Back to Journals » ImmunoTargets and Therapy » Volume 11
β-Glucan “Trained Immunity” Immunomodulatory Properties Potentiate Tissue Wound Management and Accelerate Fitness Recover
Authors Renke G ![]() , Baesso T, Paes R, Renke A
, Baesso T, Paes R, Renke A
Received 12 July 2022
Accepted for publication 4 October 2022
Published 17 October 2022 Volume 2022:11 Pages 67—73
DOI https://doi.org/10.2147/ITT.S381145
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Michael Shurin
Guilherme Renke,1,2 Thomaz Baesso,2 Raphaela Paes,2 Aline Renke2
1National Institute of Cardiology, Brazilian Ministry of Health, Rio de Janeiro, Brazil; 2Nutrindo Ideais Performance and Nutrition Research Center, Rio de Janeiro, Brazil
Correspondence: Guilherme Renke, Endocrinology, Nutrindo Ideais Performance and Nutrition Research Center, Rua Alm. Saddock de Sá 207. Ipanema, Rio de Janeiro, Brazil, Tel +55522997840836, Email [email protected]
Introduction: It is well established that moderate physical activity can improve the immune status, rather excess or high-intensity physical exercise can cause damage to the immune system. In addition, muscle injuries resulting from increased frequency and intensity of exercises compromise innate immune activity and may decrease tissue regeneration. Thus, β-glucans, a natural compound, may represent an important substance with strong immunomodulatory properties acting as an immunostimulant therapy known as “trained immunity”. This immune stimulating therapeutic is an immunological memory phenomenon linked to the innate immune system, triggering cellular changes at epigenetic, transcriptional, and functional levels, to regulate the immune system and recover its homeostasis with clinical benefits.
Conclusion: This narrative review works with the current evidence regarding β-glucans as a possible alternative therapy for wound healing and its safety and efficacy in the treatment of muscle injuries and physical recovery including other chronic conditions and diseases.
Keywords: innate immunity, metabolic-inflammation, trained immunity, beta-glucans, muscle recovery, wound healing, tissue regeneration
Introduction
Sedentary behavior may be an important determinant of health and inactive individuals are more likely to suffer from viral infections, an increase in lung inflammation, an imbalance in body weight, and abdominal fat, which relates to sarcopenia and reduces muscular function.1,2 On the other hand, individuals who practice physical activity develop better immune system responses, increasing T-cells responses, inhibiting lung inflammation, and mitigating the progress of viral infection.3,4 A sedentary lifestyle is associated with an increased incidence of infections when compared to regular and moderate exercise.1–4
However, several immunological aspects can be temporarily weakened, such as antigen presentation to monocytes and a neutrophilic respiratory response in the case of strenuous physical exercise.4,5 Excessive exercise for more than 1.5 h, of high or moderate intensity (O2 maximum uptake between 55–75%) and in a non-fed state can favor the onset of postexercise immune dysfunction. Although, we know that moderate activities have a positive immunological effect compared to sedentary individuals. Exercise can have both favorable and unfavorable effects on immune function and predisposition to minor illnesses.2–4
The practice of continuous and moderate physical exercise is associated with neuroendocrine and metabolic adaptations that lead to an improvement in the immune system.5 However, the practice of high-intensity physical exercises or high-volume training could lead to transient negative immunological adaptations. Negative changes could last from 3 to 72 hours, which may represent a major risk of infections.5,6
A systematic review looked at the effects of interval training on the immune system.7 It has been shown that after a single session of interval physical exercise, there is a slight reduction in immune function due to greater production of inflammatory cytokines, such as interleukin 6, and greater intestinal permeability leading to greater absorption of intestinal endotoxins.6–8
Rather, regular practice of training leads to positive adaptations in immune function, which improves the immune system in ways that do not require an increase in immune cell counts.7,8 Additionally, this benefit is observed in strength and resistance physical exercise where the effects of this type of training were beneficial for the cellular immune action leading to a decrease in inflammatory markers both in the elderly and in young individuals.8,9
Thus, exercise overload should bring better attention to immunosurveillance as it has been linked to the downregulation of the immune system.5,6,8 In this review we describe the advantages of an immune-stimulating therapeutic referred to as “trained immunity” using β-glucans (beta-glucans) to up-regulate the immune system and recover its homeostasis with clinical benefits, especially, in the recovery of muscle injuries and regeneration of tissue injuries.
Recently, studies have shown that β-glucans can induce trained immunity, through epigenetic changes in monocytes, promoting modifications that can protect against infections by different microorganisms.10 The long-term effects of trained immunity on circulating monocytes are still under investigation, as these cells have a relatively short lifespan in circulation.
We will also present up-to-date literature on this strategy on recurrent infections, fitness recovery, and general well-being. Also, discuss the key successful data on tissue injuries management that indicates promising potential therapeutic on tissue injuries derived from exercise injury and overtraining.9
β-Glucans
Natural substances that can treat pathological conditions have been isolated and identified, such as β-glucans. Chemically, they are heterogeneous non-starch polysaccharides that form cell walls and other items of microorganisms such as algae, mushrooms, and grains.11 β-glucans can be soluble or insoluble. Soluble β-glucan fibers consist of β-(1,3/1,6)-d-linked glucose, while insoluble β-glucan consists of β-(1,3/1,4)-d-linked glucose units.11,12
Several studies, initially published 45 years ago, undoubtedly show that β-glucans are effective substances that help the body in the important defense mechanisms of immunomodulation.11,12 This effect occurs through numerous mechanisms, including the stimulation of cellular and humoral immunity,12 control of metabolic diseases, such as diabetes,13,14 stimulation of regenerative systems, such as wound healing,15 attenuation of chronic fatigue and stress,16 cancer inhibitory stimuli, and lowering cholesterol (Table 1).17,18
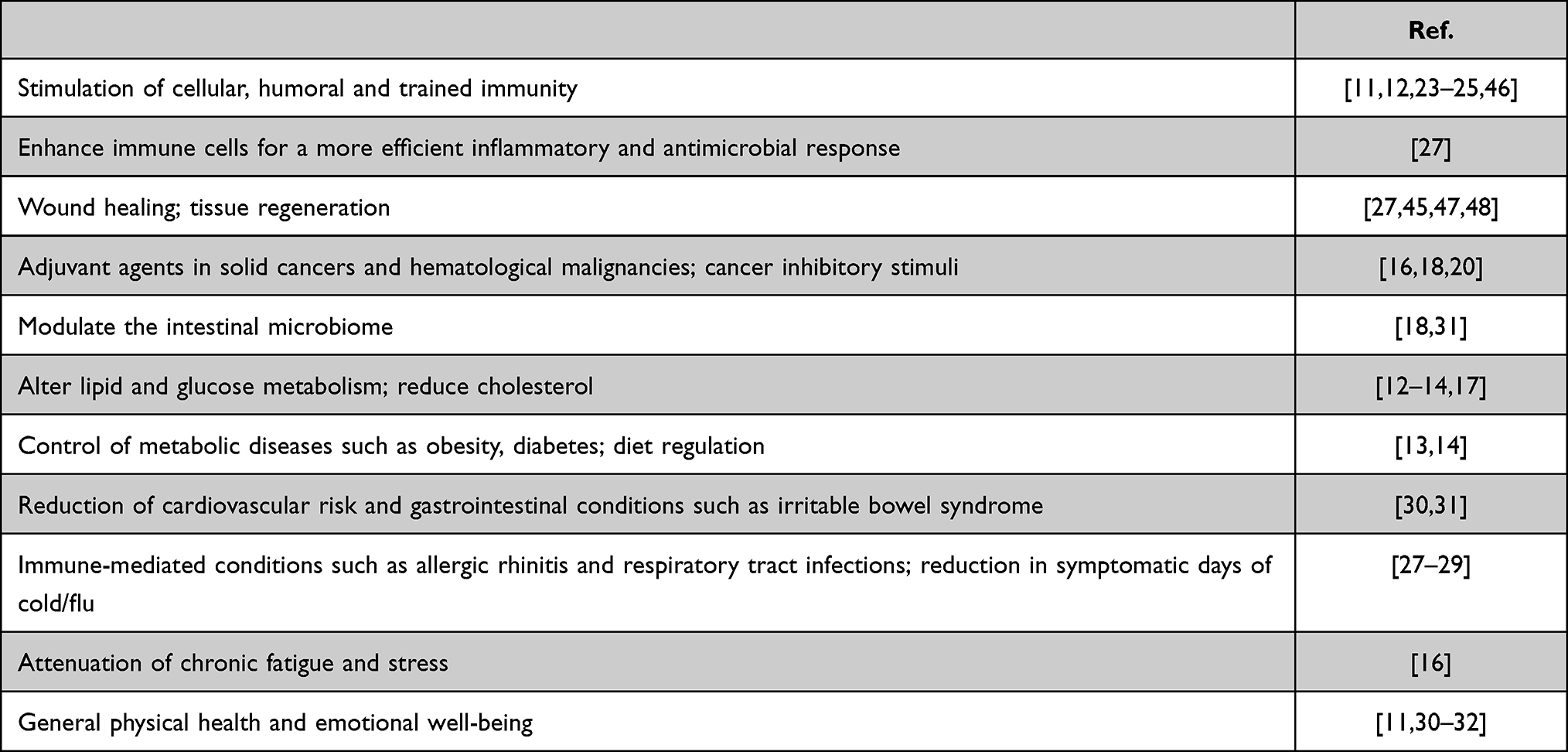 |
Table 1 Biomedical Applications of β-Glucans |
Oral supplementation with β-glucan is the most used and widely studied, being safe in doses of 35 to 500 mg a day for both children and adults.19 Vetvika et al examine several commercially available β-glucans and report that the injectable application of β-glucans, used as a single dose, showed greater effects than oral supplementation.20 In addition, authors observed that some β-glucans exhibited either longer effects or were effective only after subcutaneous injection.20,21
Regarding its effects on CD4-positive lymphocytes, a single application of β-glucans injection already increases the influx of these cells, a fact that led to their greater expression.20,21 Thus, CD8 cells were also approached and investigated and it was seen the same effect for a period of 48 hours. Additional data showed that the duration of these effects depends on the moment and intensity of the β-glucans treatment since doses when replicated, resulting in a more lasting and potent stimulus. Therefore, therapy with injected β-glucans has clinical and laboratory evidence compared to oral supplementation, to represent a possible current strategy in the development of immune modulation.21
Training Immunity and β-Glucans
Innate immunity is characterized by a rapid response to aggression independent of previous stimulus, non-specific, which does not generate immunological memory. Adaptive immunity is characterized by an antigen-specific immune response, which generates immunological memory. It is known that both act together, however, it was observed that some organisms do not have adaptive immunity, although they have immunological memory.10
“Trained immunity” is an immunological memory phenomenon linked to the innate immune system, given by the first encounter between a pathogen and classical innate immune cells, triggering cellular changes at epigenetic, transcriptional, and functional levels that accelerate future defense against the same pathogen or an unrelated pathogen (Figure 1).10,22 In the innate system, this recognition of pathogens can be done through pattern recognition receptors (PRRs) expressed in these cell types. Through this process, trained immunity can lead to greater protection, increasing the effectiveness of the non-specific response of innate immune cells against pathogens, especially monocytes and macrophages. Thus, taking advantage of the activation state of dendritic cells to increase T cell adaptive responses to specific and unrelated pathogens including the regulating of the gut microbiota.22,23 Several immunostimulants and adjuvants can already fall under the concept of trained immunity, including due to their ability to promote responses beyond their specific antigens.23
 |
Figure 1 Immune mechanism of trained immunity. Abbreviations: DC, dentritic cell; Treg, Regulatory T cells. |
Trained immunity concerns epigenetic changes resulting from the reprogramming of chromatin marks within the progenitors of innate immune cells,24,25 the mechanism by which β-glucan can enhance these cells, whereas there is evidence of a more robust innate response of monocytes and macrophages against various pathogens after exposure to this substance, with enhanced antimicrobial and inflammatory properties, derived from activation of the dectin-1/toll-like receptor (TLR).25 However, compared to the classical acquired immunity offered by antigen receptors as part of the adaptive immune response, PRRs show less specificity and duration of memory.26
β-glucans are highly conserved structural components constituting most yeast and fungal cell walls and are not produced by mammalian cells. They act as “biological response modifiers” and can improve the immune system of humans, as they are recognized as microbe-associated molecular patterns (MAMPs) by PRRs present on the surface of innate immune cells,26,27 being phagocytosed and processed by monocytes, macrophages and dendritic cells found in the lymphatic tissue of the upper intestine, to later be transported to different immune organs (such as the spleen), where soluble fragments of β-1,3-glucans are released and enhance immune cells for a more efficient inflammatory and antimicrobial response.27
Several clinical studies point to a reduction in the symptomatology of upper respiratory tract infections and symptomatic days of cold/flu after intense exercise, decreased fatigue, and increased vigor after its use.27,28 In addition, β-glucans have important metabolic and gastrointestinal effects, being able to modulate the intestinal microbiome, alter lipid and glucose metabolism, and reduce cholesterol.12,13,17 Thus, they have probable therapeutic potential for the treatment of metabolic syndrome, obesity, diabetes, diet regulation, reduction of cardiovascular risk, and gastrointestinal conditions such as irritable bowel syndrome.29,30 Due to their immunomodulatory effect, they can act as adjuvant agents in solid cancers and hematological malignancies, in immune-mediated conditions such as allergic rhinitis and respiratory tract infections, and also improve wound healing.31 At least 13 randomized controlled trials have reported improvement in allergy symptoms, general physical health, and emotional well-being with the use of β-glucans compared with placebo.30–32
β-Glucan in Tissue Repair
Topical applications of β-glucans are increasing, especially due to their pluripotent properties. The most promising area for the application of β-glucans is tissue repair, for example, in wound healing, where macrophages, keratinocytes, and fibroblasts are considered the main target cells, since they can improve wound repair, increasing macrophage infiltration, stimulating tissue granulation, collagen deposition and re-epithelialization.32,33 The wound healing process is a four-step process that includes hemostasis, inflammation, proliferation (or granulation), and remodeling (or maturation). Cells such as neutrophils, macrophages, and fibroblasts are involved in orchestrating the healing process and the inflammatory response causes these cells to secrete various cytokines that work together to induce cell migration and remodeling.34
Neutrophils and monocytes/macrophages are the main immune cells in skin wound repair, especially in the inflammatory phase, where clot formation occurs and neutrophils are recruited to the wound area to phagocytose invading pathogens and necrotic tissue.35 Monocytes enter the injury site a few days later and ingest cellular debris, including neutrophils and remaining pathogens.36 Although resident macrophages contribute to the inflammatory response to some extent, most macrophages found at the site of injury are derived from blood monocytes and recruited by the release of chemokines.37
Other immune cells with a role in tissue repair are mast cells, T cells, type 1 T helper cells (TH1), and type 2 T helper cells (TH2), the last two mentioned being mainly involved in the proliferation and remodeling phase.22,38 During the proliferation phase, re-epithelialization occurs, and the formation of new vessels (angiogenesis) and granulation tissue, responsible for covering and filling the affected area, restoring tissue integrity. Finally, the remodeling phase can last up to a year after injury and involves the reorganization of connective tissue and wound contraction with scar formation.38–41 In addition to contributing to the inflammatory response, macrophages also produce growth factors that stimulate the proliferation of non-immune cells (especially keratinocytes and fibroblasts) and extracellular matrix synthesis.41,42
Until recently, neutrophils were thought to have only pro-inflammatory functions, however, they are also capable of acquiring different phenotypes linked to their functions (pro-inflammatory, pro-healing, pro-fibrotic, anti-inflammatory, anti-fibrotic), generating successful phases in tissue repair.42 Neutrophils also release growth factors and promote angiogenesis, contributing to the resolution of inflammation.43,44
The healing of skin and skeletal muscle is influenced by local and systemic factors that can increase the likelihood of complications, causing impaired healing. The recurrence of physical trauma and tissue injuries caused by physical exercise and overtraining compromise the innate immunity and healing capacity of the patient7,9,10,35 while changes in coagulation, tissue perfusion, immune functioning, metabolism, nutrition, and drug therapies promote modification of biological responses to tissue rupture, influencing healing.35,36
Pathogen-associated molecular patterns can trigger these responses when recognized by PRRs, such as C-lectin receptors (CLRs) associated with dendritic cells (Dectin-1).43,44 Recent studies demonstrate that the activation of the Dectin-1 signal accelerated the wound healing response, inducing the accumulation of neutrophils in the initial phase.45 It is well documented that the immunomodulatory properties of β-Glucan can contribute to the upregulation of the inflammatory response and its effects on primary human dermal fibroblasts accelerate the wound healing process.46–48
Many advances have been made in the use and supplementation of β-glucan in humans either orally, transdermally, or by injection. Its use appears to be safe and efficient in a range of conditions that include tissue and muscle recovery after injury. Additionally, we can also suggest a post-exercise recovery benefit of β-glucan for athletes due to its modulating effect on the immune system. However, more studies are needed to assess the real impact of supplementation with β-glucan and improvement in sports performance.
Conclusion
Trained immunity can result in a better response of innate immune cells to infection, with effective and rapid elimination of pathogens, avoiding excessive inflammation and facilitating the succession to the proliferative phase of healing. In summary, this review presents current knowledge and progress made in characterizing the wound healing properties promoted by the use of β-glucans and their safety and effectiveness in managing non-healing injuries or other chronic conditions and diseases.
Tissue repair is the most promising area for the application of β-glucans because of the wound healing contribution by stimulating tissue regeneration, collagen biosynthesis, and deposition, increasing wound tensile strength and maintaining skin’s moisture and elasticity. β-glucans induce the production of growth factors essential for skin, re-epithelialization, and regeneration of damaged skin. In short, β-glucans benefits includes their ability to regulate the immune system, reduction of inflammatory expression in skeletal muscle, a decrease in oxidative stress and optimization of tissue regeneration, and faster healing.
Abbreviations
β-glucans, Betaglucans; CLRs, C-type lectin receptors; Dectin-1, Dendritic cell-associated C-type lectin; MAMPs, Microbial-associated molecular patterns; PRRs, pattern recognition receptors; TH1 cells, Type 1 T helper; TH2 cells, Type 2 T helper; TLR, dectin-1/toll-like receptor.
Data Sharing Statement
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis, and interpretation, or in all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors declare that they have no competing interests.
References
1. de Rezende LF, Rodrigues Lopes M, Rey-López JP, Matsudo VK, Luiz Odo C. Sedentary behavior and health outcomes: an overview of systematic reviews. PLoS One. 2014;9(8):e105620. PMID: 25144686; PMCID: PMC4140795. doi:10.1371/journal.pone.0105620
2. Prestes J, Ferreira CKO, Frollini AB, et al. Influence of acute physical exercise performed until exhaustion on the number of circulating leukocytes, lymphocytes, and cytokines. Fit Perf J. 2007;6(1):32–37.
3. Filgueira TO, Castoldi A, Santos LER, et al. The relevance of a physical active lifestyle and physical fitness on immune defense: mitigating disease burden, with focus on COVID-19 consequences. Front Immunol. 2021;12:587146. PMID: 33613573; PMCID: PMC7892446. doi:10.3389/fimmu.2021.587146
4. Gleeson M. Immune function in sport and exercise. J Appl Physiol. 2007;103(2):693–699.
5. Nieman DC, Wentz LM. The compelling link between physical activity and the body’s defense system. J Sport Health Sci. 2019;8(3):201–217. PMID: 31193280; PMCID: PMC6523821. doi:10.1016/j.jshs.2018.09.009
6. Gonçalves CAM, Dantas PMS, Dos Santos IK, et al. Effect of acute and chronic aerobic exercise on immunological markers: a systematic review. Front Physiol. 2020;10:1602. PMID: 32038286; PMCID: PMC6993577. doi:10.3389/fphys.2019.01602
7. Souza D, Vale AF, Silva A, et al. Acute and chronic effects of interval training on the immune system: a systematic review with meta-analysis. Biology. 2021;10(9):868. PMID: 34571745; PMCID: PMC8465842. doi:10.3390/biology10090868
8. Pires W, Veneroso CE, Wanner SP, et al. Association between exercise-induced hyperthermia and intestinal permeability: a systematic review. Sports Med. 2017;47(7):1389–1403. PMID: 27943148. doi:10.1007/s40279-016-0654-2
9. Proschinger S, Winker M, Joisten N, Bloch W, Palmowski J, Zimmer P. The effect of exercise on regulatory T cells: a systematic review of human and animal studies with future perspectives and methodological recommendations. Exerc Immunol Rev. 2021;27:142–166. PMID: 33965900.
10. Netea MG, Joosten LA, Latz E, et al. Trained immunity: a program of innate immune memory in health and disease. Science. 2016;352(6284):aaf1098. doi:10.1126/science.aaf1098
11. Barsanti L, Passarelli V, Evangelista V, Frassanito AM, Gualtieri P. chemistry, physics-chemistry and applications linked to biological activities of β-glucans. Nat Prod Rep. 2011;28:457–466. doi:10.1039/c0np00018c
12. Novak M, Vetvicka V. Beta-glucans, history, and the present: immunomodulatory aspects and mechanisms of action. J Immunotoxicol. 2008;5(1):47–57. doi:10.1080/15476910802019045
13. Wursch P, Pi-Sunyer FX. The role of viscous soluble fiber in the metabolic control of diabetes. A review with special emphasis on cereals rich in beta-glucan. Diabetes Care. 1997;20(11):1774–1780. doi:10.2337/diacare.20.11.1774
14. Bozbulut R, Sanlier N. Promising effects of β-glucans on glycemic control in diabetes. Trends Food Sci Technol. 2019;83:159–166. doi:10.1016/j.tifs.2018.11.018
15. Browder W, Williams D, Lucore P, Pretus H, Jones E, McNamee R. Effect of enhanced macrophage function on early wound healing. Surgery. 1988;104(2):224–230.
16. Vetvicka V, Vetvickova J. B-glucan attenuates chronic fatigue syndrome in a murine model. J Nat Sci. 2015;1:e112.
17. Braaten JT, Wood PJ, Scott FW, et al. Oat beta-glucan reduces blood cholesterol concentration in hypercholesterolemic subjects. Eur J Clin Nutr. 1994;48(7):465–474.
18. Sima P, Vannucci L, Vetvicka V. Glucan and cancer: historical perspective. Canc Transl Med. 2015;1(6):209–214. doi:10.4103/2395-3977.172860
19. De Marco Castro E, Calder PC, Roche HM. β-1,3/1,6-glucans and immunity: state of the art and future directions. Mol Nutr Food Res. 2021;65(1):e1901071. PMID: 32223047; PMCID: PMC7816268. doi:10.1002/mnfr.201901071
20. Vetvicka V, Vetvickova J. Glucans and cancer: comparison of commercially available β-glucans - part IV. Anticancer Res. 2018;38(3):1327–1333. doi:10.21873/anticanres.12355
21. Vetvicka V, Vetvickova J. A comparison of injected and orally administered β-glucans, Vaclav vetvicka, jana vetvickova, ms university of Louisville, department of pathology, Louisville, Kentucky. J Am Nutraceut Ass. 2008;11(1):42–49.
22. Mengdai X, Xiaoxing M, Huang H, et al. Yeast β-glucan alleviates cognitive deficit by regulating gut microbiota and metabolites in Aβ1–42-induced AD-like mice. Int J Biol Macromol. 2020;161:258–270. doi:10.1016/j.ijbiomac.2020.05.180
23. Sánchez-Ramón S, Conejero L, Netea MG, Sancho D, Palomares Ó, Subiza JL. Trained immunity-based vaccines: a new paradigm for the development of broad-spectrum anti-infectious formulations. Front Immunol. 2018;17(9):2936. doi:10.3389/fimmu.2018.02936
24. Netea MG, Quintin J, van der Meer JM. Trained immunity: a memory for innate host defense. Cell Host Microbe. 2011;9(5):355–361. doi:10.1016/j.chom.2011.04.006
25. Quintin J, Saeed S, Martens JHA, et al. Candida albicans infection affords protection against reinfection via functional reprogramming of monocytes. Cell Host Microbe. 2012;12(2):223–232. doi:10.1016/j.chom.2012.06.006
26. McCann F, Carmona E, Puri V, Pagano RE, Limper AH. Macrophage internalization of fungal beta-glucans is not necessary for the initiation of related inflammatory responses. Infect Immun. 2005;73(10):6340–6349. PMID: 16177305; PMCID: PMC1230895. doi:10.1128/IAI.73.10.6340-6349.2005
27. Talbott S, Talbott J. Effect of BETA 1, 3/1, 6 GLUCAN on upper respiratory tract infection symptoms and mood state in marathon athletes. J Sports Sci Med. 2009;8(4):509–515.
28. McFarlin BK, Carpenter KC, Davidson T, McFarlin MA. Baker’s yeast beta glucan supplementation increases salivary IgA and decreases cold/flu symptomatic days after intense exercise. J Diet Suppl. 2013;10(3):171–183. doi:10.3109/19390211.2013.820248
29. Zhong K, Liu Z, Lu Y, Xu X. Effects of yeast β-glucans for the prevention and treatment of upper respiratory tract infection in healthy subjects: a systematic review and meta-analysis. Eur J Nutr. 2021;60(8):4175–4187. PMID: 3390046. doi:10.1007/s00394-021-02566-4
30. Wang Q, Sheng X, Shi A, et al. β-glucans: relationships between modification, conformation and functional activities. Molecules. 2017;22:257. doi:10.3390/molecules22020257
31. Murphy EJ, Rezoagli E, Major I, Rowan NJ, Laffey JG. β-glucan metabolic and immunomodulatory properties and potential for clinical application. J Fungi. 2020;6(4):356. PMID: 33322069; PMCID: PMC7770584. doi:10.3390/jof6040356
32. Majtan J, Jesenak M. β-glucans: multi-functional modulator of wound healing. Molecules. 2018;23(4):806. PMID: 29614757; PMCID: PMC6017669. doi:10.3390/molecules23040806
33. Pasparakis M, Haase I, Nestle FO. Mechanisms regulating skin immunity and inflammation. Nat Rev Immunol. 2014;14(5):289–301. PMID: 24722477. doi:10.1038/nri3646
34. Guo S, Dipietro LA. Factors affecting wound healing. J Dent Res. 2010;89(3):219–229. PMID: 20139336; PMCID: PMC2903966. doi:10.1177/0022034509359125
35. Williams DT, Harding K. Healing responses of skin and muscle in critical illness. Crit Care Med. 2003;31(8Suppl):S547–57. PMID: 12907885. doi:10.1097/01.CCM.0000081430.34066.1D
36. Nosrati H, Khodaei M, Alizadeh Z, Banitalebi-Dehkordi M. Cationic, anionic and neutral polysaccharides for skin tissue engineering and wound healing applications. Int J Biol Macromol. 2021;192:298–322. doi:10.1016/j.ijbiomac.2021.10.013
37. Singer AJ, Clark RA. Cutaneous wound healing. N Engl J Med. 1999;341(10):738–746. PMID: 10471461. doi:10.1056/NEJM199909023411006
38. Wang J. Neutrophils in tissue injury and repair. Cell Tissue Res. 2018;371(3):531–539. PMID: 29383445; PMCID: PMC5820392. doi:10.1007/s00441-017-2785-7
39. Davies LC, Jenkins SJ, Allen JE, Taylor PR. Tissue-resident macrophages. Nat Immunol. 2013;14(10):986–995. doi:10.1038/ni.2705
40. Wynn TA, Vannella KM. Macrophages in tissue repair, regeneration, and fibrosis. Immunity. 2016;44(3):450–462. PMID: 26982353; PMCID: PMC4794754. doi:10.1016/j.immuni.2016.02.015
41. Rappolee DA, Mark D, Banda MJ, Werb Z. Wound macrophages express TGF-alpha and other growth factors in vivo: analysis by mRNA phenotyping. Science. 1988;241:708–712.
42. Peiseler M, Kubes P. More friend than foe: the emerging role of neutrophils in tissue repair. J Clin Invest. 2019;129:2629–2639.
43. Mnich ME, van Dalen R, Van Sorge NM. C-type lectin receptors in host defense against bacterial pathogens. Front Cell Infect Microbiol. 2020;10:309. doi:10.3389/fcimb.2020.00309
44. Kanazawa N. Dendritic cell immunoreceptors: c-type lectin receptors for pattern recognition and signaling on antigen-presenting cells. J Dermatol Sci. 2007;45(2):77–86. PMID: 17046204. doi:10.1016/j.jdermsci.2006.09.001
45. Fusté NP, Guasch M, Guillen P, et al. Barley β-glucan accelerates wound healing by favoring migration versus proliferation of human dermal fibroblasts. Carbohydr Polym. 2019;210:389–398. PMID: 30732776. doi:10.1016/j.carbpol.2019.01.090
46. Goodridge HS, Wolf AJ, Underhill DM. Beta-glucan recognition by the innate immune system. Immunol Rev. 2009;230(1):38–50. PMID: 19594628; PMCID: PMC6618291. doi:10.1111/j.1600-065X.2009.00793.x
47. Yamaguchi K, Kanno E, Tanno H, et al. Distinct roles for dectin-1 and dectin-2 in skin wound healing and neutrophilic inflammatory responses. J Invest Dermatol. 2021;141(1):164–176.e8. PMID: 32511980. doi:10.1016/j.jid.2020.04.030
48. Seo G, Hyun C, Choi S, et al. The wound healing effect of four types of beta-glucan. Appl Biol Chem. 2019;62:20.
 © 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2022 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.