect Immun. 2002;70(7):3969–3972. doi:10.1128/IAI.70.7.3969-3972.2002
33. Spiliotis M, Brehm K. Axenic in vitro cultivation of Echinococcus multilocularis metacestode vesicles and the generation of primary cell cultures. Methods Mol Biol. 2009;470:245–262.
34. Brehm K, Wolf M, Beland H, et al. Analysis of differential gene expression in Echinococcus multilocularis larval stages by means of spliced leader differential display. Int J Parasitol. 2003;33(11):1145–1159. doi:10.1016/S0020-7519(03)00169-3
35. Hemmings L, McManus DP. The diagnostic value and molecular characterisation of an Echinococcus multilocularis antigen gene clone. Mol Biochem Parasitol. 1991;44(1):53–61. doi:10.1016/0166-6851(91)90220-Z
36. Lightowlers MW, Lawrence SB, Gauci CG, et al. Vaccination against hydatidosis using a defined recombinant antigen. Parasite Immunol. 1996;18(9):457–462. doi:10.1111/j.1365-3024.1996.tb01029.x
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The full terms of this license are available at https://www.dovepress.com/terms.php and incorporate the Creative Commons Attribution - Non Commercial (unported, v3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted without any further permission from Dove Medical Press Limited, provided the work is properly attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.