Back to Journals » Journal of Inflammation Research » Volume 14
Altered T-Cell Subsets are Associated with Dysregulated Cytokine Secretion of CD4+ T Cells During HIV Infection
Authors Wang D, Jiang Y, Song Y, Zeng Y, Li C, Wang X, Liu Y, Xiao J, Kong Y, Zhao H ![]()
Received 17 August 2021
Accepted for publication 27 September 2021
Published 7 October 2021 Volume 2021:14 Pages 5149—5163
DOI https://doi.org/10.2147/JIR.S333902
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 4
Editor who approved publication: Professor Ning Quan
Di Wang,1,2 Yu Jiang,2 Yangzi Song,1,2 Yongqin Zeng,1 Cuilin Li,1 Xinyue Wang,2 Ying Liu,1,2 Jiang Xiao,1 Yaxian Kong,2 Hongxin Zhao1
1Clinical and Research Center of Infectious Diseases, Beijing Ditan Hospital, Capital Medical University, Beijing, People’s Republic of China; 2Beijing Key Laboratory of Emerging Infectious Diseases, Institute of Infectious Diseases, Beijing Ditan Hospital, Capital Medical University, Beijing, People’s Republic of China
Correspondence: Hongxin Zhao
Clinical and Research Center of Infectious Diseases, Beijing Ditan Hospital, Capital Medical University, Beijing, 100015, People’s Republic of China
Tel +86 10-84322583
Email [email protected]
Yaxian Kong
Beijing Key Laboratory of Emerging Infectious Diseases, Institute of Infectious Diseases, Beijing Ditan Hospital, Capital Medical University, Beijing, 100015, People’s Republic of China
Tel +86 10-84322620
Email [email protected]
Background: CD4+ T cells play a critical role in the regulation of immunopathogenesis in HIV infection. Previous studies have shown contradictory results of the CD4+ T-cell responses in people living with HIV (PLHIV).
Methods: A cross-sectional study was performed on 40 healthy controls, 134 ART-naïve PLHIV, and 34 individuals who experienced 3-year ART with low baseline CD4 count from 4 August 2016 to 23 January 2019. We determined the frequencies of CD4+ T-cell subsets and described the cytokine secretion pattern of total and subsets of CD4+ T cells in these individuals.
Results: We found that CD4+ T cells in PLHIV displayed enhanced secretion of pro-inflammation cytokines and polyfunctionality due to HIV disease progression (r = − 0.282, P = 0.0035 for IFN-γ; r = − 0.412, P = 0.0002 for TNF-α; r = − 0.243, P < 0.0001 for GM-CSF; r = − 0.252, P = 0.0093 for IFN-γ+ TNF-α+ cells). However, the altered T-cell subsets, as presented by the loss of naïve cells and expansion of memory/effector population in PLHIV, were associated with discordant results in total and subsets of CD4+ T cells. As major cytokine-producing T subsets, effector/memory CD4 subsets showed impaired cytokine production (P < 0.05). We further demonstrated that 3-year ART treatment could improve CD4 counts by increasing the pool of naïve T cells but could not restore cytokine secretion in CD4+ T-cell subsets (P < 0.05).
Conclusion: These data identified the impaired capacity of cytokine secretion in CD4+ T-cell subsets due to HIV disease progression, and the altered T-cell subsets were associated with pseudo-elevation of cytokine production in total CD4+ T cells. This study collectively suggested the importance of therapies that can preserve and/or enhance the function of CD4+ T cells in strategies of HIV remission.
Keywords: HIV infection, cytokines, T-cell dysfunction, T-cell subsets, inflammatory responses
Introduction
Chronic human immunodeficiency virus (HIV) infection causes progressive destruction of CD4+ T cells and profound immune suppression, increasing the risk for opportunistic infections.1 As a target of HIV infection, CD4+ T cells play a critical role in the modulation of innate immunity and initiation of adaptive immunity by their production of cytokines and chemokines to orchestrate the full panoply of immune responses.2–4 Recently, considerable emphasis has been placed on the cytokine immunity of CD4+ T cells regulating immunopathogenesis during HIV infection.5
CD4+ T cells, commonly referred to as T-helper (Th) cells, differentiate into Th1 and Th2 populations characterized exclusively by cytokine secretion patterns and distinct biological functions. Th1 cells synthesize IFN-γ, TNF-α, IL-2, and are associated with protective responses, whereas Th2 cells secret anti-inflammatory cytokines such as IL-10.6,7 Numerous studies showed defects in Th1 immune response with reduced expression of IL-2 and IFN-γ during HIV infection.8,9 On the contrary, some studies revealed that HIV could activate the Th1 immune response, induce inflammatory processes, and indirectly accelerate disease progression.10,11 Diverse methodological approaches could explain the contradictory results for the analysis of cytokines or Th subsets. A range of novel subsets of T helper cells, which could produce IL-17, IL-22, IL-9, and IL-27, have been characterized as Th17, Th22, Th9, and Th27 cells.12–15 Depletion of Th17 and Th22 cells were observed in PLHIV and was linked to the loss of mucosal immunity.14,16,17 Gorenec et al found a decrease of Th9 cells from acute to chronic stage of HIV infection.18 Additionally, IL-27 was demonstrated to exhibit potent anti-HIV properties in all major cell types targeted by HIV.15,19,20 More importantly, accumulating evidence suggested that polyfunctional T cells (PFC) are associated with more effective control of chronic viral infections, including HIV.21–24 However, our knowledge of the complex cytokine profile of Th cells during HIV infection was limited.
Most previous studies of cytokine profiles in PLHIV were based on serological data. However, it is impossible to determine the exact source of elevated cytokines in plasma. A plethora of cytokines is not only released by CD4+ T helper cells but also by CD8+ T cells or innate immune cells.25 CD4+ T cells are also heterogeneous and display differential proliferative capacities and effector functions by producing characteristic cytokines.26 It is necessary to consider the imbalance of distinct CD4+ T lymphocyte subsets during HIV infection. In the present study, we described the profile of multiple Th cells subsets in PLHIV. Overall, we found contradictory CD4+ T-cell responses in total and subsets of CD4+ T cells, which could be explained by the altered distribution of T-cell subsets during HIV infection.
Materials and Methods
Study Participants
We conducted a cross-sectional study, which was approved by the Committee of Ethics at Beijing Ditan Hospital, Capital Medical University in Beijing, and the study conformed to the tenets of the Declaration of Helsinki. All the human blood samples were collected with informed consent. From 4 August 2016 to 23 January 2019, we examined the cytokine secretion pattern of CD4+ T cells in 40 healthy controls, 134 ART-naïve, and 34 ART-experienced individuals with HIV infection. The precise infection time in these PLHIV is unknown. At the time of specimen collection, ART-experienced PLHIV were treated for a median of 3 years (IQR 2.0–3.4) with low baseline CD4 count (baseline CD4 <200 cells/μL) and had plasma HIV RNA levels that were below detection according to routine clinical assays. Participants co-infected with hepatitis B virus (HBV) infection, hepatitis C virus (HCV) infection, syphilis, or other opportunistic infections/cancers were excluded. Among ART-experienced PLHIV, 12 immunological responders (CD4 cell count <200 cells/μL before ART and CD4 cell count ≥350 cells/μL at 3 years following ART initiation) were included. The clinical details of all the PLHIV are shown in Table 1 and Supplementary Table 1.
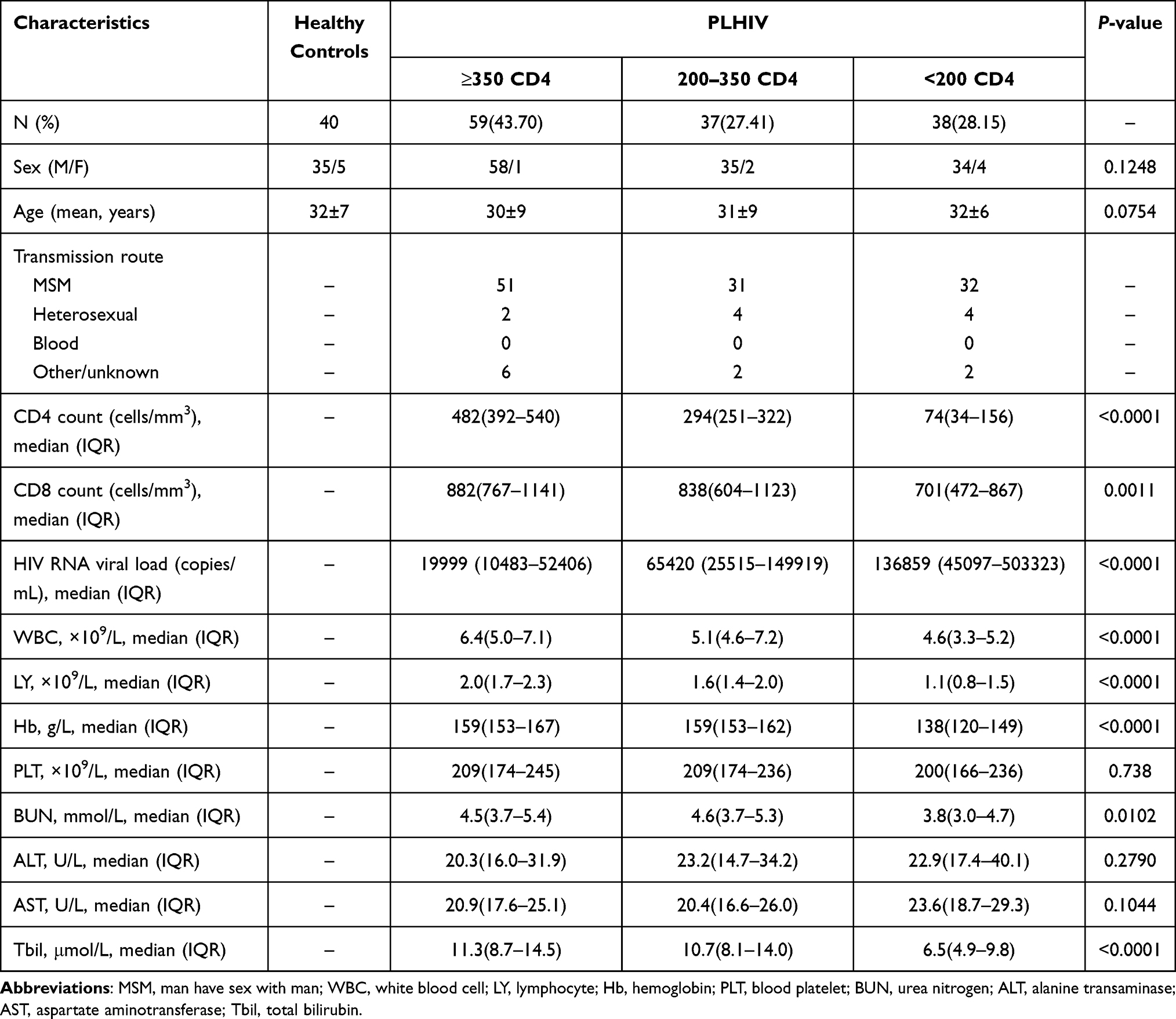 |
Table 1 Demographic and Clinical Characteristics in ART-Naïve PLHIV |
Sample Collection and Processing
Peripheral blood mononuclear cells were prepared from venous blood by density gradient centrifugation. All samples were processed and analyzed within 24 hours of collection.
Plasma HIV-1 Viral Load and CD4+ T-Cell Count
The plasma HIV-1-RNA levels were measured using a Standard Amplicor HIV Monitor assay, version 1.5 (Roche Diagnostics, Indianapolis, IN, USA), with a lower detection limit of 40 copies/mL. The CD4+ T-cell count was determined with a standard flow cytometry technique with a TruCOUNT tube in routinely equipped laboratories (BD Biosciences, San Jose, CA, USA). Both measurements were determined twice per year in a single laboratory using standard methodologies that are included in the National Quality Assurance Programs.
Flow Cytometry Cell Phenotype Analysis
PBMCs were incubated with directly conjugated antibodies for 30 min at 4°C. The cells were washed before flow cytometry analysis. Antibodies used included anti-human CD3-BV786 (clone SK7), CD8-BV510 (clone SK1), CD45RA-BV711 (clone HI100, BD Biosciences, San Diego, CA, USA), CD4-APC-Fire750 (clone SK3), CCR7-PE-CY7 (clone G043H7, BioLegend, San Diego, CA, USA), and the corresponding isotype controls including BV786 Mouse IgG1 (clone X40), BV510 Mouse IgG1 (clone X40), BV711 Mouse IgG2b (clone 27–35, BD Biosciences), APC-Fire750 Mouse IgG1 (clone MOPC-21), PE-CY7 Mouse IgG2a (clone MOPC-173, BioLegend). Flow cytometric acquisition was performed on at least 100,000 T lymphocytes on BD LSRFortessa driven by the FACS Diva software. Analysis of the acquired data was performed using FlowJo software (TreeStar, Ashland, OR, USA).
In vitro Stimulation and Intracellular Staining
Levels of Th cells were determined as the percentage of CD4+ T cells that produce these cytokines following in vitro stimulation with PMA & Ionomycin. PBMCs, isolated as described above, were resuspended to 1×106 cells/mL in RPMI 1640 medium plus 10% FBS (R10). Cells were then incubated for 5h at 37°C in a medium with 1 × Cell Stimulation Cocktail (containing 81 nM PMA and 1.34 nM ionomycin plus protein transport inhibitors, Ebioscience, San Diego, CA, USA). Following incubation, the cells were washed and surface-stained with CD3-BV786, CD4-APC-Fire750, CD8-BV510 for 30 minutes in the dark at room temperature, followed by fixation and permeabilization. After permeabilization, cells were stained with IFN-γ-AF700 (clone 4S.B3), IL-22-FITC (clone 22URTI), IL-9-PE (clone MH9D1), IL-27-APC (clone ebic6), IL-10-PE-CY7 (clone JES3-9D7, Ebioscience), TNF-α-BV711 (clone MAb11), IL-17-BV421 (clone BL168), IL-2-BV650 (clone MQ1-17H12, BioLegend), or GM-CSF-PE-CF594 (clone BVD2-21C11, BD Biosciences) antibodies for 30 minutes in the dark at room temperature. Following staining, cells were washed and acquired on an LSRFortessa.
Statistical Analysis
The data are expressed as the mean [standard deviation (SD)], median [interquartile range (IQR)], and percentage [frequency]. The normality of each variable was evaluated using the Kolmogorov–Smirnov test. In cases of two normally distributed data, the comparison of variables was performed using unpaired or paired data were specified and two-tailed Student’s t-tests for unpaired and paired data, respectively. A one-way ANOVA test followed by Tukey’s multiple comparisons test was performed to compare two more independent samples. When the data were not normally distributed, the comparison of variables was performed with a Mann–Whitney U-test or a Wilcoxon matched-pairs signed-rank test for unpaired and paired data. A Kruskal–Wallis test followed by Dunn’s multiple comparisons test was applied for comparing two more independent samples. Comparisons of participant characteristics were analyzed using Fisher’s exact test (categorical variables) or the Kruskal–Wallis test (continuous variables). Pearson’s or Spearman correlation coefficients were used to evaluating correlations for normally or non-normally distributed data, respectively. All analyses were performed using GraphPad7 (GraphPad Software, La Jolla, CA, USA) or SPSS (IBM Corporation, New York, NY, USA). P<0.05 was considered significant.
Results
Intracellular Cytokine Profile of CD4+ T Cells of PLHIV in Response to PMA Activation
We recruited 134 ART-naïve PLHIV and 40 gender- and age-matched healthy controls. According to their baseline CD4 cell count, 134 PLHIV were subdivided into three subgroups at three CD4 cell count thresholds (<200 cells/µL, 200–350 cells/µL, and ≥350 cells/µL). The clinical characteristics and laboratory parameters are presented in Table 1.
To determine the cytokine secretion pattern of CD4+ T cells after activation, we in vitro stimulated PBMCs with PMA/Ionomycin. We then analyzed a series of representative T helper cell-related cytokines (IFN-γ, TNF-α, GM-CSF, IL-17, IL-22, IL-9, IL-27, IL-10, and IL-2) by using multicolor intracellular cytokine staining. Representative flow cytometry gating strategy for cytokine-producing CD4+ T cells is illustrated in Figure S1. As shown in the heatmap of Figure 1A, CD4+ T cells in PLHIV exhibited aberrant profiles of these cytokines compared with those in healthy controls.
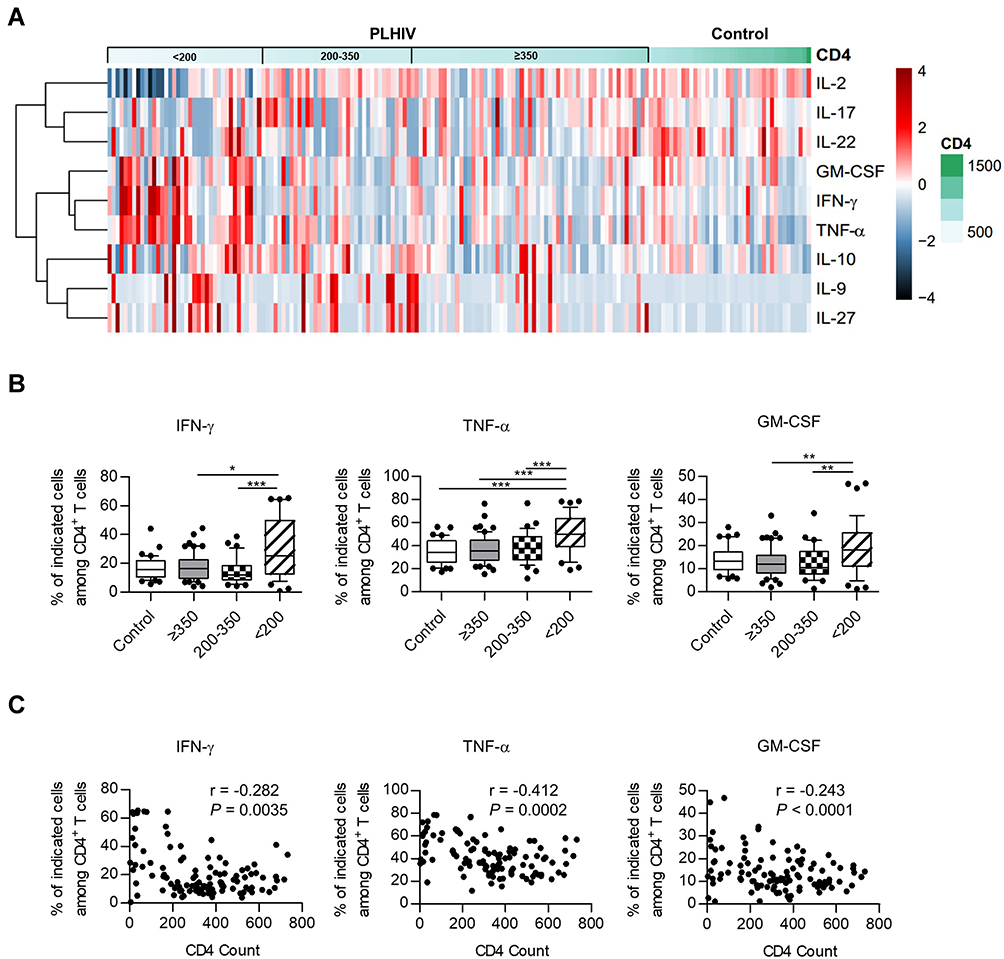 |
Figure 1 Heatmap and cluster analysis of cytokine profiles and correlation with HIV disease progression. (A) Heatmap and cluster analysis of cytokines for healthy donors and PLHIV. The percentage of cytokine-producing cells was expressed as color scales (black represents low levels; red, high levels). Dendrograms on the left side reflected proximities of cytokines based on intracellular cytokine staining. (B) The expression of IFN-γ, TNF-α, and GM-CSF in healthy controls and different PLHIV groups (CD4 count≥350 cells/µL, 200–350 cells/µL, <200 cells/µL). P values were obtained by the Kruskal–Wallis test followed by Dunn’s multiple comparisons test (IFN-γ, GM-CSF) or one-way ANOVA test followed by Tukey’s multiple comparisons test (TNF-α). (C) Correlation analysis of the percentages of cytokine-producing CD4+ T cells and CD4 count. Pearson’s (TNF-α) or spearman’s (IFN-γ, GM-CSF) test was used to test for correlations. *P < 0.05, **P < 0.01, ***P < 0.001. Abbreviations: GM-CSF, granulocyte-macrophage colony-stimulating factor; IFN, interferon; IL, interleukin; TNF, tumor necrosis factor. |
Compared with healthy donors, the expression of IFN-γ, TNF-α, and GM-CSF on CD4+ T cells was found to be significantly increased in the PLHIV with low CD4 counts (<200 cells/µL) (Figure 1B). There were also higher percentages of IL-9+ and IL-27+ cells in PLHIV than in healthy controls. In addition, we noted a decreased frequency of IL-22 on CD4+ T cells from every PLHIV group compared with those in healthy donors, suggesting IL-22 production was severely affected since a relatively early stage of HIV infection. Of interest, a decrease in IL-2 percentage was observed in CD4+ T cells from PLHIV with CD4 count <200 cells/µL. The frequencies of IL-10- and IL-17-expressing CD4+ T cells did not show a significant difference between PLHIV and healthy donors (Figures 1A and S2A).
Correlation analysis revealed that TNF-α+ cells frequencies among CD4+ T cells were negatively correlated with CD4 count (r = −0.412, P = 0.0002; Figure 1C), whereas the frequencies of IL-2+ cells exhibited a positive correlation (r = 0.437, P = 0.0033; Figure S2B). Although not statistically significant, there was a trend that suggested that frequencies of intracellular IFN-γ and GM-CSF increased with HIV disease progression (r = −0.282, P = 0.0035; r = −0.243, P = 0.0001; Figure 1C). Similar results were obtained in IL-9 and IL-27 expression, and no significant correlation was observed between the expression of CD4 count and IL-17, IL-22, or IL-10 (Figure S2B). Thus, our data revealed a tendency toward secretion of pro-inflammation cytokines in the PLHIV with low CD4 count, consistent with previous reports.27,28
Assessment of Cytokines Producing CD4+ T-Cell Polyfunctionality in PLHIV
We then assessed the ability of blood CD4+ T cells to co-produce multiple pro-inflammation cytokines in PLHIV and healthy donors, including IFN-γ, TNF-α, and GM-CSF. Representative staining for these cytokines is shown in Figure 2A. Analysis of CD4+ T-cell cytokine profiles (ie, all combinations of IFN-γ, TNF-α, and GM-CSF) revealed frequent coproduction of multiple cytokines.
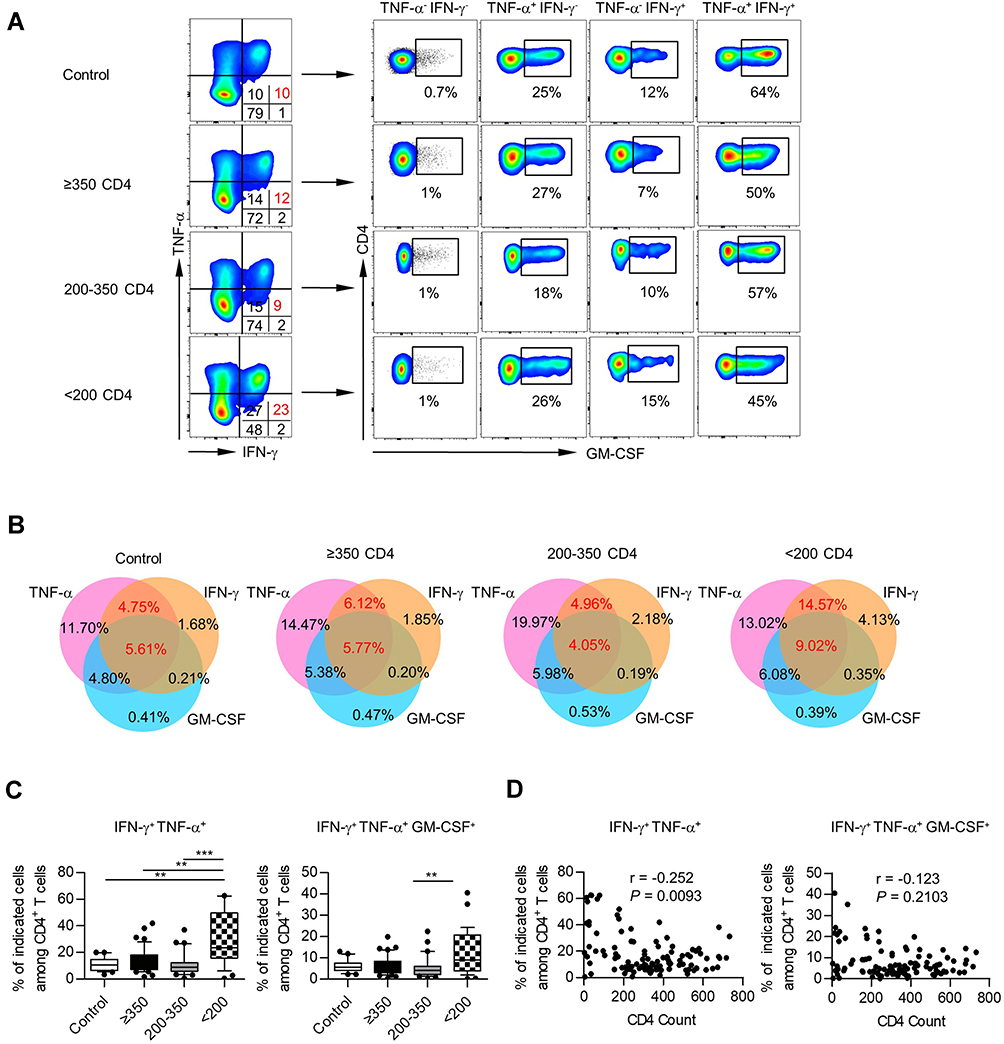 |
Figure 2 Polyfunctionality of cytokine-producing CD4+ T cells in PBMCs. (A) Representative staining for IFN-γ, TNF-α, and GM-CSF within CD4+ T cells from healthy controls and different PLHIV groups. (B) Venn diagram of the differentially double or triple-positive CD4+ T cells in healthy controls and different PLHIV groups (CD4 count≥350 cells/µL, 200–350 cells/µL, <200 cells/µL). (C) Box plots of the frequencies of IFN-γ+ TNF-α+, IFN-γ+ TNF-α+ GM-CSF+ cells on CD4+ T cells from healthy controls and different PLHIV groups (CD4 count≥350 cells/µL, 200–350 cells/µL, <200 cells/µL). P values were obtained by Kruskal–Wallis test followed by Dunn’s multiple comparisons test. (D) Correlation analysis of CD4 count and percentages of IFN-γ+ TNF-α+, IFN-γ+ TNF-α+ GM-CSF+ cells among CD4+ T cells. Spearman’s non-parametric test was used to test for correlations. **P < 0.01, ***P < 0.001. |
To better characterize these CD4+ T-cell subsets, we used a 3-set Venn diagram displaying the overlap among IFN-γ-, TNF-α- and GM-CSF-producing CD4+ T cells (Figure 2B). Specifically, all these CD4+ T-cell subsets showed remarkably larger fractions of cells producing double or triple-positive cytokines both in PLHIV and healthy controls. We also observed an increased proportion of IFN-γ+ TNF-α+ in CD4+ T cells in the PLHIV with CD4 count <200 cells/µL (Figure 2C), and correlation analysis confirmed that the percentage of IFN-γ+ TNF-α+ cells was elevated accompanying by a decrease in CD4 count (r = −0.252, P = 0.0093; Figure 2D). Of note, we found that the majority of IFN-γ+ TNF-α+ CD4+ T cells were also GM-CSF+ (Figure 2A). Therefore, the PLHIV with CD4 count <200 cells/µL also displayed an increased fraction of IFN-γ+ TNF-α+ GM-CSF+ in CD4+ T cells (Figure 2C). However, we could not find a correlation between the triple-functional CD4+ T cells and CD4 numbers (Figure 2D). A similar polyfunctionality was also found in IL-9+ and IL-27+ CD4+ T cells (Figure S3A and B).
Since the critical role of immune activation and exhaustion during HIV infection was previously stated, we further analyzed the expression of HLA-DR, CD38, and PD-1. As shown in Table 2, HLA-DR and PD-1 expression in CD4+ T cells were strongly correlated with single or multiple expression of IFN-γ, TNF-α, and GM-CSF, implying the involvement of immune activation and exhaustion in pro-inflammatory responses during HIV infection.
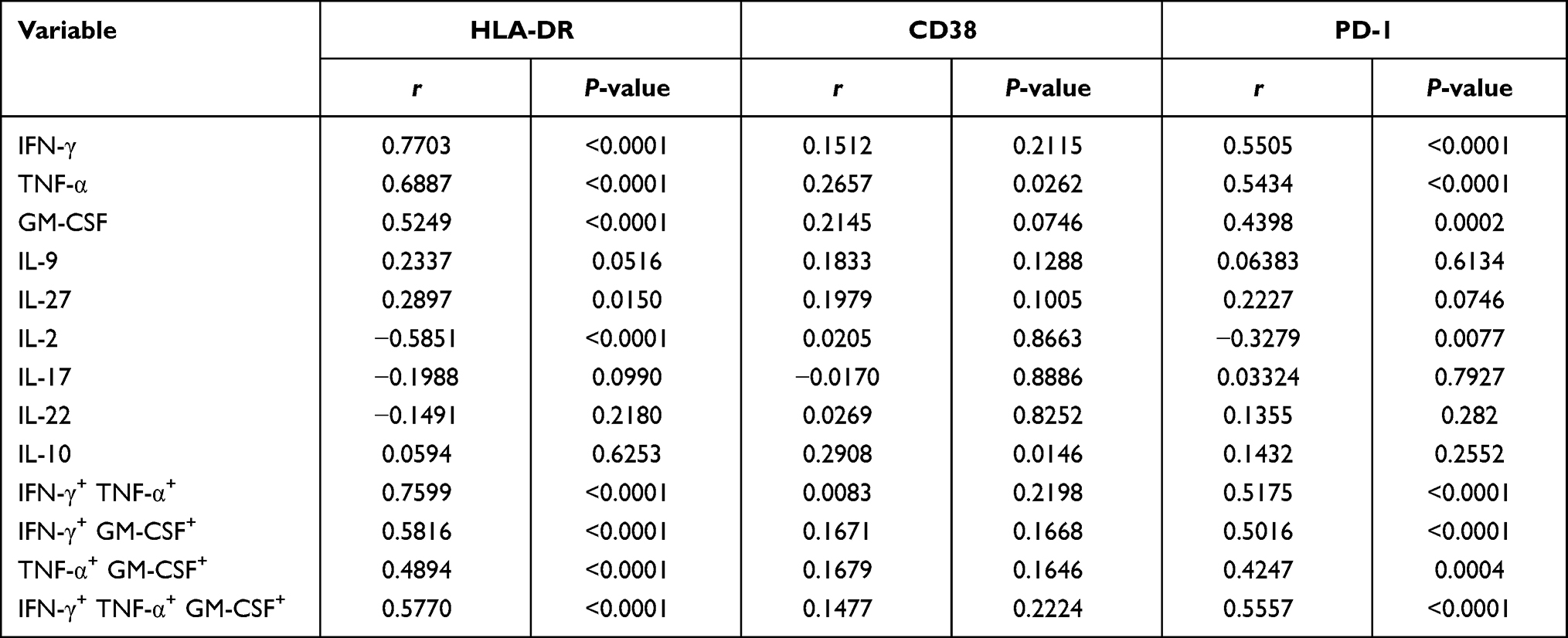 |
Table 2 Correlation Between Cytokine-Producing Cells and CD4 Immune Activation or Exhaustion Markers in ART-Naïve PLHIV |
In summary, these results showed that CD4+ T cells in PLHIV displayed a character of T-cell polyfunctionality and these strong PFC responses were a sign of the most severe stage during HIV infection (CD4 count <200 cells/µL).
Dysregulated Cytokine Secretion Was Associated with Loss of TN Fraction and the Increase of TCM and TEM CD4+ T Cells
CD4+ T cells are a heterogeneous population and could contain naïve T cells (TN, CD45RA+ CCR7+), central memory T cells (TCM, CD45RA− CCR7+), effector memory T cells (TEM, CD45RA− CCR7−), and a small fraction of terminally differentiated effector cells (TEMRA, CD45RA+ CCR7−). Prior studies have demonstrated that cytokine production capacity was also different in these T populations. In agreement with previous reports, we observed that PMA/Ionomycin induced high percentages of IFN-γ, TNF-α, and GM-CSF expression in both TCM and TEM fractions than TN fraction in healthy donors (Figure 3A). Since the dramatic decrease in both the percentages of TN fraction and absolute cell count in the PLHIV with CD4 count <200 cells/µL (Figure 3B and C), we wondered that whether the enhanced capacity of cytokine-production in PLHIV might be a result of the loss of TN fraction. To test the above hypothesis, we investigated the capacity of cytokine production for each CD4+ T-cell fraction. As shown in Figure 3B, the percentage of TEMRA CD4+ T cells was meager, and the following study would not be covering that subset.
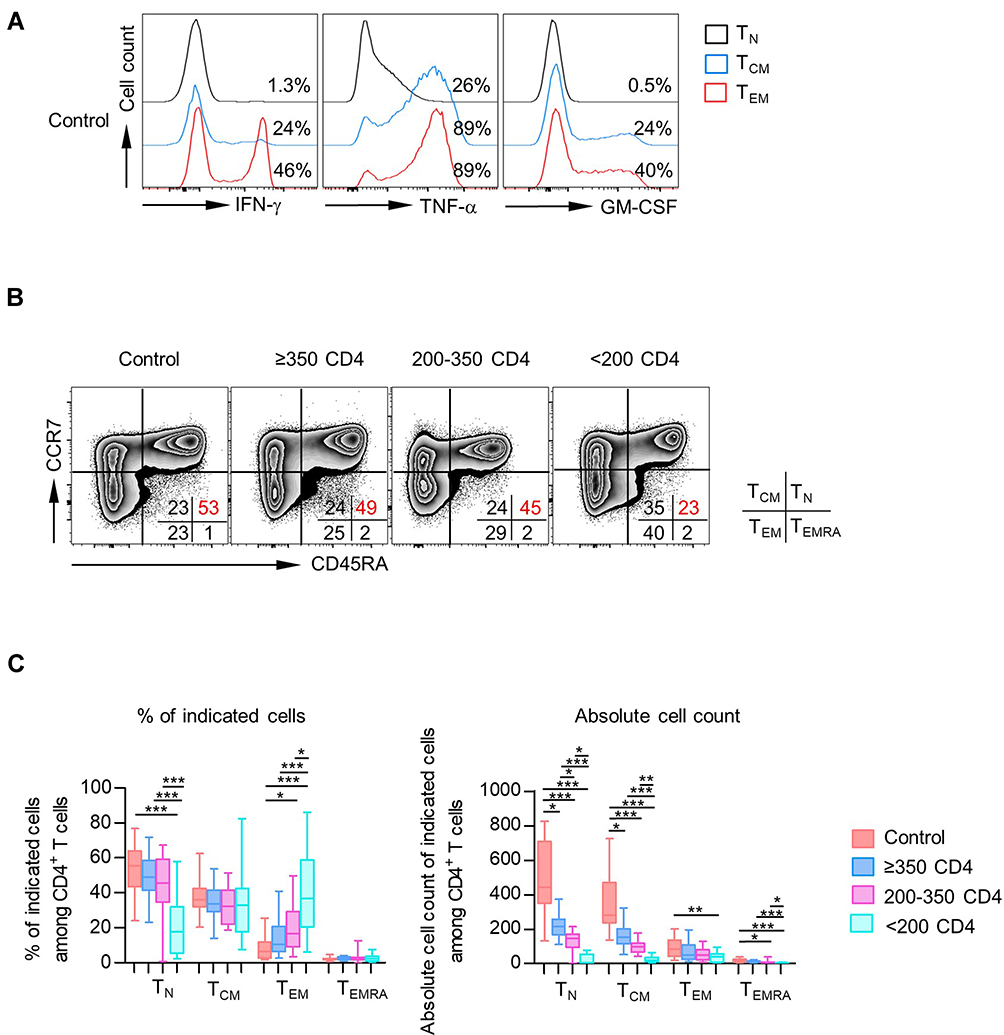 |
Figure 3 CD4 TCM and TEM populations were primary subsets of cytokine-producing CD4+ T cells. (A) Expression of IFN-γ, TNF-α, and GM-CSF among each subset (TN, TCM, and TEM) of CD4+ T cells was analyzed. Representative histograms displayed the expression of IFN-γ, TNF-α, and GM-CSF among each subset of CD4+ T cells from healthy donors. (B) Distribution of TN, TCM, TEM, and TEMRA in CD4+ T cells from healthy donors vs PLHIV. Representative flow data gated on CD4+ T cells of the percentage of each subset in healthy donors or PLHIV are shown. (C) Box plots of the frequencies (left) and absolute numbers (right) of different CD4+ T-cell compartments (TN, TCM, TEM, TEMRA) from healthy controls and PLHIV groups (CD4 count≥350 cells/µL, 200–350 cells/µL, <200 cells/µL). P values were obtained by Kruskal–Wallis test followed by Dunn’s multiple comparisons test. *P < 0.05, **P < 0.01, ***P < 0.001. |
We observed comparable IFN-γ, GM-CSF, and TNF-α frequencies in TN cells from PLHIV and healthy controls. In line with the above observation, most proliferation concentrated in the CD45RA− memory/effector populations of CD4+ T cells in healthy controls and PLHIV. Strikingly, both TCM and TEM from PLHIV displayed decreased percentages of cells producing IFN-γ, TNF-α, and GM-CSF compared with healthy controls (Figure 4A–C). Similar results were found on IL-17- and IL-22-producing T cells (Figure S4A and B), and no significant change was observed in the expression of IL-9, IL-27, IL-10, and IL-2 (Figure S4C–F).
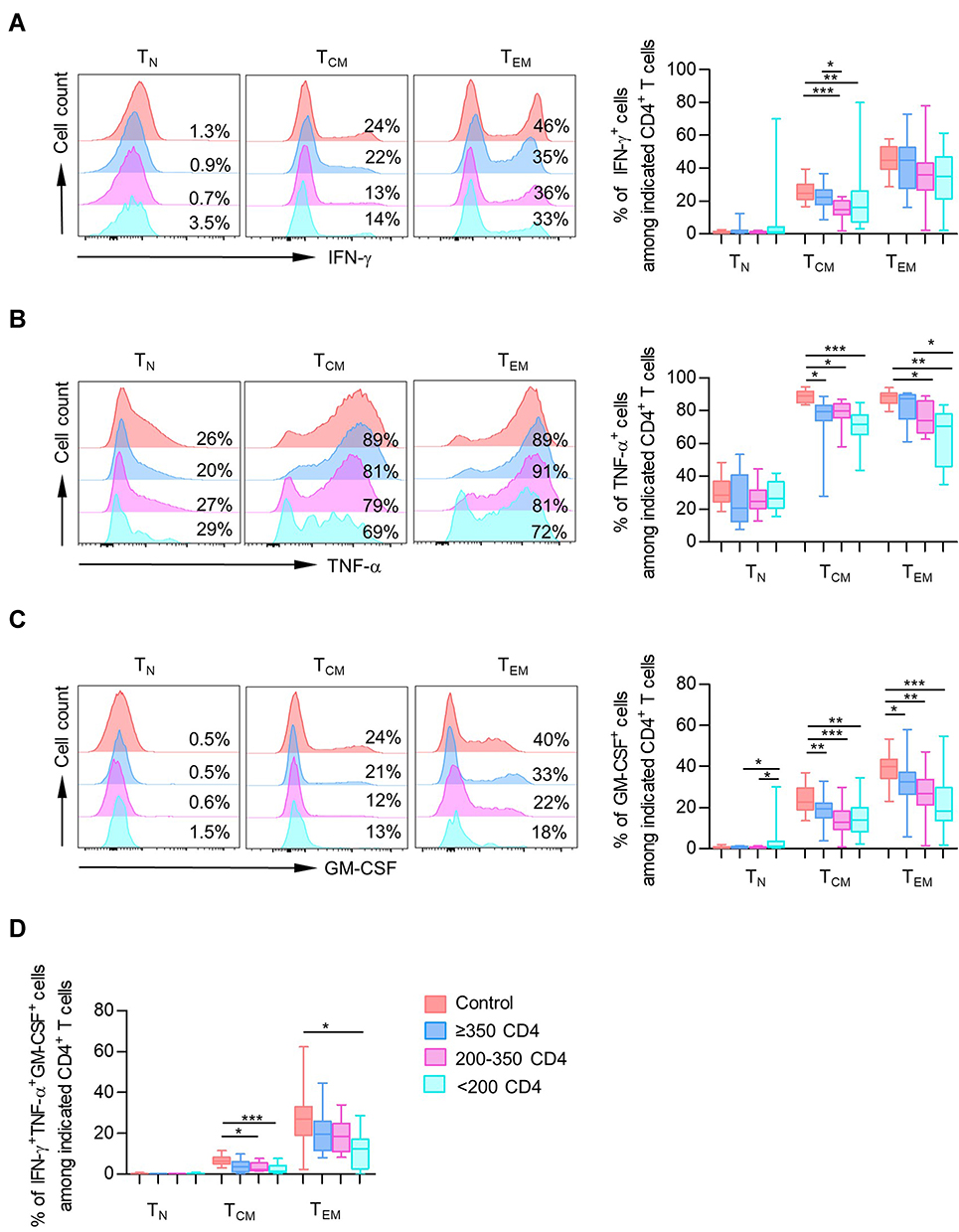 |
Figure 4 CD4+ T-cell subsets showed impaired production of pro-inflammation cytokines. (A–C) Data were represented as histograms (left) and box plots (right) comparing median frequency in CD4+ cytokine-producing (IFN-γ, TNF-α, and GM-CSF) cells in different CD4+ T-cell subsets (TN, TCM, TEM) among healthy controls and different PLHIV groups (CD4 count≥350 cells/µL, 200–350 cells/µL, <200 cells/µL). (D) Data were represented as box plots comparing median frequency in triple-positive producing CD4+ T cells in different CD4+ T-cell subsets among healthy controls and different PLHIV groups. P values were obtained by Kruskal–Wallis test followed by Dunn’s multiple comparisons test. *P < 0.05, **P < 0.01, ***P < 0.001. |
Consistent with the analysis of individual cytokine production, TCM and TEM were the main subsets contributing to polyfunctional responses (Figures 4D and S4G). Compared with healthy individuals, we also observed a declined proportion of IFN-γ+ TNF-α+ GM-CSF+ cells among TCM and TEM fractions in PLHIV (Figure 4D). Thus, CD4+ T-cell PFC responses were also impaired in PLHIV.
Indeed, we find that single or multiple IFN-γ, TNF-α, and GM-CSF expression was negatively correlated with the frequency of naïve CD4+ T cells and was positively correlated with CD4 TCM and TEM populations (Figure 5). These findings showed impaired production of pro-inflammation cytokines in CD4+ T-cell subsets, which was contradictory with that in total CD4+ T cells. It could be explained by the expansion of major cytokine-producing memory/effector T subsets as well as a severe loss of TN fraction despite impaired cytokine secretion in each subset of CD4+ T cells.
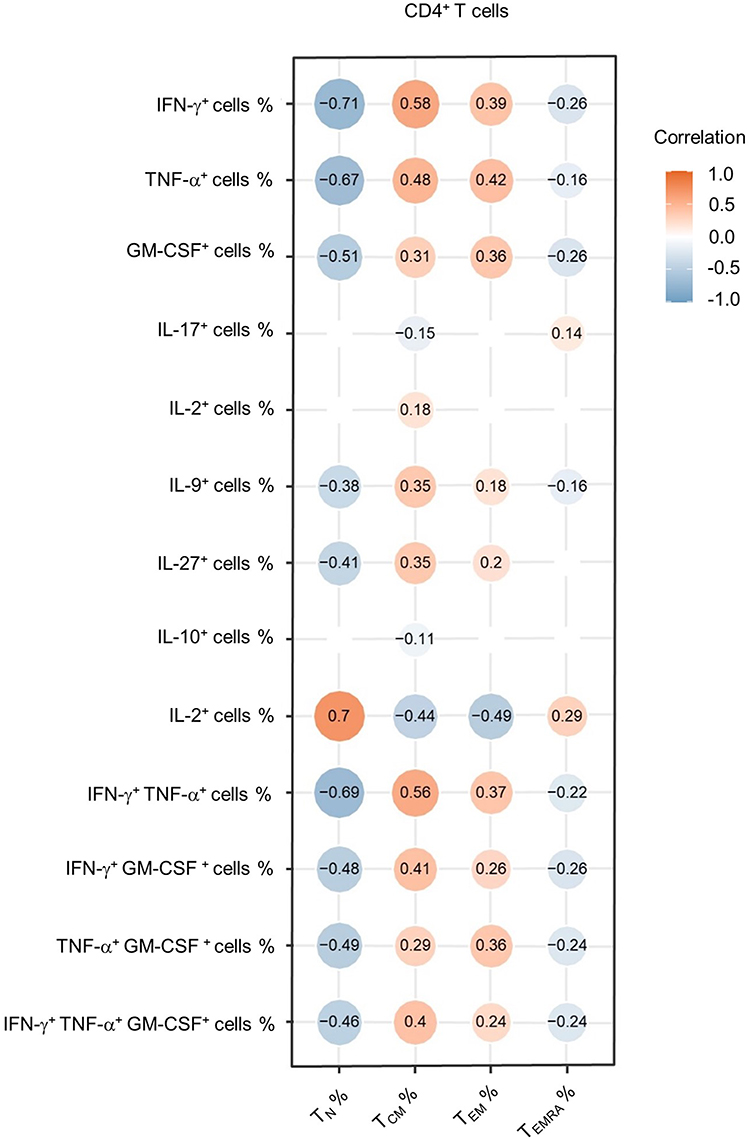 |
Figure 5 Correlation between cytokine-producing CD4+ T cells and CD4 subsets. Heatmap of correlation coefficients for percentages of cytokine-producing cells and percentages of CD4+ T-cell subsets (TN, TCM, TEM, TEMRA) among CD4+ T cells from PLHIV. The data shown are representative of 43 PLHIV with complete data. The circle size is proportional to the correlation coefficient value. Blue circle: negative correlation; Orange circle: positive correlation; values with no significant correlation are not displayed. |
ART Could Not Restore the Cytokine Secretion of CD4+ T-Cell Subsets in PBMCs
Given that antiretroviral therapy (ART) in the treatment of HIV has substantially reduced the morbidity and mortality of PLHIV by inhibiting viral replication and immune disorders,29 we then investigated how ART influences the frequency of cytokine-producing CD4+ T cells. Since PLHIV with low baseline CD4 count displayed low frequencies of inflammatory cytokines in CD4 subsets, we selected 20 ART-naïve PLHIV with a baseline CD4 count lower than 200 cells/μL and 34 PLHIV who experienced 3-year ART with matched baseline CD4 count. Supplementary Table 1 depicts the individuals’ demographic and clinical characteristics. As shown in Figure S5, ART could not alter cytokine secretion in both CD4+ T-cell subsets and total CD4+ T cells from these selected PLHIV. To determine whether ART could effectively restore the cytokine secretion of CD4+ T cells in immunological responders (IRs),30,31 we further identified 12 PLHIV with matched low baseline CD4 count who achieved successful immune reconstitution after 3 years of ART. Of note, the frequencies of IFN-γ+ producing CD4+ T-cell subsets were unchanged throughout treatment. Surprisingly, we observed a decline of TNF-α+ cells among TN and TCM fractions and decreased GM-CSF+ cells in the TN population (Figure 6A). Additionally, along with the altered ratio of CD4+ T-cell subsets after 3-year ART, the proportions of TNF-α+ and GM-CSF+ cells in total CD4+ T cells were significantly down-regulated in IRs (Figure 6B–C).
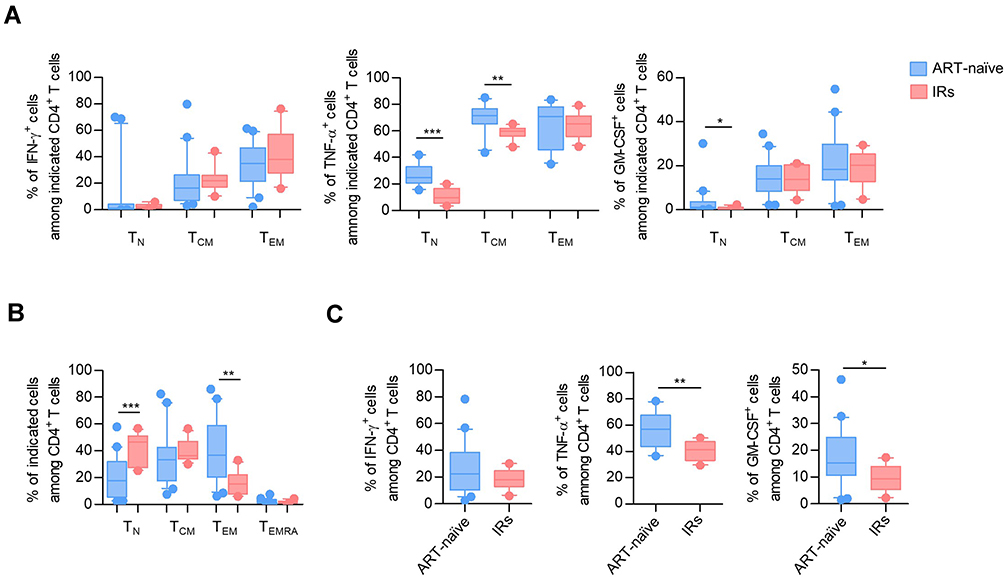 |
Figure 6 ART was instrumental in recovering naïve T cells but could not restore the function of cytokine secretion in CD4+ T-cell subsets. (A) Box plots of the frequencies of IFN-γ+, TNF-α+, and GM-CSF+ cells in TN, TCM, TEM CD4+ T cells from ART-naïve PLHIV with baseline CD4 count<200 cells/μL and IRs who had CD4+ T cell counts above 350 cells/μL after 3-year ART with matched baseline CD4 cell count. P values were obtained by the Kruskal–Wallis test, followed by Dunn’s multiple comparisons test. (B) The frequencies of TN, TCM, TEM, TEMRA CD4+ T cells in ART-naïve PLHIV and IRs. P values were obtained by the Kruskal–Wallis test, followed by Dunn’s multiple comparisons test. (C) The expression of IFN-γ, TNF-α, and GM-CSF in total CD4+ T cells from ART-naïve individuals and IRs. P values were obtained by unpaired t-test. *P < 0.05, **P < 0.01, ***P < 0.001. |
Collectively, ART was instrumental in recovering naïve T-cell subsets but did not restore the function of cytokine secretion in CD4+ T-cell subsets. Moreover, some functions of CD4+ T-cell subsets, such as TNF-α and GM-CSF secretion, became worse after ART.
Discussion
Previous studies have described discordant results of the CD4+ T-cell responses during chronic HIV infections. In the present study, we demonstrated the impaired cytokine secretion capacity such as IFN-γ, TNF-α, and GM-CSF in effector/memory CD4 subsets due to HIV disease progression. In contrast, total CD4+ T cells exhibited elevated production of these pro-inflammation cytokines. However, T cells are heterogeneous, consist of subpopulations with diverse cytokine production. These contradictory results in total and subsets of CD4+ T cells could be explained by the skewed T-cell subsets caused by loss of naïve cells and expansion of memory/effector population in PLHIV. Since the frequency of prominent cytokine-producing T subsets, TCM and TEM, remarkedly increased in PLHIV, total CD4+ T cells produced elevated cytokine despite decreased cytokine secretion in each subset of T cells. These data explained the contradictory findings in previous studies and suggested that we need to evaluate the response of CD4+ T cells at a subset level during HIV infection.
It has been demonstrated that chronic HIV infection is associated with establishing high and persistent levels of immune activation and inflammation, which contributes to disease progression and immune dysfunction.32,33 Innate cells such as natural killer cells, dendritic cells, and macrophages affected by HIV infection may cause high plasma levels of pro-inflammatory cytokines.34 Moreover, chronic activation of adaptive immunity, including CD8+ T-cell and CD4+ T-helper cell responses, is also vital against infections.35 Surprisingly, our results seemed inconsistent with these findings. We found that PLHIV harbored reduced inflammatory cytokine production and polyfunctional responses in CD4+ T-cell subsets. Accordingly, CD8+ T-cell dysfunction has also been demonstrated in various studies.36,37 Our results implied that effector cells of innate immunity, rather than activated T cells, might contribute to the systemic immune activation during HIV infection. The reduced amount and impaired function of regulatory T cells were demonstrated to facilitate immune hyperactivation during HIV infection.38,39 Of note, studies in mice models revealed that the insufficient number of T cells results in loss of control of innate immune responses, which presented a possible explanation for chronic activation of innate immunity in PLHIV.40,41 Furthermore, low-level inflammation due to innate immune cells might cause numerical depletion and progressive loss of CD4+ T cells’ functional responsiveness in a negative feedback loop. These findings supported the notion that the functional impairment and loss of CD4+ T cells leading to the increased risk of serious opportunistic infections. As such, we suggest that therapies able to reduce chronic activation of the innate immune system and preserve and/or enhance the functions of CD4+ T-cell subsets should be considered in the therapeutic regimen necessary for achieving HIV remission.
ART can suppress the virus in PLHIV to below the detection limit of the conventional clinical assay, improve CD4+ lymphocyte count and overall immunity.42,43 In the present study, we also showed that ART could restore the pool of naïve T cells in PLHIV and reconstituted their immune systems. Nevertheless, our data demonstrated that the function of cytokine secretion in CD4+ T-cell subsets could not be restored after 3 years of ART. It was consistent with others’ findings that 2 years of ART could not correct the dysfunctional phenotypes occurring in CD4+ and CD8+ T cells associated with HIV infection.44 Instead, we observed a further decline of Th1-related cytokine TNF-α and GM-CSF, indicating a persistent immune impairment. Morou et al indicated that the impaired functions of helper T cells might result from persistent changes in gene modules adapting to the environment.45 However, our previous study indicated that thymic output and CD4+ T-cell recovery could be further improved after long-term ART.46 Thus, a longer follow-up may be required after the ART to assess CD4+ T-cell function status. In addition, our results highlight the importance of developing novel, effective HIV treatments that can overcome the limitations of ART.
A key unresolved issue is the mechanism underlying the immune dysregulation of CD4+ T cells during HIV infection. Our study showed reduced production of pro-inflammation cytokines, including IFN-γ, TNF-α, and GM-CSF in CD4+ T-cell subsets. In addition, the polyfunctionality of CD4+ T-cell subsets to secrete more than one cytokine was also impaired in PLHIV. More importantly, we also observed a declined secretion of IL-17 and IL-22 in severely immunocompromised PLHIV, whose naïve CD4+ T cells were significantly reduced. Previous studies suggested that persistent virus infections (such as HBV, HCV, and HIV) are associated with a state of T-cell exhaustion in both CD4+ and CD8+ T cells.47,48 Exhausted T cells expressed multiple inhibitory receptors and exhibited functional impairment manifested by low cytokine production and impaired proliferation capacity.49,50 CD8+ T-cell exhaustion has been well described in chronic infection and tumor studies.36,37 During HIV infection, the focus of T cell exhaustion is often on CD8+ T cells that are primarily responsible for killing infected cells. However, it is now accepted that exhaustion is not restricted to CD8+ T cell responses as CD4+ T cells are also subject to exhaustion. Studies have demonstrated that exhausted CD4+ T cells shared similar features with CD8+ T cells during tumor and chronic viral infections. Goding et al showed tumor-specific CD4+ T cells upregulated PD-1, TIM-3, LAG-3, 2B4, and TIGIT, accompanied by reduced expression of IFN-γ and TNF-α.51 Moreover, CD4+ T cells with PD-1 and LAG-3 expression exhibited decreased cytokine production in chronic HBV infection, and treatment with PD-L1 and LAG-3 antibodies could restore CD4+ T-cell function.52 CD4+ helper T cells in PLHIV also expressed PD-1, CTLA-4, and TIM-3 and reduced IL-2 production,53–56 suggesting a similar exhausting state of CD4+ T cells. The notion was further confirmed by the present study, which showed over-activation and exhaustion of CD4+ T cells, accompanied by severe skewing of CD4+ T-cell subsets during HIV disease progression.
Our study has several limitations, including a lack of longitudinal data after ART and a heavily gender-skewed cohort. This analysis is almost entirely made up of men who have sex with men (MSM) due to the clinical status in our hospital. There have been essential distinctions reported between immune/inflammatory profiles in HIV-infected men vs women (generally more inflammatory in the latter).57 Therefore, more evidence was urgently needed to investigate whether the findings accurately reflect a more gender-balanced cohort.
Conclusions
Our study identified inconsistent results of cytokine secretion in total and subsets of CD4+ T cells. The capacity of cytokine production of each CD4+ T subset was impaired during HIV infection, and altered T-cell subsets were associated with pseudo-elevation of cytokine production in total CD4+ T cells. Moreover, ART could increase CD4 counts by restoring the pool of naïve T cells but could not improve the effector function of CD4+ T-cell subsets. Therefore, therapies that can preserve and/or enhance the function of CD4+ T cells should be considered as another target in strategies of HIV remission.
Data Sharing Statement
All data used to draw the conclusions in the paper are presented in the paper and/or the supplementary materials.
Ethics Approval
The study was approved by the Committee of Ethics at Beijing Ditan Hospital, Capital Medical University in Beijing, China.
Consent to Participate
Informed consents were obtained from all healthy controls and enrolled PLHIV.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Funding
This work was supported by grants from the Beijing Municipal Administration of Hospitals’ Ascent Plan (DFL20191802), Capital Clinical Characteristic Application (Z171100001017052), Beijing Municipal Administration of Hospitals Clinical Medicine Development of Special Funding Support (ZYLX202126), and the National Natural Science Foundation of China (81971307 and 81672000).
Disclosure
The authors declare that they have no conflicts of interest.
References
1. Saharia KK, Koup RA. T cell susceptibility to HIV influences outcome of opportunistic infections. Cell. 2013;155(3):505–514. doi:10.1016/j.cell.2013.09.045
2. Laidlaw BJ, Craft JE, Kaech SM. The multifaceted role of CD4(+) T cells in CD8(+) T cell memory. Nat Rev Immunol. 2016;16(2):102–111. doi:10.1038/nri.2015.10
3. Boucau J, Madouasse J, Kourjian G, et al. The activation state of CD4 T cells alters cellular Peptidase activities, HIV antigen processing, and MHC class I presentation in a sequence-dependent manner. J Immunol. 2019;202(10):2856–2872. doi:10.4049/jimmunol.1700950
4. Porichis F, Hart MG, Massa A, et al. Immune checkpoint blockade restores HIV-specific CD4 T cell help for NK cells. J Immunol. 2018;201(3):971–981. doi:10.4049/jimmunol.1701551
5. Catalfamo M, Le Saout C, Lane HC. The role of cytokines in the pathogenesis and treatment of HIV infection. Cytokine Growth Factor Rev. 2012;23(4–5):207–214. doi:10.1016/j.cytogfr.2012.05.007
6. Klein SA, Dobmeyer JM, Dobmeyer TS, et al. Demonstration of the Th1 to Th2 cytokine shift during the course of HIV-1 infection using cytoplasmic cytokine detection on single cell level by flow cytometry. AIDS. 1997;11(9):1111–1118. doi:10.1097/00002030-199709000-00005
7. Romagnani S. Biology of human TH1 and TH2 cells. J Clin Immunol. 1995;15(3):121–129. doi:10.1007/BF01543103
8. Altfeld M, Addo MM, Kreuzer KA, et al. T(H)1 to T(H)2 shift of cytokines in peripheral blood of HIV-infected patients is detectable by reverse transcriptase polymerase chain reaction but not by enzyme-linked immunosorbent assay under nonstimulated conditions. J Acquir Immune Defic Syndr. 2000;23(4):287–294. doi:10.1097/00126334-200004010-00001
9. Sindhu S, Toma E, Cordeiro P, Ahmad R, Morisset R, Menezes J. Relationship of in vivo and ex vivo levels of TH1 and TH2 cytokines with viremia in HAART patients with and without opportunistic infections. J Med Virol. 2006;78(4):431–439. doi:10.1002/jmv.20558
10. Bahbouhi B, Landay A, Al-Harthi L. Dynamics of cytokine expression in HIV productively infected primary CD4+ T cells. Blood. 2004;103(12):4581–4587. doi:10.1182/blood-2003-12-4172
11. Santinelli L, Statzu M, Pierangeli A, et al. Increased expression of IL-32 correlates with IFN-γ, Th1 and Tc1 in virologically suppressed HIV-1-infected patients. Cytokine. 2019;120:273–281. doi:10.1016/j.cyto.2019.01.012
12. Reuter MA, Pombo C, Betts MR. Cytokine production and dysregulation in HIV pathogenesis: lessons for development of therapeutics and vaccines. Cytokine Growth Factor Rev. 2012;23(4–5):181–191. doi:10.1016/j.cytogfr.2012.05.005
13. Végran F, Berger H, Boidot R, et al. The transcription factor IRF1 dictates the IL-21-dependent anticancer functions of TH9 cells. Nat Immunol. 2014;15(8):758–766. doi:10.1038/ni.2925
14. Klatt NR, Estes JD, Sun X, et al. Loss of mucosal CD103+ DCs and IL-17+ and IL-22+ lymphocytes is associated with mucosal damage in SIV infection. Mucosal Immunol. 2012;5(6):646–657. doi:10.1038/mi.2012.38
15. Fakruddin JM, Lempicki RA, Gorelick RJ, et al. Noninfectious papilloma virus-like particles inhibit HIV-1 replication: implications for immune control of HIV-1 infection by IL-27. Blood. 2007;109(5):1841–1849. doi:10.1182/blood-2006-02-001578
16. Brenchley JM, Paiardini M, Knox KS, et al. Differential Th17 CD4 T-cell depletion in pathogenic and nonpathogenic lentiviral infections. Blood. 2008;112(7):2826–2835. doi:10.1182/blood-2008-05-159301
17. Micci L, Cervasi B, Ende ZS, et al. Paucity of IL-21-producing CD4(+) T cells is associated with Th17 cell depletion in SIV infection of rhesus macaques. Blood. 2012;120(19):3925–3935. doi:10.1182/blood-2012-04-420240
18. Gorenec L, Zidovec Lepej S, Grgic I, et al. The comparison of Th1, Th2, Th9, Th17 and Th22 cytokine profiles in acute and chronic HIV-1 infection. Microb Pathog. 2016;97:125–130. doi:10.1016/j.micpath.2016.06.008
19. Imamichi T, Yang J, Huang DW, et al. IL-27, a novel anti-HIV cytokine, activates multiple interferon-inducible genes in macrophages. AIDS. 2008;22(1):39–45. doi:10.1097/QAD.0b013e3282f3356c
20. Dai L, Lidie KB, Chen Q, et al. IL-27 inhibits HIV-1 infection in human macrophages by down-regulating host factor SPTBN1 during monocyte to macrophage differentiation. J Exp Med. 2013;210(3):517–534. doi:10.1084/jem.20120572
21. Ciuffreda D, Comte D, Cavassini M, et al. Polyfunctional HCV-specific T-cell responses are associated with effective control of HCV replication. Eur J Immunol. 2008;38(10):2665–2677. doi:10.1002/eji.200838336
22. Qiu Z, Zhang M, Zhu Y, et al. Multifunctional CD4 T cell responses in patients with active tuberculosis. Sci Rep. 2012;2(1):216. doi:10.1038/srep00216
23. Casazza JP, Betts MR, Price DA, et al. Acquisition of direct antiviral effector functions by CMV-specific CD4+ T lymphocytes with cellular maturation. J Exp Med. 2006;203(13):2865–2877. doi:10.1084/jem.20052246
24. Duvall MG, Precopio ML, Ambrozak DA, et al. Polyfunctional T cell responses are a hallmark of HIV-2 infection. Eur J Immunol. 2008;38(2):350–363. doi:10.1002/eji.200737768
25. Lacy P, Stow JL. Cytokine release from innate immune cells: association with diverse membrane trafficking pathways. Blood. 2011;118(1):9–18. doi:10.1182/blood-2010-08-265892
26. Geginat J, Paroni M, Facciotti F, et al. The CD4-centered universe of human T cell subsets. Semin Immunol. 2013;25(4):252–262. doi:10.1016/j.smim.2013.10.012
27. Akase IE, Musa B, Obiako RO, Ahmad Elfulatiy A, Mohammed AA. Immune dysfunction in HIV: a possible role for pro- and anti-inflammatory cytokines in HIV staging. J Immunol Res. 2017;2017:4128398. doi:10.1155/2017/4128398
28. Godfried MH, van der Poll T, Weverling GJ, et al. Soluble receptors for tumor necrosis factor as predictors of progression to AIDS in asymptomatic human immunodeficiency virus type 1 infection. J Infect Dis. 1994;169(4):739–745. doi:10.1093/infdis/169.4.739
29. Lederman MM, Funderburg NT, Sekaly RP, Klatt NR, Hunt PW. Residual immune dysregulation syndrome in treated HIV infection. Adv Immunol. 2013;119:51–83. doi:10.1016/B978-0-12-407707-2.00002-3
30. Tincati C, Merlini E, Braidotti P, et al. Impaired gut junctional complexes feature late-treated individuals with suboptimal CD4+ T-cell recovery upon virologically suppressive combination antiretroviral therapy. AIDS. 2016;30(7):991–1003. doi:10.1097/QAD.0000000000001015
31. Younes SA, Talla A, Pereira Ribeiro S, et al. Cycling CD4+ T cells in HIV-infected immune nonresponders have mitochondrial dysfunction. J Clin Invest. 2018;128(11):5083–5094. doi:10.1172/JCI120245
32. Ipp H, Zemlin AE, Erasmus RT, Glashoff RH. Role of inflammation in HIV-1 disease progression and prognosis. Crit Rev Clin Lab Sci. 2014;51(2):98–111. doi:10.3109/10408363.2013.865702
33. Appay V, Sauce D. Immune activation and inflammation in HIV-1 infection: causes and consequences. J Pathol. 2008;214(2):231–241. doi:10.1002/path.2276
34. Boasso A, Shearer GM. Chronic innate immune activation as a cause of HIV-1 immunopathogenesis. Clin Immunol. 2008;126(3):235–242. doi:10.1016/j.clim.2007.08.015
35. Klatt NR, Chomont N, Douek DC, Deeks SG. Immune activation and HIV persistence: implications for curative approaches to HIV infection. Immunol Rev. 2013;254(1):326–342. doi:10.1111/imr.12065
36. Kostense S, Vandenberghe K, Joling J, et al. Persistent numbers of tetramer+ CD8(+) T cells, but loss of interferon-gamma+ HIV-specific T cells during progression to AIDS. Blood. 2002;99(7):2505–2511. doi:10.1182/blood.v99.7.2505
37. Lieberman J, Manjunath N, Shankar P. Avoiding the kiss of death: how HIV and other chronic viruses survive. Curr Opin Immunol. 2002;14(4):478–486. doi:10.1016/s0952-7915(02)00366-7
38. Sakaguchi S. Control of immune responses by naturally arising CD4+ regulatory T cells that express toll-like receptors. J Exp Med. 2003;197(4):397–401. doi:10.1084/jem.20030012
39. Eggena MP, Barugahare B, Jones N, et al. Depletion of regulatory T cells in HIV infection is associated with immune activation. J Immunol. 2005;174(7):4407–4414. doi:10.4049/jimmunol.174.7.4407
40. Guarda G, Dostert C, Staehli F, et al. T cells dampen innate immune responses through inhibition of NLRP1 and NLRP3 inflammasomes. Nature. 2009;460(7252):269–273. doi:10.1038/nature08100
41. Kim KD, Zhao J, Auh S, et al. Adaptive immune cells temper initial innate responses. Nat Med. 2007;13(10):1248–1252. doi:10.1038/nm1633
42. Nakagawa F, May M, Phillips A. Life expectancy living with HIV: recent estimates and future implications. Curr Opin Infect Dis. 2013;26(1):17–25. doi:10.1097/QCO.0b013e32835ba6b1
43. May M, Gompels M, Delpech V, et al. Impact of late diagnosis and treatment on life expectancy in people with HIV-1: UK Collaborative HIV Cohort (UK CHIC) Study. BMJ. 2011;343:d6016. doi:10.1136/bmj.d6016
44. Amu S, Lantto Graham R, Bekele Y, et al. Dysfunctional phenotypes of CD4+ and CD8+ T cells are comparable in patients initiating ART during early or chronic HIV-1 infection. Medicine. 2016;95(23):e3738. doi:10.1097/MD.0000000000003738
45. Morou A, Brunet-Ratnasingham E, Dubé M, et al. Altered differentiation is central to HIV-specific CD4+ T cell dysfunction in progressive disease. Nat Immunol. 2019;20(8):1059–1070. doi:10.1038/s41590-019-0418-x
46. Kong Y, Tian Y, Hao Y, et al. Two types of poor immunological responder showing distinct responses to long-term HAART. Int J Infect Dis. 2019;86:178–187. doi:10.1016/j.ijid.2019.07.037
47. Khaitan A, Unutmaz D. Revisiting immune exhaustion during HIV infection. Curr HIV/AIDS Rep. 2011;8(1):4–11. doi:10.1007/s11904-010-0066-0
48. Wherry EJ, Kurachi M. Molecular and cellular insights into T cell exhaustion. Nat Rev Immunol. 2015;15(8):486–499. doi:10.1038/nri3862
49. Crawford A, Angelosanto JM, Kao C, et al. Molecular and transcriptional basis of CD4+ T cell dysfunction during chronic infection. Immunity. 2014;40(2):289–302. doi:10.1016/j.immuni.2014.01.005
50. Wherry EJ. T cell exhaustion. Nat Immunol. 2011;12(6):492–499. doi:10.1038/ni.2035
51. Goding SR, Wilson KA, Xie Y, et al. Restoring immune function of tumor-specific CD4+ T cells during recurrence of melanoma. J Immunol. 2013;190(9):4899–4909. doi:10.4049/jimmunol.1300271
52. Dong Y, Li X, Zhang L, et al. CD4+ T cell exhaustion revealed by high PD-1 and LAG-3 expression and the loss of helper T cell function in chronic hepatitis B. BMC Immunol. 2019;20(1):27. doi:10.1186/s12865-019-0309-9
53. Kassu A, Marcus RA, D’Souza MB, et al. Regulation of virus-specific CD4+ T cell function by multiple costimulatory receptors during chronic HIV infection. J Immunol. 2010;185(5):3007–3018. doi:10.4049/jimmunol.1000156
54. Younes SA, Yassine-Diab B, Dumont AR, et al. HIV-1 viremia prevents the establishment of interleukin 2-producing HIV-specific memory CD4+ T cells endowed with proliferative capacity. J Exp Med. 2003;198(12):1909–1922. doi:10.1084/jem.20031598
55. D’Souza M, Fontenot AP, Mack DG, et al. Programmed death 1 expression on HIV-specific CD4+ T cells is driven by viral replication and associated with T cell dysfunction. J Immunol. 2007;179(3):1979–1987. doi:10.4049/jimmunol.179.3.1979
56. Kaufmann DE, Kavanagh DG, Pereyra F, et al. Upregulation of CTLA-4 by HIV-specific CD4+ T cells correlates with disease progression and defines a reversible immune dysfunction. Nat Immunol. 2007;8(11):1246–1254. doi:10.1038/ni1515
57. Griesbeck M, Scully E, Altfeld M. Sex and gender differences in HIV-1 infection. Clin Sci. 2016;130(16):1435–1451. doi:10.1042/CS20160112
 © 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2021 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.