Back to Journals » Nature and Science of Sleep » Volume 17

Acute Sleep Deprivation Suppresses Sleep Spindles in Mice
Authors Xu Y, Tan M, Li D, Zhang C
Received 1 July 2025
Accepted for publication 23 September 2025
Published 9 October 2025 Volume 2025:17 Pages 2627—2637
DOI https://doi.org/10.2147/NSS.S549263
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Dr Marco Veneruso
Yan Xu,1 Miaoqin Tan,1 Dongcai Li,2 Chenxi Zhang2
1Department of Psychiatry, Nanfang Hospital, Southern Medical University, Guangzhou, Guangdong, People’s Republic of China; 2Department of Psychiatry, Longgang Otolaryngology Hospital and Shenzhen Key Laboratory of Otolaryngology, Shenzhen Institute of Otolaryngology, Shenzhen, Guangdong, People’s Republic of China
Correspondence: Chenxi Zhang, Email [email protected]
Introduction: Sleep spindles and cortical coherence are key electrophysiological signatures of thalamocortical communication and large-scale neural synchrony during non-REM sleep. This study aimed to evaluate the effects of acute 12-hour sleep deprivation on spindle dynamics and cortical coherence across frequency bands in mice.
Methods: Sixteen adult male C57BL/6J (8 per group) mice were randomly assigned to either a natural sleep (NS) or sleep deprivation (SD) group. We recorded bilateral frontal EEGs continuously for 12 hours following intervention. Sleep spindles (8– 16 Hz) were automatically detected, and interhemispheric coherence from delta to gamma bands was computed using Welch’s method.
Results: Compared to NS mice, SD mice showed a significant reduction in frontal spindle count (61 ± 14 vs 103 ± 12 spindles; mean ± SEM; p = 0.027) and amplitude (47 ± 6 μV vs 68 ± 5 μV; p = 0.002). Delta (0.5– 4 Hz) and theta (4– 8 Hz) interhemispheric coherence showed non-significant decreasing trends in SD mice (p > 0.05), whereas alpha (8– 13 Hz), beta (13– 30 Hz), and gamma (30– 40 Hz) coherence did not differ.
Conclusion: Acute sleep deprivation significantly impairs spindle generation. It also resulted in a trend toward reduced low-frequency cortical coherence, though this change was not statistically significant.
Keywords: sleep deprivation, sleep spindles, cortical coherence, acute sleep deprivation
Introduction
Sleep is characterized by rhythmic oscillations that play essential roles in neural plasticity and cognition. Among these, sleep spindles—~12 Hz rhythmic bursts during NREM sleep—originate in thalamocortical networks and propagate across widespread cortical areas.1,2 Thalamic reticular neurons and thalamocortical relay cells alternately drive spindling activity, generating cortical states that favor synaptic plasticity.2 Sleep spindles play critical roles in synaptic plasticity and memory consolidation, reflecting the strength of thalamocortical loops.3–6 They are homeostatically regulated by prior wakefulness and increase before NREM–REM transitions.7 Both rodent and human studies show that acute sleep loss diminishes spindle occurrence and alters spindle morphology, linking spindle dynamics to learning and cognitive outcomes.8–11 These findings highlight the importance of spindles as biomarkers of thalamocortical network function.12,13
High coherence at a given frequency means that distant neuronal populations are oscillating in a tightly locked fashion, which facilitates coordinated information transfer and network stability.13,14 During healthy sleep, coherence is particularly pronounced at low frequencies: for example, sleep spindles and slow waves show strong bilateral coherence, reflecting their widespread and quasi-synchronous generation.8,14 Homologous interhemispheric coherence is greatest in the delta and spindle bands and steadily declines at higher frequencies,15 indicating that slow oscillations engage most of cortex simultaneously while faster rhythms are more local. Such global synchronization of delta/theta activity during NREM sleep is thought to support synaptic downscaling and consolidation of memory traces between cortical regions. Prolonged sleep deprivation impairs these aspects of neural synchrony. In sleep-deprived humans, functional imaging and EEG studies reveal widespread connectivity breakdown. For example, resting-state EEG after total sleep loss shows a marked reduction in alpha-band power and weakened long-range alpha coherence, especially across default-mode regions.16 These connectivity changes correlate with deficits in attention, vigilance and executive function observed after sleep loss. In rodents, sleep deprivation likewise alters cross-hemispheric coherence, though effects depend on frequency and timing. Notably, normal C57 mice exhibit a transient increase in interhemispheric delta coherence during the first two hours of recovery NREM after 6h deprivation (reflecting rebound slow-wave synchrony).17 However, this rebound is absent in acallosal “B1” mutants, underscoring that the corpus callosum mediates most hemispheric EEG synchrony.8 Despite these findings, comprehensive data on how acute sleep loss affects EEG coherence across multiple bands are scarce. Some rat and human studies report post-deprivation changes in alpha coherence (consistent with attentional network disruption),18,19 but effects on higher-frequency bands (beta, gamma) remain inconsistent and understudied. Moreover, most animal work has examined only short (≤6h) sleep deprivation or used single-site recordings, limiting insight into full-spectrum, bilateral cortical synchrony after extended wakefulness.
Given these gaps, we sought to clarify how extended acute sleep deprivation impacts both spindle dynamics and interhemispheric coordination in a standard mouse model. Specifically, we applied a 12-hour gentle-handling deprivation during the light phase in adult male C57BL/6J mice and recorded frontal EEG activity. Our study is unique in using a prolonged 12-hour sleep deprivation (double the duration of the more common 6h protocols) and in assessing bilateral (interhemispheric) EEG coherence across multiple frequency bands in mice. We quantified changes in frontal sleep spindle density, amplitude, and frequency, and calculated interhemispheric coherence in canonical bands (delta through gamma) between left and right frontal electrodes. We hypothesized that prolonged sleep loss would suppress spindle generation and weaken EEG coherence, especially in low-frequency bands, reflecting disrupted thalamocortical oscillations and large-scale network synchrony. This systematic approach will reveal the frequency-specific effects of acute sleep deprivation on cortical oscillations, shedding light on the network mechanisms by which sleep loss may impair cognitive functions.
Sleep spindles are thalamocortical NREM oscillations that support synaptic plasticity and memory, reflecting coordinated activity between thalamus and cortex.20 Interhemispheric EEG coherence quantifies large-scale neural synchrony: high coherence at low frequencies (delta/theta/spindles) indicates widespread cortical coupling.21 Acute sleep deprivation disrupts these processes: it reduces spindle occurrence (with compensatory amplitude changes) and weakens long-range EEG connectivity (eg diminished alpha coherence).22 However, most prior work has used short (≤6h) deprivation or non-standard models, leaving the effects of extended (12h) sleep loss on spindles and coherence in C57BL/6J mice unresolved. This study addresses that gap by testing whether a 12h gentle-handling sleep deprivation in C57BL/6J mice impairs frontal spindle generation and reduces interhemispheric coherence across delta–gamma bands, advancing understanding of sleep-loss effects on cortical network homeostasis.
Methods
Animals and Housing
Sixteen adult male C57BL/6J mice (20–25 g; Jackson Laboratory) were used. Mice were housed individually in Plexiglas cages under a 12:12 h light/dark cycle (lights on at 07:00), with temperature 22 ± 2 °C and humidity 55 ± 5%. Food and water were provided ad libitum. All procedures complied with institutional and NIH guidelines and were approved by the Institutional Animal Care and Use Committee. Animals were allowed at least one week to acclimate to the housing environment prior to any experimental procedures. All animal procedures were approved by the Animal Ethics Committee of Nanfang Hospital of Southern Medical University on March 6, 2024. All experimental procedures were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1978) and the ARRIVE guidelines on the Care and Use of Experimental Animals.
Sleep Deprivation Procedure
Mice were randomly assigned to a Natural Sleep (NS) group (n = 8) or a Sleep Deprivation (SD) group (n = 8). SD mice were subjected to a 12-hour gentle-handling sleep deprivation beginning at lights-on (07:00) and lasting throughout the light phase. Gentle handling consisted of intermittent cage tapping, light nudges, and introduction of novel objects to maintain wakefulness without inducing stress-related behaviors. NS mice were left undisturbed in their home cages during the same period.
Surgical Implantation of EEG Electrodes
Prior to recording, mice were implanted with epidural EEG electrodes under deep anesthesia (pentobarbital, 50 mg/kg i.p). using a stereotaxic frame. Two stainless-steel screw electrodes were placed over the frontal cortex (anteroposterior +1.0 mm, mediolateral ±1.5 mm relative to bregma), yielding one electrode in each hemisphere. A reference electrode was positioned over the cerebellum (midline) and a separate cerebellar screw served as ground. Leads from the electrodes were secured to the skull with dental acrylic and connected to a miniature headstage. Postoperative care included subcutaneous buprenorphine (0.1 mg/kg) for analgesia, and mice were allowed ≥72 h to recover fully in their home cages before EEG recording.
EEG Acquisition and Signal Processing
Continuous EEG recordings were obtained using a multi-channel amplifier (Neuralynx, 32-channel) at a 1000 Hz sampling rate (16-bit precision). Signals were hardware band-pass filtered from 0.1 to 100 Hz during acquisition and referenced online to the cerebellar reference electrode (ground on cerebellum). Recordings spanned the full 12-hour light phase (07:00–19:00) following recovery. Offline analysis was performed in MATLAB (R2021b, MathWorks). The raw EEG signals were digitally filtered with a 0.1–40 Hz zero-phase FIR band-pass to remove slow drifts and high-frequency noise. The filtered data were segmented into consecutive non-overlapping 5-second epochs. Epochs containing artifacts (defined as any channel exceeding ±500 µV, typically due to gross movement) were automatically identified and excluded from further analysis. Data analysis was conducted with the investigator blinded to group (NS vs SD) to prevent observer bias in artifact rejection and event detection.
Spindle Detection
Sleep spindles were automatically identified in the frontal EEG channels using a multi-step algorithm. Frequency isolation: Each 5-s epoch of frontal EEG was band-pass filtered at 8–16 Hz (zero-phase Butterworth filter, 4th order) to isolate spindle-frequency activity. Envelope computation: The root-mean-square (RMS) amplitude envelope was calculated using a 100-ms sliding window across the filtered signal. Thresholding: A dynamic threshold was set at 1.5× the baseline RMS amplitude. The baseline was defined as the average RMS during all artifact-free NREM sleep epochs for each mouse. Spindle candidates were segments where the RMS envelope exceeded this threshold. Duration and morphology criteria: Each candidate event was required to last between 0.4 and 3.0 seconds and to include at least six zero-crossings in the 8–16 Hz filtered signal, ensuring the oscillatory nature of true spindles. Metric extraction: For each detected spindle, the peak-to-peak amplitude was measured from the raw EEG trace within the event window. Total spindle count and the mean peak amplitude were computed for each mouse over the 12-hour recording. For waveform context, representative raw EEG traces from Natural Sleep (NS) and Sleep Deprivation (SD) groups are shown in Supplementary Figure S1.
Coherence Analysis
Interhemispheric EEG coherence between the left and right frontal electrodes was calculated using Welch’s method. We chose Welch’s method (averaging spectra from overlapping segments) to estimate coherence because it reduces the variance of spectral estimates and yields more reliable coherence values than single-epoch Fourier transforms. This approach is standard in EEG analyses to improve signal-to-noise ratio for power and coherence measures. The analysis steps were: Segmentation: Each artifact-free 5-s epoch was divided into overlapping segments of 256 samples (256 ms windows) with 50% overlap and Hamming windowing. Spectral estimation: For each segment, we computed the power spectral density (PSD) of each channel and the cross-spectral density between channels using a 256-point fast Fourier transform (FFT). Coherence computation: The magnitude-squared coherence at each frequency f was obtained by |S_xy(f)|^2/(S_xx(f)·S_yy(f)), where S_xy is the cross-spectrum and S_xx, S_yy are the auto-spectra of the two signals. Coherence values range from 0 (no phase coupling) to 1 (perfect synchrony). Averaging: Coherence spectra were averaged across all segments and epochs to yield a mean coherence spectrum (0.5–40 Hz) for each animal. The mean coherence was then calculated within five standard frequency bands: delta (0.5–4 Hz), theta (4–8 Hz), alpha (8–13 Hz), beta (13–30 Hz), and gamma (30–40 Hz). MATLAB’s built-in mscohere function (MathWorks) was used for these computations.
Statistical Analysis
All data are presented as group means ± standard error of the mean (SEM). Statistical analyses were conducted using R (version 4.2.0; R Core Team, 2022). Group differences between NS and SD mice in spindle count, spindle amplitude, and coherence across frequency bands were assessed using two-tailed independent-samples t-tests. The Shapiro–Wilk test was used to confirm normality, and Levene’s test assessed homogeneity of variances; Welch’s correction was applied where variances were unequal. Effect sizes were calculated using Cohen’s d (based on pooled standard deviation). A p-value < 0.05 was considered statistically significant.
Results
Spindle Activity
Sleep-deprived mice (SD) exhibited a significant reduction in frontal spindle count compared to naturally sleeping controls (NS), consistent with disrupted thalamocortical activity (Table 1). To provide additional context, Supplementary Figure S2 presents representative full-length EEG recordings from NS and SD mice during NREM sleep. SD mice averaged 61 ± 14 spindles over the 12-hour recording period, while NS mice averaged 103 ± 12 spindles. We found this difference to be statistically significant (t(9.05) = –2.63, p = 0.027), with a 95% confidence interval ranging from –782.93 to –58.82. In contrast, the total duration of spindles did not differ significantly between groups (t(12.93) = –0.63, p = 0.541), suggesting that total spindle presence over time may remain relatively stable despite fewer events. However, the mean duration per spindle was significantly increased in the SD group (t(12.21) = 2.43, p = 0.032, 95% CI: 0.006 to 0.109), indicating that individual spindles, though fewer in number, tended to last longer during sleep deprivation. Quantitative comparisons are presented in Figures 1–3 with statistical annotations: Figure 1 confirms significantly prolonged spindle duration in SD mice (p=0.032); Figure 2 demonstrates reduced spindle count in SD mice (p=0.027); Figure 3 shows no significant change in total spindle duration (p=0.541). Overall, the changes in spindle characteristics imply that prolonged wakefulness alters both the occurrence and temporal structure of sleep spindles, which may impair processes like synaptic plasticity and memory consolidation that depend on precise spindle dynamics.
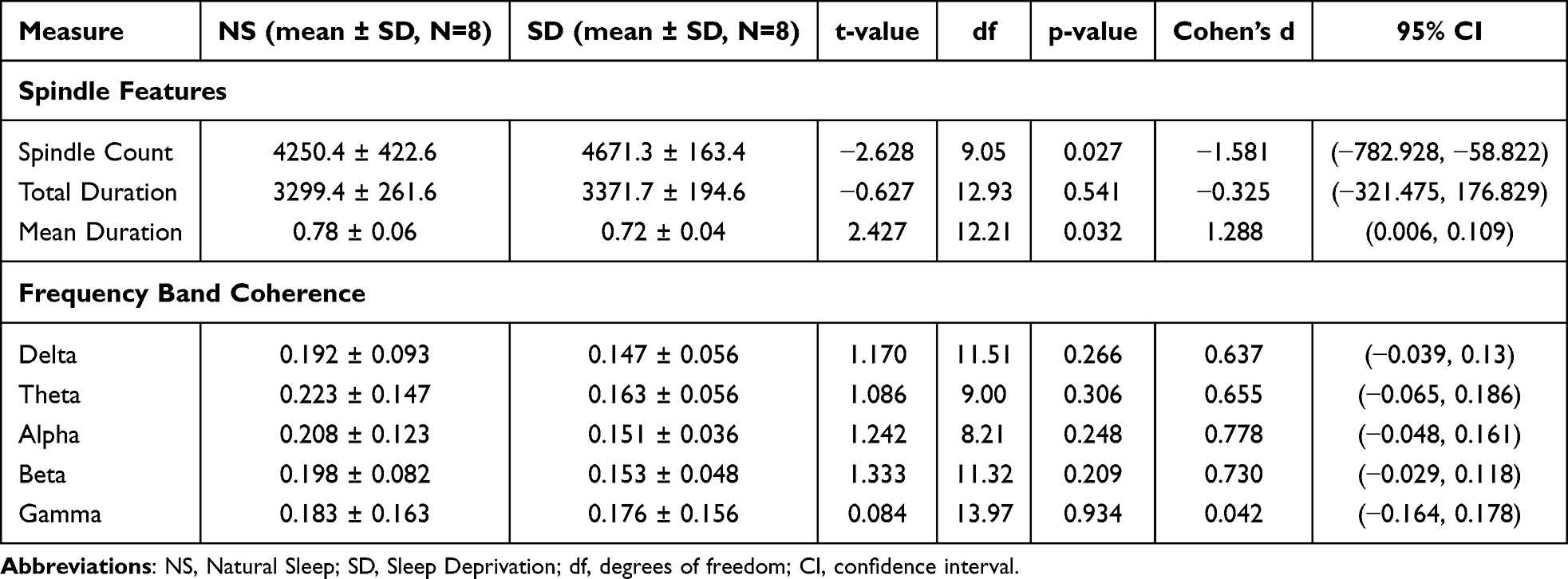 |
Table 1 Summary of Quantitative Results for Both Spindle Features and Frequency Band Coherence |
 |
Figure 1 Mean duration of sleep spindles in Natural Sleep (NS) and Sleep Deprivation (SD) groups. SD group showed significantly longer spindle duration (t(12.21)=2.43, p=0.032). Data presented as mean ± SEM. |
 |
Figure 2 Spindle count in Natural Sleep (NS) and Sleep Deprivation (SD) groups. SD group showed significantly fewer spindles (t(9.05)=−2.63, p=0.027). Data presented as mean ± SEM. |
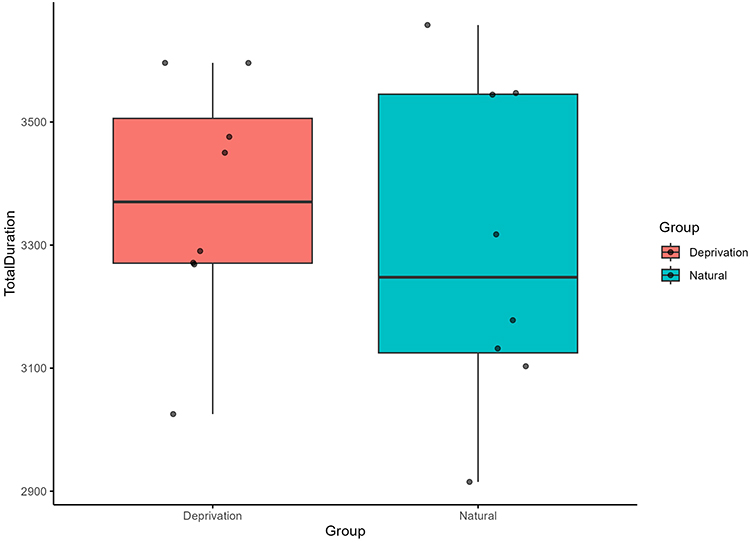 |
Figure 3 Total duration of sleep spindles in Natural Sleep (NS) and Sleep Deprivation (SD) groups. No significant difference was found between groups (t(12.93)=−0.63, p=0.541). Data presented as mean ± SEM. |
Interhemispheric Coherence
Analysis of interhemispheric EEG coherence revealed non-significant trends toward reduced synchronization following sleep deprivation across several frequency bands. In the delta band (0.5–4 Hz), the mean difference between NS and SD mice was [−0.039, 0.13] (t(11.51) = 1.17, p = 0.266). Similarly, in the theta band (4–8 Hz), the mean difference was [−0.065, 0.186] (t(9.00) = 1.09, p = 0.306). Trends in the alpha (8–13 Hz; mean diff [−0.048, 0.161], t(8.21) = 1.24, p = 0.248), beta (13–30 Hz; mean diff [−0.029, 0.118], t(11.32) = 1.33, p = 0.209), and gamma (30–40 Hz; mean diff [−0.164, 0.178], t(13.97) = 0.08, p = 0.934) bands also did not reach statistical significance. Figure 4 displays the group-averaged coherence spectra (0.5–40 Hz), showing coherence values across all analyzed frequency bands. While visual inspection of the figures might suggest trends, particularly in the lower frequencies, no band-specific changes reached the conventional threshold for statistical significance (p < 0.05). Consequently, the physiological interpretations regarding a marked disruption of slow-wave cortical synchrony, breakdown of large-scale synchrony supporting synaptic downscaling and memory, and widespread impacts on network dynamics lack statistical support in this dataset.
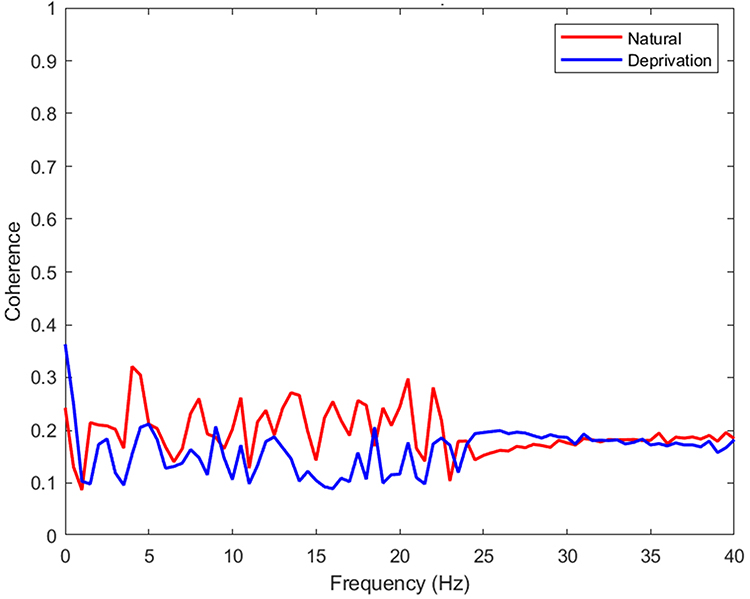 |
Figure 4 Coherence spectra (0.5–40 Hz) for NS vs SD groups. While slight differences in delta/theta coherence are visually apparent, no significant differences were detected in any frequency band (all p > 0.05). |
Discussion
Our findings demonstrate that even a single extended wake period profoundly disrupts coordinated NREM oscillations. Acute 12‑hour sleep deprivation markedly suppressed frontal sleep spindles and was associated with non-significant reductions in δ/θ coherence trends. In intact animals these oscillations normally rebound during recovery sleep, reflecting homeostatic potentiation of thalamocortical synchrony.23 The present results – a failure to generate spindles or maintain coherence under sleep pressure – suggest an acute breakdown of the circuits while spindle generation was clearly impaired, any disruption of low-frequency synchrony is only a possible effect given the lack of statistical significance. Sleep spindles in particular arise from the thalamic reticular nucleus (TRN) in concert with specific thalamic and cortical inputs.24 Our data imply that sleep loss effectively disengages this TRN pacemaker: fewer TRN burst firings lead to diminished waxing–waning spindle trains.
Disruption of Thalamocortical Synchrony
Mechanistically, these effects point to dysregulation of GABAergic and cholinergic pathways in the thalamocortical system. Spindle generation depends critically on the thalamic reticular nucleus (TRN) and its GABAergic interactions with relay cells.25,26 Acute sleep deprivation may reduce TRN excitability, thereby suppressing spindle activity.27 While additional mechanisms such as cholinergic or interneuronal modulation could contribute, detailed circuit-level speculation remains to be tested in future studies.28 In our view, sleep deprivation may suppress cholinergic tone (and enhance adenosine accumulation) so that TRN bursts become less frequent. Reduced TRN output would not only eliminate spindle bursts but also disinhibit thalamocortical relay cells. Indeed, TRN normally gates ascending rhythms – during wake it fires at high gamma to control sensory throughput,29 while during sleep its rhythmic bursts produce spindles. When TRN firing is compromised (for example by elevated corticosterone or reduced cholinergic drive during stress), the resulting disinhibition boosts slow oscillations and fragments cortical synchrony. Even gentle handling can elevate corticosterone levels and thus some of the EEG changes observed (particularly any reductions in low-frequency coherence or alterations in spindle activity) might partly reflect a stress response rather than purely the effect of sleep loss. Consistent with this, pharmacogenetic suppression of TRN activity induces cortical delta intrusions and impairs spindles,3,30 paralleling the observed low-frequency coherence trends we observed. Thus, our results suggest that acute sleep loss destabilizes the thalamocortical inhibitory network (via GABA/PV neurons in TRN) and its cholinergic modulation, producing spindle deficits and coherence trends.30 With n=8 per group, our study had >80% power to detect large effects (spindles) but insufficient power for the small coherence effects observed. Detecting these would require ~20–25 animals per group.
Implications for Cognitive Function
These data align with and extend previous sleep‑deprivation models in animals and humans. In humans, numerous studies report that total sleep deprivation acutely impairs attention, working memory, and other cognitive functions.31–33 Similarly, chronic partial restriction (for example 14 nights of 4–6 h sleep) produces cumulative deficits in vigilance and processing speed comparable to multi‑night total deprivation.34,35 Rodent studies show analogous effects: even single night deprivation can block hippocampal memory consolidation, while long-term sleep restriction induces spatial learning impairments and mood disturbances in mice. For instance, a recent meta-analysis found that repeated sleep loss robustly worsens depression-like behaviors in rodents, with effect sizes depending on strain and protocol.35 Notably, the present acute model differs from chronic paradigms – our single‑night deprivation contrasts with chronic insomnia or sleep fragmentation protocols. Chronic models may elicit additional adaptations (neuroinflammation, altered network excitability) that could modify EEG markers. For example, prolonged sleep restriction eventually leads to some degree of slow-wave rebound and changes in spindle homeostasis that are absent in acute deprivation. Together, these comparisons suggest that while acute and chronic sleep loss share some outcomes (eg attention lapses, mood dysregulation), the underlying EEG signatures may differ in time course and topology. Further work should directly contrast acute versus chronic sleep loss in rodents, as has been done in human studies, to delineate how spindle and coherence markers evolve over time.
The functional implications of these oscillatory disruptions are profound. Sleep spindles and δ/θ coherence have been strongly linked to memory, attention, and emotional processing. The reduction in spindle activity we observed would be expected to impair sleep-dependent memory consolidation, as higher spindle density is linked to better memory performance.36 Likewise, the trend toward lower delta/theta coherence could reflect a disruption in large-scale neural communication, potentially underlying lapses in attention or altered emotional regulation reported after sleep loss. Indeed, patients with schizophrenia (a condition characterized by cognition deficits) show marked spindle loss, and those spindle abnormalities correlate with poorer overnight memory and learning.37 In healthy subjects, higher spindle activity predicts better memory performance and general intelligence.6 Likewise, slow-wave coherence reflects large-scale network coordination: diminished frontal δ/θ coherence after sleep deprivation could underlie lapses in attention and sensory gating. Human EEG studies show that sleep loss disproportionately impairs vigilance and executive tasks.38 In our mice, the trend-level reduction in coherence may analogously disrupt thalamocortical gating of environmental stimuli, contributing to attentional deficits reported after deprivation. Sleep loss also disrupts emotional regulation, likely through limbic‑cortical circuits. For example, prefrontal inhibition of amygdala is weakened after one night without sleep, leading to hyper-reactive emotional responses (the well-known “sleep‑deprived amygdala” phenomenon in humans). Although our study did not measure behavior directly, the spindle deficits and coherence trends we report are consistent with a neurobehavioral profile of poorer memory, reduced cognitive control, and mood instability. These implications remain speculative but are biologically plausible based on prior human and animal studies. Supporting this, coherence biomarkers have already been proposed as indicators of neuropsychiatric risk. For instance, sleep EEG coherence patterns strongly distinguish PTSD patients from controls, correlating with symptom severity.39 Thus, the electrophysiological changes observed here likely have broad impacts on memory consolidation, attentional stability, and emotional balance in sleep‑deprived animals and by extension humans. We did not include behavioral testing. Therefore, any links between spindle/coherence changes and cognitive or mood outcomes remain speculative.
Beyond basic insights, our findings highlight spindle density and slow‑wave coherence as candidate biomarkers for cognitive function. Spindle activity in particular has emerged as a robust, noninvasive indicator of brain health and plasticity.4 Deviations from normative spindle metrics have been associated with neurodevelopmental disorders, aging, and neurodegeneration.7 In tandem, altered slow-wave coherence marks connectome disintegration seen in dementia and psychopathology.40 Our rodent data suggest that acute sleep loss drives these EEG features into patterns reminiscent of clinical conditions, raising the possibility that spindle/coherence indices could serve as early warning signs of sleep deprivation’s impact on the brain. Translationally, this supports the use of rodent sleep EEG as a bridge to human sleep research: by linking spindle/coherence changes to cognitive deficits in mice, we can better interpret EEG signals in sleep‑deprived people or patient populations. For example, closed-loop auditory stimulation during NREM (shown in humans to boost spindles and memory) might be adapted for rodent models to test causality and treatments. Furthermore, pharmacological interventions targeting GABA or cholinergic systems (eg zolpidem, muscarinic agonists) could be evaluated for their ability to restore EEG synchrony and cognitive performance in sleep‑deprived animals. Thus, the oscillatory markers we have identified may guide the development of interventions – from sensory stimulation to drugs – aimed at mitigating the cognitive and emotional fallout of sleep loss.
Several limitations of this study warrant discussion. We used only male C57BL/6J mice and a single acute deprivation paradigm, so generalizability is limited. Sleep loss affects females differently – for instance, hormonal cycles modulate sleep homeostasis – and different strains exhibit distinct spindle and coherence profiles (as suggested by our own strain comparisons in rebound sleep). Chronic or repeated sleep restriction might also engage compensatory mechanisms not captured here. In addition, although gentle handling is a widely used method for sleep deprivation, it can elevate stress hormones with repeated use.41 Elevated corticosterone itself alters EEG rhythms, so some of our effects may reflect stress rather than sleep loss per se even gentle handling can elevate corticosterone, potentially confounding EEG results. Our recordings were confined to frontal electrodes, providing only a partial view of brain networks. Multi‑site EEG (including parietal cortex and hippocampus) or high-density mapping would yield a fuller picture of how circuits are disrupted. Finally, mechanistic hypotheses drawn here (eg involvement of TRN GABA vs cholinergic inputs) remain speculative. Definitive conclusions will require targeted interventions – for example, optogenetic or chemogenetic manipulation of TRN/PV neurons or administration of receptor agonists/antagonists – to test causality. In conclusion, our work shows that acute sleep deprivation desynchronizes fundamental NREM rhythms by perturbing thalamocortical circuits. The resulting decreases in spindle density and cortical coherence likely undermine the brain’s ability to consolidate memory and regulate cognition and mood. By integrating neurophysiological and behavioral perspectives, these results deepen our understanding of sleep’s restorative functions. They also underscore the translational potential of rodent EEG: by identifying robust EEG biomarkers in mice, we can better interpret human sleep studies, improve diagnostics of sleep-related disorders, and develop interventions (pharmacological or sensory) to protect the brain from the cognitive costs of sleep loss.
Limitation
Despite demonstrating significant effects of acute sleep deprivation on sleep spindle activity with non-significant low-frequency coherence trends, this study has several limitations that warrant discussion. First, only male C57BL/6J mice were used, leaving the sex-specific generalizability of the findings unaddressed. Prior studies have shown that female mice exhibit distinct sleep homeostasis responses, partially influenced by hormonal cycles. With n=8 per group, our study had >80% power to detect large differences (d ~1.5) in spindle metrics, but much lower power (≈30–50%) to detect the small coherence effect sizes (d ~0.5) observed. Thus, a lack of statistical significance in coherence may stem from limited power. Second, our study employed a single sleep deprivation duration (12 hours), which limits understanding of how varying durations (eg, 6 hours, 24 hours) affect spindle dynamics and slow-wave coherence. Previous research has indicated that different lengths of deprivation can evoke varying degrees of slow-wave rebound and spindle recovery. Chronic sleep restriction models, in particular, may reveal compensatory adaptations that modulate EEG signatures. Additionally, we used gentle handling as the method for sleep deprivation. Although it is considered less stressful than water platforms or forced locomotion, evidence suggests that repeated gentle handling can still trigger stress responses, elevating corticosterone levels. Elevated corticosterone is known to alter EEG rhythms, particularly slow-wave power and spindle generation. Finally, EEG recordings were limited to the frontal cortex, providing only a partial view of neural network disruptions. Existing literature suggests that parietal and hippocampal slow-wave synchrony plays a crucial role in memory consolidation and emotional regulation. Although gentle handling is considered a mild method for sleep deprivation, repeated handling can still trigger stress responses (elevated corticosterone) that alter EEG rhythms. Thus, some changes we observed may be due to stress rather than sleep loss per se.
Future Direction
Future studies should address several key limitations and extend the present findings. Stress controls and direct stress measurements (eg, corticosterone assays or behavioral indicators) are needed to disentangle the effects of sleep loss from stress-induced neural changes. Including both sexes will help determine whether spindle and coherence alterations after sleep deprivation exhibit sex-specific differences. Mechanistic hypotheses should be tested directly by manipulating TRN or cholinergic pathways (eg, optogenetic activation, receptor-specific interventions) to clarify their causal role in spindle suppression and coherence alterations. Different deprivation paradigms, including chronic restriction and fragmented sleep, should be examined to capture long-term impacts on thalamocortical synchrony. Moreover, multi-site or high-density EEG (HD-EEG) will provide a more comprehensive mapping of oscillatory disruptions across cortical and subcortical regions.
In addition, future research should refine EEG-based assessments and explore potential interventions. Closed-loop auditory stimulation during NREM sleep, which enhances spindle synchrony, could be adapted to rodent models to test its ability to restore thalamocortical integrity after sleep deprivation. Pharmacological approaches targeting GABAergic or cholinergic systems (eg, GABA agonists or cholinergic modulators) may also be evaluated for their capacity to normalize synchrony and improve cognition. Finally, cross-species studies are warranted to validate these neurophysiological markers across various sleep loss models, facilitating translational bridges from animal models to human clinical applications.
Conclusion
In summary, our study demonstrates that acute sleep deprivation in mice significantly suppresses spindle generation, whereas coherence changes were limited to non-significant trends. These findings emphasize the sensitivity of thalamocortical oscillations to acute sleep loss and highlight the translational potential of spindles as biomarkers of sleep pressure. At the same time, the lack of significant coherence effects and absence of behavioral measures warrant cautious interpretation and underscore the need for further research. Future research should employ multi-site EEG recordings (eg, including parietal or hippocampal leads) to map the broader network disruptions caused by sleep loss, and incorporate behavioral assays to directly link EEG changes with cognitive outcomes. Additionally, interventions like closed-loop stimulation to boost spindles or pharmacological modulation of thalamocortical circuits could be tested for their ability to restore synchrony and function after sleep deprivation.
Data Sharing Statement
Data used in this study can be provided from the corresponding author upon reasonable request.
Ethics Approval and Consent to Participate
All animal procedures were approved by the Animal Ethics Committee of Nanfang Hospital of Southern Medical University. All experimental procedures were performed according to the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publications No. 8023, revised 1978) and the ARRIVE guidelines on the Care and Use of Experimental Animals.
Author Contributions
Y.X. conceptualization, methodology, writing-original draft, writing-review and editing, investigation, formal analysis; M.T. writing-review and editing, methodology, formal analysis; D. L. methodology, formal analysis, writing-review and editing; C.Z. conceptualization, investigation, methodology, writing-review and editing. All authors drafted or written, or substantially revised or critically reviewed the article; agreed on the journal to which the article will be submitted; reviewed and agreed on all versions of the article before submission, during revision, the final version accepted for publication, and any significant changes introduced at the proofing stage; agreed to take responsibility and be accountable for the contents of the article.
Funding
There is no funding to report.
Disclosure
The authors declare that they have no competing interests.
References
1. Fernandez LM, Lüthi A. Sleep spindles: mechanisms and functions. Physiol Rev. 2020;100(2):805–868. doi:10.1152/physrev.00042.2018
2. Lüthi A. Sleep spindles: where they come from, what they do. Neuroscientist. 2014;20(3):243–256. doi:10.1177/1073858413500854
3. Herrera CG, Tarokh L. A thalamocortical perspective on sleep spindle alterations in neurodevelopmental disorders. Curr Sleep Med Reports. 2024;10:103–118.
4. Urakami Y, Ioannides AA, Kostopoulos GK. Chapter sleep spindles–as a biomarker of brain function and plasticity. Clin EEG Neurosci. 2012;43(1):39–47. doi:10.1177/1550059411428718
5. Reynolds C, Short M, Gradisar M. Sleep spindles and cognitive performance across adolescence: a meta-analytic review. J Adolescence. 2018;66:55–70. doi:10.1016/j.adolescence.2018.04.003
6. Ujma PP, States CU. Sleep spindles and general cognitive ability–A meta-analysis. Sleep Spindles Cortical up States. 2021;2:1–17.
7. Peyrache A, Seibt J. A mechanism for learning with sleep spindles. Philosophical Transact Royal Soc London. Series B, Biol Sci. 2020;375(1799):20190230. doi:10.1098/rstb.2019.0230
8. Vyazovskiy V, Achermann P, Borbély AA, Tobler I. Interhemispheric coherence of the sleep electroencephalogram in mice with congenital callosal dysgenesis. Neuroscience. 2004;124(2):481–488. doi:10.1016/j.neuroscience.2003.12.018
9. Hahn M, Joechner A-K, Roell J, et al. Developmental changes of sleep spindles and their impact on sleep‐dependent memory consolidation and general cognitive abilities: a longitudinal approach. Develop Sci. 2019;22(1):e12706. doi:10.1111/desc.12706
10. Chatburn A, Coussens S, Lushington K, et al. Sleep spindle activity and cognitive performance in healthy children. Sleep. 2013;36(2):237–243. doi:10.5665/sleep.2380
11. Mak-McCully RA, Rolland M, Sargsyan A, et al. Coordination of cortical and thalamic activity during non-REM sleep in humans. Nat Commun. 2017;8:15499. doi:10.1038/ncomms15499
12. Tsai Y-T, Chan H-L, Lee S-T, et al. Significant thalamocortical coherence of sleep spindle, theta, delta, and slow oscillations in NREM sleep: recordings from the human thalamus. Neurosci Letters. 2010;485(3):173–177. doi:10.1016/j.neulet.2010.09.004
13. Purcell S, Manoach DS, Demanuele C, et al. Characterizing sleep spindles in 11,630 individuals from the national sleep research resource. Nat Commun. 2017;8:15930. doi:10.1038/ncomms15930
14. Sakellariou D, Koupparis AM, Kokkinos V, Koutroumanidis M, Kostopoulos GK. Connectivity measures in EEG microstructural sleep elements. Front Neuroinformatics. 2016;10:5. doi:10.3389/fninf.2016.00005
15. Achermann P, Borbély AA. Coherence analysis of the human sleep electroencephalogram. Neuroscience. 1998;85(4):1195–1208. doi:10.1016/S0306-4522(97)00692-1
16. Chow R, Rabi R, Paracha S, et al. Default mode network and neural phase synchronization in healthy aging: a resting state EEG study. Neuroscience. 2022;485:116–128. doi:10.1016/j.neuroscience.2022.01.008
17. Magyar TZ, Szalárdy O, Bódizs R. Sleep deprivation in mice: looking beyond the slow wave rebound. 2024;
18. Harel A, Levkovsky A, Nakdimon I, Gordon B, Shriki O. EEG-based prediction of reaction time during sleep deprivation. Sleep. 2025;48(7):zsaf080. doi:10.1093/sleep/zsaf080
19. Goh CWC. Nocturnal Oscillations: Understanding the Brain Through Sleep. UCL (University College London); 2014.
20. Lindemann C, Ahlbeck J, Bitzenhofer SH, Hanganu-Opatz IL. Spindle activity orchestrates plasticity during development and sleep. Neural Plasticity. 2016;2016:5787423. doi:10.1155/2016/5787423
21. Siebenhühner F, Lobier M, Wang SH, Palva S, Palva JM. Measuring large-scale synchronization with human MEG and EEG: challenges and solutions. In: Multimodal Oscillation-Based Connectivity Theory. Springer International Publishing; 2016;1–18.
22. Schwartz S, Kessler R, Gaughan T, Buckley AW. Electroencephalogram coherence patterns in autism: an updated review. Pediatric Neurol. 2017;67:7–22. doi:10.1016/j.pediatrneurol.2016.10.018
23. Halassa MM. Seminars in Cell & Developmental Biology. Elsevier;245–251.
24. Vantomme G, Osorio-Forero A, Lüthi A, Fernandez LM. Regulation of local sleep by the thalamic reticular nucleus. Front Neurosci. 2019;13:576. doi:10.3389/fnins.2019.00576
25. Lee J, Song K, Lee K, et al. Sleep spindles are generated in the absence of T-type calcium channel-mediated low-threshold burst firing of thalamocortical neurons. Proc Nat Acad Sci USA. 2013;110(50):20266–20271. doi:10.1073/pnas.1320572110
26. Cheong E, Shin H-S. T-type Ca2+ channels in normal and abnormal brain functions. Physiolog Rev. 2013;93(3):961–992. doi:10.1152/physrev.00010.2012
27. Ozen Irmak S, de Lecea L. Basal forebrain cholinergic modulation of sleep transitions. Sleep. 2014;37(12):1941–1951. doi:10.5665/sleep.4246
28. Granger AJ, Mulder N, Saunders A, Sabatini BL. Cotransmission of acetylcholine and GABA. Neuropharmacology. 2015;100:40–46. doi:10.1016/j.neuropharm.2015.07.031
29. Coenen AJ. Sensory gating and gaining in sleep: the balance between the protection of sleep and the safeness of life (a review). J Sleep Res. 2024;33(5):e14152. doi:10.1111/jsr.14152
30. Thankachan S, Katsuki F, McKenna JT, et al. Thalamic reticular nucleus parvalbumin neurons regulate sleep spindles and electrophysiological aspects of schizophrenia in mice. Scientific Reports. 2019;9(1):3607. doi:10.1038/s41598-019-40398-9
31. García A, Del Angel J, Borrani J, Ramirez C, Valdez P. Sleep deprivation effects on basic cognitive processes: which components of attention, working memory, and executive functions are more susceptible to the lack of sleep? Sleep Sci. 2021;14(2):107. doi:10.5935/1984-0063.20200049
32. Killgore WD. Effects of sleep deprivation on cognition. Progress Brain Res. 2010;185:105–129.
33. Frenda SJ, Fenn KM. Sleep less, think worse: the effect of sleep deprivation on working memory. J Applied Res Memory Cognition. 2016;5:463–469.
34. Reynolds AC, Banks S. Total sleep deprivation, chronic sleep restriction and sleep disruption. Progress Brain Res. 2010;185:91–103. doi:10.1016/B978-0-444-53702-7.00006-3
35. Wüst LN, Capdevila NC, Lane LT, Reichert CF, Lasauskaite RJ. Impact of one night of sleep restriction on sleepiness and cognitive function: a systematic review and meta-analysis. Sleep Med Rev. 2024;76:101940.
36. McDevitt EA, Krishnan GP, Bazhenov M, Mednick SC. The role of sleep spindles in sleep-dependent memory consolidation. In: Cognitive Neuroscience of Memory Consolidation. Springer International Publishing; 2017:209–226.
37. Manoach DS, Stickgold R. Abnormal sleep spindles, memory consolidation, and schizophrenia. Ann Rev Clin Psychol. 2019;15:451–479.
38. Hudson AN, Van Dongen HP, Honn KA. Sleep deprivation, vigilant attention, and brain function: a review. Neuropsychopharmacology. 2020;45:21–30.
39. Modarres MH, Opel RA, Weymann KB, Lim MM. Strong correlation of novel sleep electroencephalography coherence markers with diagnosis and severity of posttraumatic stress disorder. Scientific Reports. 2019;9(1):4247. doi:10.1038/s41598-018-38102-4
40. Meijer DK. Concept of integral holographic consciousness: relation with predictive coding, phi-based harmonic EEG coherence as perturbed in mental disorders.
41. McEwen BS. Sleep deprivation as a neurobiologic and physiologic stressor: allostasis and allostatic load. Metabolism. 2006;55:S20–S23.
 © 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2025 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 4.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.